
Estradiol Priming Potentiates the Kisspeptin-Induced Release of LH in Ovariectomized Cows

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Procedures
2.2. Hormone Quantification
2.3. Statistical Analyses
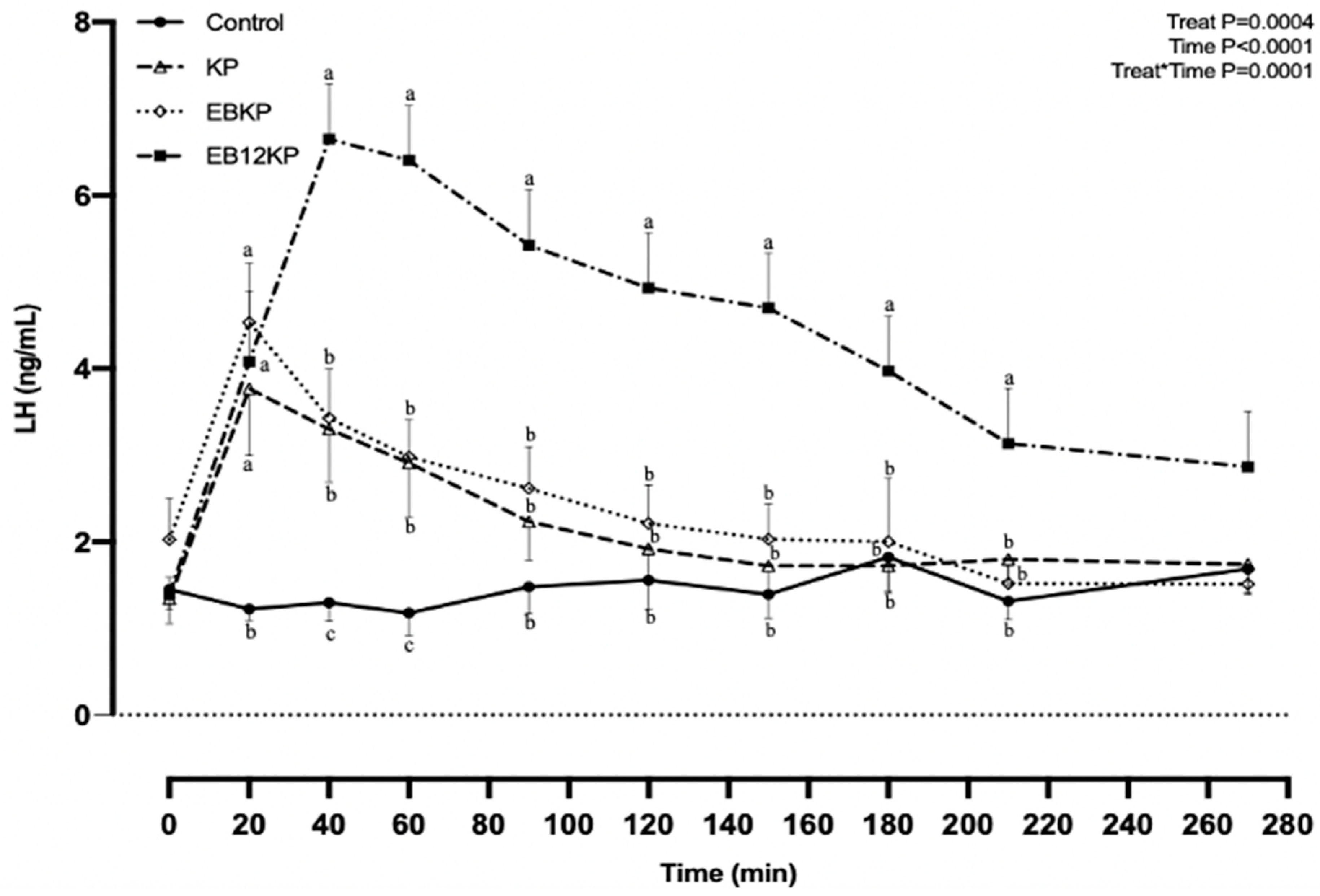
3. Results
3.1. Progesterone
3.2. LH
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Bó, G.A.; Huguenine, E.; De La Mata, J.J.; Núñez-Olivera, R.; Baruselli, P.S.; Menchaca, A. Programs for fixed-time artificial insemination in South American beef cattle. Anim. Reprod. 2018, 15, 952–962. [Google Scholar] [CrossRef]
- Burton, R.J.F.; Farstad, M. Cultural Lock-in and Mitigating Greenhouse Gas Emissions: The Case of Dairy/Beef Farmers in Norway. Sociol. Rural. 2020, 60, 20–39. [Google Scholar] [CrossRef]
- Vasconcelos, J.L.M.; Carvalho, R.; Peres, R.F.G.; Rodrigues, A.D.P.; Junior, I.C.; Meneghetti, M.; Aono, F.H.; Costa, W.M.; Lopes, C.N.; Cooke, R.F.; et al. Reproductive programs for beef cattle: Incorporating management and reproductive techniques for better fertility. Anim. Reprod. 2017, 14, 547–557. [Google Scholar] [CrossRef]
- Randel, R.D.; Short, R.E.; Christensen, D.S.; Bellows, R.A. Effects of Various Mating Stimuli on the LH Surge and Ovulation Time following Synchronization of Estrus in the Bovine. J. Anim. Sci. 1973, 37, 128–130. [Google Scholar] [CrossRef] [PubMed]
- Wiltbank, M.C.; Pursley, J.R. The cow as an induced ovulator: Timed AI after synchronization of ovulation. Theriogenology 2014, 81, 170–185. [Google Scholar] [CrossRef] [PubMed]
- Sales, J.N.S.; Carvalho, J.B.P.; Crepaldi, G.A.; Cipriano, R.S.; Jacomini, J.O.; Maio, J.R.G.; Souza, J.C.; Nogueira, G.P.; Baruselli, P.S. Effects of two estradiol esters (benzoate and cypionate) on the induction of synchronized ovulations in Bos indicus cows submitted to a timed artificial insemination protocol. Theriogenology 2012, 78, 510–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Ispierto, I.; De Rensis, F.; Casas, X.; Caballero, F.; Serrano-Pérez, B.; López-Gatius, F. Inducing ovulation with hCG in a five-day progesterone-based fixed-time AI protocol improves the fertility of anestrous dairy cows under heat stress conditions. Theriogenology 2019, 124, 65–68. [Google Scholar] [CrossRef]
- Sá Filho, M.F.; Ayres, H.; Ferreira, R.M.; Marques, M.O.; Reis, E.L.; Silva, R.C.P.; Rodrigues, C.A.; Madureira, E.H.; Bó, G.A.; Baruselli, P.S. Equine chorionic gonadotropin and gonadotropin-releasing hormone enhance fertility in a norgestomet-based, timed artificial insemination protocol in suckled Nelore (Bos indicus) cows. Theriogenology 2010, 73, 651–658. [Google Scholar] [CrossRef]
- Bó, G.A.; de la Mata, J.J.; Baruselli, P.S.; Menchaca, A. Alternative programs for synchronizing and resynchronizing ovulation in beef cattle. Theriogenology 2016, 86, 388–396. [Google Scholar] [CrossRef]
- Bó, G.A.; Baruselli, P.S. Synchronization of ovulation and fixed-time artificial insemination in beef cattle. Animal 2014, 8, 144–150. [Google Scholar] [CrossRef] [Green Version]
- Bisinotto, R.S.; Ribeiro, E.S.; Santos, J.E.P. Synchronisation of ovulation for management of reproduction in dairy cows. Animal 2014, 8, 151–159. [Google Scholar] [CrossRef] [Green Version]
- Cardoso Consentini, C.E.; Wiltbank, M.C.; Sartori, R. Factors That Optimize Reproductive Efficiency in Dairy Herds with an Emphasis on Timed Artificial Insemination Programs. Animals 2021, 11, 301. [Google Scholar] [CrossRef]
- Macedo, G.G.; Mingoti, R.D.; Batista, E.O.S.; Monteiro, B.M.; Vieira, L.M.; Barletta, R.V.; Wiltbank, M.C.; Nogueira, G.P.; Rennó, F.P.; Maio, J.R.; et al. Profile of LH release in response to intramuscular treatment with kisspeptin in Bos indicus and Bos taurus prepubertal heifers. Theriogenology 2019, 125, 64–70. [Google Scholar] [CrossRef]
- Sá Filho, M.F.; Penteado, L.; Siqueira, G.R.; Soares, J.G.; Mendanha, M.F.; Macedo, G.G.; Baruselli, P.S. Timed artificial insemination should be performed early when used norgestomet ear implants are applied for synchronizing ovulation in beef heifers. Theriogenology 2013, 80, 642–647. [Google Scholar] [CrossRef] [Green Version]
- Redmond, J.S.; Macedo, G.G.; Velez, I.C.; Caraty, A.; Williams, G.L.; Amstalden, M. Kisspeptin activates the hypothalamic-adenohypophyseal-gonadal axis in prepubertal ewe lambs. Reproduction 2011, 141, 541–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, J.T.; Rao, A.; Pereira, A.; Caraty, A.; Millar, R.P.; Clarke, I.J. Kisspeptin Is Present in Ovine Hypophysial Portal Blood But Does Not Increase during the Preovulatory Luteinizing Hormone Surge: Evidence that Gonadotropes Are Not Direct Targets of Kisspeptin in Vivo. Endocrinology 2008, 149, 1951–1959. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Pascual, E.; Martínez-Fuentes, A.J.; Pinilla, L.; Tena-Sempere, M.; Malagón, M.M.; Castaño, J.P. Direct Pituitary Effects of Kisspeptin: Activation of Gonadotrophs and Somatotrophs and Stimulation of Luteinising Hormone and Growth Hormone Secretion. J. Neuroendocrinol. 2007, 19, 521–530. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, S.; Kadokawa, H.; Hashizume, T. Direct kisspeptin-10 stimulation on luteinizing hormone secretion from bovine and porcine anterior pituitary cells. Anim. Reprod. Sci. 2008, 103, 360–365. [Google Scholar] [CrossRef]
- Gahete, M.D.; Vázquez-Borrego, M.C.; Martínez-Fuentes, A.J.; Tena-Sempere, M.; Castaño, J.P.; Luque, R.M. Role of the Kiss1/Kiss1r system in the regulation of pituitary cell function. Mol. Cell. Endocrinol. 2016, 438, 100–106. [Google Scholar] [CrossRef]
- Souza, A.H.; Cunha, A.P.; Silva, E.P.B.; Gümen, A.; Ayres, H.; Guenther, J.N.; Wiltbank, M.C. Comparison of gonadorelin products in lactating dairy cows: Efficacy based on induction of ovulation of an accessory follicle and circulating luteinizing hormone profiles. Theriogenology 2009, 72, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Skarda, R.T. Local and Regional Anesthesia in Ruminants and Swine. Vet. Clin. N. Am. Food Anim. Pract. 1996, 12, 579–626. [Google Scholar] [CrossRef]
- Bolt, D.J.; Scott, V.; Kiracofe, G.H. Plasma LH and FSH after estradiol, norgestomet and Gn-RH treatment in ovariectomized beef heifers. Anim. Reprod. Sci. 1990, 23, 263–271. [Google Scholar] [CrossRef]
- Martínez, M.F.; Kastelic, J.P.; Colazo, M.G.; Mapletoft, R.J. Effects of estradiol on gonadotrophin release, estrus and ovulation in CIDR-treated beef cattle. Domest. Anim. Endocrinol. 2007, 33, 77–90. [Google Scholar] [CrossRef]
- Schoenemann, H.M.; Humphrey, W.D.; Crowder, M.E.; Nett, T.M.; Reeves, J.J. Pituitary Luteinizing Hormone-releasing Hormone Receptors in Ovariectomized Cows after Challenge with Ovarian Steroids. Biol. Reprod. 1985, 32, 574–583. [Google Scholar] [CrossRef] [Green Version]
- Kesner, J.S.; Convey, E.M.; Anderson, C.R. Evidence that Estradiol Induces the Preovulatory LH Surge in Cattle by Increasing Pituitary Sensitivity to LHRH and then Increasing LHRH Release. Endocrinology 1981, 108, 1386–1391. [Google Scholar] [CrossRef]
- Clarke, I.J.; Cummins, J.T. GnRH pulse frequency determines LH pulse amplitude by altering the amount of releasable LH in the pituitary glands of ewes. J. Reprod. Fertil. 1985, 73, 425–431. [Google Scholar] [CrossRef]
- Klenke, U.; Constantin, S.; Wray, S. Neuropeptide Y Directly Inhibits Neuronal Activity in a Subpopulation of Gonadotropin-Releasing Hormone-1 Neurons via Y1 Receptors. Endocrinology 2010, 151, 2736–2746. [Google Scholar] [CrossRef]
- Ericson, M.D.; Lensing, C.J.; Fleming, K.A.; Schlasner, K.N.; Doering, S.R.; Haskell-Luevano, C. Bench-top to clinical therapies: A review of melanocortin ligands from 1954 to 2016. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 2414–2435. [Google Scholar] [CrossRef]
- Advis, J.P.; Klein, J.; Kuljis, R.O.; Sarkar, D.K.; McDonald, J.M.; Conover, C.A. Regulation of Gonadotropin Releasing Hormone Release by Neuropeptide Y at the Median Eminence during the Preovulatory Period in Ewes. Neuroendocrinology 2003, 77, 246–257. [Google Scholar] [CrossRef]
- Roa, J.; Herbison, A.E. Direct Regulation of GnRH Neuron Excitability by Arcuate Nucleus POMC and NPY Neuron Neuropeptides in Female Mice. Endocrinology 2012, 153, 5587–5599. [Google Scholar] [CrossRef] [Green Version]
- Bauer-Dantoin, A.C.; Urban, J.H.; Levine, J.E. Neuropeptide Y gene expression in the arcuate nucleus is increased during preovulatory luteinizing hormone surges. Endocrinology 1992, 131, 2953–2958. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Nedungadi, T.P.; Zhu, L.; Sobhani, N.; Irani, B.G.; Davis, K.E.; Zhang, X.; Zou, F.; Gent, L.M.; Hahner, L.D.; et al. Distinct Hypothalamic Neurons Mediate Estrogenic Effects on Energy Homeostasis and Reproduction. Cell Metab. 2011, 14, 453–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irwig, M.S.; Fraley, G.S.; Smith, J.T.; Acohido, B.V.; Popa, S.M.; Cunningham, M.J.; Gottsch, M.L.; Clifton, D.K.; Steiner, R.A. Kisspeptin Activation of Gonadotropin Releasing Hormone Neurons and Regulation of KiSS-1 mRNA in the Male Rat. Neuroendocrinology 2004, 80, 264–272. [Google Scholar] [CrossRef]
- Gottsch, M.L.; Cunningham, M.J.; Smith, J.T.; Popa, S.M.; Acohido, B.V.; Crowley, W.F.; Seminara, S.; Clifton, D.K.; Steiner, R.A. A Role for Kisspeptins in the Regulation of Gonadotropin Secretion in the Mouse. Endocrinology 2004, 145, 4073–4077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarro, V.M.; Castellano, J.M.; Fernndez-Fernndez, R.; Barreiro, M.L.; Roa, J.; Sanchez-Criado, J.E.; Aguilar, E.; Dieguez, C.; Pinilla, L.; Tena-Sempere, M. Developmental and Hormonally Regulated Messenger Ribonucleic Acid Expression of KiSS-1 and Its Putative Receptor, GPR54, in Rat Hypothalamus and Potent Luteinizing Hormone-Releasing Activity of KiSS-1 Peptide. Endocrinology 2004, 145, 4565–4574. [Google Scholar] [CrossRef] [PubMed]
- Dhillo, W.S.; Chaudhri, O.B.; Patterson, M.; Thompson, E.L.; Murphy, K.G.; Badman, M.K.; McGowan, B.M.; Amber, V.; Patel, S.; Ghatei, M.A.; et al. Kisspeptin-54 Stimulates the Hypothalamic-Pituitary Gonadal Axis in Human Males. J. Clin. Endocrinol. Metab. 2005, 90, 6609–6615. [Google Scholar] [CrossRef] [Green Version]
- Kadokawa, H.; Matsui, M.; Hayashi, K.; Matsunaga, N.; Kawashima, C.; Shimizu, T.; Kida, K.; Miyamoto, A. Peripheral administration of kisspeptin-10 increases plasma concentrations of GH as well as LH in prepubertal Holstein heifers. J. Endocrinol. 2008, 196, 331–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Messager, S.; Chatzidaki, E.E.; Ma, D.; Hendrick, A.G.; Zahn, D.; Dixon, J.; Thresher, R.R.; Malinge, I.; Lomet, D.; Carlton, M.B.L.; et al. Kisspeptin directly stimulates gonadotropin-releasing hormone release via G protein-coupled receptor 54. Proc. Natl. Acad. Sci. USA 2005, 102, 1761–1766. [Google Scholar] [CrossRef] [Green Version]
- Shahab, M.; Mastronardi, C.; Seminara, S.B.; Crowley, W.F.; Ojeda, S.R.; Plant, T.M. Increased hypothalamic GPR54 signaling: A potential mechanism for initiation of puberty in primates. Proc. Natl. Acad. Sci. USA 2005, 102, 2129–2134. [Google Scholar] [CrossRef] [Green Version]
- Lents, C.A.; Heidorn, N.L.; Barb, C.R.; Ford, J.J. Central and peripheral administration of kisspeptin activates gonadotropin but not somatotropin secretion in prepubertal gilts. Reproduction 2008, 135, 879–887. [Google Scholar] [CrossRef] [Green Version]
- Decourt, C.; Robert, V.; Lomet, D.; Anger, K.; Georgelin, M.; Poissenot, K.; Pellicer-Rubio, M.-T.; Aucagne, V.; Beltramo, M. The kisspeptin analog C6 is a possible alternative to PMSG (pregnant mare serum gonadotropin) for triggering synchronized and fertile ovulations in the Alpine goat. PLoS ONE 2019, 14, e0214424. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| KP | EBKP | EB12KP | p-Value | |
|---|---|---|---|---|
| AUC (ng/mL*min) | 176 ± 73 a | 241 ± 73 a | 439 ± 73 b | 0.02 |
| △LH (ng/mL) | 2.4 ± 0.9 a | 3.5 ± 0.9 a | 5.6 ± 0.9 b | <0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Macedo, G.G.; Batista, E.d.O.S.; Santos, G.M.G.d.; D’Occhio, M.J.; Baruselli, P.S. Estradiol Priming Potentiates the Kisspeptin-Induced Release of LH in Ovariectomized Cows. Animals 2021, 11, 1236. https://doi.org/10.3390/ani11051236
Macedo GG, Batista EdOS, Santos GMGd, D’Occhio MJ, Baruselli PS. Estradiol Priming Potentiates the Kisspeptin-Induced Release of LH in Ovariectomized Cows. Animals. 2021; 11(5):1236. https://doi.org/10.3390/ani11051236
Chicago/Turabian StyleMacedo, Gustavo Guerino, Emiliana de Oliveira Santana Batista, Gustavo Martins Gomes dos Santos, Michael J. D’Occhio, and Pietro Sampaio Baruselli. 2021. "Estradiol Priming Potentiates the Kisspeptin-Induced Release of LH in Ovariectomized Cows" Animals 11, no. 5: 1236. https://doi.org/10.3390/ani11051236