
Porcine Protein Hydrolysates (PEPTEIVA®) Promote Growth and Enhance Systemic Immunity in Gilthead Sea Bream (Sparus aurata)

 , , , , ,
, , , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Fish, Diets, and Sampling
2.2. Skin Mucus Collection, Biomarker Analyses, and Antibacterial Activity Measurement
2.3. Ex Vivo Immune Stimulation of Splenocytes with LPS and Gene Expression Analysis
2.4. Statistical Analyses
3. Results
3.1. Growth, Body Condition, and Feed Performance Indicators
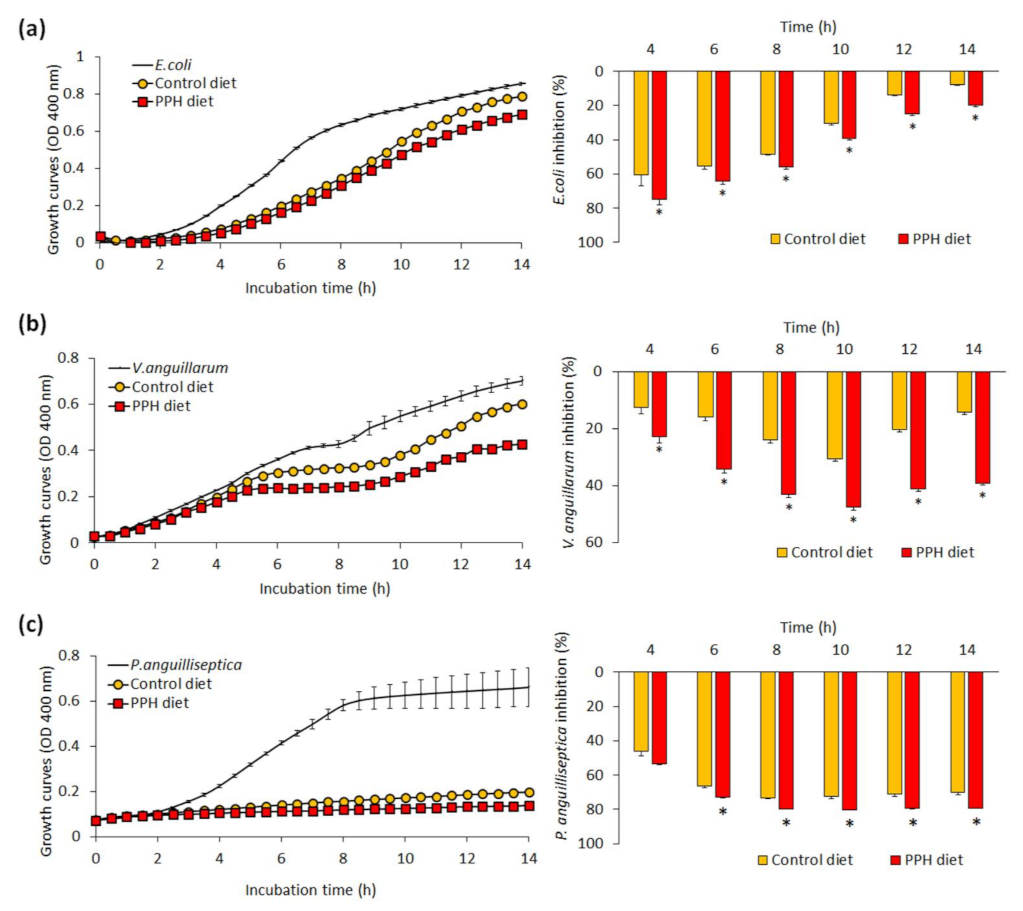
3.2. Skin Mucus Biomarkers and Bactericidal Activity
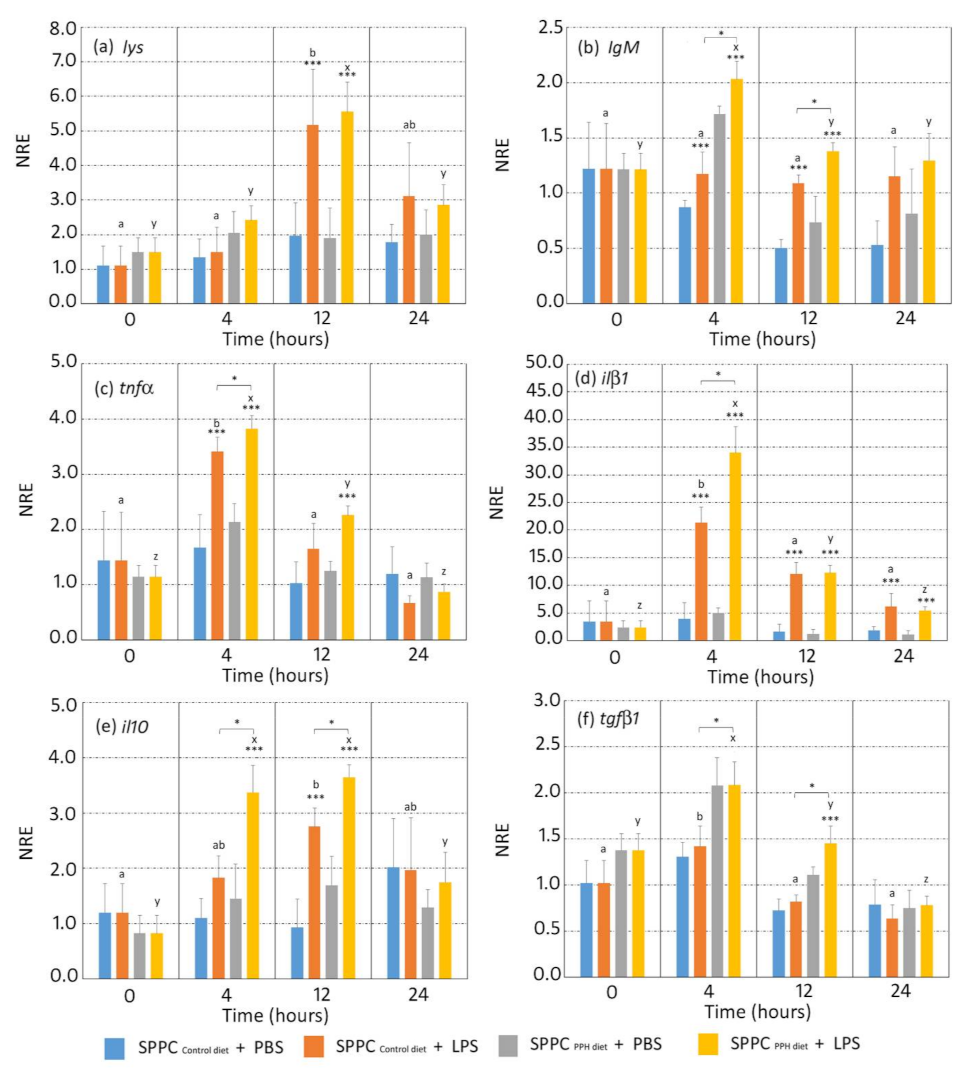
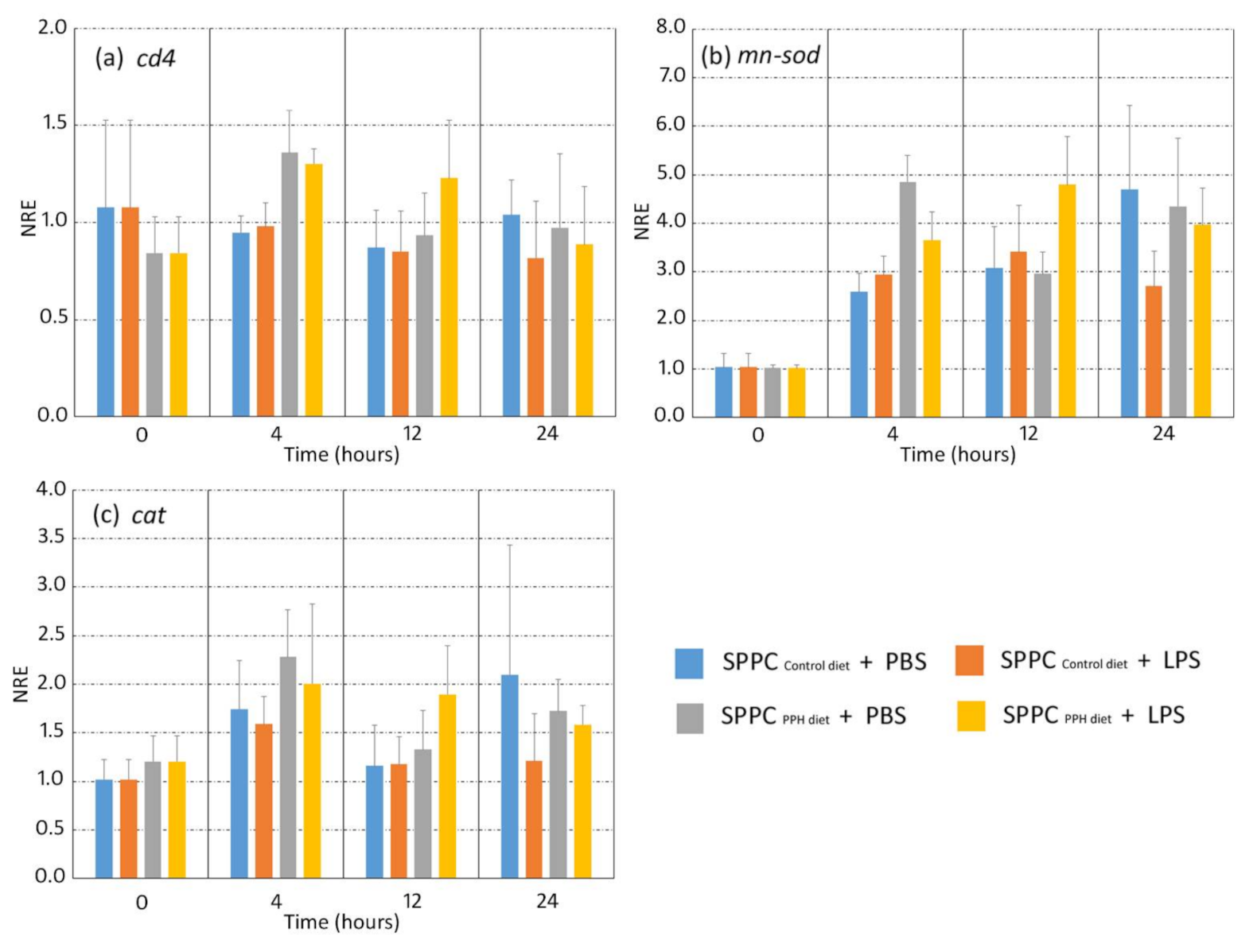
3.3. Gene Expression Analysis of Splenocytes Stimulated by LPS in an Ex Vivo Assay
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Naylor, R.L.; Hardy, R.W.; Buschmann, A.H.; Bush, S.R.; Cao, L.; Klinger, D.H.; Little, D.C.; Lubchenco, J.; Shumway, S.E.; Troell, M. A 20-year retrospective review of global aquaculture. Nat. Cell Biol. 2021, 591, 551–563. [Google Scholar] [CrossRef]
- Siddik, M.A.; Howieson, J.; Fotedar, R.; Partridge, G. Enzymatic fish protein hydrolysates in finfish aquaculture: A review. Rev. Aquac. 2021, 13, 406–430. [Google Scholar] [CrossRef]
- Stentiford, G.D.; Sritunyalucksana, K.; Flegel, T.W.; Williams, B.; Withyachumnarnkul, B.; Itsathitphaisarn, O.; Bass, D. New Paradigms to Help Solve the Global Aquaculture Disease Crisis. PLoS Pathog. 2017, 13, e1006160. [Google Scholar] [CrossRef]
- Lulijwa, R.; Rupia, E.J.; Alfaro, A.C. Antibiotic use in aquaculture, policies and regulation, health and environmental risks: A review of the top 15 major producers. Rev. Aquac. 2019, 12, 640–663. [Google Scholar] [CrossRef]
- Schar, D.; Klein, E.Y.; Laxminarayan, R.; Gilbert, M.; Van Boeckel, T.P. Global trends in antimicrobial use in aquaculture. Sci. Rep. 2020, 10, 1–9. [Google Scholar] [CrossRef]
- More, S.J. European perspectives on efforts to reduce antimicrobial usage in food animal production. Ir. Veter. J. 2020, 73, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Costello, C.; Cao, L.; Gelcich, S.; Cisneros-Mata, M.; Free, C.M.; Froehlich, H.E.; Golden, C.; Ishimura, G.; Maier, J.; Macadam-Somer, I.; et al. The future of food from the sea. Nat. Cell Biol. 2020, 588, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Dawood, M.; Koshio, S.; Esteban, M. Beneficial roles of feed additives as immunostimulants in aquaculture: A review. Rev. Aquac. 2017, 10, 950–974. [Google Scholar] [CrossRef]
- Piazzon, M.C.; Calduch-Giner, J.A.; Fouz, B.; Estensoro, I.; Simó-Mirabet, P.; Puyalto, M.; Karalazos, V.; Palenzuela, O.; Sitjà-Bobadilla, A.; Pérez-Sánchez, J. Under control: How a dietary additive can restore the gut microbiome and proteomic profile, and improve disease resilience in a marine teleostean fish fed vegetable diets. Microbiome 2017, 5, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Firmino, J.P.; Vallejos-Vidal, E.; Sarasquete, C.; Ortiz-Delgado, J.B.; Balasch, J.C.; Tort, L.; Estevez, A.; Reyes-López, F.E.; Gisbert, E. Unveiling the effect of dietary essential oils supplementation in Sparus aurata gills and its efficiency against the infestation by Sparicotyle chrysophrii. Sci. Rep. 2020, 10, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Firmino, J.P.; Fernández-Alacid, L.; Vallejos-Vidal, E.; Salomón, R.; Sanahuja, I.; Tort, L.; Antoni Ibarz, A.; Reyes-López, F.; Gisbert, E. Carvacrol, thymol, and garlic essential oil promote skin innate immunity in gilthead sea bream (Sparus aurata) through the multifactorial modulation of the secretory pathway and enhancement of mucus protective capacity. Front. Immunol. 2021, 12, 559. [Google Scholar] [CrossRef] [PubMed]
- Vallejos-Vidal, E.; Reyes-López, F.E.; Teles, M.; MacKenzie, S. The response of fish to immunostimulant diets. Fish Shellfish Immunol. 2016, 56, 34–69. [Google Scholar] [CrossRef]
- Kotzamanis, I.; Gisbert, E.; Gatesoupe, F.; Zambonino-Infante, J.L.; Cahu, C. Effects of different dietary levels of fish protein hydrolysates on growth, digestive enzymes, gut microbiota, and resistance to Vibrio anguillarum in European sea bass (Dicentrarchus labrax) larvae. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2007, 147, 205–214. [Google Scholar] [CrossRef] [Green Version]
- Robert, M.; Zatylny-Gaudin, C.; Fournier, V.; Corre, E.; Le Corguillé, G.; Bernay, B.; Henry, J. Molecular characterization of peptide fractions of a Tilapia (Oreochromis niloticus) by-product hydrolysate and in vitro evaluation of antibacterial activity. Process. Biochem. 2015, 50, 487–492. [Google Scholar] [CrossRef]
- Hou, Y.; Wu, Z.; Dai, Z.; Wang, G.; Wu, G. Protein hydrolysates in animal nutrition: Industrial production, bioactive peptides, and functional significance. J. Anim. Sci. Biotechnol. 2017, 8, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gisbert, E.; Fournier, V.; Solovyev, M.; Skalli, A.; Andree, K.B. Diets containing shrimp protein hydrolysates provided protection to European sea bass (Dicentrarchus labrax) affected by a Vibrio pelagius natural infection outbreak. Aquaculture 2018, 495, 136–143. [Google Scholar] [CrossRef]
- Gisbert, E.; Skalli, A.; Fernández, I.; Kotzamanis, Y.; Zambonino-Infante, J.L.; Fabregat, R. Protein hydrolysates from yeast and pig blood as alternative raw materials in microdiets for gilthead sea bream (Sparus aurata) larvae. Aquaculture 2012, 338, 96–104. [Google Scholar] [CrossRef] [Green Version]
- Breck, O.; Bjerkaas, P.; Campbell, P.; Arnesen, P.; Haldorsen, P.; Waagbø, R. Cataract preventive role of mammalian blood meal, histidine, iron and zinc in diets for Atlantic salmon (Salmo salar L.) of different strains. Aquac. Nutr. 2003, 9, 341–350. [Google Scholar] [CrossRef]
- Gisbert, E.; Skalli, A.; Campbell, J.; Solovyev, M.M.; Rodríguez, C.; Dias, J.; Polo, J. Spray-dried plasma promotes growth, modulates the activity of antioxidant defenses, and enhances the immune status of gilthead sea bream (Sparus aurata) fingerlings1. J. Anim. Sci. 2015, 93, 278–286. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.A.; Summerfelt, R.C. Spray-dried blood cells as a partial replacement for fishmeal in diets for rainbow trout Oncorhynchus mykiss. J. World Aquac. Soc. 2000, 31, 96–104. [Google Scholar] [CrossRef]
- De Araújo, E.P.; de Carvalho, P.L.P.F.; De Freitas, J.M.A.; Da Silva, R.L.; Rocha, M.K.H.R.; Teixeira, C.P.; Damasceno, F.M.; Sartori, M.M.P.; Pezzato, L.E.; Barros, M.M. Dietary spray-dried plasma enhances the growth performance, villus:crypt ratio and cold-induced stress resistance in Nile tilapia (Oreochromis niloticus). Aquaculture 2017, 479, 675–681. [Google Scholar] [CrossRef] [Green Version]
- Reyes-López, F.E.; Ibarz, A.; Ordóñez-Grande, B.; Vallejos-Vidal, E.; Andree, K.B.; Balasch, J.C.; Fernández-Alacid, L.; Sanahuja, I.; Sánchez-Nuño, S.; Firmino, J.P.; et al. Skin Multi-Omics-Based Interactome Analysis: Integrating the Tissue and Mucus Exuded Layer for a Comprehensive Understanding of the Teleost Mucosa Functionality as Model of Study. Front. Immunol. 2021, 11. [Google Scholar] [CrossRef]
- Bureau, D.P. Rendered Products in Fish Aquaculture Feeds. In Essential Rendering—All about the Animal by Products Industry; Meeker, D.L., Ed.; National Renderers Association: Alexandria, VR, USA, 2006; pp. 179–194. [Google Scholar]
- FAO. The State of World Fisheries and Aquaculture. In FishStatJ, A Tool for Fishery Statistics Analysis; Release: 4.01.0. Fao Fisheries and Aquaculture Department; The Food and Agriculture Organization of the United Nations: Rome, Italy, 2020. [Google Scholar]
- Fernández-Alacid, L.; Sanahuja, I.; Grande, B.O.; Sánchez, S.; Viscor, G.; Gisbert, E.; Herrera, M.; Ibarz, A. Skin mucus metabolites in response to physiological challenges: A valuable non-invasive method to study teleost marine species. Sci. Total Environ. 2018, 644, 1323–1335. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Fernández-Alacid, L.; Sanahuja, I.; Grande, B.O.; Sánchez-Nuño, S.; Herrera, M.; Ibarz, A. Skin mucus metabolites and cortisol in meagre fed acute stress-attenuating diets: Correlations between plasma and mucus. Aquaculture 2019, 499, 185–194. [Google Scholar] [CrossRef]
- Sanahuja, I.; Fernández-Alacid, L.; Grande, B.O.; Sánchez, S.; Ramos, A.; Araujo, R.M.; Ibarz, A. Comparison of several non-specific skin mucus immune defences in three piscine species of aquaculture interest. Fish Shellfish Immunol. 2019, 89, 428–436. [Google Scholar] [CrossRef] [PubMed]
- Salomón, R.; Firmino, J.P.; Reyes-López, F.E.; Andree, K.B.; González-Silvera, D.; Esteban, M.A.; Tort, L.; Quintela, J.C.; Pinilla-Rosas, J.M.; Vallejos-Vidal, E.; et al. The growth promoting and immunomodulatory effects of a medicinal plant leaf extract obtained from Salvia officinalis and Lippia citriodora in gilthead sea bream (Sparus aurata). Aquaculture 2020, 524, 735291. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Stevens, J.R.; Newton, R.W.; Tlusty, M.; Little, D.C. The rise of aquaculture by-products: Increasing food production, value, and sustainability through strategic utilisation. Mar. Policy 2018, 90, 115–124. [Google Scholar] [CrossRef] [Green Version]
- Nogueira, N.; Cordeiro, N.; Andrade, C.; Aires, T. Inclusion of low levels of blood and feather meal in practical diets for gilthead sea bream (Sparus aurata). Turk. J. Fish. Aquat. Sci. 2012, 12, 641–650. [Google Scholar] [CrossRef]
- European Commission. Commission Regulation (EC) No 1292/2005 of 5 August 2005 amending Annex IV to Regulation (EC) No 999/2001 of the European Parliament and of the Council as regards animal nutrition. Off. J. Eur. Union 2005, 48, 3–11. [Google Scholar]
- European Commission. Commission Regulation (EU) No 56/2013 of 16 January 2013 amending Annexes I and IV to Regulation (EC) No 999/2001 of the European Parliament and of the Council laying down rules for the prevention, control and eradication of certain transmissible spongiform encephalopathies. Off. J. Eur. Union 2013, 56, 3–16. [Google Scholar]
- van Dijk, A.; Everts, H.; Nabuurs, M.; Margry, R.; Beynen, A. Growth performance of weanling pigs fed spray-dried animal plasma: A review. Livest. Prod. Sci. 2001, 68, 263–274. [Google Scholar] [CrossRef]
- Booth, A.M.; Allan, G.L.; Anderson, A.J. Investigation of the nutritional requirements of Australian snapper Pagrus auratus (Bloch & Schneider, 1801): Apparent digestibility of protein and energy sources. Aquac. Res. 2005, 36, 378–390. [Google Scholar] [CrossRef]
- Sun, Z.; Ma, Q.; Li, Z.; Ji, C. Effect of Partial Substitution of Dietary Spray-dried Porcine Plasma or Fishmeal with Soybean and Shrimp Protein Hydrolysate on Growth Performance, Nutrient Digestibility and Serum Biochemical Parameters of Weanling Piglets. Asian Australas. J. Anim. Sci. 2009, 22, 1032–1037. [Google Scholar] [CrossRef]
- Campbell, J.; Polo, J.; Russell, L.; Crenshaw, J. Review of spray-dried plasma’s impact on intestinal barrier function. Livest. Sci. 2010, 133, 239–241. [Google Scholar] [CrossRef]
- Balan, P.; Staincliffe, M.; Moughan, P.J. Effects of spray-dried animal plasma on the growth performance of weaned piglets—A review. J. Anim. Physiol. Anim. Nutr. 2020. [Google Scholar] [CrossRef]
- Parra, D.; Reyes-López, F.E.; Tort, L. Mucosal Immunity and B Cells in Teleosts: Effect of Vaccination and Stress. Front. Immunol. 2015, 6, 354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuesta, A.; Meseguer, J.; Esteban, M.A. Total serum immunoglobulin M levels are affected by immunomodulators in sea bream (Sparus aurata L.). Vet. Immunol. Immunopathol. 2004, 3, 203–210. [Google Scholar] [CrossRef]
- Reyes-Cerpa, S.; Vallejos-Vidal, E.; José Gonzalez-Bown, M.; Morales-Reyes, J.; Pérez-Stuardo, D.; Vargas, D.; Imarai, M.; Cifuentes, V.; Spencer, E.; María Sandino, A.; et al. Effect of yeast (Xanthophyllomyces dendrorhous) and plant (Saint John’s wort, lemon balm, and rosemary) extract based functional diets on antioxidant and immune status of Atlantic salmon (Salmo salar) subjected to crowding stress. Fish Shellfish Immunol. 2018, 74, 250–259. [Google Scholar] [CrossRef]
- Hayden, M.S.; Ghosh, S. Regulation of NF-κB by TNF family cytokines. Semin. Immunol. 2014, 26, 253–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, J.; Secombes, C.J. The Function of Fish Cytokines. Biology 2016, 5, 23. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Hayden, M.S. New regulators of NF-κB in inflammation. Nat. Rev. Immunol. 2008, 8, 837–848. [Google Scholar] [CrossRef]
- Uribe, C.; Folch, H.; Enriquez, R.; Moran, G. Innate and adaptive immunity in teleost fish: A review. Vet. Med. 2011, 56, 486–503. [Google Scholar] [CrossRef] [Green Version]
- Luckheeram, R.V.; Zhou, R.; Verma, A.D.; Xia, B. CD4+ T cells: Differentiation and functions. Clin. Dev. Immunol. 2012, 2012, 925135. [Google Scholar] [CrossRef] [Green Version]
- Cummings, J.H.; Antoine, J.-M.; Azpiroz, F.; Bourdet-Sicard, R.; Brandtzaeg, P.; Calder, P.; Gibson, G.R.; Guarner, F.; Isolauri, E.; Pannemans, D.; et al. PASSCLAIM1? Gut health and immunity. Eur. J. Nutr. 2004, 43, ii118–ii173. [Google Scholar] [CrossRef]
- Moore, K.W.; de Waal Malefyt, R.; Coffman, R.L.; O’Garra, A. Interleukin −10 and the interleukin −10 receptor. Ann. Rev. Immunol. 2001, 19, 683–765. [Google Scholar] [CrossRef]
- Gao, Y.Y.; Jiang, Z.Y.; Lin, Y.C.; Zheng, C.T.; Zhou, G.L.; Chen, F. Effects of spray-dried animal plasma on serous and intestinal redox status and cytokines of neonatal piglets1. J. Anim. Sci. 2011, 89, 150–157. [Google Scholar] [CrossRef]
- Pérez-Bosque, A.; Miró, L.; Amat, C.; Polo, J.; Moretó, M. The Anti-Inflammatory Effect of Spray-Dried Plasma Is Mediated by a Reduction in Mucosal Lymphocyte Activation and Infiltration in a Mouse Model of Intestinal Inflammation. Nutrition 2016, 8, 657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Bosque, A.; Miró, L.; Polo, J.; Russell, L.; Campbell, J.; Weaver, E.; Crenshaw, J.; Moretó, M. Dietary Plasma Protein Supplements Prevent the Release of Mucosal Proinflammatory Mediators in Intestinal Inflammation in Rats. J. Nutr. 2009, 140, 25–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández-Chirlaque, C.; Aranda, C.; Ocón, B.; Polo, J.; Martínez-Augustin, O.; de Medina, F.S. Immunoregulatory Effects of Porcine Plasma Protein Concentrates on Rat Intestinal Epithelial Cells and Splenocytes. Animals 2021, 11, 807. [Google Scholar] [CrossRef]
- Chowdhury, S.; Saikia, S.K. Oxidative Stress in Fish: A Review. J. Sci. Res. 2020, 12, 145–160. [Google Scholar] [CrossRef] [Green Version]
- Guardiola, F.A.; Cuesta, A.; Abellán, E.; Meseguer, J.; Esteban, M.A. Comparative analysis of the humoral immunity of skin mucus from several marine teleost fish. Fish Shellfish Immunol. 2014, 40, 24–31. [Google Scholar] [CrossRef]
- Fernández-Alacid, L.; Sanahuja, I.; Grande, B.O.; Sánchez, S.; Herrera, M.; Ibarz, A. Comparison between properties of dorsal and ventral skin mucus in Senegalese sole: Response to an acute stress. Aquaculture 2019, 513, 734410. [Google Scholar] [CrossRef]
- de Mercado, E.; Larrán, A.M.; Pinedo, J.; Tomás-Almenar, C. Skin mucous: A new approach to assess stress in rainbow trout. Aquaculture 2018, 484, 90–97. [Google Scholar] [CrossRef]
- Herrera, M.; Fernández-Alacid, L.; Sanahuja, I.; Ibarz, A.; Salamanca, N.; Morales, E.; Giráldez, I. Physiological and metabolic effects of a tryptophan-enriched diet to face up chronic stress in meagre (Argyrosomus regius). Aquaclture 2020, 522, 735102. [Google Scholar] [CrossRef]
- Muniesa, A.; Basurco, B.; Aguilera, C.; Furones, D.; Reverté, C.; Sanjuan-Vilaplana, A.; Jansen, M.D.; Brun, E. Tavornpanich, S. Mapping the knowledge of the main diseases affecting sea bass and sea bream in Mediterranean. Transbound. Emerg. Diseases 2020, 67, 1089–1100. [Google Scholar] [CrossRef]
- Moretó, M.; Pérez-Bosque, A. Dietary plasma proteins, the intestinal immune system, and the barrier functions of the intestinal mucosa1. J. Anim. Sci. 2009, 87, E92–E100. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.M.; Crenshaw, J.D.; González-Esquerra, R.; Polo, J. Impact of Spray-Dried Plasma on Intestinal Health and Broiler Performance. Microorganisms 2019, 7, 219. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Ingredients (%) | Control Diet | PPH Diet |
|---|---|---|
| Fishmeal LT70 (NORVIK) | 7.0 | 2.0 |
| PPH (PEPTEIVA®, APC Europe) | - | 5.0 |
| Soy protein concentrate (Soycomil) | 21.0 | 21.0 |
| Pea protein concentrate | 12.0 | 12.0 |
| Wheat gluten | 12.0 | 12.0 |
| Corn gluten | 12.0 | 12.0 |
| Soybean meal 48 | 5.0 | 5.0 |
| Wheat meal | 10.4 | 10.4 |
| Fish oil (SAVINOR) | 15.0 | 15.0 |
| Vitamin and Mineral Premix PV01 | 1.0 | 1.0 |
| Soy lecithin—Powder | 1.0 | 1.0 |
| Binder (guar gum) | 1.0 | 1.0 |
| MCP | 2.0 | 2.0 |
| L-Lysine | 0.3 | 0.3 |
| L-Tryptophan | 0.1 | 0.1 |
| DL-Methionine | 0.2 | 0.2 |
| Proximate composition | ||
| Crude protein (%) | 48.37 ± 0.2 | 48.50 ± 0.3 |
| Crude fat (%) | 17.19 ± 0.2 | 17.11 ± 0.1 |
| Ash (%) | 5.88 ± 0.05 | 5.81 ± 0.07 |
| Gross energy (MJ kg feed−1) | 21.62 ± 0.4 | 21.77 ± 0.5 |
| Control Diet | PPH Diet | |
|---|---|---|
| Survival (%) | 96.8 ± 2.7 | 97.6 ± 1.6 |
| SLi (cm) | 10.2 ± 0.08 | 10.3 ± 0.10 |
| SLf (cm) | 18.7 ± 0.31 | 19.0 ± 0.36 |
| BWi (g) | 26.0 ± 0.03 | 25.9 ± 0.11 |
| BWf (g) | 173.8 ± 4.14 * | 182.2 ± 4.37 |
| SGRBW (% BW day−1) | 2.06 ± 0.03 * | 2.12 ± 0.02 |
| K | 2.64 ± 0.07 | 2.67 ± 0.04 |
| FCR | 0.87 ± 0.06 | 0.93 ± 0.11 |
| FI (g feed fish−1) | 115.1 ± 4.5 * | 130.8 ± 8.7 |
| Control Diet | PPH Diet | |
|---|---|---|
| Mucus biomarkers | ||
| Glucose (µg mL−1) | 14.98 ± 5.45 | 16.36 ± 5.61 |
| Lactate (µg mL−1) | 7.24 ± 3.01 | 8.57 ± 2.37 |
| Protein (mg mL−1) | 9.01 ± 3.41 | 11.76 ± 3.46 |
| Cortisol (µg mL−1) | 0.40 ± 0.45 | 0.06 ± 0.02 |
| FRAP (µmol mL−1) | 1387 ± 277 | 1610 ± 294 |
| Mucus ratios | ||
| Glucose/protein (µg mg−1) | 1.83 ± 0.53 | 1.66 ± 0.49 |
| Lactate/protein (µg mg−1) | 0.82 ± 0.17 | 0.76 ± 0.18 |
| Cortisol/protein (ng g−1) | 53.4 ± 57.6 | 7.2 ± 3.3 |
| FRAP/protein (µmol mg−1) | 146 ± 20 | 146 ± 41 |
| Glucose/Lactate | 2.41 ± 0.44 | 2.02 ± 0.46 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gisbert, E.; Ibarz, A.; Firmino, J.P.; Fernández-Alacid, L.; Salomón, R.; Vallejos-Vidal, E.; Ruiz, A.; Polo, J.; Sanahuja, I.; Reyes-López, F.E.; et al. Porcine Protein Hydrolysates (PEPTEIVA®) Promote Growth and Enhance Systemic Immunity in Gilthead Sea Bream (Sparus aurata). Animals 2021, 11, 2122. https://doi.org/10.3390/ani11072122
Gisbert E, Ibarz A, Firmino JP, Fernández-Alacid L, Salomón R, Vallejos-Vidal E, Ruiz A, Polo J, Sanahuja I, Reyes-López FE, et al. Porcine Protein Hydrolysates (PEPTEIVA®) Promote Growth and Enhance Systemic Immunity in Gilthead Sea Bream (Sparus aurata). Animals. 2021; 11(7):2122. https://doi.org/10.3390/ani11072122
Chicago/Turabian StyleGisbert, Enric, Antoni Ibarz, Joana P. Firmino, Laura Fernández-Alacid, Ricardo Salomón, Eva Vallejos-Vidal, Alberto Ruiz, Javier Polo, Ignasi Sanahuja, Felipe E. Reyes-López, and et al. 2021. "Porcine Protein Hydrolysates (PEPTEIVA®) Promote Growth and Enhance Systemic Immunity in Gilthead Sea Bream (Sparus aurata)" Animals 11, no. 7: 2122. https://doi.org/10.3390/ani11072122