
Anthropomorphism and Its Adverse Effects on the Distress and Welfare of Companion Animals

 ,
,  , , ,
, , ,  , ,
, , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Clothing and Its Effect on Thermoregulation
3. Restricted Mobility and Consequences for the Locomotor Apparatus
4. Exercise in Inadequate Places and Injuries
5. Alimentary Modifications and Effects on the Organism
5.1. Malnutrition: Effects on Skin and Fur
5.2. Obesity and Osteoarthritis
5.3. Malnutrition and Its Effect on Immunity
6. Application of Cosmetics and Their Effects
7. Effects of Anthropomorphism on Dog Emotions and Behavior
8. Anthropomorphism and Effects on Public Health
Anthropomorphism and Zoonosis
9. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Guthrie, S. Anthropomorphism. In Encyclopedia of Sciences and Religions; Runehov, A., Oviedo, L., Eds.; Springer: Dordrecht, The Netherlands, 2013; pp. 111–113. [Google Scholar]
- Sueur, C.; Forin-Wiart, M.-A.; Pelé, M. Are They Really Trying to Save Their Buddy? The Anthropomorphism of Animal Epimeletic Behaviours. Animals 2020, 10, 2323. [Google Scholar] [CrossRef] [PubMed]
- Darwin, C. The expression of the emotions in man and animals (1872). In The Portable Darwin; CiNii—National Institute of Informatics: Tokyo, Japan, 1993; pp. 364–393. [Google Scholar]
- Van Huis, A. Welfare of farmed insects. J. Insects Food Feed 2019, 5, 159–162. [Google Scholar] [CrossRef]
- Tondu, B. Anthropomorphism and service humanoid robots: An ambiguous relationship. Ind. Robot Int. J. 2012, 39, 609–618. [Google Scholar] [CrossRef]
- Urquiza-Haas, E.G.; Kotrschal, K. The mind behind anthropomorphic thinking: Attribution of mental states to other species. Anim. Behav. 2015, 109, 167–176. [Google Scholar] [CrossRef] [Green Version]
- Mota-Rojas, D.; Orihuela, A.; Strappini, A.; Cajiao, M.N.; Aguera, E.; Mora-Medina, P.; Ghezzi, M.D.; Alonso, S.M. Teaching animal welfare in veterinary schools in Latin America. Int. J. Vet. Sci. Med. 2018, 6, 131–140. [Google Scholar] [CrossRef]
- Horowitz, A.C.; Bekoff, M. Naturalizing Anthropomorphism: Behavioral Prompts to Our Humanizing of Animals. Anthrozoos 2007, 20, 23–35. [Google Scholar] [CrossRef]
- Kaminski, J.; Waller, B.M.; Diogo, R.; Hartstone-Rose, A.; Burrows, A.M. Evolution of facial muscle anatomy in dogs. Proc. Natl. Acad. Sci. USA 2019, 116, 14677–14681. [Google Scholar] [CrossRef] [Green Version]
- Paul, E.S.; Mendl, M.T. Animal emotion: Descriptive and prescriptive definitions and their implications for a comparative perspective. Appl. Anim. Behav. Sci. 2018, 205, 202–209. [Google Scholar] [CrossRef]
- Oftedal, L.K.; Harfeld, J.L. The Relevance of Shame in Dog-Human Relationships. J. Appl. Anim. Ethics Res. 2019, 2, 101–124. [Google Scholar] [CrossRef] [Green Version]
- Videla, M. El antropomorfismo en la relación humano-perro de compañía: ¿Recurso o indicador de patología? In Antrozoología. Potencial Recurso de Intervención Clínica; Díaz Videla, M., Olarte, A., Eds.; Editorial de la Universidad de Flores: Buenos Aires, Argentina, 2017; pp. 49–64. [Google Scholar]
- Mota-Rojas, D.; Broom, D.M.; Orihuela, A.; Velarde, A.; Napolitano, F.; Alonso-Spilsbury, M. Effects of human-animal relationship on animal productivity and welfare. J. Anim. Behav. Biometeorol. 2020, 8, 196–205. [Google Scholar] [CrossRef]
- Saucedo, K. Si no es un hijo, entonces ¿qué? Otras relaciones para ser-hacer familia a partir de la elección de no repro-ducción biológica masculina en la Ciudad de México. Rev. Ciencias Soc Humanidades Año 2020, 6, 283–306. [Google Scholar]
- Forbes, S.L. Pet Humanisation: What is it and Does it Influence Purchasing Behaviour? J. Dairy Vet. Sci. 2018, 5, 1–5. [Google Scholar] [CrossRef]
- Beynen, A. Vegetarian petfoods. Creat. Companion 2015, 51, 50–51. [Google Scholar]
- Jagoe, A.; Serpell, J. Owner characteristics and interactions and the prevalence of canine behaviour problems. Appl. Anim. Behav. Sci. 1996, 47, 31–42. [Google Scholar] [CrossRef]
- Butterfield, M.E.; Hill, S.E.; Lord, C.G. Mangy mutt or furry friend? Anthropomorphism promotes animal welfare. J. Exp. Soc. Psychol. 2012, 48, 957–960. [Google Scholar] [CrossRef]
- Castellanos, G.C.; Rodríguez, G.; Iregui, C.A. Estructura histológica normal de la piel del perro (Estado del arte). Rev. Med. Vet. (Bogota) 2005, 1, 109–122. [Google Scholar]
- Tang, K.-P.M.; Chau, K.-H.; Kan, C.-W.; Fan, J. Assessing the accumulated stickiness magnitude from fabric–skin friction: Effect of wetness level of various fabrics. R. Soc. Open Sci. 2018, 5, 180860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, N. Thermoregulation. In Cunningham’s Textbook of Veterinary Physiology; Bradley, G., Ed.; Elsevier España SL: Barcelona, Spain, 2014; pp. 559–568. [Google Scholar]
- Bruchim, Y.; Horowitz, M.; Aroch, I. Pathophysiology of Heatstroke in Dogs—Revisited. Temperature 2017, 4, 356–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foley, C. How to Prevent Heat Stroke in Dogs—Whole Dog Journal. Available online: https://www.whole-dog-journal.com/care/how-to-prevent-heat-stroke-in-dogs/ (accessed on 6 July 2021).
- Casas-Alvarado, A.; Mota-Rojas, D.; Hernández-Ávalos, I.; Mora-Medina, P.; Olmos-Hernández, A.; Verduzco-Mendoza, A.; Reyes-Sotelo, B.; Martínez-Burnes, J. Advances in infrared thermography: Surgical aspects, vascular changes, and pain monitoring in veterinary medicine. J. Therm. Biol. 2020, 92, 102664. [Google Scholar] [CrossRef]
- Villanueva-garcía, D.; Mota-rojas, D.; Olmos-, A. Hypothermia in newly born piglets: Mechanisms of thermoregulation and pathophysiology of death. Anim. Behav. Biometeorol. 2020, 9, 2112. [Google Scholar] [CrossRef]
- Cainzos, R.P.; Koscinczuk, P.; Ferreiro, M.C. Influencia de la temperatura ambiental sobre la presión arterial del perro. Rev. Vet. 2014, 25, 154. [Google Scholar] [CrossRef]
- Mota-Rojas, D.; Miranda-Córtes, A.; Casas-Alvarado, A.; Mora-Medina, P.; Boscato, L.; Hernández-Ávalos, I. Neurobiology and modulation of stress- induced hyperthermia and fever in animals. Abanico. Vet. 2021, 11, 1–17. [Google Scholar] [CrossRef]
- Bruchim, Y.; Loeb, E.; Saragusty, J.; Aroch, I. Pathological Findings in Dogs with Fatal Heatstroke. J. Comp. Pathol. 2009, 140, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Sotelo, B.; Mota-Rojas, D.; Martínez-Burnes, J.; Olmos-Hernández, A.; Hernández-Ávalos, I.; José, N.; Casas-Alvarado, A.; Gómez, J.; Mora-Medina, P. Thermal homeostasis in the newborn puppy: Behavioral and physiological responses. J. Anim. Behav. Biometeorol. 2021, 9, 1–25. [Google Scholar] [CrossRef]
- Holiday, B. Why you Shouldn’t Carry Your Small Dog. Available online: https://holidaybarn.com/blog/dangers-of-carrying-your-small-dog/ (accessed on 6 July 2021).
- Shelton, G.; Cardinet, G. Pathophysiologic basis of canine muscle disorders. J. Vet. Intern. Med. 1987, 1, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Aristizabal, E. Análisis Cinético de la Locomoción en Perros Como Metodología Diagnóstica de Enfermedades Ortopédicas; Informe de Práctica Rotatoria: Bogotá, Colombia, 2015; pp. 1–28. [Google Scholar]
- Musch, T.I.; Friedman, D.B.; Pitetti, K.H.; Haidet, G.C.; Stray-Gundersen, J.; Mitchell, J.H.; Ordway, G.A. Regional distribution of blood flow of dogs during graded dynamic exercise. J. Appl. Physiol. 1987, 63, 2269–2277. [Google Scholar] [CrossRef]
- Tøndevold, E.; Bülow, J. Bone blood flow in conscious dogs at rest and during exercise. Acta Orthop. Scand. 1983, 54, 53–57. [Google Scholar] [CrossRef] [Green Version]
- Sabanci, S.S.; Ocal, M.K. Comparison of the large muscle group widths of the pelvic limb in seven breeds of dogs. Anat. Histol. Embryol. 2018, 47, 358–363. [Google Scholar] [CrossRef]
- Thomas, L. Causas de la Atrofia Muscular en Perros. Available online: https://myanimals.com/es/causas-atrofia-muscular-perros/ (accessed on 16 September 2021).
- White, D.A.; Harkin, K.R.; Roush, J.K.; Renberg, W.C.; Biller, D. Fortetropin inhibits disuse muscle atrophy in dogs after tibial plateau leveling osteotomy. PLoS ONE 2020, 15, e0231306. [Google Scholar] [CrossRef] [Green Version]
- Chilibeck, P.D.; Sale, D.G.; Webber, C.E. Exercise and Bone Mineral Density. Sport Med. 1995, 19, 103–122. [Google Scholar] [CrossRef]
- Chevalier, X.; Richette, P. Cartílago articular normal: Anatomía, fisiología, metabolismo y envejecimiento. EMC Apar. Locomot. 2005, 38, 1–13. [Google Scholar] [CrossRef]
- Millis, D. Responses of Musculoskeletal Tissues to Disuse and Remobilization. In Canine Rehabilitation & Physical Therapy; Millis, D.L., Levine, D., Taylor, R.A., Eds.; W.B. Saunders: Philadelphia, PA, USA, 2004; pp. 113–159. [Google Scholar]
- Bhosale, A.M.; Richardson, J.B. Articular cartilage: Structure, injuries and review of management. Br. Med. Bull. 2008, 87, 77–95. [Google Scholar] [CrossRef] [PubMed]
- Miller, W.H.; Griffin, C.E.; Campbell, K.L. Muller & Kirk’s Small Animal Dermatology, 7th ed.; Elsevier: Maryland Heights, MO, USA, 2012; pp. 1–948. [Google Scholar]
- Viasus, P. Patitas: Sistemas Termosensibles de Prevención, Cuidado y Recuperación de Lesiones en Animales de Compañía Caninos. Ph.D. Thesis, Universidad de Bogotá, Bogota, Colombia, May 2020. [Google Scholar]
- Desachy, F. La Alimentación del Perro; De Vecchi, S., Ed.; De Vecchi Ediciones: Cuauhtémoc, Mexico, 2018; pp. 1–140. [Google Scholar]
- Carciofi, A.C.; Takakura, F.S.; De-Oliveira, L.D.; Teshima, E.; Jeremias, J.T.; Brunetto, M.A.; Prada, F. Effects of six carbohydrate sources on dog diet digestibility and post-prandial glucose and insulin response. J. Anim. Physiol. Anim. Nutr. 2008, 92, 326–336. [Google Scholar] [CrossRef]
- Fortes, C.M.L.S.; Carciofi, A.C.; Sakomura, N.K.; Kawauchi, I.M.; Vasconcellos, R.S. Digestibility and metabolizable energy of some carbohydrate sources for dogs. Anim. Feed Sci. Technol. 2010, 156, 121–125. [Google Scholar] [CrossRef]
- Axelsson, E.; Ratnakumar, A.; Arendt, M.-L.; Maqbool, K.; Webster, M.T.; Perloski, M.; Liberg, O.; Arnemo, J.M.; Hedhammar, Å.; Lindblad-Toh, K. The genomic signature of dog domestication reveals adaptation to a starch-rich diet. Nature 2013, 495, 360–364. [Google Scholar] [CrossRef]
- Arendt, M.; Cairns, K.M.; Ballard, J.W.O.; Savolainen, P.; Axelsson, E. Diet adaptation in dog reflects spread of prehistoric agriculture. Heredity 2016, 117, 301–306. [Google Scholar] [CrossRef] [Green Version]
- Van Herwijnen, I.R.; Corbee, R.J.; Endenburg, N.; Beerda, B.; van der Borg, J.A. Permissive parenting of the dog associates with dog overweight in a survey among 2303 Dutch dog owners. PLoS ONE 2020, 15, e0237429. [Google Scholar] [CrossRef]
- Sanderson, S.L. Taurine and Carnitine in Canine Cardiomyopathy. Vet. Clin. N. Am. Small Anim. Pract. 2006, 36, 1325–1343. [Google Scholar] [CrossRef]
- Overgaauw, P.A.M.; Vinke, C.M.; van Hagen, M.A.E.; Lipman, L.J.A. A One Health Perspective on the Human–Companion Animal Relationship with Emphasis on Zoonotic Aspects. Int. J. Environ. Res. Public Health 2020, 17, 3789. [Google Scholar] [CrossRef]
- Knight, A.; Leitsberger, M. Vegetarian versus Meat-Based Diets for Companion Animals. Animals 2016, 6, 57. [Google Scholar] [CrossRef] [Green Version]
- Zafalon, R.V.A.; Risolia, L.W.; Vendramini, T.H.A.; Ayres Rodrigues, R.B.; Pedrinelli, V.; Teixeira, F.A.; Rentas, M.F.; Perini, M.P.; Alvarenga, I.C.; Brunetto, M.A. Nutritional inadequacies in commercial vegan foods for dogs and cats. PLoS ONE 2020, 15, e0227046. [Google Scholar] [CrossRef] [PubMed]
- Laverde, H.J. Actualización de las Principales Dermotopatías en Perros y Gatos Diagnóstico y Tratamiento. Diploma Thesis, Universidad de Ciencias Aplicadas y Ambientales, Bogota, Colombia, 2018; p. 55. [Google Scholar]
- Watson, T.D.G. Diet and Skin Disease in Dogs and Cats. J. Nutr. 1998, 128, 2783S–2789S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marsh, K.A.; Ruedisueli, F.L.; Coe, S.L.; Watson, T.G.D. Effects of zinc and linoleic acid supplementation on the skin and coat quality of dogs receiving a complete and balanced diet. Vet. Dermatol. 2000, 11, 277–284. [Google Scholar] [CrossRef]
- Cao, M. How owners’ anthropomorphic tendencies associated with their feeding method towards their pets, and ultimately influence the pets’ weight status; Wageningen University: Wageningen, The Netherlands, 2014. [Google Scholar]
- German, A.J. The Growing Problem of Obesity in Dogs and Cats. J. Nutr. 2006, 136, 1940S–1946S. [Google Scholar] [CrossRef] [Green Version]
- Konok, V.; Marx, A.; Faragó, T. Attachment styles in dogs and their relationship with separation-related disorder—A questionnaire based clustering. Appl. Anim. Behav. Sci. 2019, 213, 81–90. [Google Scholar] [CrossRef]
- Sanderson, S.L. The epidemic of canine obesity and its role in osteoarthritis. Isr. J. Vet. Med. 2012, 67, 195–202. [Google Scholar]
- Bland, I.M.; Guthrie-Jones, A.; Taylor, R.D.; Hill, J. Dog obesity: Veterinary practices’ and owners’ opinions on cause and management. Prev. Vet. Med. 2010, 94, 310–315. [Google Scholar] [CrossRef]
- Gualillo, O. Further evidence for leptin involvement in cartilage homeostases. Osteoarthr. Cartil. 2007, 15, 857–860. [Google Scholar] [CrossRef] [Green Version]
- Marshall, W.G.; Hazewinkel, H.A.W.; Mullen, D.; De Meyer, G.; Baert, K.; Carmichael, S. The effect of weight loss on lameness in obese dogs with osteoarthritis. Vet. Res. Commun. 2010, 34, 241–253. [Google Scholar] [CrossRef] [Green Version]
- Musco, N.; Vassalotti, G.; Mastellone, V.; Cortese, L.; della Rocca, G.; Molinari, M.L.; Calabrò, S.; Tudisco, R.; Cutrignelli, M.I.; Lombardi, P. Effects of a nutritional supplement in dogs affected by osteoarthritis. Vet. Med. Sci. 2019, 5, 325–335. [Google Scholar] [CrossRef] [Green Version]
- Diehm, M.; Tünsmeyer, J. Multimodale Therapie der kaninen Osteoarthritis—Aktueller Stand und neue Optionen. Tierärztliche Prax Ausgabe K Kleintiere / Heimtiere 2018, 46, 337–345. [Google Scholar] [CrossRef]
- Lipari, L.; Gerbino, A. Expression of Gelatinases (MMP-2, MMP-9) in Human Articular Cartilage. Int. J. Immunopathol. Pharmacol. 2013, 26, 817–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vincent, T.L. IL-1 in osteoarthritis: Time for a critical review of the literature. F1000Research 2019, 8, 934. [Google Scholar] [CrossRef] [PubMed]
- Wernimont, S.M.; Radosevich, J.; Jackson, M.I.; Ephraim, E.; Badri, D.V.; MacLeay, J.M.; Jewell, D.E.; Suchodolski, J.S. The Effects of Nutrition on the Gastrointestinal Microbiome of Cats and Dogs: Impact on Health and Disease. Front. Microbiol. 2020, 11, 1–24. [Google Scholar] [CrossRef]
- Weeth, L. Focus on nutrition: Home-prepared diets for dogs and cats. Compendium 2012, 35, E1–E3. [Google Scholar]
- Blaise, M. Situating Hong Kong Pet-Dog-Child Figures within Colonialist Flows and Disjunctures. Glob. Stud. Child. 2013, 3, 380–394. [Google Scholar] [CrossRef] [Green Version]
- Brockman, B.K.; Taylor, V.A.; Brockman, C.M. The price of unconditional love: Consumer decision making for high-dollar veterinary care. J. Bus. Res. 2008, 61, 397–405. [Google Scholar] [CrossRef]
- De Lapuente, J.; Borras, M.; González-Linares, J.; Llanas, H.; Mitjans, M.; Ramos-López, D.; Vinardell, P. Los métodos alternativos en el estudio de la seguridad de cosméticos. Rev. Toxicol. 2014, 31, 140–148. [Google Scholar]
- Chomel, B.B.; Sun, B. Zoonoses in the Bedroom. Emerg. Infect. Dis. 2011, 17, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Jardes, D.J.; Ross, L.A.; Markovich, J.E. Hemolytic anemia after ingestion of the natural hair dye Lawsonia inermis (henna) in a dog. J. Vet. Emerg. Crit. Care 2013, 23, 648–651. [Google Scholar] [CrossRef]
- Li, J.; Liu, J. Orthogonal experiment design to optimize the pet dog dyeing condition of Musa basjoo Siebold (MBS) & Lithospermum erythrorhizon (LE) mixed hair dye. Med. Plant 2018, 9, 114–118. [Google Scholar]
- Buehler, J. Dogs Really Can Smell Your Fear. Available online: https://www.newscientist.com/article/2150956-dogs-really-can-smell-your-fear-and-then-they-get-scared-too/ (accessed on 16 September 2021).
- Berns, G.S.; Brooks, A.M.; Spivak, M. Scent of the familiar: An fMRI study of canine brain responses to familiar and unfamiliar human and dog odors. Behav. Process. 2015, 110, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Baran, R. Nail cosmetics: Allergies and irritations. Am. J. Clin. Dermatol. 2002, 3, 547–555. [Google Scholar] [CrossRef] [PubMed]
- Young, A.S.; Allen, J.G.; Kim, U.-J.; Seller, S.; Webster, T.F.; Kannan, K.; Ceballos, D.M. Phthalate and Organophosphate Plasticizers in Nail Polish: Evaluation of Labels and Ingredients. Environ. Sci. Technol. 2018, 52, 12841–12850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellingsen, K.; Zanella, A.J.; Bjerkås, E.; Indrebø, A. The Relationship between empathy, perception of pain and attitudes toward pets among Norwegian dog owners. Anthrozoos 2010, 23, 231–243. [Google Scholar] [CrossRef]
- Travain, T.; Colombo, E.S.; Heinzl, E.; Bellucci, D.; Prato Previde, E.; Valsecchi, P. Hot dogs: Thermography in the assessment of stress in dogs (Canis familiaris)—A pilot study. J. Vet. Behav. 2015, 10, 17–23. [Google Scholar] [CrossRef]
- Voith, V.L.; Wright, J.C.; Danneman, P.J. Is there a relationship between canine behavior problems and spoiling activities, anthropomorphism, and obedience training? Appl. Anim. Behav. Sci. 1992, 34, 263–272. [Google Scholar] [CrossRef]
- Rooney, N.; Bradshaw, J. Canine welfare science: An antidote to sentiment and myth. In Domestic Dog Cognition and Behavior: The Scientific Study of Canis Familiaris; Horowitz, A., Ed.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 241–274. [Google Scholar]
- Morris, P.H.; Doe, C.; Godsell, E. Secondary emotions in non-primate species? Behavioural reports and subjective claims by animal owners. Cogn. Emot. 2008, 22, 3–20. [Google Scholar] [CrossRef]
- Hecht, J.; Miklósi, Á.; Gácsi, M. Behavioral assessment and owner perceptions of behaviors associated with guilt in dogs. Appl. Anim. Behav. Sci. 2012, 139, 134–142. [Google Scholar] [CrossRef]
- Vollmer, P.J. Do mischievous dogs reveal their “guilt”? Vet. Med. Small Anim. Clin. 1977, 72, 1002–1005. [Google Scholar]
- Horowitz, A. Disambiguating the “guilty look”: Salient prompts to a familiar dog behaviour. Behav. Process. 2009, 81, 447–452. [Google Scholar] [CrossRef] [PubMed]
- Rajecki, D.; Rasmussen, J.L.; Sanders, C.R.; Modlin, S.J.; Holder, A.M. Good Dog: Aspects of humans’ causal attributions for a companion animal’s ocial behavior. Soc. Anim. 1999, 7, 17–34. [Google Scholar] [CrossRef]
- Mariti, C.; Ricci, E.; Zilocchi, M.; Gazzano, A. Owners as a secure base for their dogs. Behaviour 2013, 150, 1275–1294. [Google Scholar] [CrossRef]
- Solomon, J.; Beetz, A.; Schöberl, I.; Gee, N.; Kotrschal, K. Attachment security in companion dogs: Adaptation of Ainsworth’s strange situation and classification procedures to dogs and their human caregivers. Attach. Hum. Dev. 2019, 21, 389–417. [Google Scholar] [CrossRef] [PubMed]
- Riggio, G. A mini review on the dog-owner attachment bond and its implications in veterinary clinical ethology. Dog. Behav. 2020, 6, 17–26. [Google Scholar] [CrossRef]
- Riggio, G.; Gazzano, A.; Zsilák, B.; Carlone, B.; Mariti, C. Quantitative behavioral analysis and qualitative classification of attachment styles in comestic dogs: Are dogs with a secure and an Insecure-Avoidant attachment different? Animals 2020, 11, 14. [Google Scholar] [CrossRef]
- Muris, P.; Mayer, B.; Meesters, C. Self-reported attachment style, anxiety, and depression in children. Soc. Behav. Pers. Int. J. 2000, 28, 157–162. [Google Scholar] [CrossRef]
- Schimmenti, A.; Bifulco, A. Linking lack of care in childhood to anxiety disorders in emerging adulthood: The role of attachment styles. Child Adolesc. Ment. Health 2015, 20, 41–48. [Google Scholar] [CrossRef]
- Spruit, A.; Goos, L.; Weenink, N.; Rodenburg, R.; Niemeyer, H.; Stams, G.J.; Colonnesi, C. The relation between attachment and depression in children and adolescents: A multilevel meta-analysis. Clin. Child Fam. Psychol. Rev. 2020, 23, 54–69. [Google Scholar] [CrossRef] [Green Version]
- Manicavasagar, V.; Silove, D.; Marnane, C.; Wagner, R. Adult attachment styles in panic disorder with and without comorbid adult separation anxiety disorder. Aust. N. Z. J. Psychiatry 2009, 43, 167–172. [Google Scholar] [CrossRef]
- Mikulincer, M.; Shaver, P.R. Attachment, anger, and aggression. In Human Aggression and Violence: Causes, Manifestations, and Consequences; Shaver, P.R., Mikulincer, M., Eds.; American Psychological Association: Washington, DC, USA, 2010; pp. 241–257. [Google Scholar]
- van Leeuwen, W.A.; van Wingen, G.A.; Luyten, P.; Denys, D.; van Marle, H.J.F. Attachment in OCD: A meta-analysis. J. Anxiety Disord. 2020, 70, 102187. [Google Scholar] [CrossRef]
- Landsberg, G.; Hunthausen, W.; Ackerman, L. Behavior Problems of the Dog and Cat, 3rd ed.; Saunders Elsevier: Edinburgh, Scotland, 2011; pp. 1–454. [Google Scholar]
- Cavalcanti, A.L.; Porto, E.; dos Santos, B.F.; Cavalcanti, C.L.; Cavalcanti, A.F.C. Facial dog bite injuries in children: A case report. Int. J. Surg. Case Rep. 2017, 41, 57–60. [Google Scholar] [CrossRef] [PubMed]
- Meints, K.; Brelsford, V.; De Keuster, T. Teaching children and parents to understand dog signaling. Front. Vet. Sci. 2018, 5, 1–13. [Google Scholar] [CrossRef]
- Chapman, S.; Cornwall, J.; Righetti, J.; Sung, L. Preventing dog bites in children: Randomised controlled trial of an educational intervention. BMJ Br. Med. J. 2000, 320, 1512–571513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, F.; Dwyer, F.; Bennett, P.C. Prevention of dog bites: Evaluation of a brief educational intervention program for preschool children. J. Community Psychol. 2003, 31, 75–86. [Google Scholar] [CrossRef]
- De Keuster, T.; Lamoureux, J.; Kahn, A. Epidemiology of dog bites: A Belgian experience of canine behaviour and public health concerns. Vet. J. 2006, 172, 482–487. [Google Scholar] [CrossRef]
- Serpell, J. Anthropomorphism and Anthropomorphic Selection—Beyond the ‘Cute Response’. Soc. Anim. 2002, 10, 437–454. [Google Scholar] [CrossRef]
- Steinert, K.; Kuhne, F.; Kramer, M.; Hackbarth, H. People’s perception of brachycephalic breeds and breed-related welfare problems in Germany. J. Vet. Behav. 2019, 33, 96–102. [Google Scholar] [CrossRef]
- Aromaa, M.; Lilja-Maula, L.; Rajamäki, M. Assessment of welfare and brachycephalic obstructive airway syndrome signs in young, breeding age French Bulldogs and Pugs, using owner questionnaire, physical examination and walk tests. Anim. Welf 2019, 28, 287–298. [Google Scholar] [CrossRef]
- Bartels, A.; Martin, V.; Bidoli, E.; Steigmeier-Raith, S.; Brühschwein, A.; Reese, S.; Köstlin, R.; Erhard, M. Brachycephalic problems of pugs relevant to animal welfare. Anim. Welf 2015, 24, 327–333. [Google Scholar] [CrossRef]
- Schatz, K.Z.; Engelke, E.; Pfarrer, C. Comparative morphometric study of the mimic facial muscles of brachycephalic and dolichocephalic dogs. Anat. Histol. Embryol. 2021, 50, 863–875. [Google Scholar] [CrossRef] [PubMed]
- Chorpita, B.F.; Barlow, D.H. The development of anxiety: The role of control in the early environment. Psychol. Bull. 1998, 124, 3–21. [Google Scholar] [CrossRef] [Green Version]
- Harrison, M.A.; Hall, A.E. Anthropomorphism, empathy, and perceived communicative ability vary with phylogenetic relatedness to humans. J. Soc. Evol. Cult. Psychol. 2010, 4, 34–48. [Google Scholar] [CrossRef]
- Chan, A.A.Y.-H. Anthropomorphism as a conservation tool. Biodivers. Conserv. 2012, 21, 1889–1892. [Google Scholar] [CrossRef]
- Morton, D.B.; Berghardt, G.M.; Smith, J.A. Critical anthropomorphism, animal suffering, and the ecological context. Hastings Cent. Rep. 1990, 20, S13. [Google Scholar]
- Parthasarathy, V.; Crowell-Davis, S.L. Relationship between attachment to owners and separation anxiety in pet dogs (Canis lupus familiaris). J. Vet. Behav. 2006, 1, 109–120. [Google Scholar] [CrossRef] [Green Version]
- Romero, N.Y. Relación Entre el Apego Humano-Mascota y el Sentimiento de Soledad (en adultos mayores). Diploma Thesis, Facultad De Ciencias Juridicas Y Sociales Universidad Argentina de la Empresa, Buenos Aires, Argentina, March 2016. [Google Scholar]
- Monroy Gonzalez, J.C.; Almeyda Remolina, Á.G.; Bernal Loaiza, P.A. La Relación Humano-Animal en el ámbito Comunitario y Familiar: Apuestas Para el Abordaje Desde Trabajo Social. Diploma Thesis, Universidad de la Salle, Bogotá, Colombia, 2019. [Google Scholar]
- Meléndez El vínculo humano-animal y sus implicaciones para la psicología en Puerto Rico. Rev. Puertorriquena Psicol. 2014, 25, 160–182.
- Baun, M.; Bergstrom, N.; Langston, N.; Thoma, L. Physiological effects of human/companion animal bonding. Nurs. Res. 1984, 33, 126–129. [Google Scholar] [CrossRef]
- Cherniack, E.P.; Cherniack, A.R. The Benefit of Pets and Animal-Assisted Therapy to the Health of Older Individuals. Curr. Gerontol. Geriatr. Res. 2014, 2014, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Clark, K.M. Psychological Effects of Dog Ownership: Role Strain, Role Enhancement, and Depression. J. Soc. Psychol. 2010, 150, 117–131. [Google Scholar] [CrossRef]
- Kidd, A.H.; Kidd, R.M. Benefits and Liabilities of Pets for the Homeless. Psychol. Rep. 1994, 74, 715–722. [Google Scholar] [CrossRef] [PubMed]
- Kanat-Maymon, Y.; Antebi, A.; Zilcha-Mano, S. Basic psychological need fulfillment in human–pet relationships and well-being. Pers. Individ. Dif. 2016, 92, 69–73. [Google Scholar] [CrossRef]
- Day, M.J.; Breitschwerdt, E.; Cleaveland, S.; Karkare, U.; Khanna, C.; Kirpensteijn, J.; Kuiken, T.; Lappin, M.R.; McQuiston, J.; Mumford, E.; et al. Surveillance of Zoonotic Infectious Disease Transmitted by Small Companion Animals. Emerg. Infect. Dis. 2012, 18. [Google Scholar] [CrossRef]
- Breedlove, B.; Arguin, P.M. Anthropomorphism to Zoonoses: Two Inevitable Consequences of Human–Animal Relationships. Emerg. Infect. Dis. 2015, 21, 2282–2283. [Google Scholar] [CrossRef] [Green Version]
- Vega-Aragón, R.L. Zoonosis emergentes y reemergentes y principios básicos de control de zoonosis. Rev. Med. Vet. (Bogota) 2009, 1, 85–97. [Google Scholar]
- Jofré, L.; Noemí, I.; Neira, P.; Saavedra, T.; Díaz, C. Acarosis y zoonosis relacionadas. Rev. Chil. Infectología 2009, 26, 248–257. [Google Scholar] [CrossRef] [Green Version]
- Saevik, B.K.; Bredal, W.; Ulstein, T.L. Cheyletiella infestation in the dog: Observations on diagnostic methods and clinical signs. J. Small Anim. Pract. 2004, 45, 495–500. [Google Scholar] [CrossRef]
- Reynolds, H.H.; Elston, D.M. What’s eating you? Cheyletiella mites. Cutis 2017, 99, 355. [Google Scholar]
- Bourhy, H.; Dacheux, L.; Strady, C.; Mailles, A. Rabies in Europe in 2005. Eurosurveillance 2005, 10, 3–4. [Google Scholar] [CrossRef]
- Jiang, P.; Zhang, X.; Liu, R.D.; Wang, Z.Q.; Cui, J. A Human Case of Zoonotic Dog Tapeworm, Dipylidium caninum (Eucestoda: Dilepidiidae), in China. Korean J. Parasitol. 2017, 55, 61–64. [Google Scholar] [CrossRef] [Green Version]
- Butler, T. Capnocytophaga canimorsus: An emerging cause of sepsis, meningitis, and post-splenectomy infection after dog bites. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 1271–1280. [Google Scholar] [CrossRef] [PubMed]
- Peters, V.; Sottiaux, M.; Appelboom, J.; Kahn, A. Posttraumatic stress disorder after dog bites in children. J. Pediatr. 2004, 144, 121–122. [Google Scholar] [CrossRef] [PubMed]
- Mota-Rojas, D.; Calderón-Maldonado, N.; Lezama-García, K.; Sepiurka, L.; Garcia, R.C.M. Abandonment of dogs in Latin America: Strategies and ideas. Vet. World 2021, 14, 2371–2379. [Google Scholar] [CrossRef]







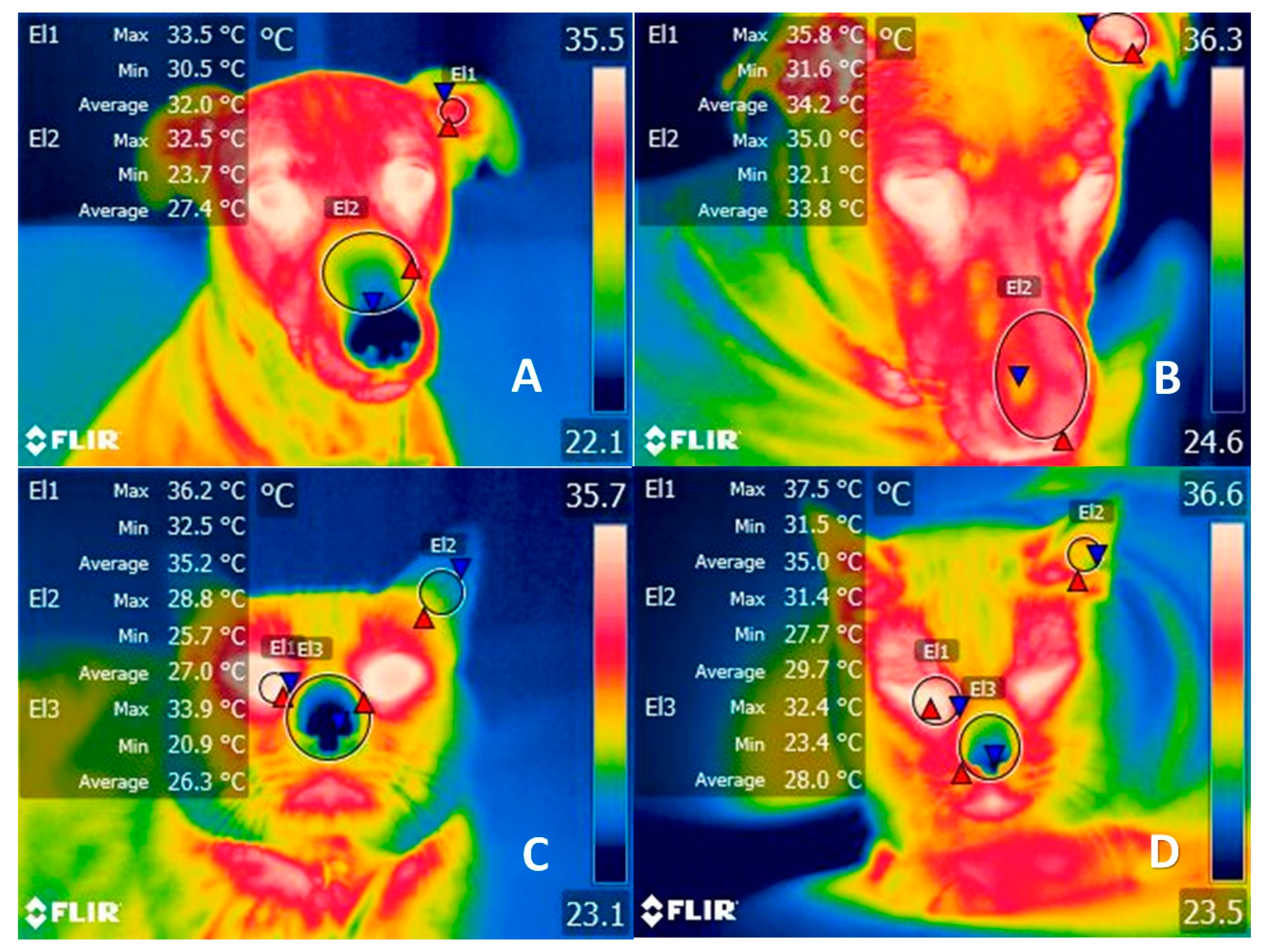{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
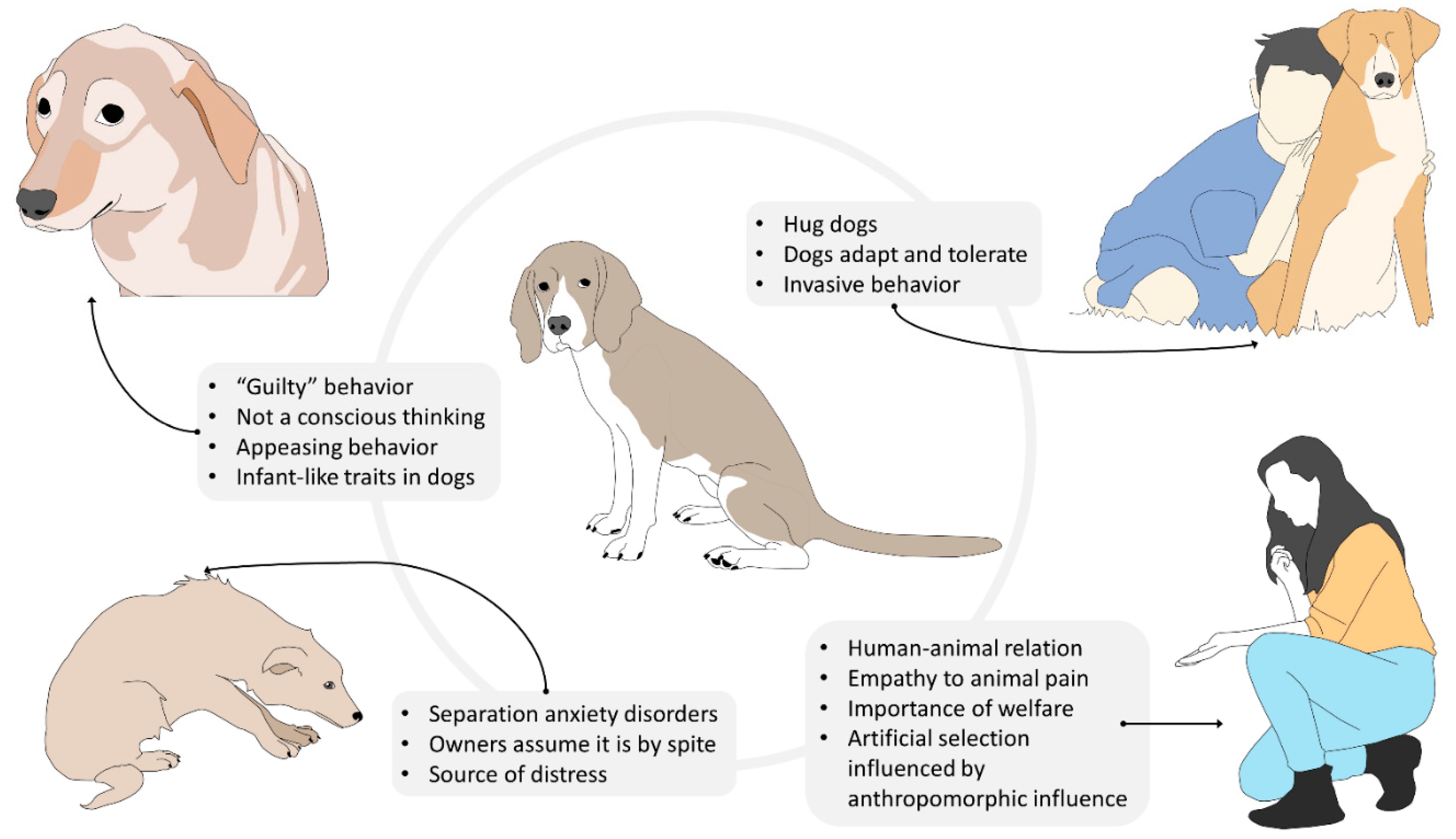{kind=link}
| Positive Effects of Interaction and Connections between Humans and Animals | References |
|---|---|
| Generate feelings of friendship | [73] |
| Hypotension | [117] |
| Reduced systolic and diastolic blood pressure | [118] |
| Reduced stress and anxiety | [117] |
| Effect on neurotransmitters and increased oxytocin concentrations | [117] |
| Provide psychological and social support | [117,119] |
| Reduced signology associated with depression | [117,120] |
| Improved self-esteem and mood, more physical exercise, and lower frequency of feelings of negativity in the face of social rejection | [117] |
| Increased social interaction | [117] |
| Sharing spare time or hours of rest as a source of psychological consolation | [73] |
| Minimized feelings of loneliness in older adults | [115] |
| Improved mental and physical health of homeless people | [121] |
| Disease | Etiologic Agent | Principal Practice | Animal Species Involved | Signology in Humans | Reference |
|---|---|---|---|---|---|
| Cheyletiellosis | Mites: Cheyletigella: Cheyletiella blackei in cats, Cheyletiella yasguri in dogs, Cheyletiella parasotivorax in rabbits | Direct contact or through fomites such as floors, towels, carpets, and beds | Dogs, cats, and rabbits | Highly contagious, non-suppurating, exfoliative dermatitis, called “walking dandruff” | [54,126,127,128] |
| Infection | Bacteria Staphylococcus intermedius (common inhabitant of saliva) | Licking of ears and face | Dogs and cats | Infection in the post-mastoidectomy mastoid cavity by medium chronic otitis with cholesteatoma; Purulent sinus infection post- endoscopic resection of pituitary adenoma | [73] |
| Infection | Methicillin-resistant bacteria Staphylococcus aureus | Licking of face and sleeping with the pet | Dogs | Recurrent nasal infection | [73] |
| Rabia | Virus Lyssavirus, genotype 1 | Bites, scratches, or licking of mucus membranes | Dogs | Lethal encephalitis | [73,129] |
| Toxocariasis | Parasite Toxocara canis Toxocara felis | Direct contact with eggs on the skin or hair, or with feces (kissing, licking, sleeping with pets) | Dogs and cats | Fever, cough, abdominal pain, anorexia, hepatomegaly, vision problems, lymphadenitis | [73] |
| Dipilidiasis | Parasite Dipylidium caninum | Direct contact with eggs due to ingestion of the infested flea Ctenocephalides felis, Ctenocephalides canis (licking or kissing the pet) | Dogs and cats | Often asymptomatic, but may show signs such as anal itching, diarrhea, moderate abdominal pain (habitually epigastric), hyporexia, indigestion, and gastrointestinal disfunctions. Occasionally, urticaria, eosinophilia, irritability, and intestinal obstruction | [130] |
| Infection | Pathogenic bacteria Capnocytophaga canimorsus (normal inhabitant of saliva) | Bites by the pet | Dogs and cats | Emergent sepsis infection, meningitis, and post-splenectomy infection after dog bites | [131] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mota-Rojas, D.; Mariti, C.; Zdeinert, A.; Riggio, G.; Mora-Medina, P.; del Mar Reyes, A.; Gazzano, A.; Domínguez-Oliva, A.; Lezama-García, K.; José-Pérez, N.; et al. Anthropomorphism and Its Adverse Effects on the Distress and Welfare of Companion Animals. Animals 2021, 11, 3263. https://doi.org/10.3390/ani11113263
Mota-Rojas D, Mariti C, Zdeinert A, Riggio G, Mora-Medina P, del Mar Reyes A, Gazzano A, Domínguez-Oliva A, Lezama-García K, José-Pérez N, et al. Anthropomorphism and Its Adverse Effects on the Distress and Welfare of Companion Animals. Animals. 2021; 11(11):3263. https://doi.org/10.3390/ani11113263
Chicago/Turabian StyleMota-Rojas, Daniel, Chiara Mariti, Andrea Zdeinert, Giacomo Riggio, Patricia Mora-Medina, Alondra del Mar Reyes, Angelo Gazzano, Adriana Domínguez-Oliva, Karina Lezama-García, Nancy José-Pérez, and et al. 2021. "Anthropomorphism and Its Adverse Effects on the Distress and Welfare of Companion Animals" Animals 11, no. 11: 3263. https://doi.org/10.3390/ani11113263