
Effects of Dietary Supplementation of Lactobacillus acidophilus on Blood Parameters and Gut Health of Rabbits

 , , , , , , , ,
, , , , , , , ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Diets
2.2. Productive Parameters
2.3. Blood Analysis
2.4. Histomorphometric Investigations
2.5. Cecal pH and Microbiota
2.6. Cecal Short-Chain Fatty Acids
2.7. Statistical Analysis
3. Results
3.1. Productive Parameters
3.2. Biochemical Parameters
3.3. Histomorphometric Investigations
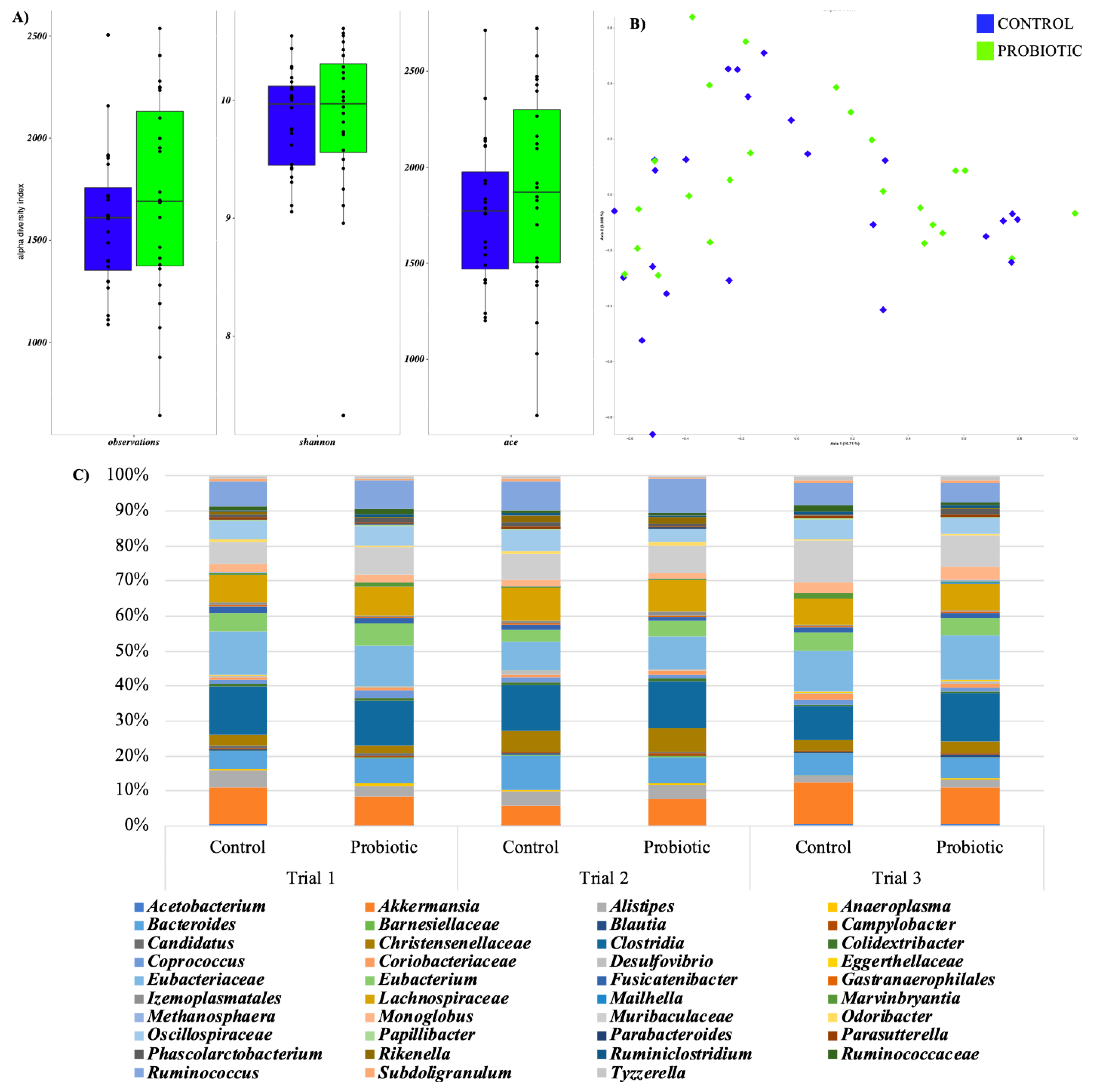
3.4. Caecal pH and Microbiota
3.5. Caecal Short-Chain Fatty Acids
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kohles, M. Gastrointestinal Anatomy and Physiology of Select Exotic Companion Mammals. Vet. Clin. N. Am.-Exot. Anim. Pract. 2014, 17, 165–178. [Google Scholar] [CrossRef] [PubMed]
- Bäuerl, C.; Collado, M.C.; Zúñiga, M.; Blas, E.; Martínez, G.P. Changes in Cecal Microbiota and Mucosal Gene Expression Revealed New Aspects of Epizootic Rabbit Enteropathy. PLoS ONE 2014, 9, e105707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatt, R.S.; Agrawal, A.R.; Sahoo, A. Effect of Probiotic Supplementation on Growth Performance, Nutrient Utilization and Carcass Characteristics of Growing Chinchilla Rabbits. J. Appl. Anim. Res. 2017, 45, 304–309. [Google Scholar] [CrossRef] [Green Version]
- Kylie, J.; Weese, J.S.; Turner, P.V. Comparison of the Fecal Microbiota of Domestic Commercial Meat, Laboratory, Companion, and Shelter Rabbits (Oryctolagus Cuniculi). BMC Vet. Res. 2018, 14, 143. [Google Scholar] [CrossRef]
- Bischoff, S.C. “Gut Health”: A New Objective in Medicine? BMC Med. 2011, 9, 24. [Google Scholar] [CrossRef]
- Bai, X.; Shi, Y.; Tang, L.; Chen, L.; Fan, H.; Wang, H.; Wang, J.; Jia, X.; Chen, S.; Lai, S. Heat Stress Affects Faecal Microbial and Metabolic Alterations of Rabbits. Front. Microbiol. 2022, 12, 817615. [Google Scholar] [CrossRef]
- Al-Shawi, S.G.; Dang, D.S.; Yousif, A.Y.; Al-Younis, Z.K.; Najm, T.A.; Matarneh, S.K. The Potential Use of Probiotics to Improve Animal Health, Efficiency, and Meat Quality: A Review. Agriculture 2020, 10, 452. [Google Scholar] [CrossRef]
- Chen, H.-J.; Yang, W.-Y.; Wang, C.-Y. The Review on the Function of Intestinal Flora and the Regulatory Effects of Probiotics on the Intestinal Health of Rabbits. In Proceedings of the 2017 2nd International Conference on Biological Sciences and Technology (BST 2017), Zhuhai, China, 17–19 November 2017; Atlantis Press: Amsterdam, The Netherlands, 2018; Volume 6, pp. 49–55. [Google Scholar] [CrossRef] [Green Version]
- Bull, M.; Plummer, S.; Marchesi, J.; Mahenthiralingam, E. The Life History of Lactobacillus Acidophilus as a Probiotic: A Tale of Revisionary Taxonomy, Misidentification and Commercial Success. FEMS Microbiol. Lett. 2013, 349, 77–87. [Google Scholar] [CrossRef] [Green Version]
- Shehata, A.A.; Yalçın, S.; Latorre, J.D.; Basiouni, S.; Attia, Y.A.; El-Wahab, A.A.; Visscher, C.; El-Seedi, H.R.; Huber, C.; Hafez, H.M.; et al. Probiotics, Prebiotics, and Phytogenic Substances for Optimizing Gut Health in Poultry. Microorganisms 2022, 10, 395. [Google Scholar] [CrossRef]
- Gadde, U.; Kim, W.H.; Oh, S.T.; Lillehoj, H.S. Alternatives to Antibiotics for Maximizing Growth Performance and Feed Efficiency in Poultry: A Review. Anim. Health Res. Rev. 2017, 18, 26–45. [Google Scholar] [CrossRef] [Green Version]
- Amber, K.; Yakout, H.; Hamed Rawya, S. Feeding Diets Containing Yucca Extract or Probiotic on Growth, Digestibility, Nitrogen Balance and Caecal Microbial Activity of Growing New Zealand Rabbits. In Proceedings of the 8th World Rabbit Congress, Puebla, Mexico, 7–10 September 2004; pp. 737–745. [Google Scholar]
- Abdelhady, D.; El-Abasy, M. Effect of Prebiotic and Probiotic on Growth, Immuno-Hematological Responses and Biochemical Parameters of Infected Rabbits with Pasteurella Multocida. Benha Vet. Med. J. 2015, 28, 40–51. [Google Scholar] [CrossRef] [Green Version]
- Simonová, M.P.; Lauková, A.; Žitňan, R.; Chrastinová, Ľ. Effect of Rabbit-Origin Enterocin-Producing Probiotic Strain Enterococcus Faecium CCM7420 Application on Growth Performance and Gut Morphometry in Rabbits. Czech J. Anim. Sci. 2015, 60, 509–512. [Google Scholar] [CrossRef] [Green Version]
- Oso, A.O.; Idowu, O.M.O.; Haastrup, A.S.; Ajibade, A.J.; Olowonefa, K.O.; Aluko, A.O.; Ogunade, I.M.; Osho, S.O.; Bamgbose, A.M. Growth Performance, Apparent Nutrient Digestibility, Caecal Fermentation, Ileal Morphology and Caecal Microflora of Growing Rabbits Fed Diets Containing Probiotics and Prebiotics. Livest. Sci. 2013, 157, 184–190. [Google Scholar] [CrossRef]
- Kadja, L.; Dib, A.L.; Lakhdara, N.; Bouaziz, A.; Espigares, E.; Gagaoua, M. Influence of Three Probiotics Strains, Lactobacillus Rhamnosus Gg, Bifidobacterium Animalis Subsp. Lactis Bb-12 and Saccharomyces Boulardii Cncm i-745 on the Biochemical and Haematological Profiles and Body Weight of Healthy Rabbits. Biology 2021, 10, 1194. [Google Scholar] [CrossRef]
- Soltan, M.; El-Katcha, M. Effect of Dietary Probiotics Supplementation on Growth Performance, Immune Response, Some Blood Parameters and Carcass Quality of Growing Rabbits. Alex. J. Vet. Sci. 2011, 34, 153–169. [Google Scholar]
- Attia, Y.A.; el Hamid, A.E.; Ismaiel, A.M.; de Oliveira, M.C.; Al-Harthi, M.A.; El-Naggar, A.S.; Simon, G.A. Nitrate Detoxification Using Antioxidants and Probiotics in the Water for Rabbits. Rev. Colomb. De Cienc. Pecu. 2018, 31, 130–138. [Google Scholar] [CrossRef]
- Nwachukwu, C.U.; Aliyu, K.I.; Ewuola, E.O. Growth Indices, Intestinal Histomorphology, and Blood Profile of Rabbits Fed Probiotics- and Prebiotics-Supplemented Diets. Transl. Anim. Sci. 2021, 5, txab096. [Google Scholar] [CrossRef]
- Shen, X.M.; Cui, H.X.; Xu, X.R. Orally Administered Lactobacillus Casei Exhibited Several Probiotic Properties in Artificially Suckling Rabbits. Asian-Australas. J. Anim. Sci. 2020, 33, 1352–1359. [Google Scholar] [CrossRef] [Green Version]
- Phuoc, T.L.; Jamikorn, U. Effects of Probiotic Supplement (Bacillus Subtilis and Lactobacillus Acidophilus) on Feed Efficiency, Growth Performance, and Microbial Population of Weaning Rabbits. Asian-Australas. J. Anim. Sci. 2017, 30, 198–205. [Google Scholar] [CrossRef] [Green Version]
- Abd El-Hamid, M.I.; Ibrahim, D.; Hamed, R.I.; Nossieur, H.H.; Elbanna, M.H.; Baz, H.; Abd-Allah, E.M.; el Oksh, A.S.A.; Ibrahim, G.A.; Khalifa, E.; et al. Modulatory Impacts of Multi-Strain Probiotics on Rabbits’ Growth, Nutrient Transporters, Tight Junctions and Immune System to Fight against Listeria Monocytogenes Infection. Animal 2022, 12, 2082. [Google Scholar] [CrossRef]
- de Cesare, A.; Sala, C.; Castellani, G.; Astolfi, A.; Indio, V.; Giardini, A.; Manfreda, G. Effect of Lactobacillus Acidophilus D2/CSL (CECT 4529) Supplementation in Drinking Water on Chicken Crop and Caeca Microbiome. PLoS ONE 2020, 15, e0228338. [Google Scholar] [CrossRef] [PubMed]
- de Cesare, A.; Sirri, F.; Manfreda, G.; Moniaci, P.; Giardini, A.; Zampiga, M.; Meluzzi, A. Effect of Dietary Supplementation with Lactobacillus Acidophilus D2/CSL (CECT 4529) on Caecum Microbioma and Productive Performance in Broiler Chickens. PLoS ONE 2017, 12, e0176309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forte, C.; Acuti, G.; Manuali, E.; Casagrande Proietti, P.; Pavone, S.; Trabalza-Marinucci, M.; Moscati, L.; Onofri, A.; Lorenzetti, C.; Franciosini, M.P. Effects of Two Different Probiotics on Microflora, Morphology, and Morphometry of Gut in Organic Laying Hens. Poult. Sci. 2016, 95, 2528–2535. [Google Scholar] [CrossRef] [PubMed]
- Cesari, V.; Mangiagalli, M.G.; Giardini, A.; Galimberti, P.; Carteri, S.; Gallazzi, D.; Toschi, I. Egg Quality and Productive Performance of Laying Hens Fed Different Levels of Skimmed Milk Powder Added to a Diet Containing Lactobacillus Acidophilus. Poult. Sci. 2014, 93, 1197–1201. [Google Scholar] [CrossRef] [PubMed]
- Gallazzi, D.; Giardini, A.; Mangiagalli, M.G.; Marelli, S.; Ferrazzi, V.; Orsi, C.; Cavalchini, L.G. Effects of Lactobacillus Acidophilus D2/CSL on Laying Hen Performance. Ital. J. Anim. Sci. 2008, 7, 27–37. [Google Scholar] [CrossRef] [Green Version]
- Forte, C.; Moscati, L.; Acuti, G.; Mugnai, C.; Franciosini, M.P.; Costarelli, S.; Cobellis, G.; Trabalza-Marinucci, M. Effects of Dietary Lactobacillus Acidophilus and Bacillus Subtilis on Laying Performance, Egg Quality, Blood Biochemistry and Immune Response of Organic Laying Hens. J. Anim. Physiol. Anim. Nutr. 2016, 100, 977–987. [Google Scholar] [CrossRef]
- Marelli, S.P.; Fusi, E.; Giardini, A.; Martino, P.A.; Polli, M.; Bruni, N.; Rizzi, R. Effects of Probiotic Lactobacillus Acidophilus D2/CSL (CECT 4529) on the Nutritional and Health Status of Boxer Dogs. Vet. Rec. 2020, 187, e28. [Google Scholar] [CrossRef] [Green Version]
- Fusi, E.; Rizzi, R.; Polli, M.; Cannas, S.; Giardini, A.; Bruni, N.; Marelli, S.P. Effects of Lactobacillus Acidophilus D2/CSL (CECT 4529) Supplementation on Healthy Cat Performance. Vet. Rec. Open 2019, 6, e000368. [Google Scholar] [CrossRef] [Green Version]
- Authority, E.F.S. Guidance on the Assessment of the Efficacy of Feed Additives. EFSA J. 2018, 16, 5274. [Google Scholar] [CrossRef]
- Hegab, I.; Eman, A.; Rania, A.; El-Azzazi, F. Effect of Probiotics on Productive, Physiological and Microbiological Parameters of New Zealand White Rabbits Reared Under Hot Summer Conditions. Egypt. Poult. Sci. J. 2019, 39, 599–614. [Google Scholar] [CrossRef]
- de Blas, C.; Wiseman, J. Nutrition of the Rabbit, 2nd ed.; CABI Publishing: Wallingford, UK, 2010; ISBN 9781845936693. [Google Scholar]
- Pet Food Manufacturers Association. Rabbit Size-O-Meter; Pet Food Manufacturers Association: London, UK, 2022. [Google Scholar]
- Weaver, L.A.; Blaze, C.A.; Linder, D.E.; Andrutis, K.A.; Karas, A.Z. A Model for Clinical Evaluation of Perioperative Analgesia in Rabbits (Oryctolagus Cuniculus). J. Am. Assoc. Lab. Anim. Sci. 2010, 49, 845–851. [Google Scholar] [PubMed]
- Kaneko, J.J.; Harvey, J.W.; Bruss, M.L. Clinical Biochemistry of Domestic Animals, 6th ed.; Academic Press: Cambridge, MA, USA, 2008. [Google Scholar]
- Gasco, L.; Dabbou, S.; Trocino, A.; Xiccato, G.; Capucchio, M.T.; Biasato, I.; Dezzutto, D.; Birolo, M.; Meneguz, M.; Schiavone, A.; et al. Effect of Dietary Supplementation with Insect Fats on Growth Performance, Digestive Efficiency and Health of Rabbits. J. Anim. Sci. Biotechnol. 2019, 10, 4. [Google Scholar] [CrossRef] [PubMed]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of General 16S Ribosomal RNA Gene PCR Primers for Classical and Next-Generation Sequencing-Based Diversity Studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, Interactive, Scalable and Extensible Microbiome Data Science Using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Guantario, B.; Giribaldi, M.; Devirgiliis, C.; Finamore, A.; Colombino, E.; Capucchio, M.T.; Evangelista, R.; Motta, V.; Zinno, P.; Cirrincione, S.; et al. A Comprehensive Evaluation of the Impact of Bovine Milk Containing Different Beta-Casein Profiles on Gut Health of Ageing Mice. Nutrients 2020, 12, 2147. [Google Scholar] [CrossRef]
- Abdel-Aziz, N.A.; El-Adawy, M.; Mariezcurrena-Berasain, M.A.; Salem, A.Z.M.; Olivares-Pérez, J.; Kholif, A.E.; Borhami, B.E. Effects of Exogenous Enzymes, Lactobacillus Acidophilus or Their Combination on Feed Performance Response and Carcass Characteristics of Rabbits Fed Sugarcane Bagasse. J. Integr. Agric. 2015, 14, 544–549. [Google Scholar] [CrossRef]
- Ebeid, T.A.; Zeweil, H.S.; Basyony, M.M.; Dosoky, W.M.; Badry, H. Fortification of Rabbit Diets with Vitamin E or Selenium Affects Growth Performance, Lipid Peroxidation, Oxidative Status and Immune Response in Growing Rabbits. Livest. Sci. 2013, 155, 323–331. [Google Scholar] [CrossRef]
- Cardinali, R.; Dal Bosco, A.; Bonanno, A.; di Grigoli, A.; Rebollar, P.G.; Lorenzo, P.L.; Castellini, C. Connection between Body Condition Score, Chemical Characteristics of Body and Reproductive Traits of Rabbit Does. Livest. Sci. 2008, 116, 209–215. [Google Scholar] [CrossRef]
- Abdel-Azeem, A.; Hassan, A.; Basyony, M.; Abu Hafsa, S. Rabbit Growth, Carcass Characteristic, Digestion, Caecal Fermentation, Microflora, and Some Blood Biochemical Components Affected By Oral Administration of Anaerobic Probiotic (Zad®). Egypt. J. Nutr. Feed. 2018, 21, 693–710. [Google Scholar] [CrossRef]
- Melillo, A. Applications of Serum Protein Electrophoresis in Exotic Pet Medicine. Vet.Clin. N. Am. Exot. Anim. Pract. 2013, 16, 211–225. [Google Scholar] [CrossRef] [PubMed]
- El-Adawy, M.M.; Abou-Zeid, A.E.; Camacho, L.M.; Salem, A.Z.M.; Cerrillo-Soto, M.A.; El-Rayes, T. Influence of Jojoba Meal Treated with Lactobacillus Acidophilus on Digestibility, Carcass Traits and Blood Metabolites in Growing Rabbits. Anim. Nutr. Feed. Technol. 2013, 13, 423–434. [Google Scholar]
- El-Shafei, A.A.; Younis, T.M.; Al-Gamal, M.A.; Hesham, A.M. Impact of Probiotic (Lactobacillus planterium L.) Supplementation on Productive And Physiological Performance of Growing Rabbits under Egyptian Conditions. Egypt. J. Rabbit Sci. 2019, 29, 125–148. [Google Scholar] [CrossRef]
- Seyidoglu, N.; Peker, S. Effects of Different Doses of Probiotic Yeast Saccharomyces Cerevisiae on the Duodenal Mucosa in Rabbits. Indian J. Anim. Res. 2015, 49, 602–606. [Google Scholar] [CrossRef]
- Combes, S.; Fortun-Lamothe, L.; Cauquil, L.; Gidenne, T. Engineering the Rabbit Digestive Ecosystem to Improve Digestive Health and Efficacy. Animal 2013, 7, 1429–1439. [Google Scholar] [CrossRef] [Green Version]
- Cotozzolo, E.; Cremonesi, P.; Curone, G.; Menchetti, L.; Riva, F.; Biscarini, F.; Marongiu, M.L.; Castrica, M.; Castiglioni, B.; Miraglia, D.; et al. Characterization of Bacterial Microbiota Composition along the Gastrointestinal Tract in Rabbits. Animals 2020, 11, 31. [Google Scholar] [CrossRef] [PubMed]
- Velasco-Galilea, M.; Piles, M.; Viñas, M.; Rafel, O.; González-Rodríguez, O.; Guivernau, M.; Sánchez, J.P. Rabbit Microbiota Changes throughout the Intestinal Tract. Front. Microbiol. 2018, 9, 2144. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, A.; Lordan, C.; Ross, R.P.; Cotter, P.D. Gut Microbes from the Phylogenetically Diverse Genus Eubacterium and Their Various Contributions to Gut Health. Gut Microbes 2020, 12, 1802866. [Google Scholar] [CrossRef] [PubMed]
- Nan, W.; Si, H.; Yang, Q.; Shi, H.; Zhang, T.; Shi, Q.; Li, G.; Zhang, H.; Liu, H. Effect of Vitamin a Supplementation on Growth Performance, Serum Biochemical Parameters, Intestinal Immunity Response and Gut Microbiota in American Mink (Neovison Vison). Animals 2021, 11, 1577. [Google Scholar] [CrossRef]
- Beshara, M.M.; Alazab, A.M.; Fahim, H.; el Desoky, A.; Ragab, M.; el Shahat, A.E.G.; El-Gamal, A. Effect of Early Dietary Supplementation of Probiotic and Feed Restriction Post Weaning on Productive and Economical Performance of Growing Rabbits. Egypt. J. Rabbit Sci. 2018, 28, 195–222. [Google Scholar] [CrossRef]
- Mancini, S.; Paci, G. Probiotics in Rabbit Farming: Growth Performance, Health Status, and Meat Quality. Animals 2021, 11, 3388. [Google Scholar] [CrossRef] [PubMed]
- Fang, S.; Chen, X.; Ye, X.; Zhou, L.; Xue, S.; Gan, Q. Effects of Gut Microbiome and Short-Chain Fatty Acids (SCFAs) on Finishing Weight of Meat Rabbits. Front. Microbiol. 2020, 11, 1835. [Google Scholar] [CrossRef] [PubMed]
- Bovera, F.; Iannaccone, F.; Mastellone, V.; Nizza, S.; Lestingi, A.; de Martino, L.; Lombardi, P.; Mallardo, K.; Ferrara, M.; Nizza, A. Effect of Spray Application of Lactobacillus Plantarum on in Vivo Performance, Caecal Fermentations and Haematological Traits of Suckling Rabbits. Ital. J. Anim. Sci. 2016, 11, 145–149. [Google Scholar] [CrossRef]


{kind=link}
| Analytical Components (%) | Dietary Treatments | |
|---|---|---|
| First Period (0–14 Day) | Second Period (15–45 Day) | |
| CP | 15.50 | 16.30 |
| Fat | 2.70 | 2.50 |
| CF | 17.50 | 15.80 |
| Ash | 7.60 | 7.80 |
| Ca | 1.10 | 1.10 |
| P | 0.50 | 0.60 |
| Na | 0.20 | 0.30 |
| Trial 1 | Trial 2 | Trial 3 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| C | L | p-Value | C | L | p-Value | C | L | p-Value | |
| LW (g), mean (SD) | 3131.00 (97.53) | 3242.00 (106.40) | 0.448 | 3430.10 (258.76) | 3188.90 (419.52) | 0.068 | 3459.4 (375.89) | 3535.9 (299.06) | 0.542 |
| DFI (g), mean (SD) | 155.20 (6.95) | 163.20 (7.15) | 0.429 | 184.27 a (5.33) | 160.59 b (7.80) | 0.018 | 160.24 (20.61) | 162.08 (19.39) | 0.807 |
| ADG (g), mean (SD) | 45.40 (2.44) | 48.01 (2.41) | 0.455 | 51.77 (1.67) | 46.30 (2.34) | 0.068 | 47.73 (6.44) | 49.44 (5.27) | 0.444 |
| FCR (g), mean (SD) | 3.47 (0.12) | 3.43 (0.07) | 0.745 | 3.57 (0.05) | 3.48 (0.06) | 0.287 | 3.36 (0.15) | 3.28 (0.17) | 0.164 |
| BCS (1–5) | 2.91 a (0.04) | 3.00 b (0.00) | 0.038 | 3.07 (0.04) | 3.02 (0.06) | 0.225 | 3.20 (0.08) | 3.14 (0.07) | 1.000 |
| FS (0–4) | 3.76 (0.10) | 3.96 (0.01) | 0.131 | 3.98 (0.02) | 3.98 (0.01) | 0.633 | 3.92 (0.04) | 3.97 (0.02) | 0.594 |
| Trial 1 | Trial 2 | Trial 3 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| C | L | p-Value | C | L | p-Value | C | L | p-Value | |
| ALT (U/L), mean (SD) | 54.50 (6.85) | 45.08 (4.16) | 0.253 | 51.08 (3.86) | 49.17 (4.76) | 0.758 | 45.58 (5.56) | 48.58 (2.55) | 0.164 |
| AST (U/L), mean (SD) | 33.25 (3.59) | 33.92 (3.83) | 0.900 | 27.58 (3.04) | 33.75 (3.56) | 0.201 | 38.33 (2.46) | 38.33 (2.22) | >0.999 |
| COL (mg/dL), mean (SD) | 48. 83 (4.15) | 48.33 (3.34) | 0.926 | 43.00 (3.34) | 47.00 (3.72) | 0.432 | 48.25 (3.76) | 49.33 (4.76) | 0.860 |
| CRE (mg/dL), mean (SD) | 0.84 (0.05) | 0.94 (0.05) | 0.162 | 0.87 (0.04) | 0.87 (0.05) | 0.909 | 1.06 (0.04) | 0.95 (0.04) | 0.102 |
| GGT (U/L), mean (SD) | 9.00 (1.23) | 10.92 (2.73) | 0.529 | 7.17 (0.66) | 8.42 (2.46) | 0.629 | 9.00 (1.00) | 7.58 (1.49) | 0.438 |
| PRTOT (g/dL), mean (SD) | 6.07 (0.15) | 6.05 (0.11) | 0.893 | 6.20 (0.09) | 6.15 (0.11) | 0.734 | 6.32 (0.16) | 6.33 (0.13) | 0.969 |
| TRIGL (mg/dL), mean (SD) | 56.67 (4.45) | 54.42 (3.33) | 0.689 | 52.00 (4.03) | 51.50 (2.49) | 0.917 | 43.33 (1.74) | 40.75 (1.75) | 0.305 |
| UREA (mg/dL), mean (SD) | 18.17 (0.95) | 18.92 (0.76) | 0.545 | 19.17 (1.21) | 17.67 (1.12) | 0.372 | 53.17 (2.70) | 50.75 (2.63) | 0.528 |
| Cl (mmol/L), mean (SD) | 109.30 (2.67) | 109.70 (1.96) | 0.887 | 99.83 (3.49) | 98.74 (3.00) | 0.816 | 95.81 (1.03) | 95.83 (0.88) | 0.985 |
| K (mmol/L), mean (SD) | 9.95 (0.45) | 10.09 (0.53) | 0.847 | 9.13 (0.42) | 14.05 (5.02) | 0.339 | 11.70 (0.55) | 11.22 (0.42) | 0.500 |
| Na (mmol/L), mean (SD) | 161.20 (3.35) | 161.80 (2.64) | 0.888 | 150.3 (4.22) | 148.2 (3.56) | 0.720 | 138.7 (1.22) | 137.9 (1.27) | 0.678 |
| Trial 1 | Trial 2 | Trial 3 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| C | L | p-Value | C | L | P-Value | C | L | p-Value | |
| ALB (%), mean (SD) | 60.90 (0.90) | 60.45 (0.81) | 0.714 | 60.45 (0.81) | 59.58 (1.35) | 0.589 | 68.88 (0.78) | 67.28 (0.57) | 0.013 |
| ALB (g/dL), mean (SD) | 3.70 (0.10) | 3.66 (0.07) | 0.732 | 3.66 (0.07) | 3.66 (0.08) | 0.976 | 4.35 (0.11) | 4.26 (0.09) | 0.522 |
| ALFA-1 (%), mean (SD) | 7.73 (0.38) | 7.46 (0.20) | 0.529 | 7.46 (0.20) | 7.25 (0.27) | 0.548 | 7.33 (0.22) | 7.97 (0.23) | 0.060 |
| ALFA-1 (g/dL), mean (SD) | 0.47 (0.02) | 0.45 (0.01) | 0.527 | 0.45 (0.01) | 0.44 (0.01) | 0.777 | 0.46 (0.01) | 0.50 (0.01) | 0.020 |
| ALFA-2 (%), mean (SD) | 6.96 (0.15) | 7.35 (0.23) | 0.168 | 7.35 (0.23) | 8.25 (0.66) | 0.210 | 7.07 (0.20) | 7.16 (0.23) | 0.765 |
| ALFA-2 (g/dL), mean (SD) | 0.42 (0.01) | 0.44 (0.01) | 0.191 | 0.44 (0.01) | 0.51 (0.04) | 0.165 | 0.45 (0.02) | 0.45 (0.02) | 0.798 |
| BETA (%), mean (SD) | 13.08 (0.91) | 12.89 (0.58) | 0.867 | 12.89 (0.58) | 11.51 (0.75) | 0.157 | 8.92 (0.19) | 8.88 (0.23) | 0.890 |
| BETA (g/dL), mean (SD) | 0.79 (0.06) | 0.78 (0.04) | 0.852 | 0.78 (0.04) | 0.71 (0.05) | 0.288 | 0.57 (0.02) | 0.56 (0.01) | 0.787 |
| GAMMA (%), mean (SD) | 11.33 (0.67) | 11.85 (0.75) | 0.612 | 11.85 (0.75) | 13.41 (0.99) | 0.223 | 7.79 (0.59) | 8.70 (0.68) | 0.327 |
| GAMMA (g/dL), mean (SD) | 0.69 (0.04) | 0.72 (0.05) | 0.680 | 0.72 (0.05) | 0.83 (0.07) | 0.218 | 0.49 (0.04) | 0.55 (0.05) | 0.354 |
| A/G, mean (SD) | 1.57 (0.05) | 1.54 (0.05) | 0.680 | 1.54 (0.05) | 1.50 (0.08) | 0.705 | 6.32 (0.16) | 6.33 (0.14) | 0.969 |
| Trial 1 | Trial 2 | Trial 3 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| C | L | p-Value | C | L | p-Value | C | L | p-Value | |
| Duodenum | |||||||||
| Vh, mean (SD) | 0.84 (0.16) | 0.84 (0.25) | 0.977 | 0.72 (0.25) | 0.81 (0.27) | 0.291 | 0.95 (0.12) | 0.94 (0.15) | 0.921 |
| Cd, mean (SD) | 0.05 (0.01) | 0.04 (0.01) | 0.339 | 0.04 (0.01) | 0.04 (0.01) | 0.629 | 0.05 (0.01) | 0.05 (0.01) | 0.599 |
| Vh/Cd, mean (SD) | 16.04 (6.07) | 18.75 (5.15) | 0.409 | 15.53 (6.55) | 18.70 (6.15) | 0.233 | 18.46 (3.22) | 18.89 (4.33) | 0.781 |
| Jejunum | |||||||||
| Vh, mean (SD) | 0.76 (0.17) | 0.82 (0.19) | 0.498 | 0.78 (0.16) | 0.81 (0.13) | 0.665 | 0.93 (0.12) | 0.97 (0.12) | 0.517 |
| Cd, mean (SD) | 0.05 (0.01) | 0.04 (0.01) | 0.400 | 0.05 (0.01) | 0.05 (0.01) | 0.986 | 0.05 (0.01) | 0.05 (0.01) | 0.589 |
| Vh/Cd, mean (SD) | 16.86 (4.49) | 18.81 (4.75) | 0.313 | 16.07 (2.73) | 17.09 (4.14) | 0.480 | 18.19 (2.70) | 19.21 (3.05) | 0.396 |
| Ileum | |||||||||
| Vh, mean (SD) | 0.61 (0.20) | 0.58 (0.16) | 0.752 | 0.63 (0.12) | 0.60 (0.16) | 0.562 | 0.54 (0.08) | 0.52 (0.07) | 0.537 |
| Cd, mean (SD) | 0.05 (0.01) | 0.04 (0.01) | 0.434 | 0.04 (0.01) | 0.04 (0.01) | 0.843 | 0.05 (0.01) | 0.04 (0.01) | 0.029 |
| Vh/Cd, mean (SD) | 12.88 (4.99) | 12.50 (3.29) | 0.826 | 14.29 (3.77) | 13.23 (4.06) | 0.516 | 11.27 (2.69) | 12.17 (2.84) | 0.434 |
| Trial 1 | Trial 2 | Trial 3 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| C | L | p-Value | C | L | p-Value | C | L | p-Value | |
| Liver, median (IR) | 0.25 (0.00–1.00) | 0.00 (0.00) | 0.651 | 0.58 (0.00–1.00) | 0.62 (0.00–1.00) | 0.999 | 0.50 (0.50–1.00) | 0.50 (0.12–0.87) | 0.286 |
| Spleen, mean (SD) | 0.22 (0.51) | 0.33 (0.49) | 0.640 | 0.00 (0.00) | 0.18 (0.60) | 0.640 | 0.00 (0.00) | 0.00 (0.00) | >0.999 |
| Duodenum, median (IR) | 0.50 (0.00–0.50) | 0.50 (0.00–0.75) | 0.477 | 0.50 (0.00–0.50) | 0.50 (0.00–1.00) | 0.115 | 0.50 (0.50–1.00) | 0.50 (0.00–0.87) | 0.882 |
| Jejunum, median (IR) | 0.50 (0.00–1.00) | 0.50 (0.00–0.50) | 0.352 | 0.50 (0.00–0.87) | 0.50 (0.00–0.50) | 0.710 | 0.50 (0.12–0.50) | 0.50 (0.00–0.50) | 0.400 |
| Ileum, median (IR) | 0.25 (0.00–0.50) | 0.50 (0.00–1.00) | 0.379 | 0.25 (0.00–0.50) | 0.50 (0.00–1.00) | 0.246 | 0.50 (0.00–0.50) | 0.50 (0.00–0.50) | 0.689 |
| Caecum, median (IR) | 0.45 (0.49) | 0.37 (0.48) | 0.840 | 0.50 (0.00–1.00) | 0.25 (0.00–1.00) | 0.576 | 0.00 (0.00–0.50) | 0.50 (0.00–0.87) | 0.146 |
| Trial 1 | Trial 2 | Trial 3 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| C | L | p-Value | C | L | p-Value | C | L | p-Value | |
| Acetic acid, mean (SD) | 139.50 (135.8) | 148.90 (73.05) | 0.346 | 80.05 (39.90) | 78.91 (37.05) | 0.944 | 84.82 (33.18) | 128.70 (93.94) | 0.198 |
| Lactic acid, mean (SD) | 111.30 (156.1) | 121.00 (72.12) | 0.167 | 142.00 (140.50) | 91.90 (50.60) | 0.315 | n.d | n.d | n.d |
| Succinic acid, mean (SD) | 81.78 (42.62) | 112.00 (66.11) | 0.258 | 58.21 (79.00) | 54.44 (20.25) | 0.143 | 38.67 (50.30) | 149.80 (83.73) | <0.001 |
| Isobutyric acid, mean (SD) | 431.10 (298.40) | 469.10 (254.50) | 0.744 | 501.50 (551.30) | 312.30 (131.70) | 0.607 | 304.20 (134.60) | 293.20 (186.70) | 0.882 |
| Propionic acid, mean (SD) | 40.22 (22.28) | 42.78 (12.81) | 0.251 | 49.59 (35.99) | 27.30 (6.58) | 0.210 | 13.77 (14.64) | 5.59 (11.19) | 0.385 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Colombino, E.; Biasato, I.; Michetti, A.; Rubino, M.G.; Franciosa, I.; Giribaldi, M.; Antoniazzi, S.; Bergagna, S.; Paliasso, G.; Ferrocino, I.; et al. Effects of Dietary Supplementation of Lactobacillus acidophilus on Blood Parameters and Gut Health of Rabbits. Animals 2022, 12, 3543. https://doi.org/10.3390/ani12243543
Colombino E, Biasato I, Michetti A, Rubino MG, Franciosa I, Giribaldi M, Antoniazzi S, Bergagna S, Paliasso G, Ferrocino I, et al. Effects of Dietary Supplementation of Lactobacillus acidophilus on Blood Parameters and Gut Health of Rabbits. Animals. 2022; 12(24):3543. https://doi.org/10.3390/ani12243543
Chicago/Turabian StyleColombino, Elena, Ilaria Biasato, Alberta Michetti, Maria Gabriella Rubino, Irene Franciosa, Marzia Giribaldi, Sara Antoniazzi, Stefania Bergagna, Giulia Paliasso, Ilario Ferrocino, and et al. 2022. "Effects of Dietary Supplementation of Lactobacillus acidophilus on Blood Parameters and Gut Health of Rabbits" Animals 12, no. 24: 3543. https://doi.org/10.3390/ani12243543