
Cis- and Trans-Encoded Small Regulatory RNAs in Bacillus subtilis

AG Bakteriengenetik, Matthias-Schleiden-Institut, Friedrich-Schiller-Universität Jena, D-07743 Jena, Germany
*
Author to whom correspondence should be addressed.
Microorganisms 2021, 9(9), 1865; https://doi.org/10.3390/microorganisms9091865
Submission received: 29 July 2021
/
Revised: 23 August 2021
/
Accepted: 24 August 2021
/
Published: 2 September 2021
(This article belongs to the Special Issue Gene Analysis in Bacillus subtilis)
Abstract
:Small regulatory RNAs (sRNAs) that act by base-pairing are the most abundant posttranscriptional regulators in all three kingdoms of life. Over the past 20 years, a variety of approaches have been employed to discover chromosome-encoded sRNAs in a multitude of bacterial species. However, although largely improved bioinformatics tools are available to predict potential targets of base-pairing sRNAs, it is still challenging to confirm these targets experimentally and to elucidate the mechanisms as well as the physiological role of their sRNA-mediated regulation. Here, we provide an overview of currently known cis- and trans-encoded sRNAs from B. subtilis with known targets and defined regulatory mechanisms and on the potential role of RNA chaperones that are or might be required to facilitate sRNA regulation in this important Gram-positive model organism.
1. Introduction
Small regulatory RNAs (sRNAs) that act by base-pairing constitute the main class of posttranscriptional regulators in bacteria. They can be divided into cis-encoded and trans-encoded sRNAs. Whereas cis-encoded sRNAs are transcribed from the same genetic locus in opposite direction to their single target RNA and are, therefore, completely complementary to this target RNA, trans-encoded sRNAs are transcribed from a promoter somewhere else on the bacterial chromosome and are only partially complementary to their target RNAs. Therefore, they can only form partial duplexes with their—often multitude of—target RNAs. In both cases, target RNA binding results in inhibition or activation of target gene expression. Cis- and trans-encoded sRNAs employ a variety of mechanisms of action for posttranscriptional gene regulation [1,2,3] which, eventually, result in altered translation or RNA stability.
A second class of sRNAs acts by protein binding. It comprises the ubiquitous 6S RNA that binds the RNA polymerase by mimicking an open promoter and helps to shift transcription from vegetative to stationary phase promoters (rev. in [4]), sRNAs like CsrB/CsrC that sequester small proteins like CsrA at conserved single-stranded GGA motifs and prevent them from binding to mRNAs where they regulate translation, RNA stability or transcript elongation (rev. in [5]), and plasmid ColE1-encoded Rcd that binds to and enhances the enzymatic activity of tryptophanase to delay cell division until plasmid dimers are resolved [6].
The first cis-encoded base-pairing sRNAs, i.e., bona fide antisense RNAs, were discovered in E. coli plasmids in 1981 and intensively studied as regulators of replication (rev. in [7]), conjugation, and maintenance of plasmids (rev. [1,8]), as well as transposition and transduction (rev. in [1,3]). Only in 2001, two bioinformatics and microarray-based approaches followed by Northern blotting led to the discovery of 14 and 17 trans-encoded sRNAs from the E. coli chromosome, respectively [9,10]. Since then, in almost all bacterial species, such approaches were used (rev. in [11]), and computer programs for target prediction were established (rev. in [12]). Hitherto-discovered mechanisms of action of base-pairing sRNAs include translation inhibition by induction of structural changes around the RBS (ribosome binding site), direct blocking of the RBS or a ribosome standby site, combined translation inhibition and mRNA decay, degradation, processing, or stabilization of mRNAs, target mRNA trapping, or induction of premature transcription termination at Rho-independent or -dependent transcription terminators (rev. in [3]). In addition, a few antisense RNAs act only in cis by transcriptional interference [2], and some plasmid-encoded antisense RNAs prevent the formation of an activator pseudoknot that is required for target mRNA translation [1]. Some of these mechanisms were found both in Gram-negative and Gram-positive bacteria, others only in either of them. However, since many more sRNAs have been investigated to date in Gram-negative species such as E. coli or Salmonella enterica in comparison to Gram-positive species, it is too early to draw conclusions about the prevalence of certain mechanisms in either of them. The only obvious major difference between sRNA regulation in Gram-negative and Gram-positive bacteria is the requirement of RNA chaperones, which is discussed further below.
Two transcriptomics approaches for the prediction of small RNAs in B. subtilis were performed in 2009 and 2010 [13,14], which in total discovered 108 putative trans-encoded sRNAs. However, in contrast to Gram-negative bacteria like E. coli or Salmonella enterica [15], the number of B. subtilis sRNAs with identified targets is still very limited. In this review, we summarize our current knowledge on all cis- and trans-encoded sRNAs from B. subtilis, for which targets have been identified and physiological functions elucidated. We do not include protein-binding sRNAs or riboswitches in our review.
2. Base-Pairing sRNAs
2.1. Cis-Encoded sRNAs (Bona Fide Antisense RNAs) in B. subtilis
Currently, 10 systems of cis-encoded sRNAs and their corresponding targets are known in Bacillus subtilis (see Table 1 and Figure 1), and five of them are type I toxin-antitoxin systems. In total, three mechanisms of target gene regulation were identified: induction of target mRNA degradation, inhibition of translation initiation, and transcriptional interference.
2.1.1. Antitoxins in Type I Toxin–Antitoxin Systems
Toxin–antitoxin (TA) systems are genetic modules comprising a toxin gene and the counteracting antitoxin. These systems are widespread among bacteria and archaea—Both on the chromosome and on plasmids or phages to ensure the stable inheritance of DNA elements. In type I TA systems, the antitoxin is a cis-encoded sRNA that interacts with the toxin mRNA. All antitoxins either induce the degradation of the toxin mRNA, inhibit the toxin translation, or combine both mechanisms. The expression of the antitoxin has to be relatively strong to efficiently suppress the toxin expression. The toxins contain typically at least one transmembrane domain and are assumed to act as a small pore-forming protein. So far, only four type I TA systems were investigated in detail: txpA/RatA, bsrG/SR4, bsrE/SR5, and yonT/yoyJ/SR6 (rev. in [16]). A further 10 potential TA systems were predicted but not yet verified [17].
The txpA/RatA system encodes the small toxin TxpA (59 aa) and its antitoxin RatA [18]. In the absence of RatA, TxpA causes cell lysis on agar plates after five days, and txpA overexpression drastically impairs growth. The toxin contains a trans-membrane domain followed by charged aa and likely disturbs membrane integrity by pore formation. The antitoxin RatA, encoded on the opposite strand, overlaps the txpA transcript by ca. 120 nt at the 3′ end. The txpA RNA–RatA complex is a target for RNase III. This role in toxin mRNA degradation makes RNase III essential for B. subtilis [19]. Binding of RatA does not influence the txpA translation efficiency but exclusively promotes the toxin RNA degradation. The txpA/RatA system is located on the skin element, which is excised during the sporulation process and could prevent loss of this region even during a longer period of vegetative growth to preserve the ability to sporulate [18]. Moreover, TxpA plays a role in biofilm formation by elimination of defective and malformed cells [20].
The bsrG/SR4 system is encoded on the SPβ prophage region [21]. The toxin BsrG (38 aa) comprises a transmembrane domain and is similar to the N-terminal part of TxpA. BsrG accumulates in the membrane but does not permeabilize it. Instead, it interferes with the cell-envelope synthesis and causes cell-wall defects, membrane invaginations, and an altered cell shape [22]. Deletion of the antitoxin gene sr4 leads to cell lysis after overnight incubation at 37 °C [21]. The antitoxin SR4 binds to the 3′ end of bsrG mRNA, forming a perfect duplex of 123 bp. SR4 is bifunctional: it induces i) RNase III-dependent degradation of the bsrG mRNA and ii) a refolding around the RBS (ribosome binding site) that inhibits translation initiation [23]. Both binding kinetics and binding pathway were investigated in-depth, indicating three consecutive interactions of SR4 stem-loops with the corresponding bsrG mRNA regions [23]. The initial interaction site of bsrG mRNA comprises a YUNR motif in a loop, a sequence previously shown to be able to form a U-turn [24] that facilitates RNA–RNA interactions. Interestingly, the toxin mRNA is rapidly degraded upon heatshock, and both RNases Y and J1 contribute to its degradation [25].
The bsrE/SR5 system is located on the prophage-like element P6 in the chromosome [17]. The toxin BsrE (30 aa) only consists of a transmembrane domain and is less toxic than the other type I TA system toxins. The absence of the antitoxin SR5 is not sufficient to induce toxic effects of BsrE. Only overexpression of bsrE from a multicopy plasmid causes cell lysis on agar plates and inhibits growth significantly [26]. The antitoxin SR5 encoded on the opposite strand overlaps bsrE by 114 nt at the 3′ end. Complex formation of both RNAs leads to degradation of bsrE mRNA by RNase III [27]. In contrast to bsrG/SR4, no direct translational inhibition of bsrE mRNA by SR5 was observed. The interaction pathway between bsrE mRNA and SR5 was investigated in detail [27]: the initial contact is formed between two loops that both contain YUNR motifs, enabling a highly efficient and rapid complex formation. The expression of bsrE and sr5 is regulated differently in response to a multitude of stress factors. The otherwise very stable bsrE mRNA is sensitive to ethanol and alkaline stress as well as heatshock, while SR5 amounts differ under acid and alkaline stress, iron limitation, and anaerobic conditions. Interestingly, SR5 is rapidly degraded under oxygen deficiency, which allows bsrE expression. Therefore, this TA system could play a role in the response of B. subtilis to anaerobic conditions [26].
The yonT/yoyJ/SR6 system is very special, because it contains two toxin genes that are regulated by the same antitoxin. YonT (59 aa) is a very strong toxin, consisting of an N-terminal transmembrane domain and a highly positively charged C-terminal region. The deletion of the antitoxin gene sr6 causes cell lysis. YoyJ (83 aa) is a very weak toxin with two transmembrane domains. In the absence of SR6, yoyJ overexpression is detrimental to the cells. The antitoxin SR6 uses two distinct mechanisms to repress yonT and yoyJ: SR6 and yonT mRNA interact at their 3′ ends inducing RNA degradation and, therefore, toxin inactivation by RNase III [19]. The interaction between SR6 and yoyJ mRNA involves both 5′ ends and does not influence the yoyJ mRNA stability. Instead, SR6 inhibits the yoyJ translation by blocking the RBS [28]. Surprisingly, the sr6 promoter is much weaker than the yonT promoter, which is unfavorable to generate an excess of the antitoxin. However, this is compensated for by an unusually stable antitoxin. The TA system is located on the SPβ prophage region of the chromosome and, like bsrE/SR5, both RNAs are impacted by several stress conditions.
An excess of yonT mRNA under heatshock or manganese stress suggests a role of the system in the cellular stress response [28].
The bsrH/as-bsrH system is a potential type I TA system [14,29]. The toxin BsrH (29 aa) is very similar to BsrE, indicating a common origin. So far, the toxicity of BsrH was not proven experimentally. The bsrH mRNA interacts with the antitoxin as-bsrH at the 3′ end and is degraded by RNase J1 [17]. The system is located on the skin element directly adjacent to, but independent of, the txpA/RatA system.
2.1.2. Other Cis-Encoded sRNAs in B. subtilis
The non-coding RNA S25 is an antisense RNA regulating the gene yabE. S25 is transcribed under control of the extracytoplasmic sigma factors σM and σX in response to cell envelope stress, e.g., the presence of antibiotics. The amount of yabE RNA is reduced during S25 expression, which may be due to RNase III-dependent degradation [30]. The function of YabE is so far unknown, but a potential G5 domain (N-acetylglucosamine binding domain) suggests a function at the cell wall and therefore a role in the response to antibiotics.
The sRNA S1559 is transcribed convergently to the gdpP gene on the opposite strand. GdpP is a membrane-localized c-di-AMP specific phosphodiesterase [31], which is involved in the control of sporulation initiation and cell-wall homeostasis. The S1559 promoter is σD-dependent, suggesting a role in or during motility regulation. The expression of S1559 reduces the amount of cellular GdpP but does not influence motility or antibiotic resistance [29]. The mechanism of S1559 action is still elusive.
The antisense RNA S1326 is transcribed from a σB-dependent promoter. The transcript was detected after phosphate stress, is of heterogeneous length, and is complementary to the cwlO gene. CwlO is an extracellular D,L-endopeptidase-type autolysin involved in the cell wall synthesis and mainly active in the exponential growth phase [32]. Due to rapid RNase Y-dependent degradation, the cwlO transcript is very unstable. How the antisense RNA influences the cwlO expression and the biological role of this regulation are so far unknown. The only effect observed was a slightly faster entry into exponential growth after resuspension in fresh medium [33].
Another σB-dependent antisense RNA is S1290, which is complementary to opuB mRNA. OpuB is a choline importer required to maintain the osmotic homeostasis under high salinity conditions. The transiently transcribed S1290 acts by transcriptional interference to repress a fast and strong opuB induction after osmotic shock. This delays the opuB expression in favor of the expression of opuC, which encodes an alternative but more general osmoprotectant import system. Therefore, S1290 is involved in fine-tuning the salt stress response [34].
The sRNA SR7 (S1136) is a dual-function antisense RNA that on the one hand interferes with rpsD transcription and on the other hand is an mRNA encoding the small protein SR7P (39 aa). This small protein interacts with the glycolytic enzyme enolase, thereby promoting its recruitment to RNase Y to enhance the RNase Y activity on at least two substrate RNAs, rpsO mRNA, and the 5′ UTR of yitJ mRNA [35]. Consequently, SR7P plays a role in RNA degradation [35,36]. SR7 only acts in cis to reduce the amount of rpsD mRNA, most likely by transcriptional interference [37]. RpsD (22.7 kDa) is the primary RNA binding protein S4 of the 30S ribosomal subunit. Therefore, a reduction of the SR4 amount finally decreases the number of active ribosomes. The σB-dependent transcription of sr7 is strongly induced under ethanol stress [37] as well as salt, acid, heat, and manganese stress conditions [35]. In addition, SR7P affects cell survival under stress conditions, indicating that it might play a specific role in stress response.
For none of the five antisense RNAs were the secondary structures determined or, in case they base-pair with their target RNAs, the binding pathways elucidated.
2.2. Trans-Encoded sRNAs in B. subtilis
At present, five trans-encoded sRNAs are known in B. subtilis for which targets were identified: SR1 [38,39], FsrA [40], RoxS [41,42], RosA [43], and RnaC [44] (see Figure 2). Among them, SR1 is the only dual-function sRNA that acts as a base-pairing sRNA and as mRNA encoding a small protein, SR1P (see below). In Table 2, an overview of all currently known trans-encoded sRNAs in B. subtilis and their targets is provided.
2.2.1. SR1
SR1 was the first trans-encoded sRNA discovered in B. subtilis [38]. A computational approach employed to search for putative sRNAs in intergenic regions of the B. subtilis chromosome yielded 20 candidate sRNAs, among them SR1, which was subsequently confirmed by Northern blotting [38]. The sr1 gene is transcribed under gluconeogenic conditions from the σA-dependent promoter psr1 and repressed under glycolytic conditions mainly by CcpN and, to a smaller extent, by CcpA [38]. Whereas CcpA binds at a cre site about 260 bp upstream of psr1, CcpN binds at two sites, directly upstream of the -35 box and in the spacer region of psr1, and requires two ligands, ATP and H+ (slightly acid pH), for a 20- to 30-fold repression of sr1 transcription [47,48]. CcpN interacts with the α-subunit of the RNA polymerase to prevent promoter escape [49]. The first identified target of SR1 was ahrC mRNA [39], encoding the transcriptional activator of the arginine catabolic operons rocABC and rocDEF and the transcriptional repressor of the arginine biosynthesis genes [50]. SR1 shares seven complementary regions with ahrC mRNA, designated A to G in SR1 and A’ to G’ in ahrC mRNA, located in the central portions of both RNAs [39]. Binding starts between region G of SR1 and region G’ 97 nt downstream of the ahrC RBS and inhibits translation initiation by a novel mechanism: SR1 induces structural changes 20 to 40 nt downstream of the ahrC RBS that obstruct binding of the ribosomal 30S subunit [45].
Although the G/G’ interaction is decisive, the complementary regions B/B’ to F/F’ also contribute to SR1/ahrC RNA complex formation [45]. SR1 barely affects the stability of ahrC mRNA [39,46], excluding the recruitment of an RNase as a primary mechanism of sRNA action. The RNA chaperone Hfq binds both SR1 and ahrC mRNA but does not promote their interaction. Instead, it is required for ahrC translation by opening a secondary structure 5′ of the RBS. By contrast, the RNA chaperone CsrA that also binds to both RNAs facilitates their interaction by slightly altering the ahrC mRNA structure around the G’ region to support SR1 binding [46] (see Figure 2).
In addition to being a trans-encoded sRNA that acts by a base-pairing mechanism, SR1 is an mRNA that codes for a small protein of 39 aa [51]. This protein, designated SR1P, interacts with the glycolytic GapA (glyceraldehyde-3 phosphatedehydrogenase A, one of the two Gap proteins in B. subtilis), promotes the interaction of GapA with RNase J1, and enhances the enzymatic activity of RNase J1 on two substrate RNAs [52]. The SR1P/GapA interaction surface has been elucidated [53]. So far, SR1 is the only dual-function sRNA with functions in different physiological pathways, arginine metabolism, and RNA degradation [36,54] (see Figure 3). An experimental analysis of 9 of the 23 SR1 homologues detected in 2012 demonstrated that both the base-pairing and the protein-encoding functions of SR1 are highly conserved over one billion years of evolution [55]. In the meantime, many more bacterial genomes have been sequenced, and a new search in 2021 yielded 139 SR1/SR1P homologues, all of them confined to the order Bacillales.
Recently, we discovered a second target of SR1, kinA mRNA, encoding the major histidine kinase of the sporulation phosphorelay [56]. Upon starvation and stress, KinA autophosphorylates and transfers its phosphate via Spo0F and Spo0B to the central sporulation regulator, the transcription factor Spo0A. Like SR1 and ahrC mRNA, SR1 and kinA mRNA share seven complementary regions termed A to G in SR1 and A’ to G’ in kinA mRNA, the latter including regions upstream and downstream of the RBS as well as within the 5′ part of the ORF. The SR1–kinA RNA interaction starts at region D’ 10 nt downstream of the kinA RBS and causes translation inhibition, although the RBS itself is not complementary to SR1.
The SR1 D region is located in the central single-stranded part of the molecule, whereas the SR1-ahrC mRNA interaction starts within the left arm of the SR1 transcription terminator (see Figure 2A). Similar to the SR1/ahrC system, SR1 does not affect the half-life of kinA mRNA. Accordingly, SR1P that stimulated the degradation of two RNase J1 substrates via its binding to GapA did not affect kinA regulation. Surprisingly, although CsrA binds both SR1 and kinA mRNA, it neither promotes their interaction nor affects the regulation of KinA/Spo0A downstream targets or sporulation in vivo. Hfq was also dispensable for SR1-mediated kinA regulation and sporulation. Therefore, it cannot be ruled out that another, still unidentified RNA chaperone might fulfil the function of Hfq or CsrA in the SR1/kinA system. Interestingly, by controlling kinA, SR1 increases the time window for sporulation, allowing the formation of high-quality spores with a proper coat and crust. This function is only required under starvation conditions.
2.2.2. FsrA
The search for additional transcription units regulated by the ferric uptake regulator Fur yielded FsrA, the second small trans-encoded B. subtilis sRNA, as well as three small basic proteins FbpA, FbpB, and FbpC [40]. The 84 nt long FsrA is a functional homolog of the E. coli sRNA RyhB. Iron-deplete conditions lead to derepression of the Fur-dependent fsrA promoter and allow FsrA to inhibit translation of target mRNAs involved in iron metabolism and storage. Among the first identified FsrA targets were the succinate dehydrogenase sdhCAB, the aconitase citB and lutABC encoding oxidases important for growth on lactate as sole carbon source.
A transcriptome analysis revealed in 2012 that FsrA is a global regulator with many targets, among them the gltAB (glutamate synthase), dicarboxylate transporter (dctP), the extracytoplasmic thioreductase resA, leuABCD (leucine biosynthesis), and menaquinolcytochrome c oxidoreductase qcrA genes [57]. On the basis of RNAhybrid and M-fold predictions, the authors suggested that a C-rich single-stranded (CRR) region of FsrA base-pairs with the RBS of its target mRNAs [58]. Hitherto, in vivo base-pairing could be only demonstrated between FsrA and the 5′ UTR of gltAB mRNA (Figure 2B). However, mutations that confirmed base-pairing did not involve CRR1 or CRR2, but three complementary regions upstream of them. In addition, EMSAs showed in vitro base-pairing between FsrA and sdhCAB mRNA. For the other target mRNAs, effects on mRNA and protein levels were reported, but the predicted base-pairing interactions [40] have yet to be confirmed in vivo to demonstrate that they are primary and not downstream targets of FsrA.
Whereas the functionally related Fur-regulated sRNA RyhB from E. coli requires the RNA chaperone Hfq, FsrA cooperates with one, two, or three Fur-regulated small basic proteins FbpA (54 aa), FbpB (48 aa), and FbpC (29 aa) postulated to be RNA chaperones [40]. However, RNA binding has not yet been confirmed for any of them. A concerted action of FsrA and FbpB in the inhibition of lutABC mRNA has been shown, which allows the direction of iron to higher priority target proteins [58]. The lutABC mRNA levels were affected approximately two-fold by both FsrA and FbpB with FbpB having a higher effect on RNA levels. Only small effects on the LutA, LutB, and LutC protein levels were observed. Whereas FsrA might directly repress lutABC translation by a base-pairing interaction, FbpB might facilitate the FsrA-lutABC RNA interaction since the role of FbpB could be bypassed by modest upregulation of fsrA. Alternatively, FbpB might recruit an RNase for lutABC mRNA degradation. So far, experimental evidence for any of these mechanisms is still lacking. For the repression of sdhCAB mRNA and citB mRNA by FsrA, all three Fbp proteins seem to be dispensable as has been shown by a proteomics study with fsrA and fbp mutants [58].
2.2.3. RoxS
RoxS (originally termed RsaE, 115 nt) is, so far, the only base-pairing sRNA that is conserved between B. subtilis and S. aureus. Transcription of roxS is activated in response to nitric oxide by the two-component system ResDE [41] and inhibited by the NADH-sensitive transcription repressor Rex [42]. The repression is released by malate. The physiological function of RoxS is to help restore the NAD+/NADH balance by temporarily turning down part of the TCA (tricarboxylic acid) cycle. So far, four direct RoxS targets have been experimentally confirmed: ppnKB mRNA encoding an NAD+/NADH kinase, sucCD mRNA encoding succinyl-CoA synthase [41], yflS encoding one of four malate transporters transcribed under control of MalKR that senses malate [42], and, recently, acsA encoding acetyl CoA synthase [43]. In all cases, RoxS uses several of its four C-rich regions for target binding, among them CRR3 (nt 51-64), which was shown to be decisive. However, RoxS employs different mechanisms of action: It inhibits translation and promotes degradation of ppnKB mRNA (Figure 2C) and sucCD mRNA. The RoxS–ppnKB mRNA duplex is substrate for the double-strand specific RNase III, whereas RNase Y cleaves ppnKB mRNA both RoxS-dependently and -independently and RoxS at nt +20, yielding the truncated species RoxS(D). The latter is required for inhibition of sucCD translation but can also efficiently regulate ppnKB. For acsA, fourfold-reduced mRNA levels in the presence of RoxS have been observed [43], but effects on translation were not analyzed. In the case of yflS, RoxS disrupts a structure at the 5′ UTR that obstructs ribosome binding to activate translation. Furthermore, it stabilizes yflS mRNA about 3.6-fold against 5′-3′ exoribonucleolytic degradation by RNase J1 [42] (Figure 2C). This effect of an sRNA on target mRNA stability is rather an exception, as the majority of bacterial sRNAs examined to date, among them also SR1 and FsrA, directly impact mRNA translation with only an indirect or no effect on the mRNA half-life (rev. in [3]). Translation activation of yflS mRNA is independent of its protection against 5′-3′ exoribonucleolytic degradation by RNase J1 [42]. Likewise, the RoxS effects on translation and stability of sucCD mRNA can be uncoupled [42].
Similar to SR1 and FsrA, RoxS does not require Hfq, at least for the regulation of ppnKB mRNA [41].
2.2.4. RosA
Recently, a sponge RNA, RosA (regulator of sRNA A) was identified that is transcribed from a σA-dependent promoter under control of the central regulator of carbon catabolism, CcpA [43]. Northern blotting revealed four RosA species (225, 193, 128, and 92 nt), the generation of which requires a complex network of different, partially yet unknown, endo- and exoribonucleases. The 225 and the 193 nt species contain three G-rich regions (GRR1-3) with potential complementarity to the C-rich regions of RoxS and FsrA. EMSAs showed that RosA can interact with RoxS and FsrA in vitro. IntaRNA predicted two complementary regions between RosA and RoxS, one comprising 15 consecutive bases involving nt 28-42 (with C-rich region CRR1) of RoxS, the other 10 consecutive bases involving nt 58-68 of RoxS with CCR3 that is decisive for binding of target mRNAs. Mutations in RoxS and RosA species revealed that in vitro, RoxS CRR1 complementary to RosA GRR2 is key to their interaction. In vivo, RosA binding impacts RoxS levels, reduces its half-life fivefold from 30 to 6 min, and promotes processing into the truncated RoxS(D) species. The decreased amount and half-life of three RoxS target RNAs, ppnKB mRNA, acsA mRNA, and yrhF mRNA (a new target RNA) in a ΔrosA strain corroborated the hypothesis that RosA sequesters RoxS [43]. Accordingly, an in vivo RNA quantification in B. subtilis grown in LB medium yielded a 1:1 RoxS/RosA ratio, indicating that even a complete out-titration of RoxS by RosA would be possible. The RosA effects on RoxS target mRNA levels were substantiated by proteomics confirming altered protein levels for PpnKB, SucC, SucD, AcsA, and YrhF in the absence of RosA. Interestingly, RoxS also plays a role in RosA processing, affecting at least the abundance of the 128 nt RosA species.
Another interesting question raised by the authors is whether the RosA/RoxS duplex might be a reservoir for RoxS or the truncated RoxS(D) species, from which the sRNA can be recycled for base-pairing with new target RNAs for which it has a greater affinity.
The CcpA-dependent RosA regulation places RoxS that is controlled by malate via transcription factor Rex at the key position of central metabolism in response to different carbon sources. The importance of RosA for B. subtilis under oxidative respiration conditions was demonstrated by co-cultivation of a wild-type and a ΔrosA strain: the ΔrosA strain was rapidly outcompeted due to the RoxS- and FsrA-mediated reduction of TCA cycle enzyme levels, which did not allow the mutant to generate ATP as quickly as the wild-type [43].
Why does RoxS, which is regulated by two transcription factors, ResD and Rex, need an additional level of posttranscriptional control by a sponge RNA? The authors argue that the long half-life of 30 min in a ΔrosA strain would not allow RoxS to respond quickly to altered environmental conditions and that, therefore, RosA might be required to rapidly downregulate RoxS activity first by neutralizing the C-rich regions required for target RNA binding and afterwards by promoting RoxS degradation [43].
The RosA–FsrA interaction involves RosA GRR2 as well, which binds to CRR2 of FsrA, as also shown in vitro by EMSAs. Proteomics revealed the expected alterations in the amount of proteins encoded by FsrA target mRNAs, among them CitB, SdhA, and CitZ.
Whereas the currently known sponge RNAs from Gram-negative bacteria are either mRNAs [59] or derive from the 3′ UTR of mRNAs [60], a similar stand-alone sRNA sponge named RsaI has been proposed for the staphylococcal RoxS homologue RsaE [61].
RosA contains a seven codon ORF that was associated with ribosomes in a ribosome profiling study [62]. Since the SD for this ORF is located in GRR2, RoxS binding would repress translation of this short ORF.
2.2.5. RnaC
RnaC/S1022 was first identified in a microarray screen of B. subtilis intergenic regions. However, so far it has not been visualized in Northern blots. This sRNA is only transcribed during the exponential growth phase under control of σD. Compensatory mutations introduced into predicted complementary loop regions revealed that RnaC base-pairs with abrB mRNA at the RBS and the first six codons to modulate the cellular level of transition state regulator AbrB [44] (Figure 3D). The RnaC-abrB RNA interaction that did not require Hfq was shown to support degradation of abrB mRNA and proposed to inhibit its translation. However, in ΔrnaC strains grown in LB medium, only very small alterations (≈33%) of abrB mRNA and AbrB levels were detected. The authors observed an enhanced cell-to-cell variation of AbrB levels, which finally resulted in growth rate heterogeneity within one population of logarithmically growing cells. They hypothesized that the subpopulations of fast- and slow-growing B. subtilis cells reflect a bet-hedging strategy for enhanced survival of unfavorable conditions since slowly growing cells are less susceptible to harsh environmental conditions [44].
3. Conclusions
Although more than 100 sRNAs have been discovered by transcriptome analyses in B. subtilis [13,14], and a small number have been found by other approaches [29,38,63], only 10 cis-encoded and five trans-encoded sRNAs have been characterized in detail. For each of the trans-encoded sRNAs, several interaction partners have been identified and biological functions in different areas elucidated. However, for the majority of the sRNAs, targets have yet to be identified and perhaps new mechanisms of action have yet to be unraveled. In addition, the recent discovery of the sponge RNA RosA [43] indicates that large networks exist that include both novel and already known regulatory RNAs.
In contrast to Gram-negative bacteria, B. subtilis does not seem to rely on two general RNA chaperones to promote the majority of sRNA-target RNA interactions or to stabilize sRNAs but seems to employ several RNA-binding proteins depending not only on the corresponding sRNA, but the specific sRNA/target RNA system, as illustrated by SR1/ahrC and SR1/kinA. A global approach is required to find out if the function of CsrA is confined to the SR1/ahrC system or if it plays a broader role in B. subtilis similar to Hfq and ProQ in Gram-negative bacteria like E. coli or Salmonella enteric [64]. Moreover, it has to be experimentally validated if FbpA, FbpB, and FbpC [40,58] are indeed RNA chaperones. Hitherto, only two bifunctional regulatory RNAs, SR1 and SR7, have been identified in B. subtilis (see Figure 3), but an increase in the number of such RNAs can be anticipated. Dual-function sRNAs and antisense RNAs comprising small ORFs that are translated under certain conditions will allow incorporating small proteins into extensive regulatory RNA–RNA networks. Beyond that, we hypothesize that sRNAs will be discovered that bind and modulate enzymes or act directly on the Bacillus subtilis chromosome like a number of eukaryotic siRNAs.
Author Contributions
Funding
This research was funded by the DFG (Deutsche Forschungsgemeinschaft), grant number BR1552/12-1 (to Sabine Brantl).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We acknowledge our colleague, Inam Ul Haq, for the experimental work on the role of SR7P and for critical reading of the manuscript. In addition, we thank all former coworkers of the AG Bakteriengenetik for their substantial experimental contributions.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Brantl, S. Regulatory mechanisms employed by cis-encoded antisense RNAs. Curr. Opin. Microbiol. 2007, 10, 102–109. [Google Scholar] [CrossRef]
- Brantl, S. Acting antisense: Plasmid- and chromosome-encoded sRNAs from Gram-positive bacteria. Future Microbiol. 2012, 7, 853–871. [Google Scholar] [CrossRef]
- Wagner, E.G.; Romby, P. Small RNAs in bacteria and archaea: Who they are, what they do, and how they do it. Adv. Genet. 2015, 90, 133–208. [Google Scholar] [PubMed]
- Ul Haq, I.; Müller, P.; Brantl, S. Intermolecular communication in Bacillus subtilis: RNA-RNA, RNA-protein and small protein-protein interactions. Front. Mol. Biosci. 2020, 7, 178. [Google Scholar] [CrossRef] [PubMed]
- Pourciau, C.; Lai, Y.J.; Gorelik, M.; Babitzke, P.; Romeo, T. Diverse mechanisms and circuitry for global regulation by the RNA binding protein CsrA. Front. Microbiol. 2020, 11, 601352. [Google Scholar] [CrossRef] [PubMed]
- Chant, E.L.; Summers, D.K. Indole signalling contributes to the stable maintenance of Escherichia coli multicopy plasmids. Mol. Microbiol. 2007, 63, 35–43. [Google Scholar] [CrossRef]
- Brantl, S. Plasmid-replication control by antisense RNAs. Microbiol. Spec. 2014, 2, PLAS-0001-2013. [Google Scholar] [CrossRef] [Green Version]
- Brantl, S. Antisense RNAs in plasmids: Control of replication and maintenance. Plasmid 2002, 48, 165–173. [Google Scholar] [CrossRef]
- Argaman, L.; Herhsberg, R.; Vogel, J.; Bejerano, G.; Wagner, E.G.; Margalit, H.; Altuvia, S. Novel small RNA-encoding genes in the intergenic regions of Escherichia coli. Curr. Biol. 2001, 11, 941–950. [Google Scholar] [CrossRef] [Green Version]
- Wassarman, K.M.; Repoila, F.; Rosenow, C.; Storz, G.; Gottesman, S. Identification of novel small RNAs using comparative genomics and microarrays. Genes Dev. 2001, 15, 1637–1651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, P.; Storz, G. Prevalence of small base-pairing RNAs derived from diverse genomic loci. Biochim. Biophys. Acta Gene Regul. Mech. 2020, 1863, 194524. [Google Scholar] [CrossRef]
- Georg, J.; Lalaouna, G.J.; Hou, S.; Lott, S.C.; Caldelari, I.; Marzi, S.; Hess, W.R.; Romby, P. The power of cooperation. Experimental and computational approaches in the functional characterization of bacterial sRNAs. Mol. Microbiol. 2020, 113, 603–612. [Google Scholar] [CrossRef]
- Rasmussen, S.; Nielsen, H.B.; Jarmer, H. The transcriptionally active regions in the genome of Bacillus subtilis. Mol. Microbiol. 2009, 73, 1043–1057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irnov, I.; Sharma, C.M.; Vogel, J.; Winkler, W.C. Identification of regulatory RNAs in Bacillus subtilis. Nucleic Acids Res. 2010, 38, 6637–6651. [Google Scholar] [CrossRef]
- Hör, J.; Matera, G.; Vogel, J.; Gottesman, S.; Storz, G. Trans-acting small RNAs and their effects on gene expression in Escherichia coli and Salmonella enterica. EcoSal Plus 2020, 9, 1–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brantl, S.; Müller, P. Toxin-antitoxin systems in Bacillus subtilis. Toxins 2019, 11, 262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durand, S.; Jahn, N.; Condon, C.; Brantl, S. Type I toxin-antitoxin systems in Bacillus subtilis. RNA Biol. 2012, 9, 1491–1497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silvaggi, J.M.; Perkins, J.B.; Losick, R. Small untranslated RNA antitoxin in Bacillus subtilis. J. Bacteriol. 2005, 187, 6641–6650. [Google Scholar] [CrossRef] [Green Version]
- Durand, S.; Gillet, L.; Condon, C. The essential function of B. subtilis RNase III is to silence foreign toxin genes. PLoS Genet. 2012, 8, e1003181. [Google Scholar] [CrossRef] [Green Version]
- Bloom-Ackermann, Z.; Steinberg, N.; Rosenberg, G.; Oppenheimer-Shaanan, Y.; Pollack, D.; Ely, S.; Storzi, N.; Levy, A.; Kolodkin-Gal, I. Toxin-Antitoxin systems eliminate defective cells and preserve symmetry in Bacillus subtilis biofilms. Environ. Microbiol. 2016, 18, 5032–5047. [Google Scholar] [CrossRef] [Green Version]
- Jahn, N.; Preis, H.; Wiedemann, C.; Brantl, S. BsrG/SR4 from Bacillus subtilis - the first temperature-dependent type I toxin-antitoxin system. Mol. Microbiol. 2012, 83, 579–598. [Google Scholar] [CrossRef]
- Jahn, N.; Brantl, S.; Strahl, H. Against the mainstream: The membrane-associated type I toxin BsrG from Bacillus subtilis interferes with cell envelope biosynthesis without increasing membrane permeability. Mol. Microbiol. 2015, 98, 651–666. [Google Scholar] [CrossRef] [PubMed]
- Jahn, N.; Brantl, S. One antitoxin—Two functions: SR4 controls toxin mRNA decay and translation. Nucleic Acids Res. 2013, 41, 9870–9880. [Google Scholar] [CrossRef] [PubMed]
- Heidrich, N.; Brantl, S. Antisense-RNA mediated transcriptional attenuation: Importance of a U-turn loop structure in the target RNA of plasmid pIP501 for efficient inhibition by the antisense RNA. J. Mol. Biol. 2003, 333, 917–929. [Google Scholar] [CrossRef]
- Jahn, N.; Brantl, S. Heat-shock-induced refolding entails rapid degradation of bsrG toxin mRNA by RNases Y and J1. Microbiology 2016, 162, 590–599. [Google Scholar] [CrossRef] [PubMed]
- Müller, P.; Jahn, N.; Ring, C.; Maiwald, C.; Neubert, R.; Meißner, C.; Brantl, S. A multistress responsive type I toxin-antitoxin system: bsrE/SR5 from the B. subtilis chromosome. RNA Biol. 2016, 13, 511–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meißner, C.; Jahn, N.; Brantl, S. In vitro characterization of the type I toxin-antitoxin system bsrE/SR5 from Bacillus subtilis. J. Biol. Chem. 2016, 291, 560–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reif, C.; Löser, C.; Brantl, S. Bacillus subtilis type I antitoxin SR6 promotes degradation of toxin yonT mRNA and is required to prevent toxic yoyJ overexpression. Toxins 2018, 10, 74. [Google Scholar] [CrossRef] [Green Version]
- Saito, S.; Kakeshita, H.; Nakamura, K. Novel small RNA-encoding genes in the intergenic regions of Bacillus subtilis. Gene 2009, 428, 2–8. [Google Scholar] [CrossRef]
- Eiamphungporn, W.; Helmann, J.D. Extracytoplasmic function sigma factors regulate expression of the Bacillus subtilis yabE gene via a cis-acting antisense RNA. J. Bacteriol. 2009, 191, 1101–1105. [Google Scholar] [CrossRef] [Green Version]
- Rao, F.; See, R.Y.; Zhang, D.; Toh, D.C.; Ji, Q.; Liang, Z.-X. YybT is a signaling protein that contains a cyclic dinucleotide phosphodiesterase domain and a GGDEF domain with ATPase activity. J. Biol. Chem. 2010, 285, 473–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Y.; Helmann, J.D. A σD-dependent antisense-transcript modulates expression of the cyclic-di-AMP hydrolase GdpP in Bacillus subtilis. Microbiology 2012, 158, 2732–2741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noone, D.; Salzberg, L.I.; Botella, E.; Basell, K.; Becher, D.; Antelmann, H.; Devine, K.M. A highly unstable transcript makes CwlO D,L-endopeptidase expression responsive to growth conditions in Bacillus subtilis. J. Bacteriol. 2014, 196, 237–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rath, H.; Reder, A.; Hoffmann, T.; Hammer, E.; Seubert, A.; Bremer, E.; Völker, U.; Mäder, U. Management of osmoprotectant uptake hierarchy in Bacillus subtilis via a SigB-dependent antisense RNA. Front. Microbiol. 2020, 11, 622. [Google Scholar] [CrossRef] [PubMed]
- Ul Haq, I.; Müller, P.; Brantl, S. SR7—A dual-function antisense RNA from Bacillus subtilis. RNA Biol. 2021, 18, 104–117. [Google Scholar] [CrossRef] [PubMed]
- Ul Haq, I.; Brantl, S. Moonlighting in Bacillus subtilis: The small proteins SR1P and SR7P regulate the moonlighting activity of glyceraldehyde 3-phosphate dehydrogenase A (GapA) and enolase in RNA degradation. Microorganisms 2021, 9, 1046. [Google Scholar] [CrossRef] [PubMed]
- Mars, R.A.; Mendonça, K.; Denham, E.L.; van Dijl, J.M. The reduction in small ribosomal subunit abundance in ethanol-stressed cells of Bacillus subtilis is mediated by a SigB-dependent antisense RNA. Biochim. Biophys. Acta 2015, 1853, 2553–2559. [Google Scholar] [CrossRef] [Green Version]
- Licht, A.; Preis, S.; Brantl, S. Implication of CcpN in the regulation of a novel untranslated RNA (SR1) in Bacillus subtilis. Mol. Microbiol. 2005, 58, 189–206. [Google Scholar] [CrossRef]
- Heidrich, N.; Chinali, A.; Gerth, U.; Brantl, S. The small untranslated RNA SR1 from the Bacillus subtilis genome is involved in the regulation of arginine catabolism. Mol. Microbiol. 2006, 62, 520–536. [Google Scholar] [CrossRef]
- Gaballa, A.; Antelmann, H.; Aguilar, C.; Khakh, S.K.; Song, K.B.; Smaldone, G.T.; Helmann, J.D. The Bacillus subtilis iron sparing response is mediated by a Fur regulated small RNA and three small, basic proteins. Proc. Natl. Acad. Sci. USA 2008, 105, 11927–11932. [Google Scholar] [CrossRef] [Green Version]
- Durand, S.; Braun, F.; Lioliou, E.; Romilly, C.; Helfer, A.C.; Kuhn, L.; Quittot, N.; Nicolas, P.; Romby, P.; Condon, C. A nitric oxide regulated small RNA controls expression of genes involved in redox homeostasis in Bacillus subtilis. PLoS Genet. 2015, 11, e1004957. [Google Scholar] [CrossRef] [Green Version]
- Durand, S.; Braun, F.; Helfer, A.C.; Romby, P.; Condon, C. sRNA-mediated activation of gene expression by inhibition of 5′-3′ exonucleolytic mRNA degradation. Elife 2017, 6, e23602. [Google Scholar] [CrossRef] [PubMed]
- Durand, S.; Callan-Sidat, A.; McKeown, J.; Li, S.; Kostova, G.; Hernandez-Fernaud, J.R.; Alam, M.T.; Millard, A.; Allouche, D.; Constantinidou, C.; et al. Identification of an RNA sponge that controls the RoxS riboregulatory of central metabolism in Bacillus subtilis. Nucleic Acids Res. 2021, 49, 6399–6419. [Google Scholar] [CrossRef]
- Mars, R.; Nicolas, P.; Ciccolini, M.; Rielman, E.; Reder, A.; Schaffer, M.; Mäder, U.; Völker, U.; van Dijl, J.M.; Denham, E.L. Small regulatory RNA-induced growth rate heterogeneity of Bacillus subtilis. PLoS Genet. 2015, 11, e1005046. [Google Scholar] [CrossRef] [Green Version]
- Heidrich, N.; Moll, I.; Brantl, S. In vitro analysis of the interaction between the small RNA SR1 and its primary target ahrC mRNA. Nucleic Acids Res. 2007, 35, 4331–4346. [Google Scholar] [CrossRef]
- Müller, P.; Gimpel, M.; Wildenhain, T.; Brantl, S. A new role for CsrA: Promotion of complex formation between an sRNA and its mRNA target in Bacillus subtilis. RNA Biol. 2019, 16, 972–987. [Google Scholar] [CrossRef] [Green Version]
- Licht, A.; Brantl, S. Transcriptional repressor CcpN from Bacillus subtilis compensates asymmetric contact distribution by cooperative binding. J. Mol. Biol. 2006, 364, 434–448. [Google Scholar] [CrossRef] [PubMed]
- Licht, A.; Golbik, R.; Brantl, S. Identification of ligands affecting the activity of the transcriptional repressor CcpN from Bacillus subtilis. J. Mol. Biol. 2008, 380, 17–30. [Google Scholar] [CrossRef]
- Licht, A.; Brantl, S. The transcriptional repressor CcpN from Bacillus subtilis uses different repression mechanisms at different promoters. J. Biol. Chem. 2009, 284, 30032–30038. [Google Scholar] [CrossRef] [Green Version]
- Czaplewski, L.G.; North, A.I.; Smith, M.C.; Baumberg, S.; Stockley, P.G. Purification and initial characterization of AhrC: The regulator of arginine metabolism genes in Bacillus subtilis. Mol. Microbiol. 1992, 6, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Gimpel, M.; Heidrich, N.; Mäder, U.; Krügel, H.; Brantl, S. A dual-function sRNA from B. subtilis: SR1 acts as a peptide encoding mRNA on the gapA operon. Mol. Microbiol. 2010, 76, 880–1009. [Google Scholar] [CrossRef] [Green Version]
- Gimpel, M.; Brantl, S. Dual-function sRNA encoded peptide SR1P modulates moonlighting activity of B. subtilis GapA. RNA Biol. 2016, 13, 916–926. [Google Scholar] [CrossRef] [Green Version]
- Gimpel, M.; Maiwald, C.; Wiedemann, C.; Görlach, M.; Brantl, S. Characterization of the interaction between the small RNA-encoded peptide SR1P and GapA from Bacillus subtilis. Microbiology 2017, 163, 1248–1259. [Google Scholar] [CrossRef]
- Gimpel, M.; Brantl, S. Dual-function small regulatory RNAs in bacteria. Mol. Microbiol. 2017, 103, 387–397. [Google Scholar] [CrossRef] [Green Version]
- Gimpel, M.; Preis, H.; Barth, E.; Gramzow, L.; Brantl, S. SR1—A small RNA with two remarkably conserved functions. Nucleic Acids Res. 2012, 40, 11659–11672. [Google Scholar] [CrossRef] [PubMed]
- Ul Haq, I.; Brantl, S.; Müller, P. A new role for SR1 from Bacillus subtilis - regulation of sporulation by inhibition of kinA translation. Nucleic Acids Res. 2021, in press. [Google Scholar] [CrossRef]
- Smaldone, G.T.; Revelles, O.; Gaballa, A.; Sauer, U.; Antelmann, H.; Helmann, J.D. A global investigation of the Bacillus subtilis iron-sparing response identifies major changes in metabolism. J. Bacteriol. 2012, 194, 2594–2605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smaldone, G.T.; Antelmann, H.; Gaballa, A.; Helmann, J.D. The FsrA sRNA and FbpB protein mediate the iron-dependent induction of the Bacillus subtilis lutABC iron-sulfur-containing oxidases. J. Bacteriol. 2012, 194, 2586–2593. [Google Scholar] [CrossRef]
- Figueroa-Bossi, N.; Valentini, M.; Malleret, L.; Fiorini, F.; Bossi, L. Caught at its own game: Regulatory small RNA inactivated by an inducible transcript mimicking its target. Genes Dev. 2009, 23, 2004–2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyakoshi, M.; Chao, Y.; Vogel, J. Cross talk between ABC transporter mRNAs via a target mRNA-derived sponge of the GcvB small RNA. EMBO J. 2015, 34, 1478–1492. [Google Scholar] [CrossRef]
- Bronesky, D.; Desgranges, E.; Corvaglia, A.; François, P.; Caballero, C.J.; Prado, L.; Toledo-Arana, A.; Lasa, I.; Moreau, K.; Vandenesch, F.; et al. A multifaceted small RNA modulates gene expression upon glucose limitation in Staphylococcus aureus. EMBO J. 2019, 38, e99363. [Google Scholar] [CrossRef] [PubMed]
- Li, G.W.; Oh, E.; Weissmann, J.S. The anti-Shine Dalgarno sequence drives translational pausing and codon choice in bacteria. Nature 2012, 484, 538–541. [Google Scholar] [CrossRef]
- Silvaggi, J.M.; Perkins, J.B.; Losick, R. Genes for small, noncoding RNAs under sporulation control in Bacillus subtilis. J. Bacteriol. 2006, 188, 532–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smirnov, A.; Schneider, C.; Hör, J.; Vogel, J. Discovery of new RNA classes and global RNA-binding proteins. Curr. Opin. Microbiol. 2017, 39, 152–160. [Google Scholar] [CrossRef]
Figure 1.
Overview of cis-encoded sRNAs in Bacillus subtilis and their mechanisms of action. (A) SR4 is a bifunctional antitoxin: It promotes the degradation of bsrG mRNA and inhibits bsrG translation. The SR5/bsrE mRNA interaction is highly similar, but SR5 exclusively facilitates degradation of bsrE mRNA; (B) SR6 represses two toxin genes by two distinct mechanisms: it inhibits the yoyJ translation and induces the degradation of yonT mRNA; (C) RatA promotes txpA mRNA degradation; (D) S1290 delays the opuB expression after salt shock by transcriptional interference; (E) SR7 reduces the rpsD expression by transcriptional interference under several stress conditions; (F) S25 reduces the amount of yabE mRNA; (G) S1326 represses the cwlO expression; (H) S1559 represses the gdpP expression. The two genomic strands are illustrated as black lines, genes as boxes, and promoter regions as black boxes. The sRNAs and their genes are depicted in red and target genes and mRNAs in blue, other functionally important genes in purple, and unrelated genes in gray; RNaseIII cleavage is represented by a green lightning. In the case of translational inhibition, the RBS is depicted as a grey box.
Figure 1.
Overview of cis-encoded sRNAs in Bacillus subtilis and their mechanisms of action. (A) SR4 is a bifunctional antitoxin: It promotes the degradation of bsrG mRNA and inhibits bsrG translation. The SR5/bsrE mRNA interaction is highly similar, but SR5 exclusively facilitates degradation of bsrE mRNA; (B) SR6 represses two toxin genes by two distinct mechanisms: it inhibits the yoyJ translation and induces the degradation of yonT mRNA; (C) RatA promotes txpA mRNA degradation; (D) S1290 delays the opuB expression after salt shock by transcriptional interference; (E) SR7 reduces the rpsD expression by transcriptional interference under several stress conditions; (F) S25 reduces the amount of yabE mRNA; (G) S1326 represses the cwlO expression; (H) S1559 represses the gdpP expression. The two genomic strands are illustrated as black lines, genes as boxes, and promoter regions as black boxes. The sRNAs and their genes are depicted in red and target genes and mRNAs in blue, other functionally important genes in purple, and unrelated genes in gray; RNaseIII cleavage is represented by a green lightning. In the case of translational inhibition, the RBS is depicted as a grey box.

Figure 2.
Interactions of the currently known trans-encoded sRNAs with their target mRNAs. (A) Left half: SR1 interacts with ahrC mRNA about 100 nt downstream of the RBS, which induces structural changes around the ahrC RBS that inhibit translation initiation. Hfq (dark-green) binds immediately upstream of the ahrC RBS (grey rectangle) to make it accessible to the 30S SU. By binding to GGA motives 1–3 of ahrC mRNA, CsrA (light-green) induces a slight structural change that makes region G’ (turquois) accessible to complementary region G of SR1. SR1 binding causes a structural change that renders the ahrC RBS inaccessible to 30S binding, thereby inhibiting translation initiation [45,46] (based on [4]). Right half: SR1 inhibits translation initiation of kinA mRNA by occluding the kinA RBS. The initial SR1–kinA mRNA interaction occurs between complementary regions D/D’ (violet) followed by the interaction between complementary regions E/E’ (grey). (B) Interaction of FsrA with gltAB mRNA. FsrA has numerous targets, but in vivo base-pairing has only been shown for gltAB mRNA. Most probably, FsrA inhibits translation of all targets (see Table 2). The two C-rich regions (CRR) are highlighted in pink. (C) Interaction of RoxS with two target mRNAs. Whereas RoxS and its truncated derivative repress ppnKB translation, full-length RoxS activates yflS translation, both by disrupting a structure occluding the RBS and by inhibiting RNase J1 degradation from the 5′ end. RoxS interacts with its targets via CRR’s (shown in pink), CRR3 was found to be decisive. (D) Interaction of RnaC with abrB mRNA, which promotes RNA degradation and, most probably, inhibits abrB translation. Red, sRNAs; blue, target mRNAs; beige oval, 30S SU; grey box, RBS; green arrow, RNase III; black arrow, RNase Y; yellow, RNase J1.
Figure 2.
Interactions of the currently known trans-encoded sRNAs with their target mRNAs. (A) Left half: SR1 interacts with ahrC mRNA about 100 nt downstream of the RBS, which induces structural changes around the ahrC RBS that inhibit translation initiation. Hfq (dark-green) binds immediately upstream of the ahrC RBS (grey rectangle) to make it accessible to the 30S SU. By binding to GGA motives 1–3 of ahrC mRNA, CsrA (light-green) induces a slight structural change that makes region G’ (turquois) accessible to complementary region G of SR1. SR1 binding causes a structural change that renders the ahrC RBS inaccessible to 30S binding, thereby inhibiting translation initiation [45,46] (based on [4]). Right half: SR1 inhibits translation initiation of kinA mRNA by occluding the kinA RBS. The initial SR1–kinA mRNA interaction occurs between complementary regions D/D’ (violet) followed by the interaction between complementary regions E/E’ (grey). (B) Interaction of FsrA with gltAB mRNA. FsrA has numerous targets, but in vivo base-pairing has only been shown for gltAB mRNA. Most probably, FsrA inhibits translation of all targets (see Table 2). The two C-rich regions (CRR) are highlighted in pink. (C) Interaction of RoxS with two target mRNAs. Whereas RoxS and its truncated derivative repress ppnKB translation, full-length RoxS activates yflS translation, both by disrupting a structure occluding the RBS and by inhibiting RNase J1 degradation from the 5′ end. RoxS interacts with its targets via CRR’s (shown in pink), CRR3 was found to be decisive. (D) Interaction of RnaC with abrB mRNA, which promotes RNA degradation and, most probably, inhibits abrB translation. Red, sRNAs; blue, target mRNAs; beige oval, 30S SU; grey box, RBS; green arrow, RNase III; black arrow, RNase Y; yellow, RNase J1.
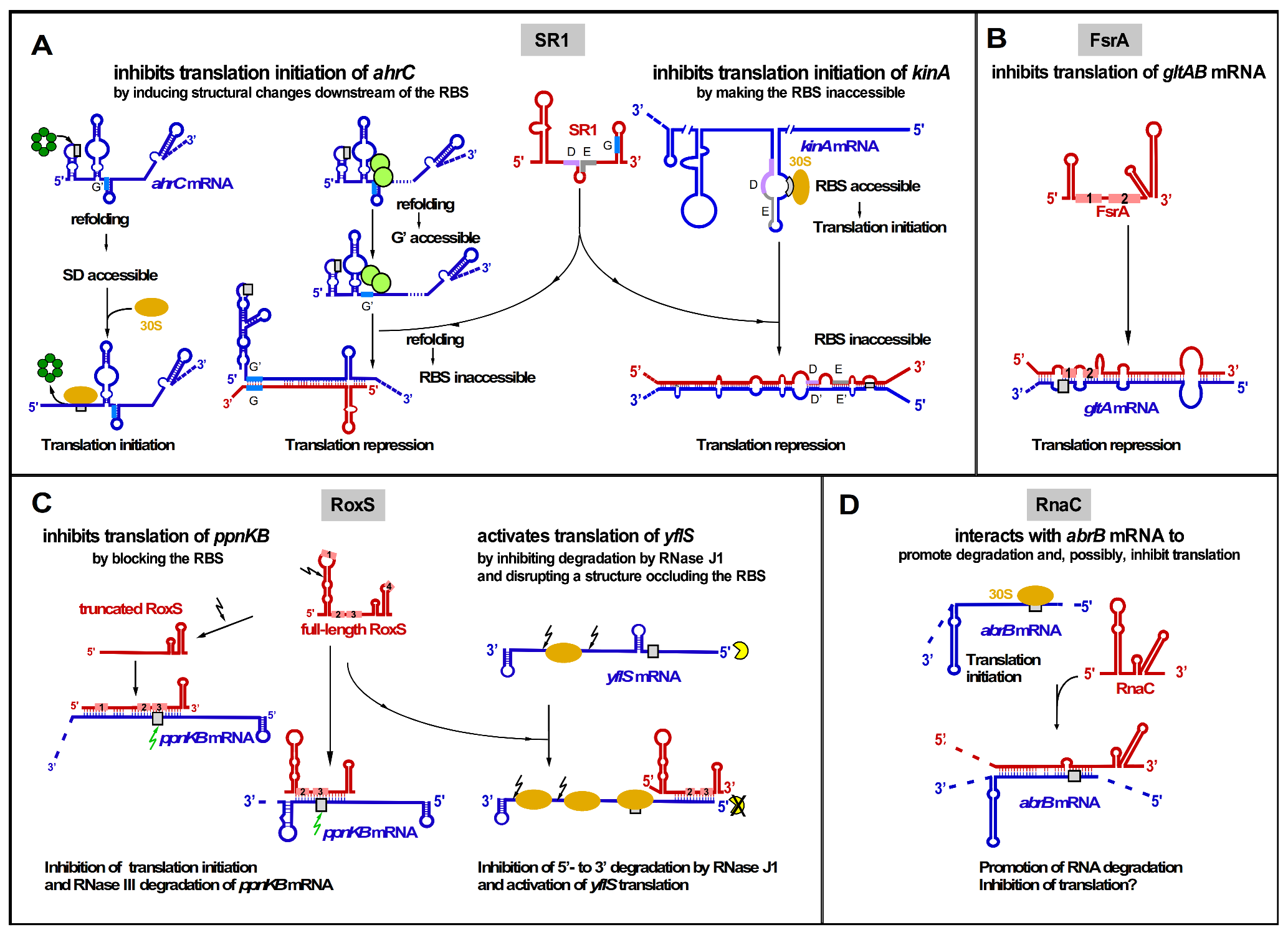
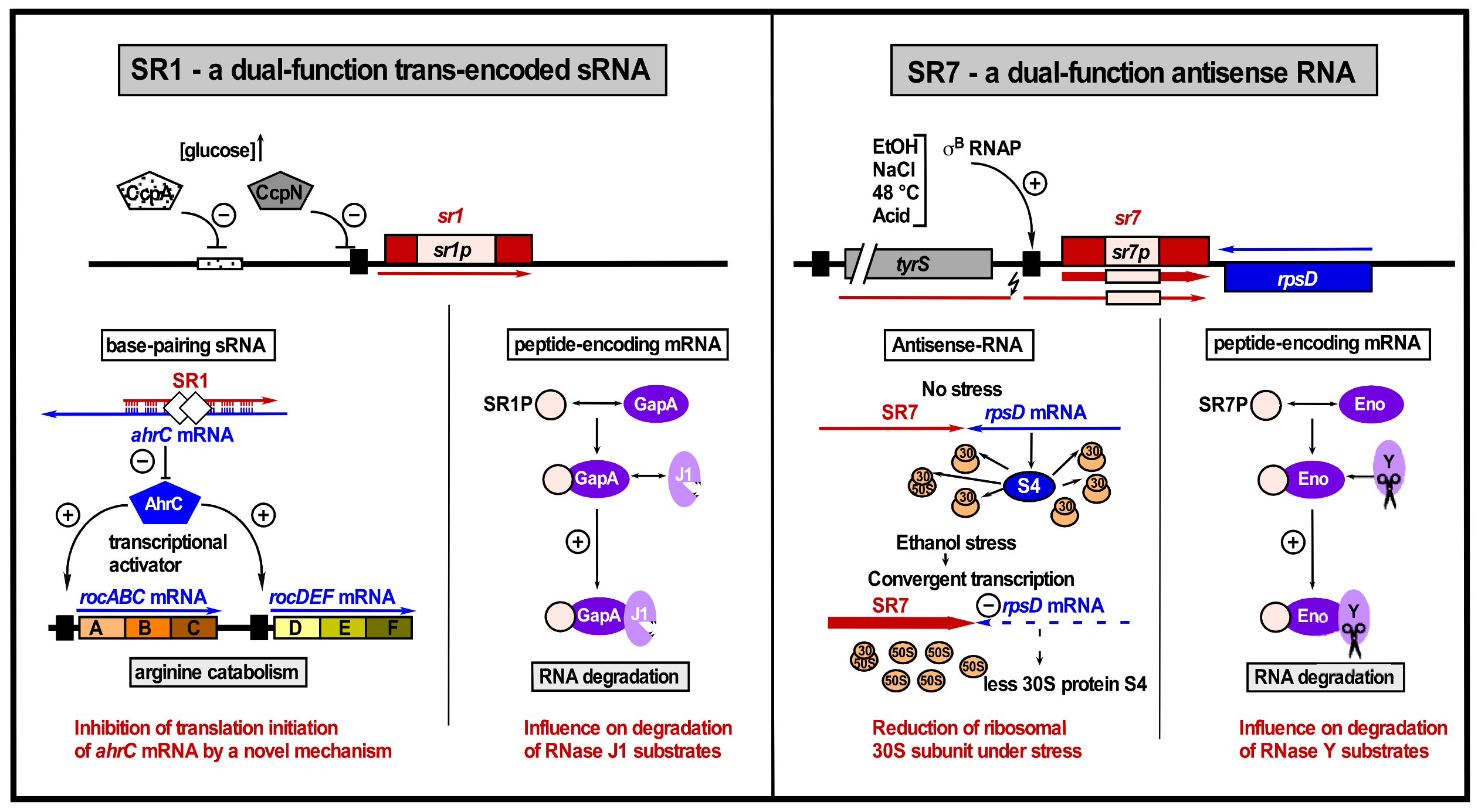
Figure 3.
Two dual-function regulatory RNAs from B. subtilis, SR1 and SR7. Left: SR1 acts on the one hand as base-pairing sRNA in arginine catabolism by inhibiting translation of ahrC mRNA encoding the transcriptional activator of the arginine catabolic operons and on the other hand as mRNA encoding SR1P that modulates the B. subtilis DLN (degradosome-like network) by interacting with GapA to promote the GapA-RNase J1 interaction and enhance the RNase J1 activity. Transcription of sr1 is repressed by CcpA and CcpN. The RNA chaperone CsrA (white diamonds) promotes the SR1-ahrC mRNA interaction (adapted with permission from [4]). Right: On the one hand, SR7 reduces by transcriptional interference the amount of the convergently transcribed rpsD mRNA and, on the other hand, encodes SR7P that also modulates the DLN by interacting with enolase, increasing the recruitment of RNase Y and enhancing the RNase Y activity. Black line, DNA; red and blue lines, antisense and target RNAs; pink rectangle, SR1P or SR7P ORF.
Figure 3.
Two dual-function regulatory RNAs from B. subtilis, SR1 and SR7. Left: SR1 acts on the one hand as base-pairing sRNA in arginine catabolism by inhibiting translation of ahrC mRNA encoding the transcriptional activator of the arginine catabolic operons and on the other hand as mRNA encoding SR1P that modulates the B. subtilis DLN (degradosome-like network) by interacting with GapA to promote the GapA-RNase J1 interaction and enhance the RNase J1 activity. Transcription of sr1 is repressed by CcpA and CcpN. The RNA chaperone CsrA (white diamonds) promotes the SR1-ahrC mRNA interaction (adapted with permission from [4]). Right: On the one hand, SR7 reduces by transcriptional interference the amount of the convergently transcribed rpsD mRNA and, on the other hand, encodes SR7P that also modulates the DLN by interacting with enolase, increasing the recruitment of RNase Y and enhancing the RNase Y activity. Black line, DNA; red and blue lines, antisense and target RNAs; pink rectangle, SR1P or SR7P ORF.

{kind=link}
{kind=link}
{kind=link}
Table 1.
Overview and characteristics of all known cis-encoded sRNAs and their target RNAs in Bacillus subtilis.
Table 1.
Overview and characteristics of all known cis-encoded sRNAs and their target RNAs in Bacillus subtilis.
| sRNA | sRNA Length | Mechanism of Action of sRNA | Target RNA | Target Gene Function | Regulation, Peculiarity |
|---|---|---|---|---|---|
| RatA | 222 nt | RD | txpA | Toxin | Glucose dependent |
| SR4 | 180 nt | RD + TI | bsrG | Toxin | Temperature dependent |
| SR5 | 163 nt | RD | bsrE | Toxin | Multistress responsive |
| SR6 | 100/215 nt * | RD TI | yonT yoyJ | Toxin Toxin | Multistress responsive |
| as-bsrH | 200 nt | RD | bsrH | Toxin | Multistress responsive |
| S25 | ≈1350 nt | unknown | yabE | Autolysin | sRNA under control of σM and σX |
| S1559 | 667 nt | unknown | gdpP | c-di-AMP PD | sRNA under σD control |
| S1326 | 700–2200 nt # | unknown | cwlO | Autolysin | sRNA under σB control |
| S1290 | 300–3800 nt # | T interference | opuB | Choline transporter | sRNA under σB control |
| SR7 | 185/259 nt | T interference | rpsD | Ribosomal protein S4 | sRNA under σB control; dual-function antisense RNA |
RD, promotion of RNA degradation; TI, translational inhibition; T interference, transcriptional interference; PD, phosphodiesterase; * longer SR6 species due to read-through of the SR6 terminator; # several longer RNA species due to read-through of the terminator.
Table 2.
Overview and characteristics of all known trans-encoded sRNAs and their target genes in Bacillus subtilis.
Table 2.
Overview and characteristics of all known trans-encoded sRNAs and their target genes in Bacillus subtilis.
| sRNA | sRNA Length | Mechanism of Action of sRNA | Target RNA | Target Gene Function | Regulation |
|---|---|---|---|---|---|
| SR1 | 205 nt | TI TI | ahrC kinA | Arginine catabolism Sporulation initiation | CcpN, CcpA, sporulation |
| FsrA | 84 nt | TI TI TI TI TI TI | sdhCAB citB gltA lutABC dctP leuCD | Iron sparing response Aconitase Glutamate synthase Iron-sulfur oxidase Dicarboxylate permease Leucine biosynthesis | Fur, iron; Some targets need FbpA, B, or C |
| RnaC | 125 nt ? | RD + TI ? | abrB | Transition state regulation | Growth phase |
| RoxS | 115 nt | TI + RD TI + RD RD RS + TA | ppnKB sucC acsA yflS | Redox regulation, TCA cycle, Acetyl-CoA synthetase Malate transporter | ResD (NO), Rex (malate) |
| RosA | 92/128/ 193/225 nt * | Seq | RoxS FsrA | Trans-encoded sRNA Trans-encoded sRNA | CcpA |
TI, translational inhibition; TA, translational activation; RD, promotion of RNA degradation; RS, RNA stabilization; Seq, sequestration of other RNAs; * several distinct RNA species were detected in Northern blotting; ? length unknown.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Brantl, S.; Müller, P. Cis- and Trans-Encoded Small Regulatory RNAs in Bacillus subtilis. Microorganisms 2021, 9, 1865. https://doi.org/10.3390/microorganisms9091865
AMA Style
Brantl S, Müller P. Cis- and Trans-Encoded Small Regulatory RNAs in Bacillus subtilis. Microorganisms. 2021; 9(9):1865. https://doi.org/10.3390/microorganisms9091865
Chicago/Turabian StyleBrantl, Sabine, and Peter Müller. 2021. "Cis- and Trans-Encoded Small Regulatory RNAs in Bacillus subtilis" Microorganisms 9, no. 9: 1865. https://doi.org/10.3390/microorganisms9091865
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.