
Intestinal Immune Cell Populations, Barrier Function, and Microbiomes in Broilers Fed a Diet Supplemented with Chlorella vulgaris
 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Microalgae Preparation
2.2. Animals, Diets, and Experimental Design
2.3. Sample Collection
2.4. Jejunal Lamina Propria Isolation
2.5. Flow Cytometry Analysis
2.6. RNA Extraction and Reverse Transcription
2.7. Quantitative Real-Time PCR Analysis
2.8. Serum Antioxidant Enzyme Analysis
2.9. Jejunal Histology
2.10. Cecal Short-Chain Fatty Acids Analysis
2.11. Cecal Microbiome Analysis
2.12. Statistical Analysis
3. Results
3.1. Growth Performance
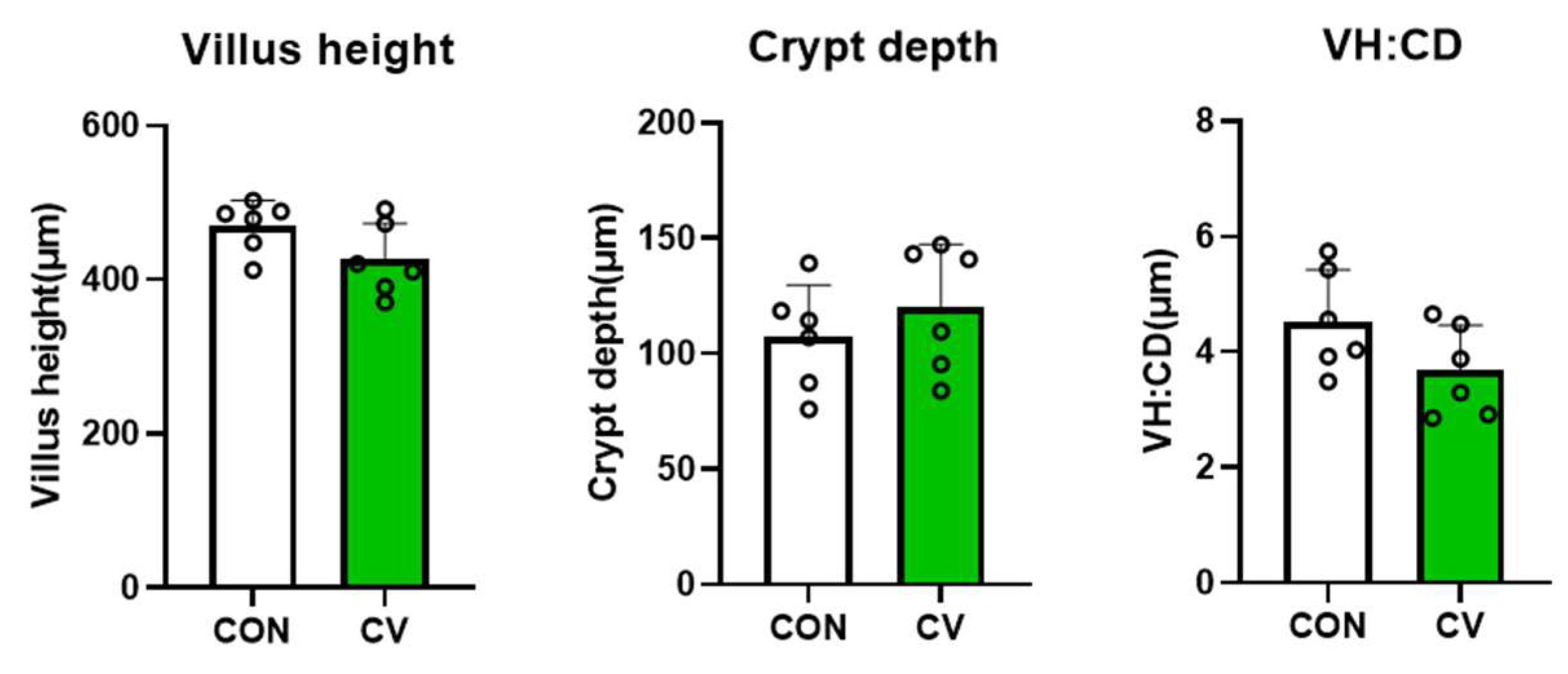
3.2. Jejunal Morphology
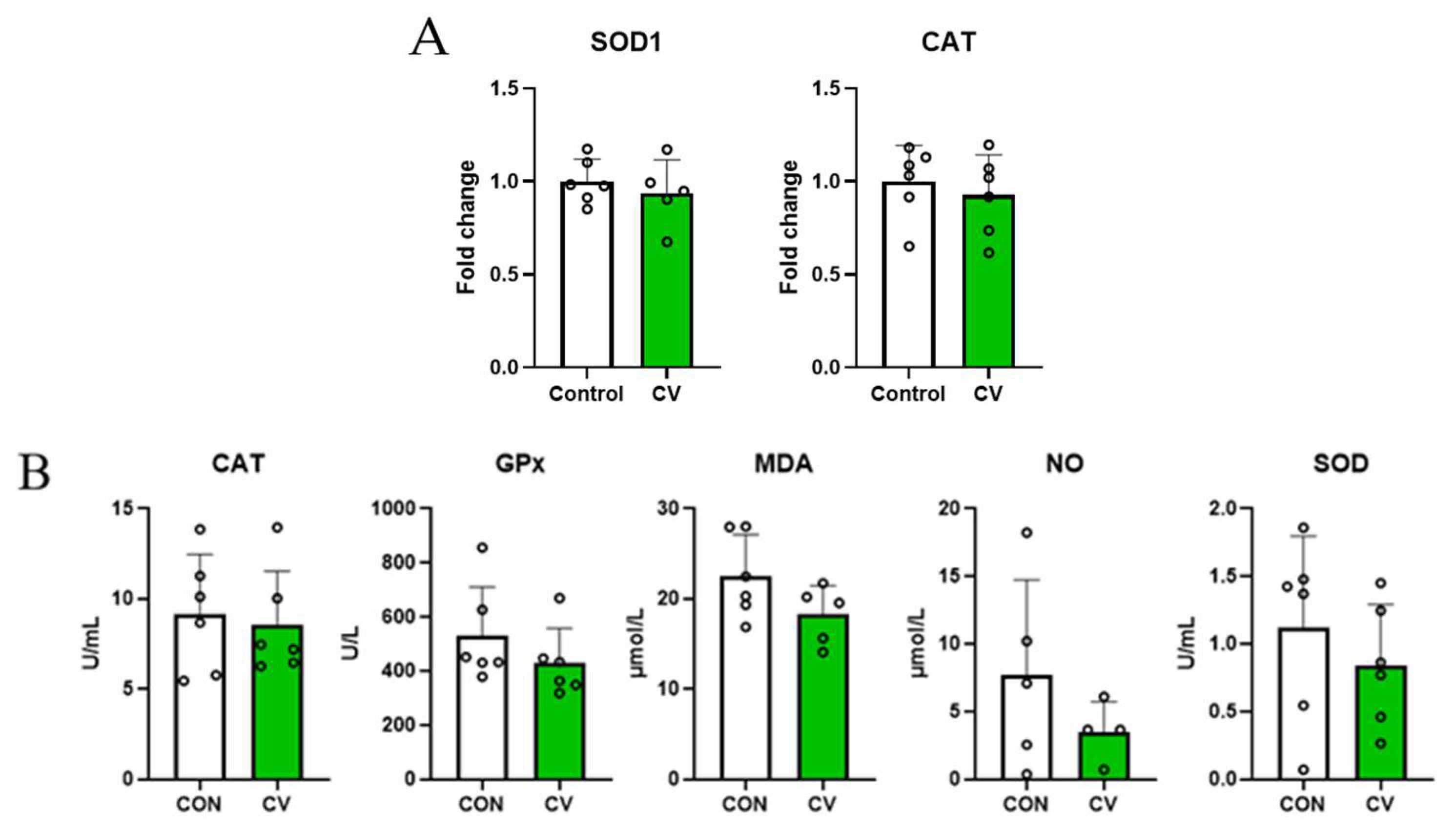
3.3. Relative mRNA Expression of Antioxdant Enzymes in Jejunal Tissues and Antioxidant Enzyme Concentrations in Serum
3.4. Relative mRNA Expression of Intestinal Epithelial Barrier Functions in Jejunal Tissues
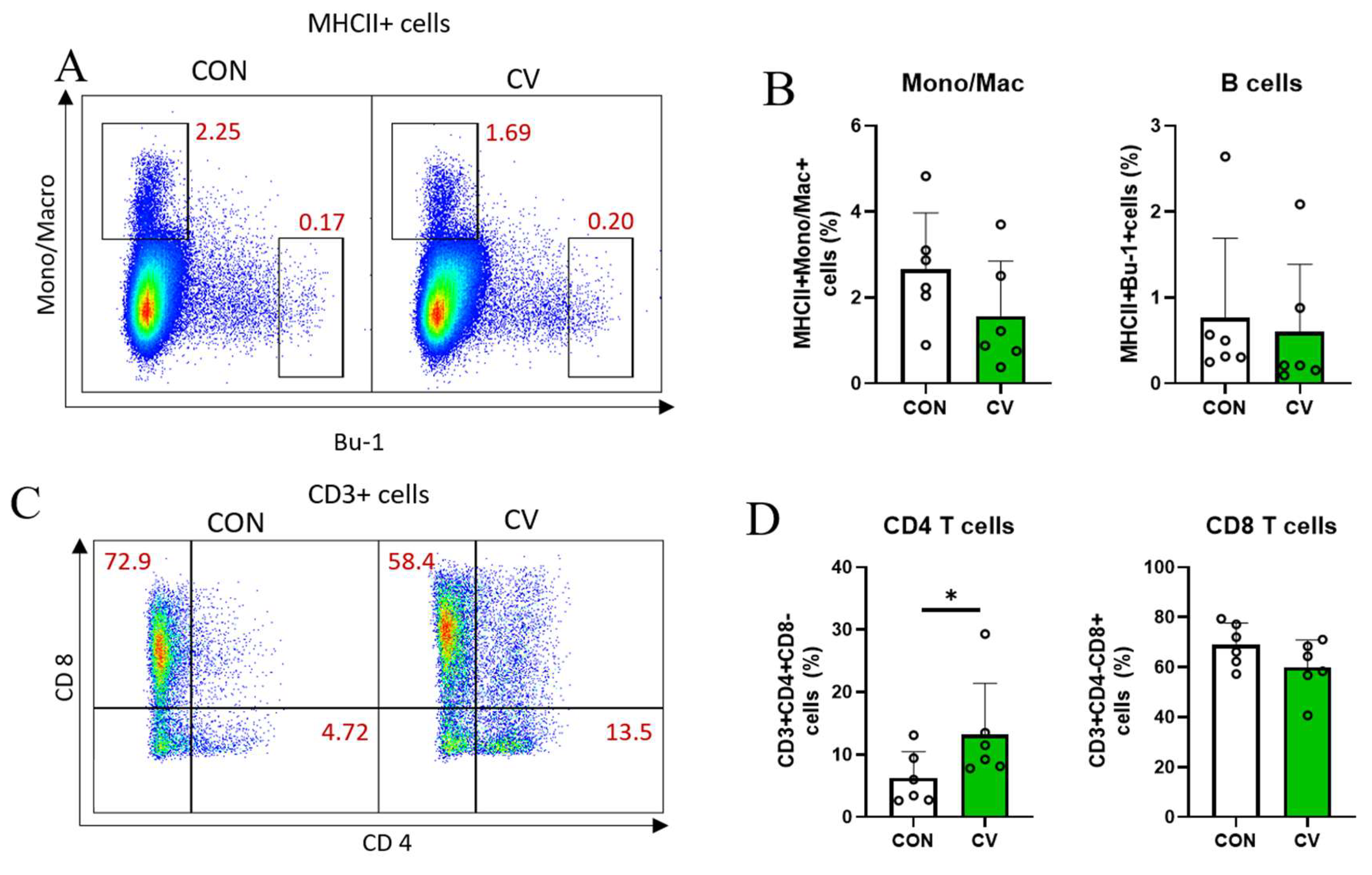
3.5. Immune Cell Populations in Jejunal Lamina Propria
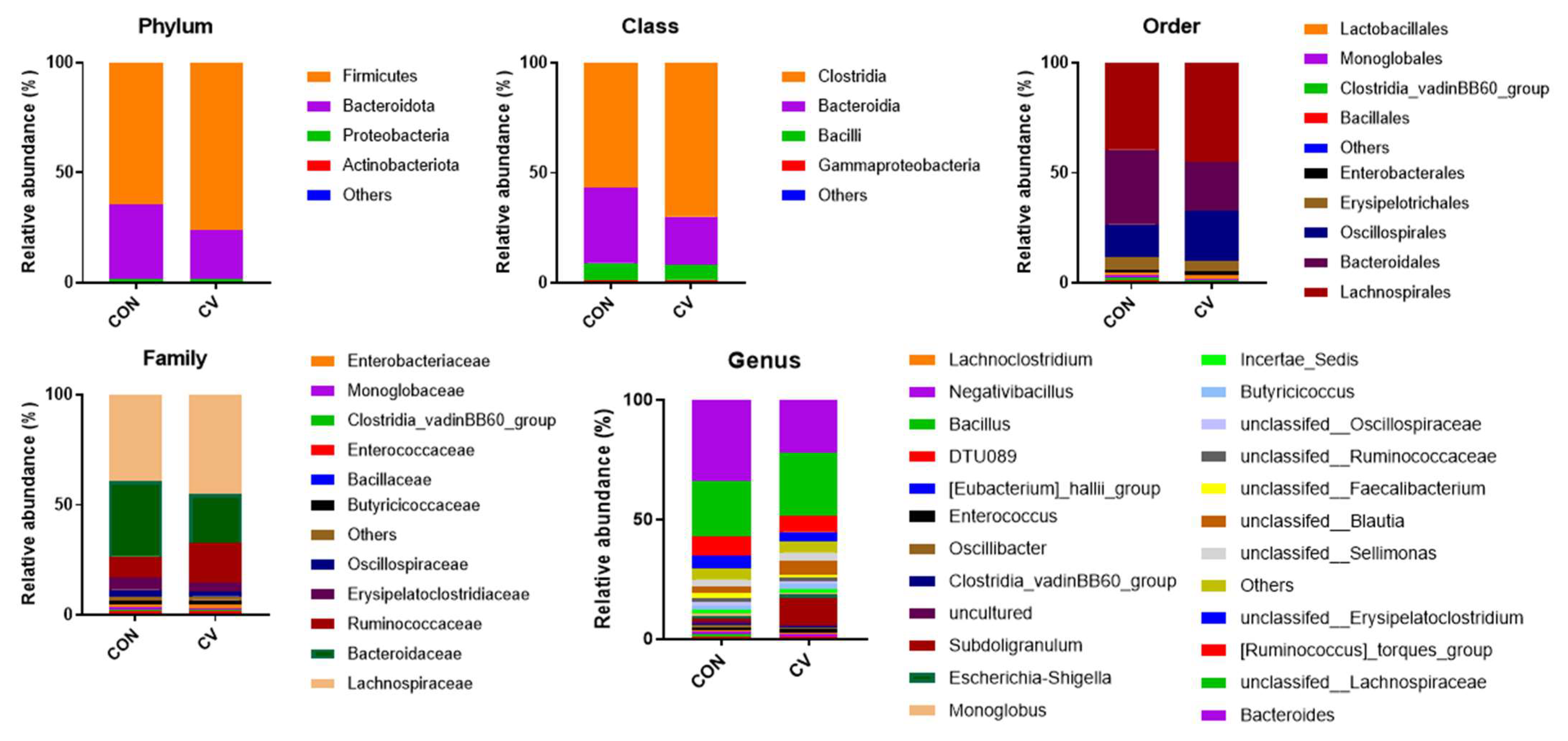
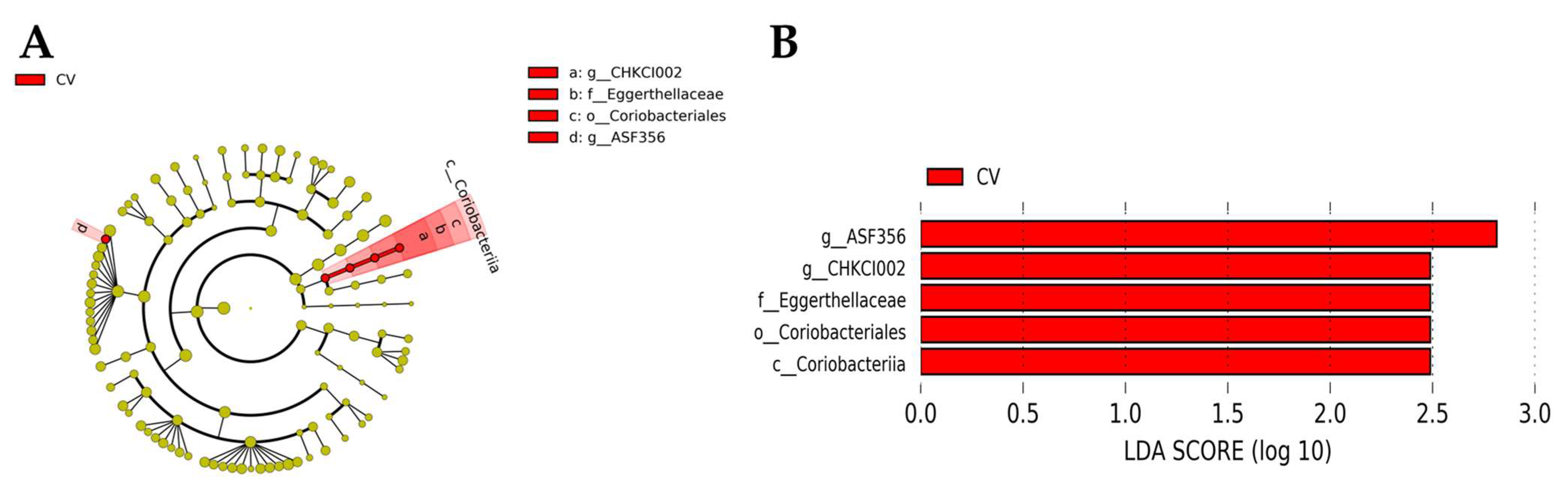
3.6. Cecal Microbiome
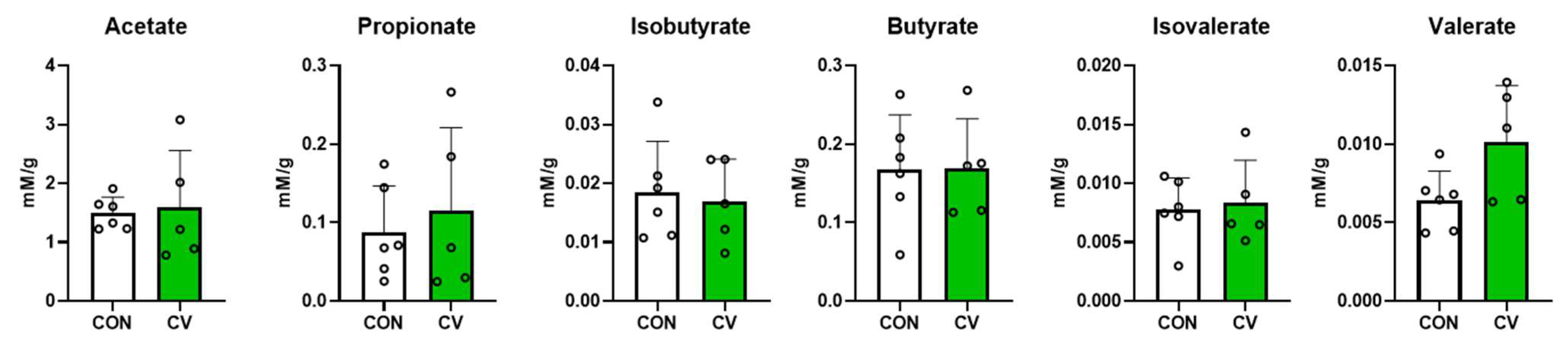
3.7. Concentrations of Short-Chain Fatty Acids in Cecal Digesta
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Van der Poel, A.; Abdollahi, M.; Cheng, H.; Colovic, R.; Den Hartog, L.; Miladinovic, D.; Page, G.; Sijssens, K.; Smillie, J.; Thomas, M.; et al. Future directions of animal feed technology research to meet the challenges of a changing world. Anim. Feed. Sci. Technol. 2020, 270, 114692. [Google Scholar] [CrossRef]
- Te Pas, M.F.; Veldkamp, T.; de Haas, Y.; Bannink, A.; Ellen, E.D. Adaptation of livestock to new diets using feed components without competition with human edible protein sources—A review of the possibilities and recommendations. Animals 2021, 11, 2293. [Google Scholar] [CrossRef]
- Martins, C.F.; Ribeiro, D.M.; Costa, M.; Coelho, D.; Alfaia, C.M.; Lordelo, M.; Almeida, A.M.; Freire, J.P.; Prates, J.A. Using microalgae as a sustainable feed resource to enhance quality and nutritional value of pork and poultry meat. Foods 2021, 10, 2933. [Google Scholar] [CrossRef]
- Wang, B.; Li, Y.; Wu, N.; Lan, C.Q. CO 2 bio-mitigation using microalgae. Appl. Microbiol. Biotechnol. 2008, 79, 707–718. [Google Scholar] [CrossRef]
- Li, D.; Wang, L.; Zhao, Q.; Wei, W.; Sun, Y. Improving high carbon dioxide tolerance and carbon dioxide fixation capability of Chlorella sp. by adaptive laboratory evolution. Bioresour. Technol. 2015, 185, 269–275. [Google Scholar] [CrossRef]
- Coudert, E.; Baéza, E.; Berri, C. Use of algae in poultry production: A review. World’s Poult. Sci. J. 2020, 76, 767–786. [Google Scholar] [CrossRef]
- Abdelnour, S.; Abd El-Hack, M.; Arif, M.; Khafaga, A.; Taha, A. The application of the microalgae Chlorella spp. as a supplement in broiler feed. World’s Poult. Sci. J. 2019, 75, 305–318. [Google Scholar] [CrossRef]
- Madeira, M.S.; Cardoso, C.; Lopes, P.A.; Coelho, D.; Afonso, C.; Bandarra, N.M.; Prates, J.A. Microalgae as feed ingredients for livestock production and meat quality: A review. Livest. Sci. 2017, 205, 111–121. [Google Scholar] [CrossRef]
- Kusmayadi, A.; Leong, Y.K.; Yen, H.-W.; Huang, C.-Y.; Chang, J.-S. Microalgae as sustainable food and feed sources for animals and humans–biotechnological and environmental aspects. Chemosphere 2021, 271, 129800. [Google Scholar] [CrossRef] [PubMed]
- Niccolai, A.; Zittelli, G.C.; Rodolfi, L.; Biondi, N.; Tredici, M.R. Microalgae of interest as food source: Biochemical composition and digestibility. Algal Res. 2019, 42, 101617. [Google Scholar] [CrossRef]
- Oh, S.T.; Zheng, L.; Kwon, H.; Choo, Y.; Lee, K.; Kang, C.; An, B.-K. Effects of dietary fermented Chlorella vulgaris (CBT®) on growth performance, relative organ weights, cecal microflora, tibia bone characteristics, and meat qualities in Pekin ducks. Asian-Australas. J. Anim. Sci. 2015, 28, 95. [Google Scholar] [CrossRef]
- Alfaia, C.; Pestana, J.; Rodrigues, M.; Coelho, D.; Aires, M.; Ribeiro, D.; Major, V.; Martins, C.; Santos, H.; Lopes, P.; et al. Influence of dietary Chlorella vulgaris and carbohydrate-active enzymes on growth performance, meat quality and lipid composition of broiler chickens. Poult. Sci. 2021, 100, 926–937. [Google Scholar] [CrossRef]
- El-Bahr, S.; Shousha, S.; Shehab, A.; Khattab, W.; Ahmed-Farid, O.; Sabike, I.; El-Garhy, O.; Albokhadaim, I.; Albosadah, K. Effect of dietary microalgae on growth performance, profiles of amino and fatty acids, antioxidant status, and meat quality of broiler chickens. Animals 2020, 10, 761. [Google Scholar] [CrossRef]
- An, S.H.; Joo, S.S.; Lee, H.G.; Kim, Z.-H.; Lee, C.S.; Kim, M.; Kong, C. Supplementation of Indigenous Green Microalga (Parachlorella sp.) to Pre-starter Diet for Broiler Chickens. Korean J. Poult. Sci. 2020, 47, 49–59. [Google Scholar] [CrossRef] [Green Version]
- An, B.-K.; Kim, K.-E.; Jeon, J.-Y.; Lee, K.W. Effect of dried Chlorella vulgaris and Chlorella growth factor on growth performance, meat qualities and humoral immune responses in broiler chickens. Springerplus 2016, 5, 718. [Google Scholar] [CrossRef] [Green Version]
- Martins, C.F.; Trevisi, P.; Coelho, D.F.; Correa, F.; Ribeiro, D.M.; Alfaia, C.M.; Pinho, M.; Pestana, J.M.; Mourato, M.P.; Almeida, A.M.; et al. Influence of Chlorella vulgaris on growth, digestibility and gut morphology and microbiota of weaned piglet. Sci. Rep. 2022, 12, 6012. [Google Scholar] [CrossRef]
- Awad, W.A.; Mann, E.; Dzieciol, M.; Hess, C.; Schmitz-Esser, S.; Wagner, M.; Hess, M. Age-related differences in the luminal and mucosa-associated gut microbiome of broiler chickens and shifts associated with Campylobacter jejuni infection. Front. Cell. Infect. Microbiol. 2016, 6, 154. [Google Scholar] [CrossRef] [Green Version]
- Kim, Z.-H.; Kim, K.; Park, H.; Lee, C.S.; Nam, S.W.; Yim, K.J.; Jung, J.Y.; Hong, S.-J.; Lee, C.-G. Enhanced fatty acid productivity by Parachlorella sp., a freshwater microalga, via adaptive laboratory evolution under salt stress. Biotechnol. Bioprocess Eng. 2021, 26, 223–231. [Google Scholar] [CrossRef]
- Czerkawski, J. The use of pivalic acid as a reference substance in measurements of production of volatile fatty acids by rumen micro-organisms in vitro. Br. J. Nutr. 1976, 36, 311–315. [Google Scholar] [CrossRef] [Green Version]
- Kang, H.; Salim, H.; Akter, N.; Kim, D.; Kim, J.; Bang, H.; Kim, M.; Na, J.; Hwangbo, J.; Choi, H.; et al. Effect of various forms of dietary Chlorella supplementation on growth performance, immune characteristics, and intestinal microflora population of broiler chickens. J. Appl. Poult. Res. 2013, 22, 100–108. [Google Scholar] [CrossRef]
- Kang, H.; Park, S.; Kim, C. Effects of dietary supplementation with a chlorella by-product on the growth performance, immune response, intestinal microflora and intestinal mucosal morphology in broiler chickens. J. Anim. Physiol. Anim. Nutr. 2017, 101, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Cabrol, M.B.; Martins, J.C.; Malhão, L.P.; Alves, S.P.; Bessa, R.J.; Almeida, A.M.; Raymundo, A.; Lordelo, M. Partial replacement of soybean meal with Chlorella vulgaris in broiler diets influences performance and improves breast meat quality and fatty acid composition. Poult. Sci. 2022, 101, 101955. [Google Scholar] [CrossRef] [PubMed]
- Safi, C.; Zebib, B.; Merah, O.; Pontalier, P.-Y.; Vaca-Garcia, C. Morphology, composition, production, processing and applications of Chlorella vulgaris: A review. Renew. Sustain. Energy Rev. 2014, 35, 265–278. [Google Scholar] [CrossRef] [Green Version]
- Awad, W.; Ghareeb, K.; Böhm, J. Intestinal structure and function of broiler chickens on diets supplemented with a synbiotic containing Enterococcus faecium and oligosaccharides. Int. J. Mol. Sci. 2008, 9, 2205–2216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schat, K.; Myers, T. Avian intestinal immunity. Crit. Rev. Poult. Biol. 1991, 3, 19–34. [Google Scholar]
- Olukosi, O.; Dono, N. Modification of digesta pH and intestinal morphology with the use of benzoic acid or phytobiotics and the effects on broiler chicken growth performance and energy and nutrient utilization. J. Anim. Sci. 2014, 92, 3945–3953. [Google Scholar] [CrossRef] [Green Version]
- Montagne, L.; Pluske, J.; Hampson, D. A review of interactions between dietary fibre and the intestinal mucosa, and their consequences on digestive health in young non-ruminant animals. Anim. Feed. Sci. Technol. 2003, 108, 95–117. [Google Scholar] [CrossRef]
- Mirzaie, S.; Sharifi, S.D.; Zirak-Khattab, F. The effect of a Chlorella by-product dietary supplement on immune response, antioxidant status, and intestinal mucosal morphology of broiler chickens. J. Appl. Phycol. 2020, 32, 1771–1777. [Google Scholar] [CrossRef]
- Roques, S.; Koopmans, S.-J.; Mens, A.; van Harn, J.; van Krimpen, M.; Kar, S.K. Effect of Feeding 0.8% Dried Powdered Chlorella vulgaris Biomass on Growth Performance, Immune Response, and Intestinal Morphology during Grower Phase in Broiler Chickens. Animals 2022, 12, 1114. [Google Scholar] [CrossRef]
- Roushdy, E.M.; Zaglool, A.W.; El-Tarabany, M.S. Effects of chronic thermal stress on growth performance, carcass traits, antioxidant indices and the expression of HSP70, growth hormone and superoxide dismutase genes in two broiler strains. J. Therm. Biol. 2018, 74, 337–343. [Google Scholar] [CrossRef]
- Goel, A.; Ncho, C.M.; Choi, Y.-H. Regulation of gene expression in chickens by heat stress. J. Anim. Sci. Biotechnol. 2021, 12, 11. [Google Scholar] [CrossRef]
- Fellenberg, M.; Speisky, H. Antioxidants: Their effects on broiler oxidative stress and its meat oxidative stability. World’s Poult. Sci. J. 2006, 62, 53–70. [Google Scholar] [CrossRef]
- Nabi, F.; Arain, M.A.; Rajput, N.; Alagawany, M.; Soomro, J.; Umer, M.; Soomro, F.; Wang, Z.; Ye, R.; Liu, J. Health benefits of carotenoids and potential application in poultry industry: A review. J. Anim. Physiol. Anim. Nutr. 2020, 104, 1809–1818. [Google Scholar] [CrossRef] [Green Version]
- Lauretani, F.; Semba, R.D.; Dayhoff-Brannigan, M.; Corsi, A.M.; Di Iorio, A.; Buiatti, E.; Bandinelli, S.; Guralnik, J.M.; Ferrucci, L. Low total plasma carotenoids are independent predictors of mortality among older persons: The InCHIANTI study. Eur. J. Nutr. 2008, 47, 335–340. [Google Scholar] [CrossRef] [Green Version]
- Coelho, D.; Alfaia, C.M.; Lopes, P.A.; Pestana, J.M.; Costa, M.M.; Pinto, R.M.; Almeida, J.M.; Moreira, O.; Fontes, C.M.; Prates, J.A. Impact of Chlorella vulgaris as feed ingredient and carbohydrases on the health status and hepatic lipid metabolism of finishing pigs. Res. Vet. Sci. 2022, 144, 44–53. [Google Scholar] [CrossRef]
- Van der Sluis, M.; De Koning, B.A.; De Bruijn, A.C.; Velcich, A.; Meijerink, J.P.; Van Goudoever, J.B.; Büller, H.A.; Dekker, J.; Van Seuningen, I.; Renes, I.B. Muc2-deficient mice spontaneously develop colitis, indicating that MUC2 is critical for colonic protection. Gastroenterology 2006, 131, 117–129. [Google Scholar] [CrossRef]
- Van Itallie, C.M.; Anderson, J.M. The molecular physiology of tight junction pores. Physiology 2004, 19, 331–338. [Google Scholar] [CrossRef]
- Feldman, G.J.; Mullin, J.M.; Ryan, M.P. Occludin: Structure, function and regulation. Adv. Drug Deliv. Rev. 2005, 57, 883–917. [Google Scholar] [CrossRef]
- Paul, S.S.; Vantharam Venkata, H.G.R.; Raju, M.V.; Rama Rao, S.V.; Nori, S.S.; Suryanarayan, S.; Kumar, V.; Perveen, Z.; Prasad, C.S. Dietary supplementation of extracts of red sea weed (Kappaphycus alvarezii) improves growth, intestinal morphology, expression of intestinal genes and immune responses in broiler chickens. J. Sci. Food Agric. 2021, 101, 997–1008. [Google Scholar] [CrossRef]
- Guo, W.; Zhu, S.; Feng, G.; Wu, L.; Feng, Y.; Guo, T.; Yang, Y.; Wu, H.; Zeng, M. Microalgae aqueous extracts exert intestinal protective effects in Caco-2 cells and dextran sodium sulphate-induced mouse colitis. Food Funct. 2020, 11, 1098–1109. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, L.; Yu, C.; Yang, X.-F.; Wang, H. Monocyte and macrophage differentiation: Circulation inflammatory monocyte as biomarker for inflammatory diseases. Biomark. Res. 2014, 2, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wynn, T.A.; Chawla, A.; Pollard, J.W. Macrophage biology in development, homeostasis and disease. Nature 2013, 496, 445–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dal Pont, G.C.; Lee, A.; Bortoluzzi, C.; Farnell, Y.; Gougoulias, C.; Kogut, M. Novel model for chronic intestinal inflammation in chickens: (2) Immunologic mechanism behind the inflammatory response. Dev. Comp. Immunol. 2023, 138, 104524. [Google Scholar] [CrossRef] [PubMed]
- Tay, R.E.; Richardson, E.K.; Toh, H.C. Revisiting the role of CD4+ T cells in cancer immunotherapy—New insights into old paradigms. Cancer Gene Ther. 2021, 28, 5–17. [Google Scholar] [CrossRef] [PubMed]
- Fries-Craft, K.; Meyer, M.; Bobeck, E. Algae-based feed ingredient protects intestinal health during Eimeria challenge and alters systemic immune responses with differential outcomes observed during acute feed restriction. Poult. Sci. 2021, 100, 101369. [Google Scholar] [CrossRef] [PubMed]
- Song, B.; Tang, D.; Yan, S.; Fan, H.; Li, G.; Shahid, M.S.; Mahmood, T.; Guo, Y. Effects of age on immune function in broiler chickens. J. Anim. Sci. Biotechnol. 2021, 12, 42. [Google Scholar] [CrossRef]
- Ao, Z.; Kocher, A.; Choct, M. Effects of dietary additives and early feeding on performance, gut development and immune status of broiler chickens challenged with Clostridium perfringens. Asian-Australas. J. Anim. Sci. 2012, 25, 541. [Google Scholar] [CrossRef] [Green Version]
- Borda-Molina, D.; Seifert, J.; Camarinha-Silva, A. Current perspectives of the chicken gastrointestinal tract and its microbiome. Comput. Struct. Biotechnol. J. 2018, 16, 131–139. [Google Scholar] [CrossRef]
- Kim, B.-R.; Shin, J.; Guevarra, R.B.; Lee, J.H.; Kim, D.W.; Seol, K.-H.; Lee, J.-H.; Kim, H.B.; Isaacson, R.E. Deciphering diversity indices for a better understanding of microbial communities. J. Microbiol. Biotechnol. 2017, 27, 2089–2093. [Google Scholar] [CrossRef] [Green Version]
- Biggs, M.B.; Medlock, G.L.; Moutinho, T.J.; Lees, H.J.; Swann, J.R.; Kolling, G.L.; Papin, J.A. Systems-level metabolism of the altered Schaedler flora, a complete gut microbiota. ISME J. 2017, 11, 426–438. [Google Scholar] [CrossRef] [Green Version]
- Farkas, V.; Csitári, G.; Menyhárt, L.; Such, N.; Pál, L.; Husvéth, F.; Rawash, M.A.; Mezőlaki, Á.; Dublecz, K. Microbiota composition of mucosa and interactions between the microbes of the different gut segments could be a factor to modulate the growth rate of broiler chickens. Animals 2022, 12, 1296. [Google Scholar] [CrossRef]
- Kim, M.; Qie, Y.; Park, J.; Kim, C.H. Gut microbial metabolites fuel host antibody responses. Cell Host Microbe 2016, 20, 202–214. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Diets 1 | ||
|---|---|---|
| Item | Control | Chlorella vulgaris |
| Ingredient compositions, % | ||
| Corn | 53.54 | |
| Soybean meal | 38.80 | |
| Cornstarch | 0.50 | – |
| Microalgae (Chlorella vulgaris) | – | 0.50 |
| Soybean oil | 2.00 | |
| L-arginine | 0.09 | |
| L-histidine | 0.03 | |
| L-isoleucine | 0.10 | |
| L-lysine-HCl | 0.35 | |
| L-methionine | 0.24 | |
| L-cysteine | 0.14 | |
| L-threonine | 0.14 | |
| L-valine | 0.2 | |
| Limestone | 1.06 | |
| Dicalcium phosphate | 1.91 | |
| Sodium chloride | 0.40 | |
| Vitamin premix 2 | 0.20 | |
| Mineral premix 3 | 0.20 | |
| Choline chloride | 0.10 | |
| Calculated chemical compositions, % | ||
| MEn, kcal/kg | 2976 | 2972 |
| Crude protein | 23.0 | 23.0 |
| Total calcium | 0.96 | 0.96 |
| Non-phytate phosphorus | 0.48 | 0.48 |
| Calculated amino acids compositions, % | ||
| SID arginine | 1.31 | |
| SID histidine | 0.47 | |
| SID isoleucine | 0.87 | |
| SID leucine | 1.53 | |
| SID lysine | 1.28 | |
| SID methionine | 0.48 | |
| SID cysteine | 0.40 | |
| SID phenylalanine | 0.92 | |
| SID threonine | 0.81 | |
| SID tryptophan | 0.23 | |
| SID valine | 1.01 | |
| Item | Experimental Diets | SEM | p-Value | |
|---|---|---|---|---|
| Control | Chlorella vulgaris | |||
| Body weight gain, g/bird | 156.2 | 150.9 | 5.38 | 0.520 |
| Feed intake, g/bird | 175.0 | 169.1 | 3.27 | 0.258 |
| Gain to feed ratio, g/g | 0.89 | 0.89 | 0.016 | 0.929 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.Y.; Yoon, J.H.; An, S.H.; Cho, I.H.; Lee, C.W.; Jeon, Y.J.; Joo, S.S.; Ban, B.C.; Lee, J.-Y.; Jung, H.J.; et al. Intestinal Immune Cell Populations, Barrier Function, and Microbiomes in Broilers Fed a Diet Supplemented with Chlorella vulgaris. Animals 2023, 13, 2380. https://doi.org/10.3390/ani13142380
Lee JY, Yoon JH, An SH, Cho IH, Lee CW, Jeon YJ, Joo SS, Ban BC, Lee J-Y, Jung HJ, et al. Intestinal Immune Cell Populations, Barrier Function, and Microbiomes in Broilers Fed a Diet Supplemented with Chlorella vulgaris. Animals. 2023; 13(14):2380. https://doi.org/10.3390/ani13142380
Chicago/Turabian StyleLee, Ji Young, June Hyeok Yoon, Su Hyun An, In Ho Cho, Chae Won Lee, Yun Ji Jeon, Sang Seok Joo, Byeong Cheol Ban, Jae-Yeong Lee, Hyun Jung Jung, and et al. 2023. "Intestinal Immune Cell Populations, Barrier Function, and Microbiomes in Broilers Fed a Diet Supplemented with Chlorella vulgaris" Animals 13, no. 14: 2380. https://doi.org/10.3390/ani13142380