
Molecular Analysis of East African Lumpy Skin Disease Viruses Reveals a Mixed Isolate with Features of Both Vaccine and Field Isolates

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
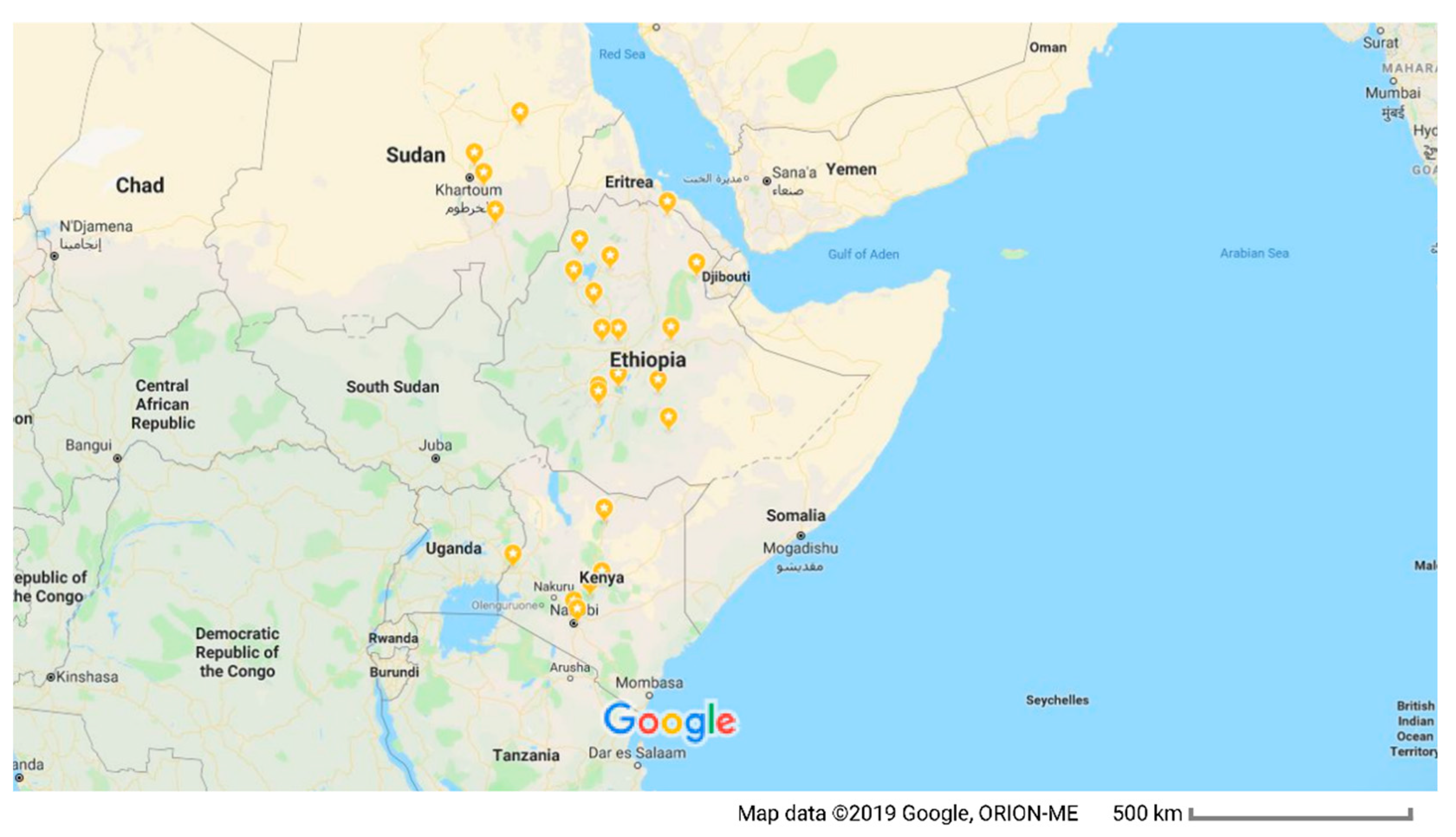
2.1. Samples and DNA Extraction
2.2. Amplification of RPO30, GPCR, EEV Glycoprotein, and B22R Genes
2.3. Sequencing and Phylogenetic Analysis
2.4. Targeted Next-Generation Sequencing of Selected Variable Sites in the LSDV Genome
3. Results
3.1. PCR Amplification and Sequencing
3.2. Analysis of the RPO30 Gene
3.3. Analysis of the GPCR Gene
3.4. Analysis of the EEV Glycoprotein and the B22R Genes of LSDV Embu/B338/2011
3.5. Analysis of Additional Variable Sites in the LSDV Genome by Targeted Next-Generation Sequencing
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Buller, R.M.; Arif, B.M.; Black, D.N.; Dumbell, K.R.; Esposito, J.J.; Lefkowitz, E.J.; McFadden, G.; Moss, B.; Mercer, A.A.; Moyer, R.W.; et al. Family poxviridae. In Virus Taxonomy: Classification and Nomenclature of Viruses. Eighth Report of the International Committee on Taxonomy of Viruses; Mayo, M.A., Maniloff, J., Desselberger, U., Ball, L.A., Eds.; Elsevier Academic Press: San Diego, CA, USA, 2005; pp. 117–133. [Google Scholar] [CrossRef]
- Babiuk, S.; Bowden, T.R.; Boyle, D.B.; Wallace, D.B.; Kitching, R.P. Capripoxviruses: An Emerging Worldwide Threat to Sheep, Goats and Cattle. Transbound. Emerg. Dis. 2008, 55, 263–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tulman, E.R.; Afonso, C.L.; Lu, Z.; Zsak, L.; Kutish, G.F.; Rock, D.L. Genome of Lumpy Skin Disease Virus. J. Virol. 2001, 75, 7122–7130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Goff, C.; Lamien, C.E.; Fakhfakh, E.; Chadeyras, A.; Aba-Adulugba, E.; Libeau, G.; Tuppurainen, E.; Wallace, D.B.; Adam, T.; Silber, R.; et al. Capripoxvirus G-protein-coupled chemokine receptor: A host-range gene suitable for virus animal origin discrimination. J. Gen. Virol. 2009, 90, 1967–1977. [Google Scholar] [CrossRef]
- OIE. Lumpy skin disease. In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals; OIE: Paris, France, 2017. [Google Scholar]
- Davies, F.G. Characteristics of a virus causing a pox disease in sheep and goats in Kenya, with observations on the epidemiology and control. J. Hyg. (Lond) 1976, 76, 163–171. [Google Scholar] [CrossRef]
- Ali, B.B.H. Investigation of the First Outbreaks of Lumpy Skin Disease in the Sudan. Br. Vet. J. 1977, 133, 184–189. [Google Scholar] [CrossRef]
- Mebratu, C.Y. Observation on the outbreak of lumpy skin disease in Ethiopia. Rev. Elev. Med. Vet. Pays Trop. 1984, 37, 395–399. [Google Scholar]
- Tuppurainen, E.S.M.; Oura, C.A.L. Review: Lumpy Skin Disease: An Emerging Threat to Europe, the Middle East and Asia. Transbound. Emerg. Dis. 2012, 59, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Mercier, A.; Arsevska, E.; Bournez, L.; Bronner, A.; Calavas, D.; Cauchard, J.; Falala, S.; Caufour, P.; Tisseuil, C.; Lefrançois, T.; et al. Spread rate of lumpy skin disease in the Balkans, 2015–2016. Transbound. Emerg. Dis. 2018, 65, 240–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badhy, S.C.; Chowdhury, M.G.A.; Settypalli, T.B.K.; Cattoli, G.; Lamien, C.E.; Fakir, M.A.U.; Akter, S.; Osmani, M.G.; Talukdar, F.; Begum, N.; et al. Molecular characterization of lumpy skin disease virus (LSDV) emerged in Bangladesh reveals unique genetic features compared to contemporary field strains. BMC Vet. Res. 2021, 17, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Lu, G.; Xie, J.; Luo, J.; Shao, R.; Jia, K.; Li, S. Lumpy skin disease outbreaks in China, since 3 August 2019. Transbound. Emerg. Dis. 2020. [Google Scholar] [CrossRef]
- Sudhakar, S.B.; Mishra, N.; Kalaiyarasu, S.; Jhade, S.K.; Hemadri, D.; Sood, R.; Bal, G.C.; Nayak, M.K.; Pradhan, S.K.; Singh, V.P. Lumpy skin disease (LSD) outbreaks in cattle in Odisha state, India in August 2019: Epidemiological features and molecular studies. Transbound. Emerg. Dis. 2020, 67, 2408–2422. [Google Scholar] [CrossRef] [PubMed]
- Tran, H.T.T.; Truong, A.D.; Dang, A.K.; Ly, D.V.; Nguyen, C.T.; Chu, N.T.; Hoang, T.V.; Nguyen, H.T.; Nguyen, V.T.; Dang, H.V. Lumpy skin disease outbreaks in vietnam, 2020. Transbound. Emerg. Dis. 2021. [Google Scholar] [CrossRef] [PubMed]
- El-Kholy, A.A.; Soliman, H.M.T.; Abdelrahman, K.A. Polymerase chain reaction for rapid diagnosis of a recent lumpy skin disease virus incursion to Egypt. Arab J. Biotechnol. 2008, 11, 293–302. [Google Scholar]
- Alkhamis, M.A.; VanderWaal, K. Spatial and temporal epidemiology of lumpy skin disease in the Middle East, 2012–2015. Front. Vet. Sci. 2016, 3, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamien, C.E.; Le Goff, C.; Silber, R.; Wallace, D.B.; Gulyaz, V.; Tuppurainen, E.; Madani, H.; Caufour, P.; Adam, T.; Harrak, M.E.; et al. Use of the Capripoxvirus homologue of Vaccinia virus 30 kDa RNA polymerase subunit (RPO30) gene as a novel diagnostic and genotyping target: Development of a classical PCR method to differentiate Goat poxvirus from Sheep poxvirus. Vet. Microbiol. 2011, 149, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Gelaye, E.; Belay, A.; Ayelet, G.; Jenberie, S.; Yami, M.; Loitsch, A.; Tuppurainen, E.; Grabherr, R.; Diallo, A.; Lamien, C.E. Capripox disease in Ethiopia: Genetic differences between field isolates and vaccine strain, and implications for vaccination failure. Antivir. Res. 2015, 119, 28–35. [Google Scholar] [CrossRef]
- Menasherow, S.; Erster, O.; Rubinstein-Giuni, M.; Kovtunenko, A.; Eyngor, E.; Gelman, B.; Khinich, E.; Stram, Y. A high-resolution melting (HRM) assay for the differentiation between Israeli field and Neethling vaccine lumpy skin disease viruses. J. Virol. Methods 2016, 232, 12–15. [Google Scholar] [CrossRef]
- Chibssa, T.R.; Settypalli, T.B.K.; Berguido, F.J.; Grabherr, R.; Loitsch, A.; Tuppurainen, E.; Nwankpa, N.; Tounkara, K.; Madani, H.; Omani, A.; et al. An HRM Assay to Differentiate Sheeppox Virus Vaccine Strains from Sheeppox Virus Field Isolates and other Capripoxvirus Species. Sci. Rep. 2019, 9, 6646. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0. molecular biology and evolution. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesian phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 2012. [Google Scholar] [CrossRef] [Green Version]
- Yu, G.; Smith, D.K.; Zhu, H.; Guan, Y.; Lam, T.T.Y. Ggtree: An r package for visualization and annotation of phylogenetic trees with their covariates and other associated data. Methods Ecol. Evol. 2017. [Google Scholar] [CrossRef]
- Van Schalkwyk, A.; Kara, P.; Ebersohn, K.; Mather, A.; Annandale, C.H.; Venter, E.H.; Wallace, D.B. Potential link of single nucleotide polymorphisms to virulence of vaccine-associated field strains of lumpy skin disease virus in South Africa. Transbound. Emerg. Dis. 2020, 67, 2946–2960. [Google Scholar] [CrossRef]
- Thorvaldsdóttir, H.; Robinson, J.T.; Mesirov, J.P. Integrative Genomics Viewer (IGV): High-performance genomics data visualization and exploration. Brief. Bioinform. 2013, 14, 178–192. [Google Scholar] [CrossRef] [Green Version]
- Sprygin, A.; Babin, Y.; Pestova, Y.; Kononova, S.; Wallace, D.B.; Van Schalkwyk, A.; Byadovskaya, O.; Diev, V.; Lozovoy, D.; Kononov, A. Analysis and insights into recombination signals in lumpy skin disease virus recovered in the field. PLoS ONE 2018, 13, e0207480. [Google Scholar] [CrossRef] [PubMed]
- Kononov, A.; Byadovskaya, O.; Kononova, S.; Yashin, R.; Zinyakov, N.; Mischenko, V.; Perevozchikova, N.; Sprygin, A. Detection of vaccine-like strains of lumpy skin disease virus in outbreaks in Russia in 2017. Arch. Virol. 2019, 164, 1575–1585. [Google Scholar] [CrossRef] [PubMed]
- Sprygin, A.; Pestova, Y.; Bjadovskaya, O.; Prutnikov, P.; Zinyakov, N.; Kononova, S.; Ruchnova, O.; Lozovoy, D.; Chvala, I.; Kononov, A. Evidence of recombination of vaccine strains of lumpy skin disease virus with field strains, causing disease. PLoS ONE 2020, 15. [Google Scholar] [CrossRef] [PubMed]
- Gershon, P.D.; Kitching, R.P.; Hammond, J.M.; Black, D.N. Poxvirus genetic recombination during natural virus transmission. J. Gen. Virol. 1989, 70, 485–489. [Google Scholar] [CrossRef]
- Agianniotaki, E.I.; Tasioudi, K.E.; Chaintoutis, S.C.; Iliadou, P.; Mangana-Vougiouka, O.; Kirtzalidou, A.; Alexandropoulos, T.; Sachpatzidis, A.; Plevraki, E.; Dovas, C.I.; et al. Lumpy skin disease outbreaks in Greece during 2015–16, implementation of emergency immunization and genetic differentiation between field isolates and vaccine virus strains. Vet. Microbiol. 2017, 201, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Yeruham, I.; Perl, S.; Nyska, A.; Abraham, A.; Davidson, M.; Haymovitch, M.; Zamir, O.; Grinstein, H. Adverse reactions in cattle to a capripox vaccine. Vet. Rec. 1994, 135, 330–332. [Google Scholar] [CrossRef]
- Tuppurainen, E.S.M.; Pearson, C.R.; Bachanek-bankowska, K.; Knowles, N.J.; Amareen, S.; Frost, L.; Henstock, M.R.; Lamien, C.E.; Diallo, A.; Mertens, P.P.C. Characterization of sheep pox virus vaccine for cattle against lumpy skin disease virus. Antivir. Res. 2014, 109, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Abutarbush, S.M.; Hananeh, W.M.; Ramadan, W.; Al Sheyab, O.M.; Alnajjar, A.R.; Al Zoubi, I.G.; Knowles, N.J.; Bachanek-Bankowska, K.; Tuppurainen, E.S.M. Adverse Reactions to Field Vaccination Against Lumpy Skin Disease in Jordan. Transbound. Emerg. Dis. 2016, 63, e213–e219. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Strain Name | Host | Origin | Sample Type | Vaccination History | Year | Accession Number | |
|---|---|---|---|---|---|---|---|---|
| RPO30 | GPCR | |||||||
| 1 | * Marsabit/B291/2007 | Cattle | Kenya | Skin lesion | Non-vaccinated | 2007 | MK302092 | MK302070 |
| 2 | * Embu/B338/2011 | Cattle | Kenya | Skin lesion | Vaccinated | 2011 | MK302093 | MK302071 |
| 3 | * Bungoma/B624/2010 | Cattle | Kenya | Skin lesion | Non-vaccinated | 2010 | MK302094 | MK302072 |
| 4 | * Arsi/B1/2011 | Cattle | Ethiopia | Skin lesion | Non-vaccinated | 2011 | MK302095 | MK302073 |
| 5 | * Asella/B2/2011 | Cattle | Ethiopia | Skin lesion | Non-vaccinated | 2011 | MK302096 | MK302074 |
| 6 | * Ziway/B3/2011 | Cattle | Ethiopia | Skin lesion | Non-vaccinated | 2011 | MK302097 | MK302075 |
| 7 | * Adama/B4/2011 | Cattle | Ethiopia | Skin lesion | Non-vaccinated | 2011 | MK302098 | MK302076 |
| 8 | * Guder/B5/2008 | Cattle | Ethiopia | Skin lesion | Non-vaccinated | 2008 | MK302099 | MK302077 |
| 9 | * Sodo/B22/2010 | Cattle | Ethiopia | Skin lesion | Non-vaccinated | 2010 | MK302100 | MK302078 |
| 10 | * Humbo/B23/2010 | Cattle | Ethiopia | Skin lesion | Non-vaccinated | 2010 | MK302101 | MK302079 |
| 11 | * Sodo/B24/2010 | Cattle | Ethiopia | Skin lesion | Non-vaccinated | 2010 | MK302102 | MK302080 |
| 12 | * Sundus/1970 | Cattle | Sudan | Skin lesion | Non-vaccinated | 1971 | MK302103 | MK302081 |
| 13 | * Sudan North/2008 | Cattle | Sudan | Skin lesion | Non-vaccinated | 2008 | MK302104 | MK302082 |
| 14 | * Sinnar/2006 | Cattle | Sudan | Skin lesion | Non-vaccinated | 2006 | MK302105 | MK302083 |
| 15 | * Toke/B6/2008 | Cattle | Ethiopia | Swab samples | Non-vaccinated | 2008 | MK302106 | MK302084 |
| 16 | * Gindo/B7/2008 | Cattle | Ethiopia | Swab samples | Non-vaccinated | 2008 | MK302107 | MK302085 |
| 17 | * Ambo/B8/2008 | Cattle | Ethiopia | Swab samples | Non-vaccinated | 2008 | MK302108 | MK302086 |
| 18 | * Holeta/B9/2008 | Cattle | Ethiopia | Swab samples | Non-vaccinated | 2008 | MK302109 | MK302087 |
| 19 | * Ginchi/B10/2008 | Cattle | Ethiopia | Swab samples | Non-vaccinated | 2008 | MK302110 | MK302088 |
| 20 | * Chilimo/B11/2008 | Cattle | Ethiopia | Swab samples | Non-vaccinated | 2008 | MK302111 | MK302089 |
| 21 | * Galesa/B12/2008 | Cattle | Ethiopia | Swab samples | Non-vaccinated | 2008 | MK302112 | MK302090 |
| 22 | * Massalamia/P04/1971 | Cattle | Sudan | Cell culture | Non-vaccinated | 1971 | MK302113 | MK302091 |
| 23 | Sudan/99-Atbara | Cattle | Sudan | 1999 | GU119944 | FJ869367 | ||
| 24 | Sudan/06-Obied | Cattle | Sudan | 2006 | GU119938 | FJ869369 | ||
| 25 | KSGP 0240 | Sheep | Kenya | 1976 | KX683219 | KX683219 | ||
| 26 | KS-1 | Sheep | Kenya | 1976 | KJ818290 | KJ818283 | ||
| 27 | NI-2490 | Cattle | Kenya | 1958 | AF325528 | AF325528 | ||
| 28 | Kenya | Cattle | Kenya | 1950 | MN072619 | MN072619 | ||
| 29 | Adama/B01/2011 | Cattle | Ethiopia | 2011 | KP663667 | KP663690 | ||
| 30 | Andassa/B04/2012 | Cattle | Ethiopia | 2012 | KP663671 | KP663694 | ||
| 31 | Kadjima/B01/2009 | Cattle | Ethiopia | 2009 | KP663681 | KP663704 | ||
| 32 | Mojo/B01/2011 | Cattle | Ethiopia | 2011 | KP663683 | KP663706 | ||
| 33 | Wenji/B01/2011 | Cattle | Ethiopia | 2011 | KP663687 | KP663710 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chibssa, T.R.; Sombo, M.; Lichoti, J.K.; Adam, T.I.B.; Liu, Y.; Elraouf, Y.A.; Grabherr, R.; Settypalli, T.B.K.; Berguido, F.J.; Loitsch, A.; et al. Molecular Analysis of East African Lumpy Skin Disease Viruses Reveals a Mixed Isolate with Features of Both Vaccine and Field Isolates. Microorganisms 2021, 9, 1142. https://doi.org/10.3390/microorganisms9061142
Chibssa TR, Sombo M, Lichoti JK, Adam TIB, Liu Y, Elraouf YA, Grabherr R, Settypalli TBK, Berguido FJ, Loitsch A, et al. Molecular Analysis of East African Lumpy Skin Disease Viruses Reveals a Mixed Isolate with Features of Both Vaccine and Field Isolates. Microorganisms. 2021; 9(6):1142. https://doi.org/10.3390/microorganisms9061142
Chicago/Turabian StyleChibssa, Tesfaye Rufael, Melaku Sombo, Jacqueline Kasiiti Lichoti, Tajelser Idris Badri Adam, Yang Liu, Yazeed Abd Elraouf, Reingard Grabherr, Tirumala Bharani K. Settypalli, Francisco J. Berguido, Angelika Loitsch, and et al. 2021. "Molecular Analysis of East African Lumpy Skin Disease Viruses Reveals a Mixed Isolate with Features of Both Vaccine and Field Isolates" Microorganisms 9, no. 6: 1142. https://doi.org/10.3390/microorganisms9061142