
Procaryotic Diversity and Hydrogenotrophic Methanogenesis in an Alkaline Spring (La Crouen, New Caledonia)

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
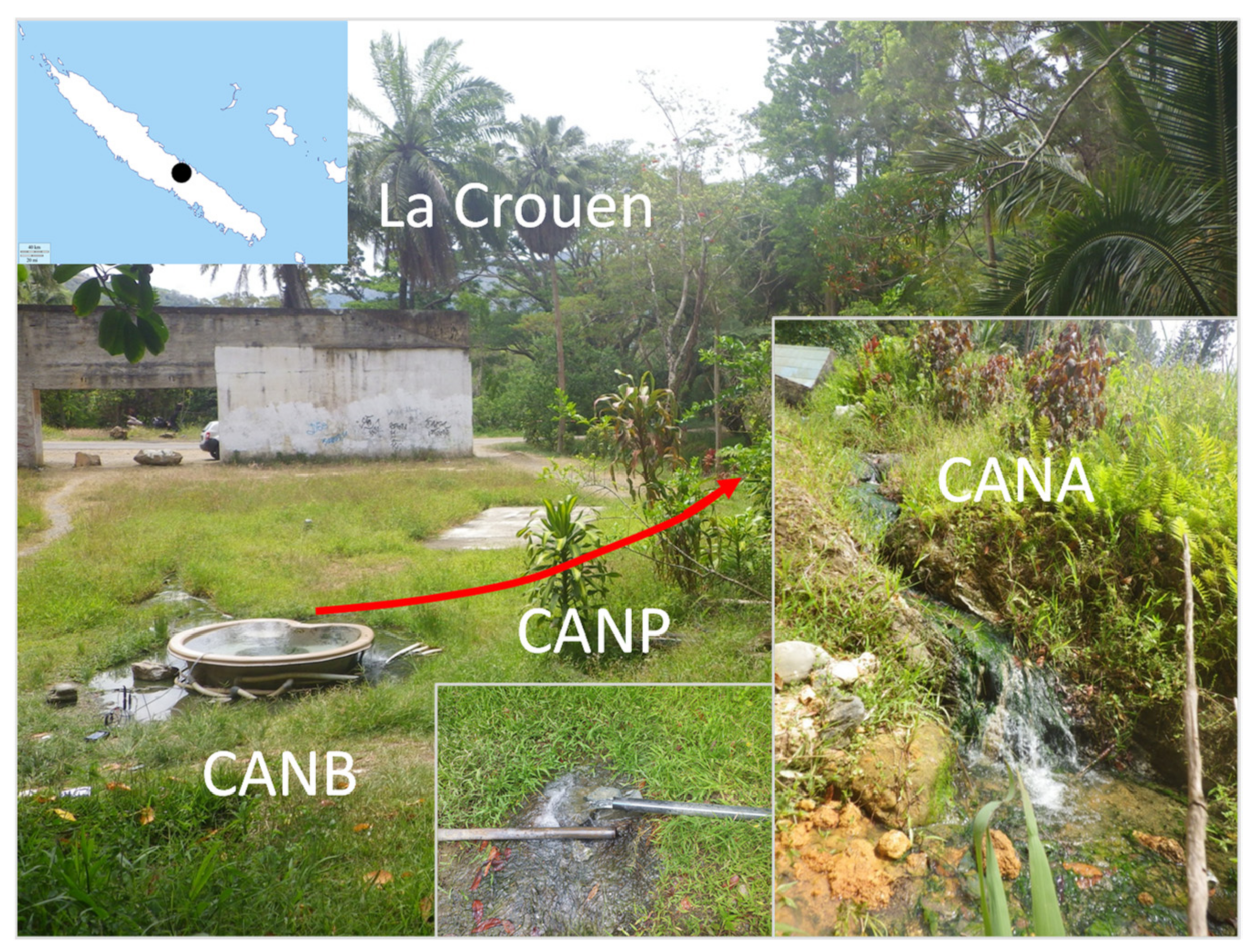
2.1. Study Site
2.2. Sample Collection
2.3. Chemical Analysis
2.4. Methane Production in Anaerobic Enrichment Cultures
2.5. Isolation and Identification of Methanogens
2.6. DNA Extraction, PCR, and MiSeq Illumina Sequencing of 16S rRNA Genes from Sediment and Water Samples
2.7. Statistical Analyses
3. Results
3.1. Chemical Composition of the Waters
3.2. Abundance and Diversity of Prokaryotic Communities
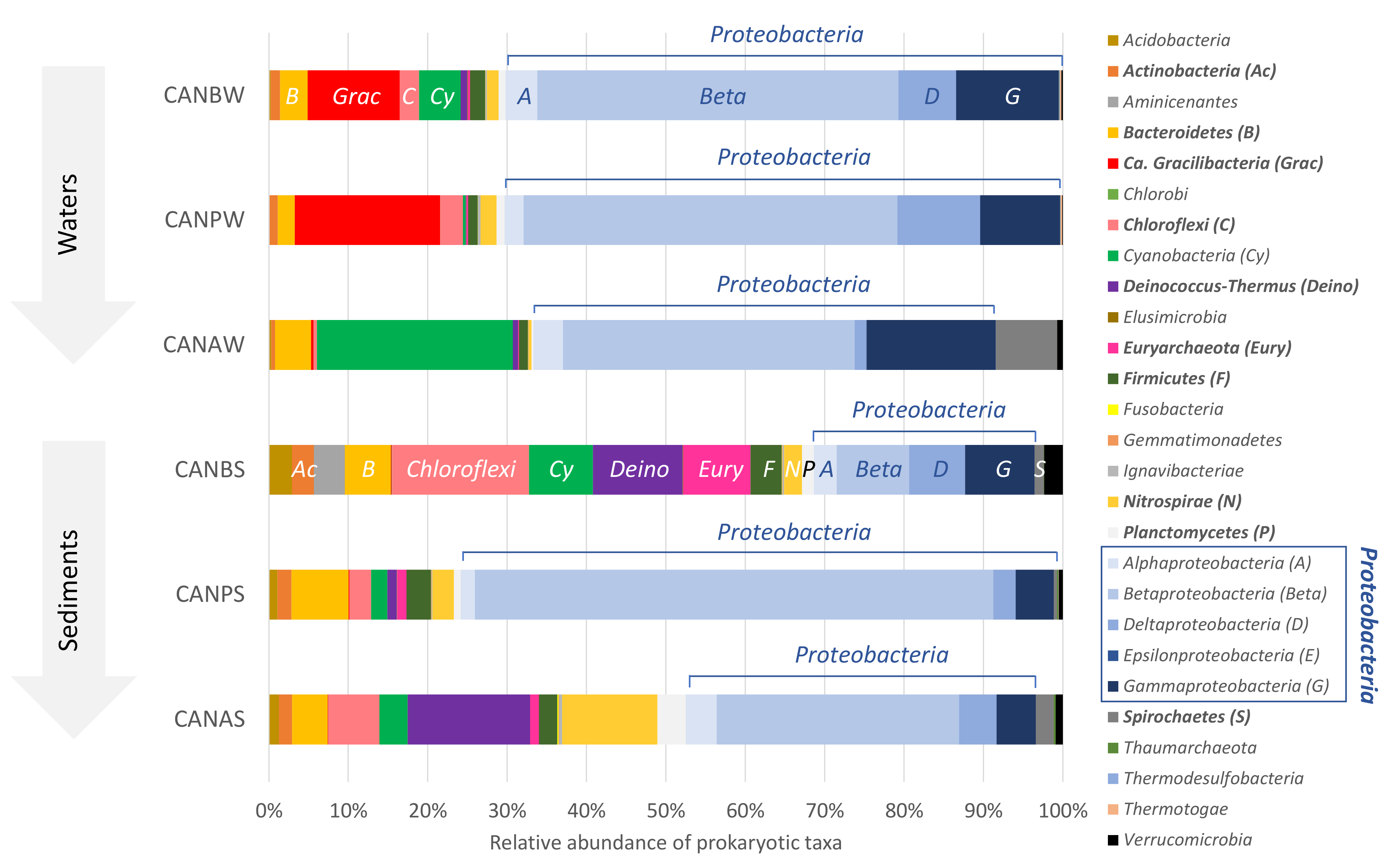
3.3. Composition and Distribution of Prokaryotic Communities
3.4. Distribution and Diversity of Dominant Bacterial OTUs in La Crouen Spring
3.5. Diversity of Dominant Archaeal OTUs in La Crouen Spring
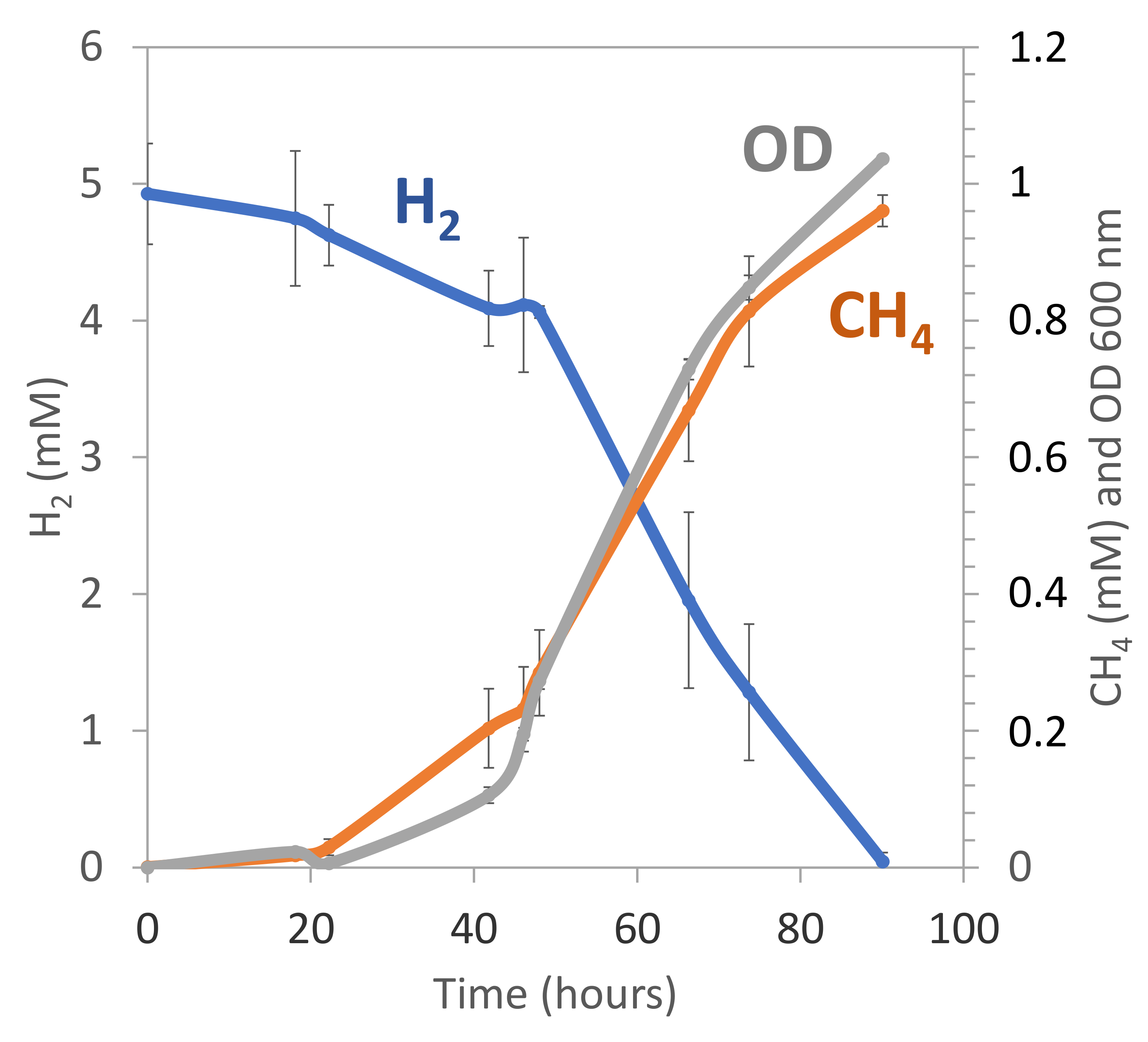
3.6. Cultivation and Isolation of Alkaliphilic Methanogens from La Crouen
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Launay, J.; Fontes, J.C. Les sources thermales de Prony (Nouvelle Calédonie) et leurs précipités chimiques, exemple de formation de brucite primaire. Geol. Fr. 1985, 1, 83–100. [Google Scholar]
- Monnin, C.; Chavagnac, V.; Boulart, C.; Ménez, B.; Gérard, M.; Pisapia, C.; Quéméneur, M.; Erauso, G.; Postec, A.; Guentas-Dombrowski, L.; et al. Fluid chemistry of the low temperature hyperalkaline hydrothermal system of the Prony Bay (New Caledonia). Biogeosciences 2014, 11, 5687–5706. [Google Scholar] [CrossRef] [Green Version]
- Deville, E.; Prinzhofer, A. The origin of N2-H2-CH4-rich natural gas seepages in ophiolitic context: A major and noble gases study of fluid seepages in New Caledonia. Chem. Geol. 2016, 440, 139–147. [Google Scholar] [CrossRef]
- Maurizot, P.; Sevin, B.; Lesimple, S.; Collot, J.; Jeanpert, J.; Bailly, L.; Robineau, B.; Patriat, M.; Etienne, S.; Monnin, C. Mineral resources and prospectivity of non-ultramafic rocks of New-Caledonia. Mortimer, N., Ed.; New Caledonia: Geology, Geodynamic Evolution and Mineral Resources. Geol. Soc. Lond. Mem. 2020, 51, 215–245. [Google Scholar] [CrossRef]
- Cox, M.E.; Launay, J.; Paris, J.P. Geochemistry of low temperature geothermal systems in New Caledonia. In Proceedings of the Pacific Geothermal Conference, Aukland, New Zeland, 8 November 1982; pp. 453–459. [Google Scholar]
- Kelley, D.S.; Karson, J.A.; Früh-Green, G.L.; Yoerger, D.R.; Shank, T.M.; Butterfield, D.A.; Hayes, J.M.; Schrenk, M.O.; Olson, E.J.; Proskurowski, G.; et al. A serpentinite-hosted ecosystem: The Lost City hydrothermal field. Science 2005, 307, 1428–1434. [Google Scholar] [CrossRef]
- Schrenk, M.O.; Kelley, D.S.; Bolton, S.A.; Baross, J.A. Low archaeal diversity linked to subseafloor geochemical processes at the Lost City Hydrothermal Field, Mid-Atlantic Ridge. Environ. Microbiol. 2004, 6, 1086–1095. [Google Scholar] [CrossRef] [PubMed]
- Brazelton, W.J.; Schrenk, M.O.; Kelley, D.S.; Baross, J.A. Methane- and sulfur-metabolizing microbial communities dominate the Lost City hydrothermal field ecosystem. Appl. Environ. Microbiol. 2006, 72, 6257–6270. [Google Scholar] [CrossRef] [Green Version]
- Brazelton, W.J.; Mehta, M.P.; Kelley, D.S.; Baross, J.A. Physiological differentiation within a single-species biofilm fueled by serpentinization. mBio 2011, 2, 2. [Google Scholar] [CrossRef] [Green Version]
- Postec, A.; Quéméneur, M.; Bes, M.; Mei, N.; Aissa, F.B.; Payri, C.; Pelletier, B.; Monnin, C.; Guentas-Dombrowsky, L.; Ollivier, B.; et al. Microbial diversity in a submarine carbonate edifice from the serpentinizing hydrothermal system of the Prony Bay (New Caledonia) over a 6-year period. Front. Microbiol. 2015, 6, 857. [Google Scholar] [CrossRef] [PubMed]
- Quéméneur, M.; Bes, M.; Postec, A.; Mei, N.; Hamelin, J.; Monnin, C.; Chavagnac, V.; Payri, C.; Pelletier, B.; Guentas-Dombrowsky, L.; et al. Spatial distribution of microbial communities in the shallow submarine alkaline hydrothermal field of the Prony Bay, New Caledonia. Environ. Microbiol. Rep. 2014, 6, 665–674. [Google Scholar] [CrossRef] [Green Version]
- Frouin, E.; Bes, M.; Ollivier, B.; Quéméneur, M.; Postec, A.; Debroas, D.; Armougom, F.; Erauso, G. Diversity of rare and abundant prokaryotic phylotypes in the Prony Hydrothermal Field and comparison with other serpentinite-hosted ecosystems. Front. Microbiol. 2018, 9, 102. [Google Scholar] [CrossRef] [Green Version]
- Pisapia, C.; Gérard, E.; Gérard, M.; Lecourt, L.; Pelletier, B.; Payri, C.; Monnin, C.; Guentas, L.; Postec, A.; Quéméneur, M.; et al. Mineralizing filamentous bacteria from the Prony Bay Hydrothermal Field give new insights into the functioning of serpentinization-based subseafloor ecosystems. Front. Microbiol. 2017, 8, 57. [Google Scholar] [CrossRef] [Green Version]
- Ben Aissa, F.; Postec, A.; Erauso, G.; Payri, C.; Pelletier, B.; Hamdi, M.; Ollivier, B.; Fardeau, M.-L. Vallitalea pronyensis sp. nov., isolated from a marine alkaline hydrothermal chimney. Int. J. Syst Evol. Microbiol. 2014, 64, 1160–1165. [Google Scholar] [CrossRef]
- Ben Aissa, F.; Postec, A.; Erauso, G.; Payri, C.; Pelletier, B.; Hamdi, M.; Fardeau, M.-L.; Ollivier, B. Characterization of Alkaliphilus hydrothermalis sp. nov., a novel alkaliphilic anaerobic bacterium, isolated from a carbonaceous chimney of the Prony hydrothermal field, New Caledonia. Extremophiles 2015, 19, 183–188. [Google Scholar] [CrossRef] [Green Version]
- Bes, M.; Merrouch, M.; Joseph, M.; Quéméneur, M.; Payri, C.; Pelletier, B.; Ollivier, B.; Fardeau, M.-L.; Erauso, G.; Postec, A. Acetoanaerobium pronyense sp. nov., an anaerobic mesophilic bacterium isolated from the Prony alkaline Hydrothermal Field, New Caledonia. Int. J. Syst Evol. Microbiol. 2015, 65, 2574–2580. [Google Scholar] [CrossRef]
- Mei, N.; Zergane, N.; Postec, A.; Erauso, G.; Ollier, A.; Payri, C.; Pelletier Fardeau, M.-L.; Ollivier, B.; Quéméneur, M. Fermentative hydrogen production by a new alkaliphilic Clostridium sp. (strain PROH2) isolated from a shallow submarine hydrothermal chimney in Prony Bay, New Caledonia. Int. J. Hydrog. Energy 2014, 39, 19465–19473. [Google Scholar] [CrossRef]
- Mei, N.; Postec, A.; Monnin, C.; Pelletier, B.; Payri, C.E.; Ménez, B.; Frouin, E.; Ollivier, B.; Erauso, G.; Quéméneur, M. Metagenomic and PCR-based diversity surveys of [FeFe]-hydrogenases combined with isolation of alkaliphilic hydrogen-producing bacteria from the serpentinite-hosted Prony Hydrothermal Field, New Caledonia. Front. Microbiol. 2016, 7, 1301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mei, N.; Postec, A.; Erauso, G.; Joseph, M.; Pelletier, B.; Payri, C.; Ollivier, B.; Quéméneur, M. Serpentinicella alkaliphila gen. nov., sp. nov., a novel alkaliphilic anaerobic bacterium isolated from the serpentinite-hosted Prony hydrothermal field, New Caledonia. Int. J. Syst. Evol. Microbiol. 2016, 66, 4464–4470. [Google Scholar] [CrossRef] [PubMed]
- Postec, A.; Quéméneur, M.; Lecoeuvre, A.; Chabert, N.; Joseph, M.; Erauso, G. Alkaliphilus serpentinus sp. nov. and Alkaliphilus pronyensis sp. nov., two novel anaerobic alkaliphilic species isolated from the serpentinite-hosted Prony Bay Hydrothermal Field (New Caledonia). Syst. Appl. Microbiol. 2021, 44, 126175. [Google Scholar] [CrossRef] [PubMed]
- Naganuma, T. Geo-Biological Coupling of Authigenic Carbonate Formation and Autotrophic Faunal Colonization at Deep-Sea Methane Seeps I: Geo-Biological Settings, Marine Ecology—Biotic and Abiotic Interactions, Muhammet Türkoğlu, Umur Önal and Ali Ismen. IntechOpen 2018. [Google Scholar] [CrossRef] [Green Version]
- Reeves, E.P.; Fiebig, J. Abiotic Synthesis of Methane and Organic-Compounds in Earth’s Lithosphere. Elements 2020, 16, 25–31. [Google Scholar] [CrossRef]
- Garcia, J.L.; Patel, B.K.; Ollivier, B. Taxonomic, phylogenetic, and ecological diversity of methanogenic Archaea. Anaerobe 2000, 6, 205–226. [Google Scholar] [CrossRef]
- Lyu, Z.; Shao, N.; Akinyemi, T.; Whitman, W.B. Methanogenesis. Curr. Biol. 2018, 28, 727–732. [Google Scholar] [CrossRef] [Green Version]
- Karl, D.M.; Beversdorf, L.; Björkman, K.M.; Church, M.J.; Martinez, A.; Delong, E.F. Aerobic production of methane in the sea. Nat. Geosci. 2008, 1, 473–478. [Google Scholar] [CrossRef]
- Carini, P.; White, A.E.; Campbell, E.O.; Giovannoni, S.J. Methane production by phosphate-starved SAR11 chemoheterotrophic marine bacteria. Nat. Commun. 2014, 5, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Brazelton, W.J.; Thornton, C.N.; Hyer, A.; Twing, K.I.; Longino, A.A.; Lang, S.Q.; Lilley, M.D.; Früh-Green, G.L.; Schrenk, M.O. Metagenomic identification of active methanogens and methanotrophs in serpentinite springs of the Voltri Massif, Italy. PeerJ 2017, 5, e2945. [Google Scholar] [CrossRef] [Green Version]
- Kraus, E.A.; Nothaft, D.; Stamps, B.W.; Rempfert, K.R.; Ellison, E.T.; Matter, J.M.; Templeton, A.S.; Boyd, E.S.; Spear, J.R. Molecular evidence for an active microbial methane cycle in subsurface serpentinite-hosted groundwaters in the Samail Ophiolite, Oman. Appl. Environ. Microbiol. 2021, 87, e02068-20. [Google Scholar] [CrossRef]
- Fones, E.M.; Colman, D.R.; Kraus, E.A.; Stepanauskas, R.; Templeton, A.S.; Spear, J.R.; Boyd, E.S. Diversification of methanogens into hyperalkaline serpentinizing environments through adaptations to minimize oxidant limitation. ISME J. 2020, 15, 1–15. [Google Scholar]
- Whi Milkov, A.V.; Etiope, G. Revised genetic diagrams for natural gases based on a global dataset of >20,000 samples. Org. Geochem. 2018, 125, 109–120. [Google Scholar] [CrossRef]
- Magen, C.; Lapham, L.L.; Pohlman, J.W.; Marshall, K.; Bosman, S.; Casso, M.; Chanton, J.P. A simple headspace equilibration method for measuring dissolved methane. Limnol. Oceanogr. Methods 2014, 12, 637–650. [Google Scholar] [CrossRef]
- Vongvichiankul, C.; Deebao, J.; Khongnakorn, W. Relationship between pH, oxidation reduction potential (ORP) and biogas production in mesophilic screw anaerobic digester. Energy Procedia 2017, 138, 877–882. [Google Scholar] [CrossRef]
- Fetzer, S.; Conrad, R. Effect of redox potential on methanogenesis by Methanosarcina barkeri. Arch. Microbiol. 1993, 160, 108–113. [Google Scholar] [CrossRef]
- Widdel, F.; Kohring, G.W.; Mayer, F. Studies on dissimilatory sulfate-reducing bacteria that decompose fatty acids. Arch. Microbiol. 1983, 134, 286–294. [Google Scholar] [CrossRef]
- Hungate, R.E. A roll tube method for cultivation of strict anaerobes. Methods Microbiol. 1969, 3, 117–132. [Google Scholar]
- Balch, W.E.; Fox, G.E.; Magrum, L.J.; Woese, C.R.; Wolfe, R.S. Methanogens: Reevaluation of a unique biological group. Microbiol. Rev. 1979, 43, 260–296. [Google Scholar] [CrossRef]
- Muyzer, G.; De Waal, E.C.; Uitterlinden, A.G. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl. Environ. Microbiol. 1993, 59, 695–700. [Google Scholar] [CrossRef] [Green Version]
- Großkopf, R.; Janssen, P.H.; Liesack, W. Diversity and structure of the methanogenic community in anoxic rice paddy soil microcosms as examined by cultivation and direct 16S rRNA gene sequence retrieval. Appl. Environ. Microbiol. 1998, 64, 960–969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicol, G.W.; Leininger, S.; Schleper, C.; Prosser, J.I. The influence of soil pH on the diversity, abundance and transcriptional activity of ammonia oxidizing archaea and bacteria. Environ. Microbiol. 2008, 10, 2966–2978. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Felsenstein, J. Phylogenies from restriction sites: A maximum-likelihood approach. Evolution 1992, 46, 159–173. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casamayor, E.O.; Massana, R.; Benlloch, S.; Øvreås, L.; Díez, B.; Goddard, V.J.; Gasol, J.M.; Joint, I.; Rodriguez-Valera, F.; Pedros-Alio, C. Changes in archaeal, bacterial and eukaryal assemblages along a salinity gradient by comparison of genetic fingerprinting methods in a multipond solar saltern. Environ. Microbiol. 2002, 4, 338–348. [Google Scholar] [CrossRef] [PubMed]
- Ovreås, L.; Forney, L.; Daae, F.L.; Torsvik, V. Distribution of bacterioplankton in meromictic Lake Saelenvannet, as determined by denaturing gradient gel electrophoresis of PCR-amplified gene fragments coding for 16S rRNA. Appl. Environ. Microbiol. 1997, 63, 3367–3373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nunoura, T.; Oida, H.; Miyazaki, J.; Miyashita, A.; Imachi, H.; Takai, K. Quantification of mcrA by fluorescent PCR in methanogenic and methanotrophic microbial communities. FEMS Microbiol. Ecol. 2008, 64, 240–247. [Google Scholar] [CrossRef] [Green Version]
- Hales, B.A.; Edwards, C.; Ritchie, D.A.; Hall, G.; Pickup, R.W.; Saunders, J.R. Isolation and identification of methanogen-specific DNA from blanket bog peat by PCR amplification and sequence analysis. Appl. Environ. Microbiol. 1996, 62, 668–675. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108, 4516–4522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dowd, S.E.; Callaway, T.R.; Wolcott, R.D.; Sun, Y.; McKeehan, T.; Hagevoort, R.G.; Edrington, T.S. Evaluation of the bacterial diversity in the feces of cattle using 16S rDNA bacterial tag-encoded FLX amplicon pyrosequencing (bTEFAP). BMC Microbiol. 2008, 8, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; University Illinois Press: Urbana, IL, USA, 1949. [Google Scholar]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Worakit, S.; Boone, D.R.; Mah, R.A.; Abdel-Samie, M.E.; El-Halwagi, M.M. Methanobacterium alcaliphilum sp. nov., an H2-utilizing methanogen that grows at high pH values. Int. J. Syst. Evol. Microbiol. 1986, 36, 380–382. [Google Scholar] [CrossRef] [Green Version]
- Ma, K.; Liu, X.; Dong, X. Methanosaeta harundinacea sp. nov., a novel acetate-scavenging methanogen isolated from a UASB reactor. Int. J. Syst. Evol. Microbiol. 2006, 56, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Mori, K.; Iino, T.; Suzuki, K.I.; Yamaguchi, K.; Kamagata, Y. Aceticlastic and NaCl-requiring methanogen “Methanosaeta pelagica” sp. nov., isolated from marine tidal flat sediment. Appl. Environ. Microbiol. 2012, 78, 3416–3423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, S.; Ishii, S.I.; Wu, A.; Cheung, A.; Tenney, A.; Wanger, G.; Nealson, K.H. Microbial diversity in The Cedars, an ultrabasic, ultrareducing, and low salinity serpentinizing ecosystem. Proc. Natl. Acad. Sci. USA 2013, 110, 15336–15341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrill, P.L.; Kuenen, J.G.; Johnson, O.J.; Suzuki, S.; Rietze, A.; Sessions, A.L.; Nealson, K.H. Geochemistry and geobiology of a present-day serpentinization site in California: The Cedars. Geochim. Cosmochim. Act. 2013, 109, 222–240. [Google Scholar] [CrossRef] [Green Version]
- Kotelnikova, S.; Macario, A.J.; Pedersen, K. Methanobacterium subterraneum sp. nov., a new alkaliphilic, eurythermic and halotolerant methanogen isolated from deep granitic groundwater. Int. J. Syst. Evol. Microbiol. 1998, 48, 357–367. [Google Scholar] [CrossRef] [Green Version]
- Joulian, C.; Patel, B.K.; Ollivier, B.; Garcia, J.L.; Roger, P.A. Methanobacterium oryzae sp. nov., a novel methanogenic rod isolated from a Philippines ricefield. Int. J. Syst. Evol. Microbiol. 2000, 50, 525–528. [Google Scholar] [CrossRef] [Green Version]
- Quéméneur, M.; Palvadeau, A.; Postec, A.; Monnin, C.; Chavagnac, V.; Ollivier, B.; Erauso, G. Endolithic microbial communities in carbonate precipitates from serpentinite-hosted hyperalkaline springs of the Voltri massif (Ligurian Alps, Northern Italy). Environ. Sci. Pollut. Res. 2015, 22, 13613–13624. [Google Scholar] [CrossRef]
- Boulart, C.; Chavagnac, V.; Monnin, C.; Delacour, A.; Ceuleneer, G.; Hoareau, G. Differences in gas venting from ultramafic-hosted warm springs: The example of Oman and Voltri ophiolites. Ofioliti 2013, 38, 143–156. [Google Scholar]
- Blank, J.G.; Green, S.J.; Blake, D.; Valley, J.W.; Kita, N.T.; Treiman, A.; Dobson, P.F. An alkaline spring system within the Del Puerto Ophiolite (California, USA): A Mars analog site. Planet. Space. Sci. 2009, 57, 533–540. [Google Scholar] [CrossRef] [Green Version]
- Woycheese, K.M.; Meyer-Dombard, D.A.R.; Cardace, D.; Argayosa, A.M.; Arcilla, C.A. Out of the dark: Transitional subsurface-to-surface microbial diversity in a terrestrial serpentinizing seep (Manleluag, Pangasinan, the Philippines). Front. Microbiol. 2015, 6, 44. [Google Scholar] [CrossRef]
- Rempfert, K.R.; Miller, H.M.; Bompard, N.; Nothaft, D.; Matter, J.M.; Kelemen, P.; Fierer, N.; Templeton, A.S. Geological and geochemical controls on subsurface microbial life in the Samail Ophiolite, Oman. Front. Microbiol. 2017, 8, 56. [Google Scholar] [CrossRef] [Green Version]
- Miller, H.M.; Matter, J.M.; Kelemen, P.; Ellison, E.T.; Conrad, M.E.; Fierer, N.; Ruchala, T.; Tominiga, M.; Templeton, A.S. Modern water/rock reactions in Oman hyperalkaline peridotite aquifers and implications for microbial habitability. Geochim Cosmochim. Act. 2016, 179, 217–241. [Google Scholar] [CrossRef] [Green Version]
- Moser, D.P.; Gihring, T.M.; Brockman, F.J.; Fredrickson, J.K.; Balkwill, D.L.; Dollhopf, M.E.; Sherwood Lollar, B.; Pratt, L.M.; Boice, E.; Southam, G.; et al. Desulfotomaculum and Methanobacterium spp. dominate a 4-to 5-kilometer-deep fault. Appl. Environ. Microbiol. 2005, 71, 8773–8783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purkamo, L.; Bomberg, M.; Kietäväinen, R.; Salavirta, H.; Nyyssönen, M.; Nuppunen-Puputti, M.; Ahonen, L.; Kukkonen, I.; Itävaara, M. Microbial co-occurrence patterns in deep Precambrian bedrock fracture waters. Biogeosciences 2016, 13, 3091–3108. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Liu, X.; Dong, X. Methanobacterium movens sp. nov. and Methanobacterium flexile sp. nov., isolated from lake sediment. Int. J. Syst. Evol. Microbiol. 2011, 61, 2974–2978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wormald, R.M.; Rout, S.P.; Mayes, W.; Gomes, H.; Humphreys, P.N. Hydrogenotrophic Methanogenesis Under Alkaline Conditions. Front. Microbiol. 2020, 11, 614227. [Google Scholar] [CrossRef] [PubMed]
- Miller, H.M.; Chaudhry, N.; Conrad, M.E.; Bill, M.; Kopf, S.H.; Templeton, A.S. Large carbon isotope variability during methanogenesis under alkaline conditions. Geochim. Cosmochim. Acta 2018, 237, 18–31. [Google Scholar] [CrossRef] [Green Version]
- Ley, R.E.; Harris, J.K.; Wilcox, J.; Spear, J.R.; Miller, S.R.; Bebout, B.M.; Maresca, J.A.; Bryant, D.A.; Sogin, M.L.; Pace, N.R. Unexpected diversity and complexity of the Guerrero Negro hypersaline microbial mat. Appl. Environ. Microbiol. 2006, 72, 3685–3695. [Google Scholar] [CrossRef] [Green Version]
- Rinke, C.; Schwientek, P.; Sczyrba, A.; Ivanova, N.N.; Anderson, I.J.; Cheng, J.F.; Darling, A.; Malfatti, S.; Swan, B.K.; Gies, E.A.; et al. Insights into the phylogeny and coding potential of microbial dark matter. Nature 2013, 499, 431–437. [Google Scholar] [CrossRef] [Green Version]
- Peoples, L.M.; Donaldson, S.; Osuntokun, O.; Xia, Q.; Nelson, A.; Blanton, J.; Allen, E.E.; Church, M.J.; Bartlett, D.H. Vertically distinct microbial communities in the Mariana and Kermadec trenches. PLoS ONE 2018, 13, e0195102. [Google Scholar] [CrossRef] [Green Version]
- Borsodi, A.K.; Anda, D.; Makk, J.; Krett, G.; Dobosy, P.; Büki, G.; Eross, A.; Madl-Szonyi, J. Biofilm forming bacteria and archaea in thermal karst springs of Gellért Hill discharge area (Hungary). J. Basic Microbiol. 2018, 58, 928–937. [Google Scholar] [CrossRef] [PubMed]
- Zakharenko, A.S.; Galachyants, Y.P.; Morozov, I.V.; Shubenkova, O.V.; Morozov, A.A.; Ivanov, V.G.; Pimenov, N.V.; Krasnopeev, A.Y.; Zemskaya, T.I. Bacterial communities in areas of oil and methane seeps in pelagic of Lake Baikal. Microb. Ecol. 2019, 78, 269–285. [Google Scholar] [CrossRef]
- Sieber, C.M.; Paul, B.G.; Castelle, C.J.; Hu, P.; Tringe, S.G.; Valentine, D.L.; Anderson, G.L.; Banfield, J.F. Unusual metabolism and hypervariation in the genome of a Gracilibacterium (BD1-5) from an oil-degrading community. MBio 2019, 10, e02128-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.; Li, P.; Liu, S.; Miao, B.; Zeng, B.; Jiang, Y.; Li, L.; Wang, L.; Chen, Y.; Zhang, H. Characterization of the rumen microbiota and volatile fatty acid profiles of weaned goat kids under shrub-grassland grazing and indoor feeding. Animals 2020, 10, 176. [Google Scholar] [CrossRef] [Green Version]
- Cahill, A.G.; Steelman, C.M.; Forde, O.; Kuloyo, O.; Ruff, S.E.; Mayer, B.; Mayer, K.U.; Strous, M.; Ryan, M.C.; Cherry, J.A. Mobility and persistence of methane in groundwater in a controlled-release field experiment. Nat. Geosci. 2017, 10, 289–294. [Google Scholar] [CrossRef]
- Kuloyo, O.; Ruff, S.E.; Cahill, A.; Connors, L.; Zorz, J.K.; Hrabe de Angelis, I.; Nightingale, M.; Mayer, B.; Strous, M. Methane oxidation and methylotroph population dynamics in groundwater mesocosms. Environ. Microbiol. 2020, 22, 1222–1237. [Google Scholar] [CrossRef]
- Hanke, A.; Hamann, E.; Sharma, R.; Geelhoed, J.S.; Hargesheimer, T.; Kraft, B.; Meyer, V.; Lenk, S.; Osmers, H.; Wu, R.; et al. Recoding of the stop codon UGA to glycine by a BD1-5/SN-2 bacterium and niche partitioning between Alpha-and Gammaproteobacteria in a tidal sediment microbial community naturally selected in a laboratory chemostat. Front. Microbiol. 2014, 5, 231. [Google Scholar] [CrossRef]
- Hedlund, B.P.; Dodsworth, J.A.; Murugapiran, S.K.; Rinke, C.; Woyke, T. Impact of single-cell genomics and metagenomics on the emerging view of extremophile “microbial dark matter”. Extremophiles 2014, 18, 865–875. [Google Scholar] [CrossRef]
- Suzuki, S.; Kuenen, J.G.; Schipper, K.; Van Der Velde, S.; Ishii, S.I.; Wu, A.; Sorokin, D.Y.; Tenney, A.; Meng, X.Y.; Morill, P.L.; et al. Physiological and genomic features of highly alkaliphilic hydrogen-utilizing Betaproteobacteria from a continental serpentinizing site. Nat. Commun. 2014, 5, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brazelton, W.J.; Morrill, P.L.; Szponar, N.; Schrenk, M.O. Bacterial communities associated with subsurface geochemical processes in continental serpentinite springs. Appl. Environ. Microbiol. 2013, 79, 3906–3916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiago, I.; Veríssimo, A. Microbial and functional diversity of a subterrestrial high pH groundwater associated to serpentinization. Environ. Microbiol. 2013, 15, 1687–1706. [Google Scholar] [CrossRef]
- Marques, J.M.; Etiope, G.; Neves, M.O.; Carreira, P.M.; Rocha, C.; Vance, S.D. Linking serpentinization, hyperalkaline mineral waters and abiotic methane production in continental peridotites: An integrated hydrogeological-bio-geochemical model from the Cabeço de Vide CH4-rich aquifer (Portugal). Appl. Geochem. 2018, 96, 287–301. [Google Scholar] [CrossRef]
- Willis, G.; Nancucheo, I.; Hedrich, S.; Giaveno, A.; Donati, E.; Johnson, D.B. Enrichment and isolation of acid-tolerant sulfate-reducing microorganisms in the anoxic, acidic hot spring sediments from Copahue volcano, Argentina. FEMS Microbiol. Ecol. 2019, 95, fiz175. [Google Scholar] [CrossRef]
- Diez-Ercilla, M.; Sánchez-España, J.; Yusta, I.; Wendt-Potthoff, K.; Koschorreck, M. Formation of biogenic sulphides in the water column of an acidic pit lake: Biogeochemical controls and effects on trace metal dynamics. Biogeochemistry 2014, 121, 519–536. [Google Scholar] [CrossRef]
- Okubo, T.; Toyoda, A.; Fukuhara, K.; Uchiyama, I.; Harigaya, Y.; Kuroiwa, M.; Suzuki, T.; Murakami, Y.; Suwa, Y.; Takami, H. The physiological potential of anammox bacteria as revealed by their core genome structure. DNA Res. 2021, 28, dsaa028. [Google Scholar] [CrossRef] [PubMed]
- Mori, K.; Suzuki, K.I. Thiofaba tepidiphila gen. nov., sp. nov., a novel obligately chemolithoautotrophic, sulfur-oxidizing bacterium of the Gammaproteobacteria isolated from a hot spring. Int. J. Syst. Evol. Microbiol. 2018, 58, 1885–1891. [Google Scholar] [CrossRef] [Green Version]
- Ito, T.; Sugita, K.; Yumoto, I.; Nodasaka, Y.; Okabe, S. Thiovirga sulfuroxydans gen. nov., sp. nov., a chemolithoautotrophic sulfur-oxidizing bacterium isolated from a microaerobic waste-water biofilm. Int. J. Syst. Evol. Microbiol. 2005, 55, 1059–1064. [Google Scholar] [CrossRef] [Green Version]
- Huber, J.A.; Johnson, H.P.; Butterfield, D.A.; Baross, J.A. Microbial life in ridge flank crustal waters. Environ. Microbiol. 2006, 8, 88–99. [Google Scholar] [CrossRef] [PubMed]
- Kimura, H.; Nashimoto, H.; Shimizu, M.; Hattori, S.; Yamada, K.; Koba, K.; Yoshida, N.; Kato, K. Microbial methane production in deep aquifer associated with the accretionary prism in Southwest Japan. ISME J. 2010, 4, 531–541. [Google Scholar] [CrossRef] [PubMed]
- Adjeroud, M.; Escuder-Rodríguez, J.J.; González-Siso, M.I.; Kecha, M. Metagenomic Investigation of Bacterial and Archaeal Diversity of Hammam Essalihine Hot Spring from Khenchela, Algeria. GeoMicrobiol. J. 2020, 37, 804–817. [Google Scholar] [CrossRef]
- Boomer, S.M.; Noll, K.L.; Geesey, G.G.; Dutton, B.E. Formation of multilayered photosynthetic biofilms in an alkaline thermal spring in Yellowstone National Park, Wyoming. Appl. Environ. Microbiol. 2009, 75, 2464–2475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spear, J.R.; Walker, J.J.; McCollom, T.M.; Pace, N.R. Hydrogen and bioenergetics in the Yellowstone geothermal ecosystem. Proc. Natl. Acad. Sci. USA 2005, 102, 2555–2560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamran, A.; Sauter, K.; Reimer, A.; Wacker, T.; Reitner, J.; Hoppert, M. Cyanobacterial mats in calcite-precipitating serpentinite-hosted alkaline springs of the Voltri Massif, Italy. Microorganisms 2021, 9, 62. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, T.L.; Klatt, J.M.; De Beer, D.; Macalady, J.L. Cyanobacterial photosynthesis under sulfidic conditions: Insights from the isolate Leptolyngbya sp. strain hensonii. ISME J. 2018, 12, 568–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henry, E.A.; Devereux, R.; Maki, J.S.; Gilmour, C.C.; Woese, C.R.; Mandelco, L.; Schauder, R.; Remsen, C.C.; Mitchell, R. Characterization of a new thermophilic sulfate-reducing bacterium. Arch. Microbiol. 1994, 161, 62–69. [Google Scholar] [CrossRef]
- Sonne-Hansen, J.; Ahring, B.K. Thermodesulfobacterium hveragerdense sp. nov., and Thermodesulfovibrio islandicus sp. nov., two thermophilic sulfate reducing bacteria isolated from a Icelandic hot spring. Syst. Appl. Microbiol. 1999, 22, 559–564. [Google Scholar] [CrossRef]
- Haouari, O.; Fardeau, M.L.; Cayol, J.L.; Fauque, G.; Casiot, C.; Elbaz-Poulichet, F.; Hamdi, M.; Ollivier, B. Thermodesulfovibrio hydrogeniphilus sp. nov., a new thermophilic sulphate-reducing bacterium isolated from a Tunisian hot spring. Syst. Appl. Microbiol. 2008, 31, 38–42. [Google Scholar] [CrossRef]
- Frank, Y.A.; Kadnikov, V.V.; Lukina, A.P.; Banks, D.; Beletsky, A.V.; Mardanov, A.V.; Sen’kina, E.I.; Avakyan, M.R.; Karnachuk, O.V.; Ravin, N.V. Characterization and genome analysis of the first facultatively alkaliphilic Thermodesulfovibrio isolated from the deep terrestrial subsurface. Front. Microbiol. 2016, 7, 2000. [Google Scholar] [CrossRef] [PubMed]
- Tindall, B.J.; Sikorski, J.; Lucas, S.; Goltsman, E.; Copeland, A.; Del Rio, T.G.; Nolan, M.; Tice, H.; Cheng, J.F.; Han, C.; et al. Complete genome sequence of Meiothermus ruber type strain (21 T). Stand. Genom. Sci. 2010, 3, 26–36. [Google Scholar] [CrossRef] [Green Version]
- Loginova, L.G.; Egorova, L.A.; Golovacheva, R.S.; Seregina, L.M. Thermus ruber sp. nov., nom. rev. Int. J. Syst. Evol. Microbiol. 1984, 34, 498–499. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples 1 | CANBW | CANPW | CANAW | CANBS | CANPS | CANAS |
|---|---|---|---|---|---|---|
| Types | Water | Water | Water | Sediment | Sediment | Sediment |
| pH 2 | 9.0–9.1 | 9.0–9.1 | 9.0–9.1 | N/A 3 | N/A 3 | N/A 3 |
| Temperature (°C) 2 | 41 | 41 | 40 | N/A 3 | N/A 3 | N/A 3 |
| O2 (mg/L) 2 | 0.7 | 5.3 | 4.9 | N/A 3 | N/A 3 | N/A 3 |
| ORP (mV; ref. Ag/AgCl) 2 | −351 | −333 | −260 | N/A 3 | N/A 3 | N/A 3 |
| Shannon index (H) | 4.60 ± 0.03 | 3.70 ± 0.02 | 4.16 ± 0.02 | 5.46 ± 0.06 | 4.45 ± 0.60 | 5.02 ± 0.19 |
| Simpson index (D) | 0.965 ± 0.001 | 0.928 ± 0.001 | 0.916 ± 0.007 | 0.974 ± 0.008 | 0.941 ± 0.021 | 0.962 ± 0.020 |
| Bacterial 16S rDNA (copies/g or copies/L) | 5.50 × 107 | 3.04 × 107 | 1.06 × 109 | 6.97 × 109 | 2.42 × 109 | 6.02 × 109 |
| Archaeal 16S rDNA (copies/g or copies/L) | 2.31 × 106 | 1.92 × 106 | 1.63 × 107 | 5.38 × 107 | 6.59 × 107 | 8.69 × 107 |
| mcrA (copies/g or copies/L) | 2.23 × 105 | 1.17 × 105 | 3.14 × 106 | 1.95 × 107 | 8.17 × 106 | 2.19 × 107 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quéméneur, M.; Mei, N.; Monnin, C.; Postec, A.; Wils, L.; Bartoli, M.; Guasco, S.; Pelletier, B.; Erauso, G. Procaryotic Diversity and Hydrogenotrophic Methanogenesis in an Alkaline Spring (La Crouen, New Caledonia). Microorganisms 2021, 9, 1360. https://doi.org/10.3390/microorganisms9071360
Quéméneur M, Mei N, Monnin C, Postec A, Wils L, Bartoli M, Guasco S, Pelletier B, Erauso G. Procaryotic Diversity and Hydrogenotrophic Methanogenesis in an Alkaline Spring (La Crouen, New Caledonia). Microorganisms. 2021; 9(7):1360. https://doi.org/10.3390/microorganisms9071360
Chicago/Turabian StyleQuéméneur, Marianne, Nan Mei, Christophe Monnin, Anne Postec, Laura Wils, Manon Bartoli, Sophie Guasco, Bernard Pelletier, and Gael Erauso. 2021. "Procaryotic Diversity and Hydrogenotrophic Methanogenesis in an Alkaline Spring (La Crouen, New Caledonia)" Microorganisms 9, no. 7: 1360. https://doi.org/10.3390/microorganisms9071360