Fallow Deer (Dama dama) as a Reservoir of Shiga Toxin-Producing Escherichia coli (STEC)


Department of Epizootiology, Faculty of Veterinary Medicine, University of Warmia and Mazury in Olsztyn, Oczapowskiego 13, 10-718 Olsztyn, Poland
*
Author to whom correspondence should be addressed.
Animals 2020, 10(5), 881; https://doi.org/10.3390/ani10050881
Submission received: 21 April 2020
/
Revised: 18 May 2020
/
Accepted: 18 May 2020
/
Published: 19 May 2020
(This article belongs to the Special Issue Wild Animals' Infectious Disease: The Wild–Domestic–Human Interface)
Abstract
:Simple Summary
Infections caused by Shiga toxin-producing Escherichia (E.) coli (STEC) are the third most typically reported zoonosis within the European Union after campylobacteriosis and salmonellosis. STEC pathogens are responsible for the outbreaks of serious diseases in humans, including haemolytic uraemic syndrome (HUS), haemorrhagic colitis (HC) and diarrhoea (D). Red deer, roe deer and wild boars are important environmental reservoirs of foodborne pathogens that may cause serious diseases in humans and contaminate fresh food products. The occurrence of STEC and attaching and effacing (AE)-STEC in the Polish population of wild fallow deer was analysed in this study. The presence of potentially pathogenic STEC/AE-STEC in fallow deer poses a risk of carcass contamination, which could have serious consequences because venison can also be consumed raw as carpaccio or steak tartare. Only a few reports on wild animals as a reservoir of foodborne pathogens in European countries have been published to date, and the present study attempts to fill in this knowledge gap by assessing the possible epidemiological risk related to STEC/AE-STEC in fallow deer. Three isolates had the virulence profile that is associated with HUS/D/HC according to the FAO/WHO report. The results of this study suggest that fallow deer are carriers of STEC/AE-STEC that are potentially pathogenic to humans.
Abstract
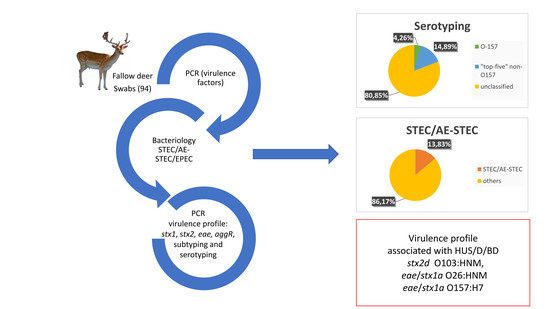
Shiga toxin-producing Escherichia (E.) coli (STEC) are responsible for the outbreaks of serious diseases in humans. Only a few reports on fallow deer as a reservoir of foodborne pathogens have been published to date. The purpose of this study was to determine the occurrence of STEC strains in the fallow deer population in Poland. In all, 94 fallow deer swabs were tested. Polymerase chain reaction (PCR) was performed to detect the virulence profile of stx1, stx2 and eae or aggR genes, to identify the subtypes of stx1 and stx2 genes and to perform O and H serotyping. STEC and attaching and effacing (AE)-STEC were identified in 13 isolates (13.83%). The most hazardous virulence profile was detected in three strains, namely stx2d serotype O103:HNM, eae/stx1a serotype O26:HNM and eae/stx1a serotype O157:H7. The predominant stx gene was stx2, which was identified in 76.92% of isolates. E. coli O157 was detected in 4/94 (4.26%). Other E. coli serogroups, O26, O103, O111 and O145, were identified in 14/94 fallow deer (14.89%). The present findings suggest that fallow deer are carriers of STEC/AE-STEC that are potentially pathogenic to humans.
1. Introduction
Shiga toxin-producing Escherichia (E.) coli (STEC) are generally recognised as a significant cause of foodborne diseases, such as haemolytic uraemic syndrome (HUS), haemorrhagic colitis (HC) and diarrhoea (D) [1]. In 2018, 8161 confirmed cases of STEC infections were registered in the European Union (EU) [2], and the number of cases and the notification rate of STEC increased significantly from 2017 (n = 6073), which made STEC the third most frequently reported zoonosis in the EU after campylobacteriosis (n = 246,571) and salmonellosis (n = 91,857) [2,3]. Between 2009 and 2018, the prevalence of human STEC infections increased steadily, principally due to a large STEC outbreak in 2011 [2,4]. The observed increase can be partially attributed to higher detection rates following the STEC outbreak, as well as advanced laboratory methods, including polymerase chain reaction (PCR), for direct extraction of bacterial DNA from specimens and strain characterisation [2]. According to the Food and Agriculture Organization (FAO) of the United Nations and the World Health Organization (WHO), the pathogenicity of STEC to humans depends on the occurrence of selected virulence factors, in particular Shiga toxin 1 (stx1), Shiga toxin 2 (stx2), attaching and effacing E. coli (eae), or a transcriptional activator of aggregative adherence fimbria I (aggR) [1]. Shiga toxin genes are subdivided into three stx1 subtypes, namely a, c and d, and seven stx2 subtypes, namely a, b, c, d, e, f and g [5]. The latest studies have demonstrated that STEC strains classified as subtype stx2a and possessing adherence genes eae or aggR pose the most serious health risk to humans and have the highest potential to induce HUS [1,6,7]. According to the strategy for assessing health risks based on an analysis of STEC virulence genes, strains containing stx2a or stx2d and eae or aggR genes are most harmful to humans. Such strains have the highest capacity to cause D, HC and HUS. In D and HC patients, strains with the stx2c/eae or stx1a/eae virulence profiles have been identified [1]. Most STEC infections are caused by the ingestion of faecal-contaminated food or water, indirect or direct exposure to animal vectors, or secondary transmission from humans to humans [8]. Household ruminants, especially cattle, are recognised as the major natural STEC reservoir [8]. Large wildlife, such as deer (Cervus elaphus), roe deer (Capreolus capreolus) and wild boars (Sus scrofa), can also be healthy hosts of O157:H7 and non-O157 STEC, but the number of publications on the virulence profiles of strains isolated from these animals is not significant [9,10,11,12,13,14,15,16]. According to the European Food Safety Authority (EFSA) and the European Centre for Disease Prevention and Control (ECDC) [2], both the number of reports and the number of tested animal samples have decreased steadily in recent years. Animal samples are still frequently assayed with the use of techniques that reveal only E. coli O157, which puts into question the reliability of the reports on STEC detection in animals in the EU. In our previous study, 21 rectal swabs from fallow deer were analysed, and the occurrence rate of STEC/AE-STEC strains in the fallow deer population in Poland was determined at 9.52%. The prevalence of enteropathogenic E. coli (EPEC) reached 33.33%, and the prevalence of the “top five” serogroups was determined at 9.52%. One strain with the stx1a/stx2g virulence profile and one with stx2b were identified. However, the results of that study could not be used to formulate reliable conclusions due to the small number of rectal swabs (21 samples) [15]. In the present study, additional swabs collected during the 2017–2018 and 2018–2019 hunting seasons were analysed to identify virulence profile, stx1 and stx2 subtypes and the prevalence of O-serogroups in STEC obtained from the Polish population of fallow deer (Dama dama).
2. Materials and Methods
2.1. Sampling
During two autumn–winter hunting seasons of 2017–2018 and 2018–2019 (from 1 September to the end of February for males and fawns, and from 1 September to 15 January for females) [17,18] in Poland, a total of 94 rectal swabs were obtained from fallow deer (Dama dama). The swabs were collected in collaboration with hunters. Unfortunately, nothing is known about the health status of the animals from which the swabs were taken. The swabs were collected from each animal before evisceration, placed in tubes and transported refrigerated to the laboratory within 48 h. All swabs were collected as part of a standard procedure; therefore, ethics approval was not required.
2.2. Detection of STEC Strains and stx1 and stx2 Subtypes
Swabs were mixed in 5 mL of buffered peptone water (BPW; BTL) in aseptic conditions and incubated for 20–24 h at 37 °C. DNA was extracted from 1 mL of culture using the Genomic Mini kit (A&A Biotechnology, Gdynia, Poland) following the manufacturer’s recommendations. The samples were screened for the presence of stx1, stx2, eae and aggR genes following the protocol proposed by the European Union Reference Laboratory for E. coli (EU-RL VTEC_Method 01 for E. coli) and the method described by Schmidt et al. [19,20,21,22]. In order to isolate a single STEC/AE-STEC strain, all samples that tested positive for the analysed genes in PCR were treated as previously described by Szczerba-Turek et al. [15]. The stx1 and stx2 subtypes were determined following the protocol developed by Scheutz et al. [5] and the protocol proposed by the European Union Reference Laboratory for E. coli (EU-RL VTEC_Method 006) [23].
2.3. Serotyping by Polymerase Chain Reaction (PCR)
The STEC/AE-STEC/EPEC serotypes were determined by the PCR assay based on the EU-RL protocol for E. coli (EU-RL VTEC_Method 003) [24,25], for O-antigen-encoding genes (wzx)—O26, O103, O111, O145 and O157—and the protocols proposed by Durso et al., Gannon et al. and Mora et al. [26,27,28] for H antigens encoding the fliC gene (specific to flagellar genes)—H7, H8, H11, H21 and H28. All PCR amplifications were performed with the HotStartTaq Plus DNA Polymerase Kit (Qiagen, Venlo, Netherlands) and the HotStartTaq Plus Master Mix Kit (Qiagen), according to the manufacturer’s recommendations. The annealing temperature for every primer was described previously [15]. The PCR products were separated by electrophoresis in 2% agarose gel with the Midori Green Advanced DNA Stain (Nippon Genetics Europe GmbH, Düren, Germany).
2.4. Statistics
For basic statistical analysis, the binomial (Clopper–Pearson) “exact” method based on beta distribution at a significance level of α = 0.05 and 95% confidence interval was applied. All statistical analyses were performed with free EpiTools epidemiological calculators (http://epitools.ausvet.com.au) [29].
3. Results
In the studied population of 94 fallow deer (Dama dama), STEC were identified in four isolates (4.26%, 95% CI = 1.17–10.54), AE-STEC were detected in nine isolates (9.57%, 95% CI = 5.99–19.97), and EPEC were identified in eight isolates (8.51%, 95% CI = 3.75–16.08). STEC/AE-STEC were identified in 13 isolates (13.83%, 95% CI = 7.57–22.49). In the examined group of STEC/AE-STEC, stx2 was the predominant gene and was identified in 10 isolates (76.92%, 95% CI = 46.19–94.96), whereas the stx1 gene was detected in seven isolates (53.85%, 95% CI = 25.13–80.78). Three strains harboured the stx1a gene, one strain harboured the stx2a gene, one strain harboured the stx2d gene and four strains harboured the stx2b gene. The occurrence of stx1 and stx2 subtypes is shown in Table 1. E. coli O157 strains were detected in 4/94 rectal swabs from fallow deer (Dama dama) (4.26%, 95% CI = 1.17–10.54) and three of them were identified as O157:H7 (3.19%, 95% CI = 0.66–9.04). E. coli O26, O103, O111 and O145 were identified in 14/94 fallow deer (14.89%, 95% CI = 8.391–23.72). The “top five” were identified in 18/94 samples (19.15%, 95% CI = 11.76–28.56). The results of serotyping are presented in Table 1.
4. Discussion
According to the concept of “One Health” developed in the early 2000s, human health and animal health (livestock, pets and wildlife) are interdependent and bound to the health of the ecosystems in which they exist, which is why their overall health status should be closely monitored [30,31,32]. Human infections with STEC have been reported after the consumption of deer meat or the meat of other game animals [33,34,35,36,37,38,39]. This study analysed the occurrence of STEC/AE-STEC in fallow deer that can be an important natural reservoir of STEC/AE-STEC strains. The above has significant implications for public health because the number of fallow deer farms has increased due to a decline in the profitability of livestock farming. The global population of farmed deer is estimated at nearly 5 million [40]. In Europe, the highest number of fallow deer breeding farms are located in Germany and the United Kingdom [41], but other European countries such as Sweden, Czechia, Austria, Spain, Italy, Switzerland, Norway, Slovakia, Hungary and Poland have well-established traditions relating to deer breeding, venison production and consumption as well as game hunting [42]. Fallow deer are reared in Europe on account of their long lifespan and their ability to withstand disease and low temperatures in winter and produce high-quality meat that is valued for its unique taste and high nutritional value [42,43,44,45]. Fallow deer are hunter-harvested for meat, skin and antlers. The fact that STEC/AE-STEC strains that are potentially pathogenic to humans were present in swabs collected from free-living fallow deer points to the possibility of environmental contamination, particularly in grasslands grazed by cattle, sheep and other farm animals, which may result in the cross-transmission of STEC/AE-STEC between different animal species. Such a situation was observed in the USA where potentially pathogenic STEC/AE-STEC strains derived from deer were found in cows and their cross-transmission was reported [46]. In addition, STEC/AE-STEC have also been isolated from water bodies and vegetable crops, posing a real risk to human health. Therefore, it is important to monitor free-living animals for the occurrence of zoonotic strains that are potentially pathogenic to humans. STEC should also be classified based on type (stx1 or stx2). The prevalence of STEC/AE-STEC that are potentially pathogenic to humans in the fallow deer population is an important consideration. Hunters have to comply with the regulations laying down specific hygiene rules for food of animal origin and specific rules for the organisation of official controls on products of animal origin intended for human consumption [47,48].
There are no previously published reports of the occurrence of STEC/AE-STEC, containing stx1 and stx2 subtypes, and the virulence genes of strains isolated from fallow deer. In our previous study, which analysed only 21 faecal swabs from the Polish population of fallow deer, the occurrence of STEC/AE-STEC was estimated at 9.52%, whereas E. coli O157 was not detected [15]. In the present investigation, the occurrence of STEC/AE-STEC was estimated at 13.8% (13/94), the occurrence of E. coli O157 strains was estimated at 4.2% (4/94) and three strains were identified as O157:H7 (3.2%) (3/94). One EPEC strain had the eae/stx1a virulence profile, whereas the eae/stx1NS profile was determined in two AE-STEC strains. Three isolates had the virulence profile that is associated with HUS/D/HC according to the FAO/WHO report [1], namely stx2d serotype O103:HNM, eae/stx1a serotype O26:HNM and eae/stx1a serotype O157:H7.
A total of 115 samples were analysed in the current study and our previous research [15]. The occurrence of STEC/AE-STEC was estimated at 13% (15/115), the occurrence of E. coli O157 strains was estimated at 3.48% (4/115) and three strains were identified as O157:H7 (2.6%) (3/115). The prevalence of EPEC strains was determined at 6.1% (7/115). The predominant stx subtype was stx2 that was identified in 10.43% of the samples (12/115), whereas subtype stx1 was found in 6.95% of the analysed swabs (8/115). The presence of potentially pathogenic STEC/AE-STEC in fallow deer poses a risk of carcass contamination, which could have serious consequences because venison can also be consumed raw as carpaccio or steak tartare. For this reason, the microbiological safety of meat from fallow deer has to be ensured. However, STEC that are potentially pathogenic to humans were found in the examined samples, which points to the risk of carcass and environmental contamination.
5. Conclusions
Fallow deer can carry STEC/AE-STEC that are potentially pathogenic to humans. Three strains isolated in this study had the virulence profile associated with human infections, namely stx2d serotype O103:HNM, eae/stx1a serotype O26:HNM and eae/stx1a serotype O157:H7. This is an important finding that indicates that high hygiene standards have to be observed in the process of dressing fallow deer carcasses in the field and preparing fallow deer meat. The occurrence of STEC/AE-STEC in the rectal swabs of wild fallow deer should also be taken into account when developing strategies aiming to limit and/or control this pathogen in water, livestock, pets and wildlife. This is the first complete report describing the virulence of STEC/AE-STEC isolated from fallow deer.
Author Contributions
Conceptualization, A.S.-T.; methodology, A.S.-T. and B.K.; formal analysis, A.S.-T.; investigation, A.S.-T.; writing—original draft preparation, A.S.-T. and B.K.; writing—review and editing, A.S.-T. and B.K.; visualization A.S.-T.; supervision A.S.-T. All authors have read and agreed to the published version of the manuscript.
Funding
This research received financial support from the Ministry of Science and Higher Education, Poland, under the “Regional Initiative of Excellence” programme for the years 2019–2022 (Project No. 010/RID/2018/19, amount of funding: PLN 12,000,000).
Acknowledgments
The authors would like to thank Piotr Socha for coordinating sample collection and all staff of the EU-RL for Escherichia coli, including verotoxigenic E. coli (VTEC), Istituto Superiore di Sanità, Roma, for their help.
Conflicts of Interest
The authors declare no conflict of interest.
References
- FAO/WHO. Shiga toxin-producing Escherichia coli (STEC) and food: Attribution, characterization, and monitoring. Microbiological Risk Assessment Series, No. 31, Food and Agriculture Organization of the United Nations and World Health Organization. 2018. Available online: https://apps.who.int/iris/handle/10665/272871 (accessed on 20 September 2019).
- EFSA and ECDC (European Food Safety Authority and European Centre for Disease Prevention and Control). The European Union One Health 2018 Zoonoses Report. EFSA J. 2019, 17. [Google Scholar] [CrossRef] [Green Version]
- EFSA and ECDC (European Food Safety Authority and European Centre for Disease Prevention and Control). The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2017. EFSA J. 2018, 16. [Google Scholar] [CrossRef]
- Bielaszewska, M.; Mellmann, A.; Zhang, W.; Koeck, R.; Fruth, A.; Bauwens, A.; Peters, G.; Karch, H. Characterisation of the Escherichia coli strain associated with an outbreak of haemolytic uraemic syndrome in Germany, 2011: A microbiological study. Lancet Infect. Dis. 2011, 11, 671–676. [Google Scholar] [CrossRef] [Green Version]
- Scheutz, F.; Teel, L.D.; Beutin, L.; Pierard, D.; Buvens, G.; Karch, H.; Mellmann, A.; Caprioli, A.; Tozzoli, R.; Morabito, S.; et al. Multicenter Evaluation of a Sequence-Based Protocol for Subtyping Shiga Toxins and Standardizing Stx Nomenclature. J. Clin. Microbiol. 2012, 50, 2951–2963. [Google Scholar] [CrossRef] [Green Version]
- Asakura, H.; Boisen, N.; Chinen, I.; Cook, R.; Dallman, T.; Devleesschauwer, B.; Feng, P.; Franz, E.; Fratamico, P.; Gill, A.; et al. Hazard Identification and Characterization: Criteria for Categorizing Shiga Toxin-Producing Escherichia coli on a Risk Basis. J. Food Prot. 2019, 82, 7–21. [Google Scholar]
- Haddad, N.; Johnson, N.; Kathariou, S.; Metris, A.; Phister, T.; Pielaat, A.; Tassou, C.; Wells-Bennikh, M.H.J.; Zwietering, M.H. Next generation microbiological risk assessment-Potential of omics data for hazard characterisation. Int. J. Food Microbiol. 2018, 287, 28–39. [Google Scholar] [CrossRef]
- Caprioli, A.; Morabito, S.; Brugere, H.; Oswald, E. Enterohaemorrhagic Escherichia coli: Emerging issues on virulence and modes of transmission. Vet. Res. 2005, 36, 289–311. [Google Scholar] [CrossRef] [Green Version]
- Dias, D.; Caetano, T.; Torres, R.T.; Fonseca, C.; Mendo, S. Shiga toxin-producing Escherichia coli in wild ungulates. Sci. Total Environ. 2019, 651, 203–209. [Google Scholar] [CrossRef]
- Diaz-Sanchez, S.; Sanchez, S.; Herrera-Leon, S.; Porrero, C.; Blanco, J.; Dahbi, G.; Blanco, J.E.; Mora, A.; Mateo, R.; Hanning, I.; et al. Prevalence of Shiga toxin-producing Escherichia coli, Salmonella spp. and Campylobacter spp. in large game animals intended for consumption: Relationship with management practices and livestock influence. Vet. Microbiol. 2013, 163, 274–281. [Google Scholar] [CrossRef]
- Frank, E.; Bonke, R.; Drees, N.; Heurich, M.; Martlbauer, E.; Gareis, M. Shiga toxin-producing Escherichia coli (STEC) shedding in a wild roe deer population. Vet. Microbiol. 2019, 239, 8. [Google Scholar] [CrossRef]
- Garcia-Sanchez, A.; Sanchez, S.; Rubio, R.; Pereira, G.; Alonso, J.M.; de Mendoza, J.H.; Rey, J. Presence of Shiga toxin-producing E-coli O157: H7 in a survey of wild artiodactyls. Vet. Microbiol. 2007, 121, 373–377. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, S.; Garcia-Sanchez, A.; Martinez, R.; Blanco, J.; Blanco, J.E.; Blanco, M.; Dahbi, G.; Mora, A.; de Mendoza, J.H.; Alonso, J.M.; et al. Detection and characterisation of Shiga toxin-producing Escherichia coli other than Escherichia coli O157:H7 in wild ruminants. Vet. J. 2009, 180, 384–388. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, S.; Martinez, R.; Garcia, A.; Vidal, D.; Blanco, J.; Blanco, M.; Blanco, J.E.; Mora, A.; Herrera-Leon, S.; Echeita, A.; et al. Detection and characterisation of O157:H7 and non-O157 Shiga toxin-producing Escherichia coli in wild boars. Vet. Microbiol. 2010, 143, 420–423. [Google Scholar] [CrossRef] [PubMed]
- Szczerba-Turek, A.; Siemionek, J.; Socha, P.; Bancerz-Kisiel, A.; Platt-Samoraj, A.; Lipczynska-Ilczuk, K.; Szweda, W. Shiga toxin-producing Escherichia coli isolates from red deer (Cervus elaphus), roe deer (Capreolus capreolus) and fallow deer (Dama dama) in Poland. Food Microbiol. 2020, 86, 7. [Google Scholar] [CrossRef]
- Szczerba-Turek, A.; Socha, P.; Bancerz-Kisiel, A.; Platt-Samoraj, A.; Lipczynska-Ilczuk, K.; Siemionek, J.; Konczyk, K.; Terech-Majewska, E.; Szweda, W. Pathogenic potential to humans of Shiga toxin-producing Escherichia coli isolated from wild boars in Poland. Int. J. Food Microbiol. 2019, 300, 8–13. [Google Scholar] [CrossRef]
- Regulation of the Minister of the Environment of 16 March 2005 on the definition of hunting periods for wildlife, Polish Law Gazette no. 48. 459, 2005. Internet System of Legal Documents. Available online: http://prawo.sejm.gov.pl/isap.nsf/download.xsp/WDU20050480459/O/D20050459.pdf (accessed on 30 April 2020). (In Polish)
- Regulation of the Minister of the Environment of 1 August 2017 amending the Regulation of the Minister of the Environment of 16 March 2005 on the definition of hunting periods for wildlife. Polish Law Gazette 1487, 2017. Internet System of Legal Documents. Available online: http://isap.sejm.gov.pl/isap.nsf/download.xsp/WDU20170001487/O/D20171487.pdf (accessed on 30 April 2020). (In Polish)
- European Union Reference Laboratory for E. coli. Identification and characterisation of Verotoxin-producing Escherichia coli (VTEC) by PCR amplification of the main virulence genes. 2013a. Available online: http://old.iss.it/binary/vtec/cont/EU_RL_VTEC_Method_01_Rev_0.pdf (accessed on 20 June 2017).
- Paton, A.W.; Paton, J.C. Detection and characterization of shiga toxigenic Escherichia coli by using multiplex PCR assays for stx(1), stx(2), eaeA, enterohemorrhagic E-coli hlyA, rfb(O111), and rfb(O157). J. Clin. Microbiol. 1998, 36, 598–602. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, H.; Scheef, J.; Morabito, S.; Caprioli, A.; Wieler, L.H.; Karch, H. A new Shiga toxin 2 variant (Stx2f) from Escherichia coli isolated from pigeons. Appl. Environ. Microbiol. 2000, 66, 1205–1208. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, H.; Plaschke, B.; Franke, S.; Russmann, H.; Schwarzkopf, A.; Heesemann, J.; Karch, H. Differentiation in virulenece patternes of Escherichia coli possessing eae genes. Med. Microbiol. Immun. 1994, 183, 23–31. [Google Scholar] [CrossRef]
- European Union Reference Laboratory for E. coli. Identification of VTEC serogroups mainly associated with human infections by conventional PCR amplification of O-associated genes. 2013b. Available online: http://old.iss.it/binary/vtec/cont/EU_RL_VTEC_Method_03_Rev_1.pdf (accessed on 20 June 2017).
- European Union Reference Laboratory for E. coli. Identification of the subtypes of Verocytotoxin encoding genes (vtx) of Escherichia coli by conventional PCR. 2013c. Available online: http://old.iss.it/binary/vtec/cont/EU_RL_VTEC_Method_06_Rev_1.pdf (accessed on 20 June 2017).
- Monday, S.R.; Beisaw, A.; Feng, P.C.H. Identification of Shiga toxigenic Escherichia coli seropathotypes A and B by multiplex PCR. Mol. Cell. Probe. 2007, 21, 308–311. [Google Scholar] [CrossRef]
- Durso, L.M.; Bono, J.L.; Keen, J.E. Molecular serotyping of Escherichia coli O26:H11. Appl. Environ. Microbiol. 2005, 71, 4941–4944. [Google Scholar] [CrossRef] [Green Version]
- Gannon, V.P.J.; Dsouza, S.; Graham, T.; King, R.K.; Rahn, K.; Read, S. Use of the flagellar H7 gene as a target in multiplex PCR assays and improved specificity in identification of enterohemorrhagic Escherichia coli strains. J. Clin. Microbiol. 1997, 35, 656–662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mora, A.; Lopez, C.; Dhabi, G.; Lopez-Beceiro, A.M.; Fidalgo, L.E.; Diaz, E.A.; Martinez-Carrasco, C.; Mamani, R.; Herrera, A.; Blanco, J.E.; et al. Seropathotypes, Phylogroups, Stx Subtypes, and Intimin Types of Wildlife-Carried, Shiga Toxin-Producing Escherichia coli Strains with the Same Characteristics as Human-Pathogenic Isolates. App. Environ. Microbiol. 2012, 78, 2578–2585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, L.D.; Cai, T.T.; DasGupta, A.; Agresti, A.; Coull, B.A.; Casella, G.; Corcoran, C.; Mehta, C.; Ghosh, M.; Santner, T.J. Interval estimation for a binomial proportion—Comment—Rejoinder. Stat. Sci. 2001, 16, 101–133. [Google Scholar]
- Gibbs, E.P.J. The evolution of One Health: A decade of progress and challenges for the future. Vet. Record 2014, 174, 85–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruegg, S.R.; McMahon, B.J.; Hasler, B.; Esposito, R.; Nielsen, L.R.; Speranza, C.I.; Ehlinger, T.; Peyre, M.; Aragrande, M.; Zinsstag, J.; et al. A Blueprint to evaluate One Health. Front. Public Health 2017, 5, 20. [Google Scholar] [CrossRef]
- Zinsstag, J.; Schelling, E.; Wyss, K.; Mahamat, M.B. Potential of cooperation between human and animal health to strengthen health systems. Lancet 2005, 366, 2142–2145. [Google Scholar] [CrossRef]
- Diaz-Sanchez, S.; Sanchez, S.; Sanchez, M.; Herrera-Leon, S.; Hanning, I.; Vidal, D. Detection and characterization of Shiga toxin-producing Escherichia coli in game meat and ready-to-eat meat products. Int. J. Food Microbiol. 2012, 160, 179–182. [Google Scholar] [CrossRef]
- Espinosa, L.; Gray, A.; Duffy, G.; Fanning, S.; McMahon, B.J. A scoping review on the prevalence of Shiga-toxigenic Escherichia coli in wild animal species. Zoonoses Public Health 2018, 65, 911–920. [Google Scholar] [CrossRef] [Green Version]
- Langholz, J.A.; Jay-Russell, M.T. Potential role of wildlife in pathogenic contamination of fresh produce. Hum-Wildl Interact. 2013, 7, 140–157. [Google Scholar]
- Rounds, J.M.; Rigdon, C.E.; Muhl, L.J.; Forstner, M.; Danzeisen, G.T.; Koziol, B.S.; Taylor, C.; Shaw, B.T.; Short, G.L.; Smith, K.E. Non-O157 Shiga Toxin-producing Escherichia coli Associated with Venison. Emerg. Infect. Dis. 2012, 18, 279–282. [Google Scholar] [CrossRef]
- Sanno, A.; Jacobson, M.; Sterner, S.; Thisted-Lambertz, S.; Aspan, A. The development of a screening protocol for Salmonella spp. and enteropathogenic Yersinia spp. in samples from wild boar (Sus scrofa) also generating MLVA-data for Y. enterocolitica and Y. pseudotuberculosis. J. Microbiol. Meth. 2018, 150, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Sauvala, M.; Laaksonen, S.; Laukkanen-Ninios, R.; Jalava, K.; Stephan, R.; Fredriksson-Ahomaa, M. Microbial contamination of moose (Alces alces) and white-tailed deer (Odocoileus virginianus) carcasses harvested by hunters. Food microbiol. 2019, 78, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Smith-Palmeri, A.; Hawkinsi, G.; Browningi, L.; Allison, L.; Hanson, M.; Bruce, R.; McElhiney, J.; Horne, J.; Incident Management, T. Outbreak of Escherichia coli O157 Phage Type 32 linked to the consumption of venison products. Epidemiol. Infect. 2018, 146, 1922–1927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daszkiewicz, T.; Hnatyk, N.; Dabrowski, D.; Janiszewski, P.; Gugolek, A.; Kubiak, D.; Smiecinska, K.; Winarski, R.; Koba-Kowalczyk, M. A comparison of the quality of the Longissimus lumborum muscle from wild and farm-raised fallow deer (Dama dama L.). Small Ruminant Res. 2015, 129, 77–83. [Google Scholar] [CrossRef]
- Hoffman, L.C.; Wiklund, E. Game and venison—meat for the modern consumer. Meat Sci. 2006, 74, 197–208. [Google Scholar] [CrossRef]
- Zochowska-Kujawska, J.; Kotowicz, M.; Sobczak, M.; Lachowicz, K.; Wojcik, J. Age-related changes in the carcass composition and meat quality of fallow deer (Dama Dama). Meat Sci. 2019, 147, 37–43. [Google Scholar] [CrossRef]
- Bures, D.; Barton, L.; Kotrba, R.; Hakl, J. Quality attributes and composition of meat from red deer (Cervus elaphus), fallow deer (Dama dama) and Aberdeen Angus and Holstein cattle (Bos taurus). J. Sci. Food Agric. 2015, 95, 2299–2306. [Google Scholar] [CrossRef]
- Kudrnacova, E.; Barton, L.; Bures, D.; Hoffman, L.C. Carcass and meat characteristics from farm-raised and wild fallow deer (Dama dama) and red deer (Cervus elaphus): A review. Meat Sci. 2018, 141, 9–27. [Google Scholar] [CrossRef]
- Kwiecinska, K.; Kosicka-Gebska, M.; Gebski, J.; Gutkowska, K. Prediction of the conditions for the consumption of game by Polish consumers. Meat Sci. 2017, 131, 28–33. [Google Scholar] [CrossRef]
- Singh, P.; Sha, Q.; Lacher, D.W.; Del Valle, J.; Mosci, R.E.; Moore, J.A.; Scribner, K.T.; Manning, S.D. Characterization of enteropathogenic and Shiga toxin-producing Escherichia coli in cattle and deer in a shared agroecosystem. Front. Cell. Infect. Microbiol. 2015, 5, 29. [Google Scholar] [CrossRef] [Green Version]
- Regulation (EC) No. 853/2004, 2004. of the European Parliament and of the Council of 29 April 2004 laying down specific hygiene rules for food of animal origin. Official Journal of the European Union, L 139/55. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:32004R0853&from=EN (accessed on 1 February 2019).
- Regulation (EC) No. 854/2004, 2004. of the European Parliament and of the Council of 29 April 2004 laying down specific rules for the organisation of official controls on products of animal origin intended for human consumption. Official Journal of the European Union, L 155/206. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:32004R0854&from=en (accessed on 1 February 2019).

{kind=link}
Table 1.
Pathotypes and O serogroups of Shiga toxin-producing Escherichia coli / attaching and effacing (STEC/AE)-STEC and enteropathogenic Escherichia coli (EPEC) isolated from fallow deer (Dama dama).
Table 1.
Pathotypes and O serogroups of Shiga toxin-producing Escherichia coli / attaching and effacing (STEC/AE)-STEC and enteropathogenic Escherichia coli (EPEC) isolated from fallow deer (Dama dama).
| Virulence Genes | Number of Samples | Number of Strains | stx1 Subtype (Number) | stx2 Subtype (Number) | Serogroup (Number) | Time of Collection Year/Month |
|---|---|---|---|---|---|---|
| STEC stx1 | 0 | -- | -- | -- | -- | |
| STEC stx2 | 4 | 2 | stx2a (1) stx2d (1) | ONT:HNM (1) O103:HNM (1) | 2019/Jan. 2018/Nov | |
| STEC stx1 stx2 | 2 | 2 | stx1a (1) stx1NS (1) | stx2NS (1) stx2b (1) | ONT:H7 (1) ONT:HNM (1) | 2019/Jan. 2018/Dec. |
| AE-STEC stx1 eae | 8 | 3 | stx1a (2) stx1NS (1) | O26:HNM (1), O157:H7 (1) O157:H7 (1) | 2018/Oct., 2018/Nov. 2018/Nov. | |
| AE-STEC stx2 eae | 11 | 4 | stx2b (2) stxNS (2) | O103:HNM (2) O103:HNM (1), O26:HNM (1) | 2017/Nov., 2018/Oct. 2018/Feb., 2018/Dec. | |
| AE-STEC stx1 stx2 eae | 4 | 2 | stx1NS (1) stx1NS (1) | stx2b (1) stxNS (1) | O103:HNM (1) O26:HNM (1) | 2019/Jan. 2018/Oct. |
| EPEC eae | 34 | 8 | O26:HNM (1), O103:HNM (2), O145:H7 (3), O157:H7 (1), O157:HNM (1) | 2018/Dec., 2018/Oct., 2018/Oct. 2018/Jan. (2), 2018/Feb., 2019/Jan. 2018/Nov. | ||
| Total | 63 | 21 | 7 | 10 | 21 |
NS, non-subtype; HNM, non-motile H antigen; ONT, untypable O antigen.
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Szczerba-Turek, A.; Kordas, B. Fallow Deer (Dama dama) as a Reservoir of Shiga Toxin-Producing Escherichia coli (STEC). Animals 2020, 10, 881. https://doi.org/10.3390/ani10050881
AMA Style
Szczerba-Turek A, Kordas B. Fallow Deer (Dama dama) as a Reservoir of Shiga Toxin-Producing Escherichia coli (STEC). Animals. 2020; 10(5):881. https://doi.org/10.3390/ani10050881
Chicago/Turabian StyleSzczerba-Turek, Anna, and Bernard Kordas. 2020. "Fallow Deer (Dama dama) as a Reservoir of Shiga Toxin-Producing Escherichia coli (STEC)" Animals 10, no. 5: 881. https://doi.org/10.3390/ani10050881
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.