
Characterization of the Caudal Ventral Tubercle in the Sixth Cervical Vertebra in Modern Equus ferus caballus



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Statement
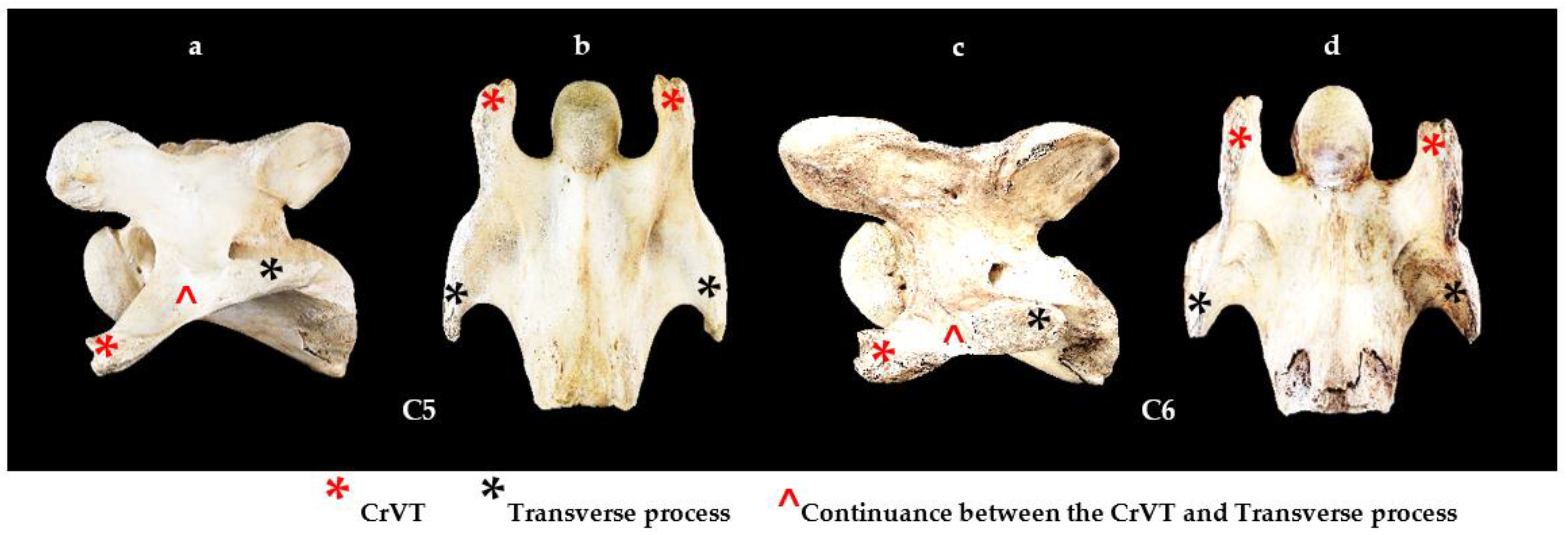
2.2. Terminology
2.3. Materials
2.4. Methods
2.5. Statistical Analysis
3. Results
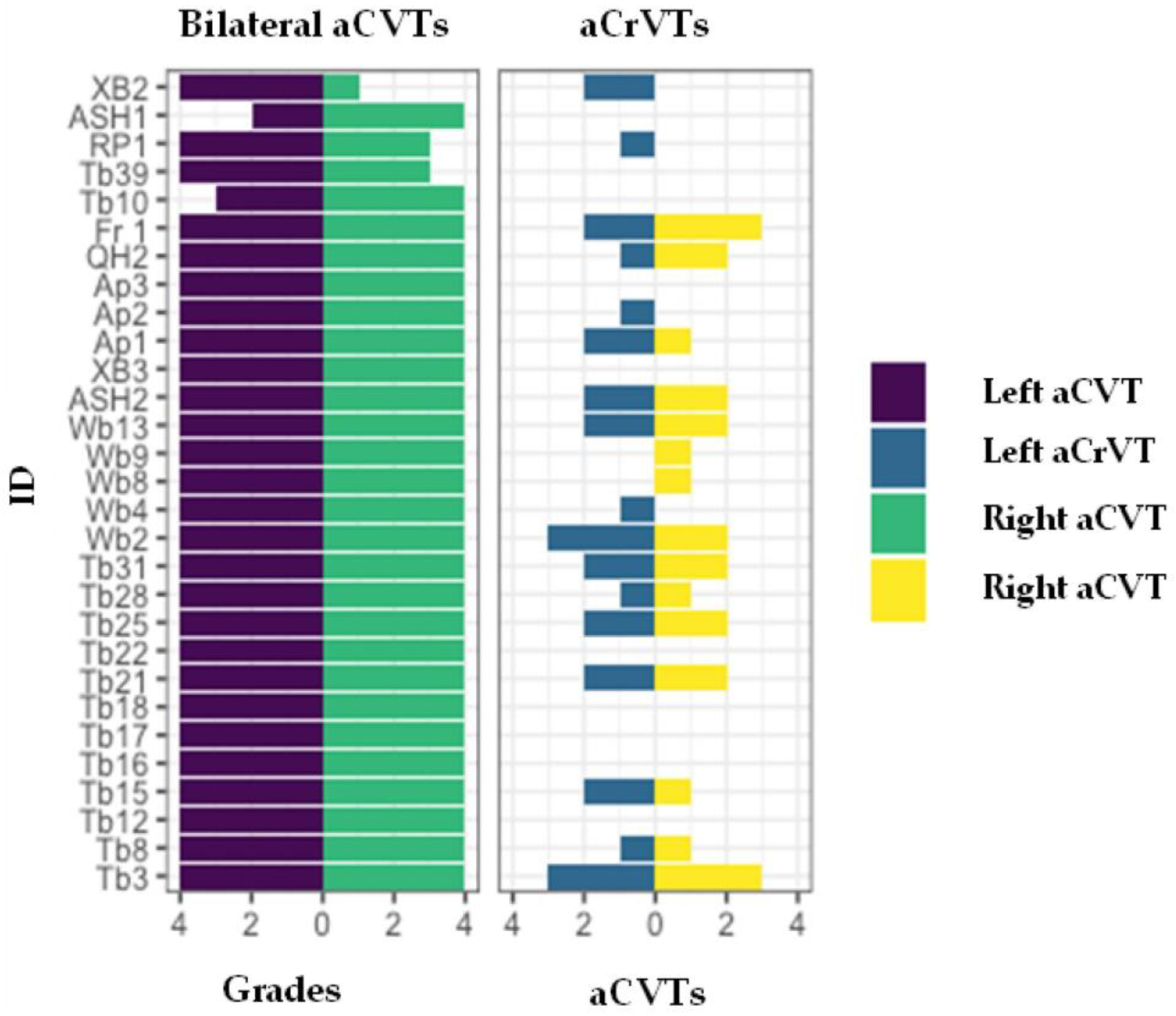
3.1. Individual Grades
3.2. Statistical Analysis: Absent CVTs by Morphology—Bilateral or Unilateral
3.3. Statistical Analysis: Absent CrVT Patterns of Loss
3.4. Bilateral Presentations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arnold, P. Evolution of the Mammalian Neck and Development, Morpho-functional, and Paleontological Perspectives. J. Mammal. Evol. 2021, 28, 173–183. [Google Scholar] [CrossRef]
- Arnold, P.; Amson, E.; Fischer, M.S. Differential Scaling Patterns of Vertebrae and the Evolution of the Neck Length in Mammals. Evolution 2017, 71, 587–1599. [Google Scholar] [CrossRef]
- Narita, Y.; Kuratani, S. Evolution of the Vertebral Formulae in Mammals: A Perspective on Developmental Constraints. J. Exp. Zool. Part B Mol. Dev. Evol. 2005, 304, 91–106. [Google Scholar] [CrossRef]
- Prothero, D.R. Evolutionary Transitions in the Fossil Record of Terrestrial Hoofed Mammals. Evol. Educ. Outreach 2009, 2, 289–302. [Google Scholar] [CrossRef] [Green Version]
- Wood, A.R.; Bebej, R.M.; Manz, C.L.; Begun, D.L.; Gingerich, P.D. Postcranial Functional Morphology of Hyracotherium (Equidae, Perissodactyla) and Locomotion in the Earliest Horses. J. Mamm. Evol. 2011, 18, 1–32. [Google Scholar] [CrossRef]
- May-Davis, S.; Hunter, R.; Brown, W. A Comparative Morphological Study of the Sixth and Seventh Spinous Processes of the Cervical Vertebrae in Extinct and Extant Species of Equus. Vertebr. Anat. Morphol. Palaeontol. 2021, 9, 40–51. [Google Scholar] [CrossRef]
- May-Davis, S.; Hunter, R.; White, R. Morphology of the Ventral Process of the Sixth Cervical Vertebra in Extinct and Extant Equus: Functional Implications. Animals 2023, 13, 1672. [Google Scholar] [CrossRef] [PubMed]
- Sisson, S. Osteology. In A Textbook of Veterinary Anatomy; W.B. Saunders Company: Philadelphia, PA, USA, 1910; pp. 31–35. [Google Scholar]
- Getty, R. Equine Osteology. In The Anatomy of the Domestic Animals, 5th ed.; Sisson, S., Grossman, J.D., Eds.; W.B. Saunders: Philadelphia, PA, USA, 1975; pp. 255–348. [Google Scholar]
- Denoix, J.-M.; Pailloux, J.-P. Anatomy and Basic Biomechanical Concepts. In Physical Therapy and Massage for the Horse, 2nd ed.; Trafalgar Square Publishing: North Pomfret, VT, USA, 2001; pp. 19–69. [Google Scholar]
- Bainbridge, D. The Normal Anatomy of the Neck. In Equine Neck and Back Pathology, 2nd ed.; Henson, F.M.D., Ed.; John Wiley and Sons Ltd.: Hoboken, UK, 2018; pp. 1–9. [Google Scholar]
- Popesko, P. Atlas of Topographical Anatomy of the Domestic Animals, 4th ed.; W.B. Saunders: Philadelphia, PA, USA, 1985; Volume 4, p. 185. [Google Scholar]
- Bradley, O. The Topical Anatomy of the Head and Neck of the Horse, 2nd ed.; Green and Sons: Edinburgh, UK, 1947. [Google Scholar]
- Gasse, H.; Van den Broeck, W.; Simeons, P. Nomina Anatomica Veterinaria, 6th ed.; World Association of Veterinary Anatomists: Hanover, Germany, 2017; p. 17. [Google Scholar]
- May-Davis, S. The Occurrence of a Congenital Malformation in the Sixth and Seventh Cervical Vertebrae Predominantly Observed in Thoroughbred Horses. J. Equine Vet. Sci. 2014, 18, 1313–1317. [Google Scholar] [CrossRef]
- DeRouen, A.; Spriet, M.; Aleman, M. Prevalence of Anatomical Variation of the Sixth Cervical Vertebra and Association with Vertebral Canal Stenosis and Articular Process Osteoarthritis in the Horse. Vet. Radiol. Ultrasound 2016, 57, 253–258. [Google Scholar] [CrossRef]
- Santinelli, I.; Beccati, F.; Arcelli, R. Anatomical Variation of the Spinous and Transverse Processes in the Caudal Cervical Vertebrae and the First Thoracic Vertebra in Horses. Equine Vet. J. 2016, 48, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Rombach, N.; Stubbs, N.; Clayton, H. Gross Anatomy of the Deep Perivertebral Musculature in Horses. Am. J. Vet. Res. 2014, 75, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Veraa, S.; Bergmann, W.; van den Belt, A.-J.; Wijnberg, I.; Back, W. Ex Vivo Computed Tomographic Evaluation of Morphology Variations in Equine Cervical Vertebrae. Vet. Radiol. Ultrasound 2016, 57, 482–488. [Google Scholar] [CrossRef] [PubMed]
- Veraa, S.; de Graaf, K.; Wijnberg, I.D.; Back, W.; Vernooij, H.; Nielen, M.; Belt, A.J. Caudal Cervical Vertebral Morphological Variation is not Associated with Clinical Signs in Warmblood Horses. Equine Vet. J. 2019, 52, 210–224. [Google Scholar] [CrossRef]
- Beccati, F.; Pepe, M.; Santinelli, I.; Gialletti, R.; Di Meo, A.; Romero, J.M. Radiographic Findings and Anatomical Variations of the Caudal Cervical Area in Horses with Neck Pain and Ataxia: Case–Control Study on 116 Horses. Vet. Rec. 2020, 187, e79. [Google Scholar] [CrossRef]
- May-Davis, S. Congenital Malformations of the 1st Sternal Rib. J. Equine Vet. Sci. 2014, 49, 92–100. [Google Scholar] [CrossRef]
- May-Davis, S.; Walker, C. Variations and Implications of the Gross Morphology in the Longus colli Muscle in Thoroughbred and Thoroughbred Derivative Horses Presenting with a Congenital Malformation of the Sixth and Seventh Cervical Vertebrae. J. Equine Vet. Sci. 2015, 35, 560–568. [Google Scholar] [CrossRef]
- Agerholm, J.S.; Bendixen, C.; Andersen, O.; Arnbjerg, J. Complex vertebral malformation in Holstein calves. J. Vet. Diagn. Invest. 2001, 13, 283–289. [Google Scholar] [CrossRef] [Green Version]
- Agerholm, J.S. Complex Vertebral Malformation Syndrome in Holstein cattle: The Story so Far. Acta Vet. Scand. 2007, 49, S5. [Google Scholar] [CrossRef] [Green Version]
- Thomsen, B.; Horn, P.; Panitz, F.; Bendixen, E.; Petersen, A.H.; Holm, L.-E.; Nielsen, V.H.; Agerholm, J.S.; Arnbjerg, J.; Bendixen, C. A Missense Mutation in the Bovine SLC35A3 Gene, Encoding a UDP-N-acetylglucosamine Transporter, causes Complex Vertebral Malformation. Genome Res. 2006, 16, 97–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 31 October 2022).
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.D.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. “Welcome to the Tidyverse”. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef] [Green Version]
- Daniel, D.S.; Whiting, K.; Curry, M.; Lavery, J.A.; Larmarange, J. Reproducible Summary Tables with the Gtsummary Package. R J. 2021, 13, 570–580. [Google Scholar] [CrossRef]
- Chauveau, A. The Comparative Anatomy of the Domesticated Animals, 2nd ed.; Fleming, G., Ed.; D. Appleton and Company: New York, NY, USA, 1908; pp. 53–80. [Google Scholar]
- Haussler, K.K.; Pool, R.R.; Clayton, H.M. Characterization of the Bony Changes Localized to the Cervical Articular Processes in a Mixed Population of Horse. PLoS ONE 2019, 14, e0222989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitwell, K.E.; Dyson, S. Interpreting Radiographs 8: Equine Cervical Vertebrae. Equine Vet. J. 1987, 19, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Gee, C.; Small, A.; Shorter, K.; Brown, W.Y. A Radiographic Technique for Assessment of Morphologic Variations of the Equine Caudal Cervical Spine. Animals 2020, 10, 667. [Google Scholar] [CrossRef] [Green Version]
- Shiratori, H.; Sakuma, R.; Watanabe, M.; Hashiguchi, H.; Mochida, L.; Sakai, Y.; Nishino, I.; Saijoh, Y.; Whitman, M.; Hamada, H. Two-Step Regulation of Left–Right Asymmetric Expression of Pitx2: Initiation by Nodal Signaling and Maintenance by Nkx2. Mol. Cell 2001, 7, 137–149. [Google Scholar] [CrossRef]
- Hamada, H.; Meno, C.; Watanabe, D.; Saijoh, Y. Establishment of Vertebrate Left–right Asymmetry. Nat. Rev. 2002, 3, 103–112. [Google Scholar] [CrossRef]
- Blum, M.; Ott, T. Animal Left–right Asymmetry. Curr. Biol. 2018, 28, R293–R305. [Google Scholar] [CrossRef] [Green Version]
- Witzmann, F.; Haridy, Y.; Hilger, A.; Manke, I.; Asbach, P. Rarity of Congenital Malformation and Deformity in the Fossil Record 474 of Vertebrates–A Non-Human Perspective. Int. J. Paleopathol. 2021, 33, 30–42. [Google Scholar] [CrossRef] [PubMed]
- Larson, G.; Fuller, D.Q. The Evolution of Animal Domestication. Annu. Rev. Ecol. Evol. Syst. 2014, 45, 115–136. [Google Scholar] [CrossRef] [Green Version]
- Schubert, M.; Jónsson, H.; Chang, D.; Sarkissian, C.D.; Ermini, L.; Ginolhac, A.; Albrechtsen, A.; Dupanloup, I.; Foucal, A.; Petersen, B.; et al. Prehistoric Genomes Reveal the Genetic Foundation and Cost of Horse Domestication. Proc. Natl. Acad. Sci. USA 2014, 111, E5661–E5669. [Google Scholar] [CrossRef] [PubMed]
- Fages, A.; Hanghøj, K.; Khan, N.; Gaunitz, C.; Sequin-Orlando, A.; Leonardi, M.; Constantz, C.M.; Gamba, C.; Al-Rasheid, K.A.S.; Albizuir, S.; et al. Tracking Five Millennia of Horse Management with Extensive Ancient Genome Time Series. Cell 2019, 177, 1419–1435. [Google Scholar] [CrossRef]
- Orlando, L.; Linbrado, P. Origin and Evolution of Deleterious Mutations in Horses. Genes 2019, 10, 649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valberg, S. Genetics of Equine Muscle Disease. Vet. Clin. N. Am. 2020, 36, 353–378. [Google Scholar] [CrossRef]
- Sandmeyer, L.S.; Grahn, B.H.; Breaux, C. Diagnostic Ophthalmology. Can. Vet. J. 2006, 47, 1131–1133. [Google Scholar] [PubMed]
- Figueirido, B.; Pérez-Ramos, A.; Martín-Serra, A. Intravertebral vs. Intervertebral Integration and Modularity in the Vertebral Column of Mammalian Carnivorans. J. Anat. 2022, 4, 642–656. [Google Scholar] [CrossRef] [PubMed]
- Randau, M.; Goswani, A. Morphological Modularity in the Vertebral Column of Felidae (Mammalia, Carnivora). BMC Evol. Biol. 2017, 17, 133. [Google Scholar] [CrossRef] [Green Version]
- Böhmer, C.; Amson, E.; Arnold, P.; van Heteren, A.H.; Nyakatura, J.A. Homeotic Transformations Reflect Departure from the Mammalian ‘Rule of Seven’ Cervical Vertebrae in Sloths: Inferences on the Hox Code and Morphological Modularity of the Mammalian Neck. BMC Evol. Biol. 2018, 18, 84. [Google Scholar] [CrossRef] [Green Version]
- Arnold, P.; Forterre, F.; Lang, J.; Fischer, M.S. Morphological Disparity, Conservatism, and Integration in the Canine Lower Cervical Spine: Insights into Mammalian Neck Function and Regionalization. Mamm. Biol. 2016, 81, 153–162. [Google Scholar] [CrossRef]
- Arnold, P.; Esteve-Altava, B.; Fischer, M.S. Musculoskeletal Networks reveal Topological Disparity in Mammalian Neck Evolution. BMC Evol. Biol. 2017, 17, 251. [Google Scholar] [CrossRef] [Green Version]
- Heude, E.; Tesarova, M.; Sefton, E.M.; Jullian, E.; Adachi, N.; Grimaldi, A.; Zikmund, T.; Kaiser, J.; Kardon, G.; Kelly, R.G.; et al. Unique Morphogenetic Signatures Define Mammalian Neck Muscles and Associated Connective Tissues. eLife 2018, 7, e40179. [Google Scholar] [CrossRef]
- Zsoldos, R.R.; Groesel, M.; Kotschwar, A.; Kotschwar, A.B.; Licka, T.; Peham, C. A Preliminary Modelling Study on the Equine Cervical Spine with Inverse Kinematics at Walk. Equine Vet. J. 2010, 38, 516–522. [Google Scholar] [CrossRef]
- Vidal, P.P.; Graf, W.; Berthoz, A. The Orientation of the Cervical Vertebral Column in Unrestrained Awake Animals. Exp. Brain Res. 1986, 61, 549–559. [Google Scholar] [CrossRef] [PubMed]
- Graf, W.; de Waele, C.; Vidal, P.; Wang, D.; Evinger, C. The Orientation of the Cervical Vertebral Column in Unrestrained Awake Animals. II. Movement Strategies. Brain Behav. Evol. 1995, 45, 209–231. [Google Scholar] [CrossRef]
- Kyriazis, C.C.; Wayne, R.K.; Lohmueller, K.E. Strongly Deleterious Mutations are a Primary Determinant of Extinction Risk due to Inbreeding Depression. Evol. Lett. 2020, 5, 33–47. [Google Scholar] [CrossRef]
- Thirstrup, J.P.; Bach, L.A.; Loeschcke, V.; Pertoldi, C. Population Viability Analysis on Domestic Horse Breeds (Equus caballus). J. Anim. Sci. 2009, 87, 3525–3535. [Google Scholar] [CrossRef] [PubMed]
- Kessel, M.; Gruss, P. Homeotic Transformations of Murine Vertebrae and Concomitant Alteration of Hox Codes Induced by Retinoic Acid. Cell 1991, 67, 89–104. [Google Scholar] [CrossRef] [Green Version]
- Krumlauf, R. Hox Genes in Vertebrate Development. Cell 1994, 78, 191–201. [Google Scholar] [CrossRef]
- Cordes, R.; Schuster-Gossler, K.; Sereh, K.; Gossler, A. Specification of Vertebral Identity is Coupled to Notch Signaling and the Segmentation Clock. Development 2004, 131, 1221–1233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mallo, M. Of Necks, Trunks and Tails: Axial Skeletal Diversity among Vertebrates. Diversity 2021, 13, 289. [Google Scholar] [CrossRef]
- Kostic, D.; Capecchi, M.R. Targeted Disruptions of the Murine Hoxa-4 and Hoxa-6 Genes Result in Homeotic Transformations of Components of the Vertebral Column. Mech. Dev. 1994, 46, 231–247. [Google Scholar] [CrossRef]
- Hubert, K.; Wellik, D.M. Hox Genes in Development and Beyond. Development 2023, 150, dev192476. [Google Scholar] [CrossRef] [PubMed]
- Rancourt, D.E.; Suzuki, T.; Capecchi, M.R. Genetic Interaction between Hoxb-5 and Hoxb-6 is Revealed by Nonallelic Noncomplementation. Genes Dev. 1995, 9, 108–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horan, G.S.; Ramírez-Solis, R.; Featherstone, M.S.; Wolgemuth, D.J.; Bradley, A.; Berhringer, R.R. Compound Mutants for the Paralogous Hoxa-4, Hoxb-4 and Hoxd-4 show More Complete Homeotic Transformations and a Dose-dependent Increase in the Number of Vertebrae Transformed. Genes Dev. 1995, 9, 1667–1677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aubin, J.; Lemieux, M.; Tremblay, M.; Behringer, R.R.; Jeannotte, L. Transcriptional Interferences at the Hoxa4/Hoxa5 Locus: Importance of Correct Hoxa5 Expression for the Proper Specification of the Axial Skeleton. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 1998, 212, 141–156. [Google Scholar] [CrossRef]
- Jeannotte, L.; Gotti, F.; Landry-Truchon, K. Hoxa-5: A Key Player in Development and Disease. J. Dev. Biol. 2016, 4, 13. [Google Scholar] [CrossRef] [Green Version]



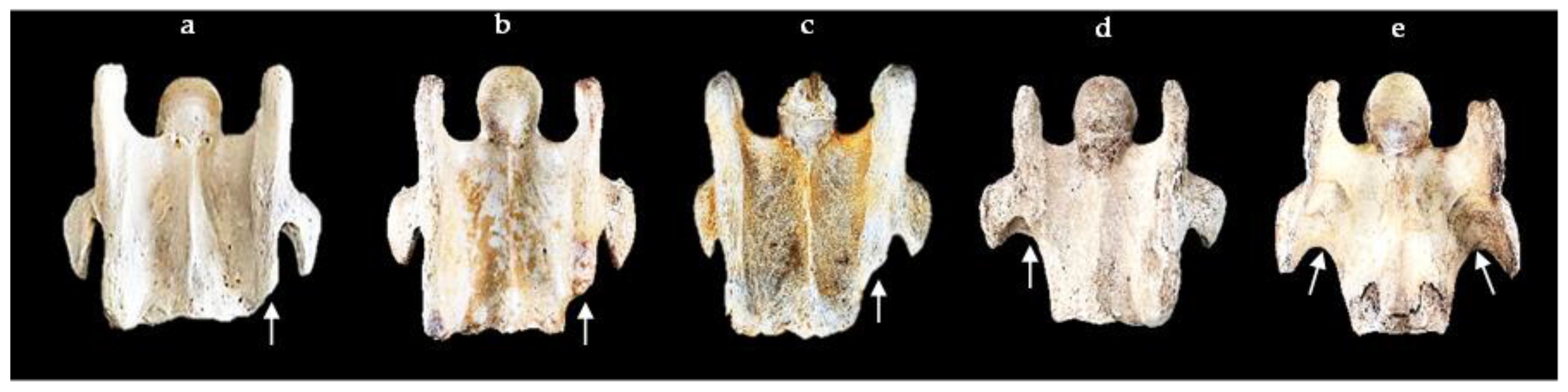





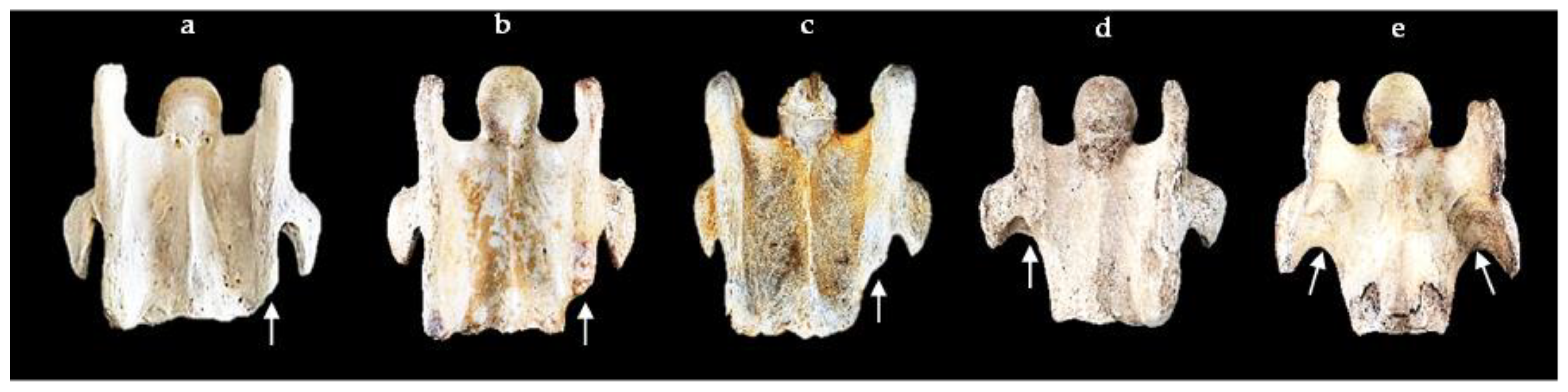
{kind=link}
{kind=link}
{kind=link}
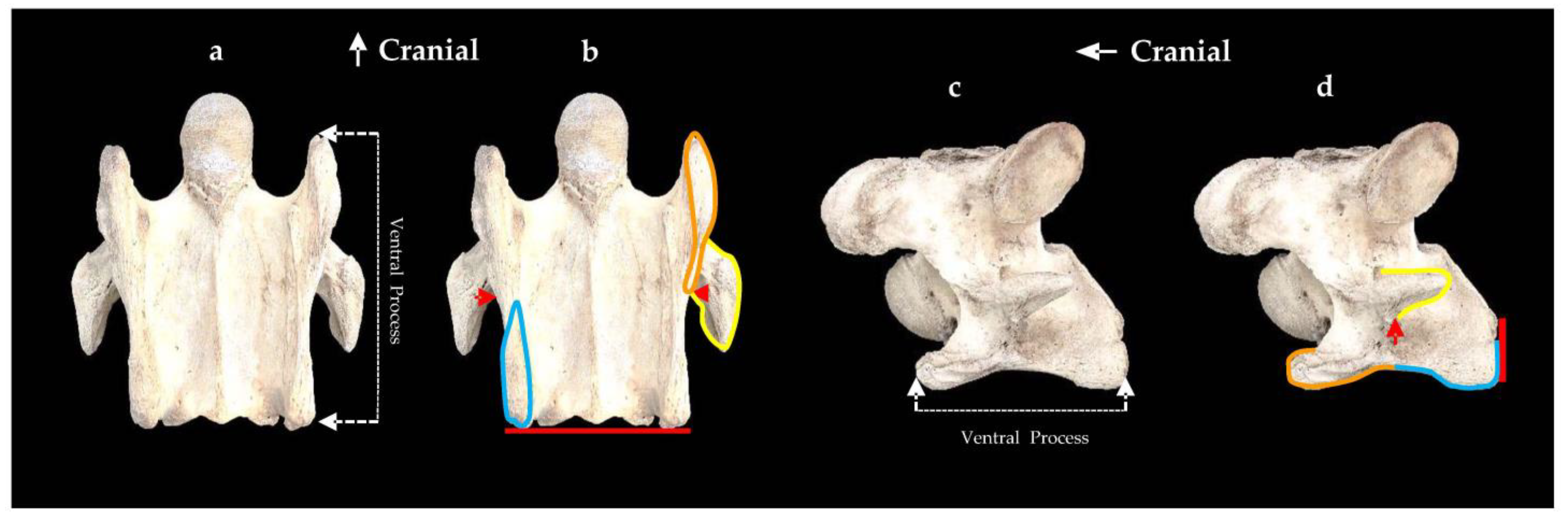
{kind=link}
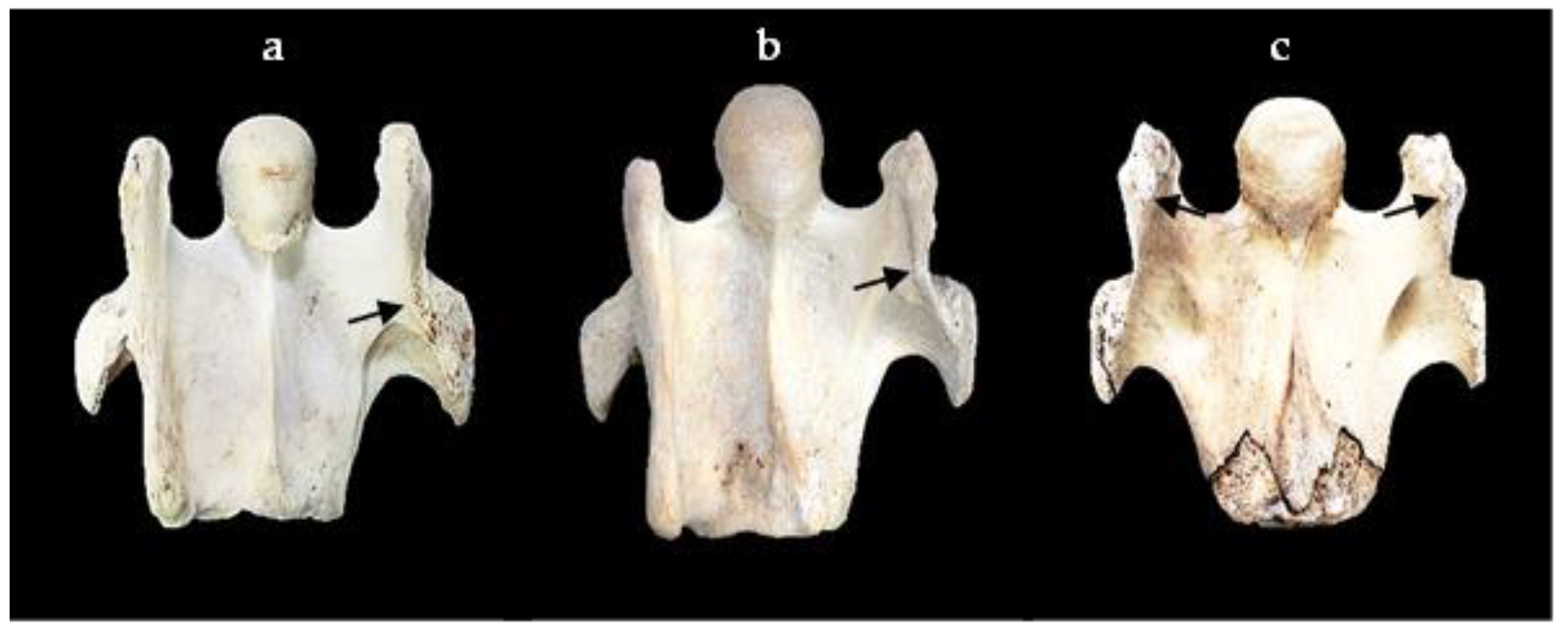{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Laterality | Grade | aCVT Left | aCVT Right | aCrVT Left | aCrVT Right |
|---|---|---|---|---|---|
| Unilateral | 1/4 | 3 | 1 | 4 | |
| 2/4 | 5 | 4 | 14 | 2 | |
| 3/4 | 5 | 1 | 2 | ||
| 4/4 | 22 | 6 | |||
| Bilateral | 1/4 | 2 | 7 | 6 | |
| 2/4 | 1 | 8 | 6 | ||
| 3/4 | 1 | 1 | 4 | 3 | |
| 4/4 | 27 | 26 |
| Characteristic | Bilateral, n = 29 | Unilateral Left n = 35 | Unilateral Right n = 12 | p-Value |
|---|---|---|---|---|
| Origin | 0.0787 1 | |||
| Asia | 0 (0%) | 3 (8.6%) | 1 (8.3%) | |
| Australasia | 18 (62%) | 23 (66%) | 6 (50%) | |
| Europe | 2 (6.9%) | 2 (5.7%) | 4 (33%) | |
| North America | 9 (31%) | 7 (20%) | 1 (8.3%) | |
| Breed (purpose) | 0.3970 2 | |||
| Racing | 15 (52%) | 24 (69%) | 7 (58%) | |
| Riding | 14 (48%) | 11 (31%) | 5 (42%) | |
| Average Age | 11 | 12 | 11 | 0.8023 3 |
| Sex | 0.9508 2 | |||
| F | 12 (41%) | 13 (37%) | 5 (42%) | |
| M | 17 (59%) | 22 (63%) | 7 (58%) |
| Grade (0–4) | 0/4 (%) | 1/4 (%) | 2/4 (%) | 3/4 (%) | 4/4 (%) | Total |
|---|---|---|---|---|---|---|
| Left aCVT | 12 (15%) | 3 (4%) | 6 (8%) | 6 (8%) | 49 (65%) | 76 |
| Right aCVT | 35 (46%) | 2 (4%) | 4 (4.5%) | 4 (4.5%) | 31 (41%) | 76 |
| Total | 47 (30.5%) | 5 (4%) | 10 (6.25%) | 10 (6.25%) | 80 (53%) | 152 |
| Left aCVT Grade (n) | 0/4 n = 12 | 1/4 n= 3 | 2/4 n= 6 | 3/4 n= 6 | 4/4 n= 49 |
| Left aCrVT Grade | |||||
| 0/4 | 12 (100%) | 3 (100%) | 6 (100%) | 6 (100%) | 12 (24%) |
| 1/4 | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) | 9 (18%) |
| 2/4 | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) | 24 (49%) |
| 3/4 | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) | 4 (8.2%) |
| Right aCVT Grade | 0/4 n = 35 | 1/4 n= 2 | 2/4 n= 4 | 3/4 n= 4 | 4/4 n= 31 |
| Right aCrVT Grade | |||||
| 0/4 | 35 (100%) | 2 (100%) | 4 (100%) | 4 (100%) | 14 (45%) |
| 1/4 | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) | 6 (19%) |
| 2/4 | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) | 9 (29%) |
| 3/4 | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) | 2 (6.5%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
May-Davis, S.; Dzingle, D.; Saber, E.; Blades Eckelbarger, P. Characterization of the Caudal Ventral Tubercle in the Sixth Cervical Vertebra in Modern Equus ferus caballus. Animals 2023, 13, 2384. https://doi.org/10.3390/ani13142384
May-Davis S, Dzingle D, Saber E, Blades Eckelbarger P. Characterization of the Caudal Ventral Tubercle in the Sixth Cervical Vertebra in Modern Equus ferus caballus. Animals. 2023; 13(14):2384. https://doi.org/10.3390/ani13142384
Chicago/Turabian StyleMay-Davis, Sharon, Diane Dzingle, Elle Saber, and Pamela Blades Eckelbarger. 2023. "Characterization of the Caudal Ventral Tubercle in the Sixth Cervical Vertebra in Modern Equus ferus caballus" Animals 13, no. 14: 2384. https://doi.org/10.3390/ani13142384