

Comparison of Cortisol Concentrations in Different Matrices in Alpine Ibex (Capra ibex) at the Zoo
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Collection of Samples
2.2. Laboratory Tests
2.3. Data Analysis
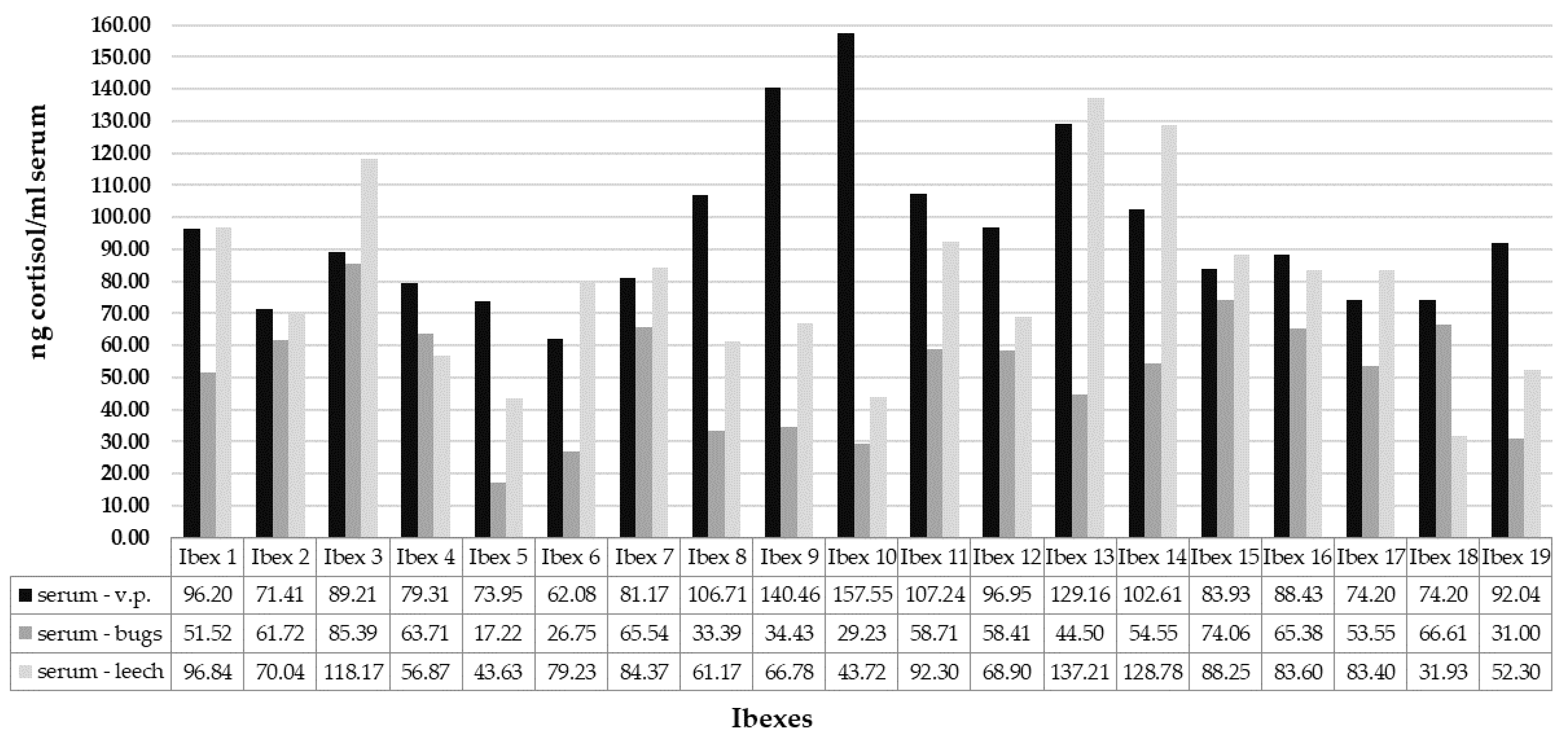
3. Results
4. Discussion
4.1. Blood Collection
4.2. Saliva, Faeces, and Hair Collection
4.3. Tear and Urine Collection
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grossen, C.; Biebach, I.; Angelone-Alasaad, S.; Keller, L.F.; Croll, D. Population genomics analyses of European ibex species show lower diversity and higher inbreeding in reintroduced populations. Evol. Appl. 2018, 11, 123–139. [Google Scholar] [CrossRef] [Green Version]
- Stüwe, M.; Nievergelt, B. Recovery of alpine ibex from near extinction: The result of effective protection, captive breeding, and reintroductions. Appl. Anim. Behav. Sci. 1991, 29, 379–387. [Google Scholar] [CrossRef]
- Parrini, F.; Cain, J.W., III; Krausman, P.R. Capra ibex (Artiodactyla: Bovidae). Mamm. Species 2009, 830, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Espinosa, J.; López-Olvera, J.R.; Cano-Manuel, F.J.; Fandos, P.; Pérez, J.M.; López-Graells, C.; Ráez-Bravo, A.; Mentaberre, G.; Romero, D.; Soriguer, R.C.; et al. Guidelines for managing captive Iberian ibex herds for conservation purposes. J. Nat. Conserv. 2017, 40, 24–32. [Google Scholar] [CrossRef]
- Gilbert, T.; Gardner, R.; Kraaijeveld, A.R.; Riordan, P. Contributions of zoos and aquariums to reintroductions: Historical reintroduction efforts in the context of changing conservation perspectives. Int. Zoo Yearb. 2017, 51, 15–31. [Google Scholar] [CrossRef]
- Zooinstitutes. Alpine Ibex/Capra Ibex at Zoos Worldwide on Zooinstitutes. Available online: https://zooinstitutes.com/animals/alpine-ibex-334/ (accessed on 31 July 2023).
- Gardocka, T.; Gruszczynska, A.; Maslak, R.; Sergiel, A. The Welfare of Animals in Zoos and EU Legal Standards, 1st ed.; Elipsa: Warsaw, Poland, 2014; p. 172. [Google Scholar]
- Wolfensohn, S.; Shotton, J.; Bowley, H.; Davies, S.; Thompson, S.; Justice, W.S.M. Assessment of welfare in zoo animals: Towards optimum quality of life. Animals 2018, 8, 110. [Google Scholar] [CrossRef] [Green Version]
- Schilling, A.K.; Mazzamuto, M.V.; Romeo, C. A review of non-Invasive sampling in wildlife disease and health research: What’s new? Animals 2022, 12, 1719. [Google Scholar] [CrossRef] [PubMed]
- Kastelic, M.; Pšeničnik, I.; Gregurić Gračner, G.; Čebulj Kadunc, N.; Lindtner Knific, R.; Slavec, B.; Krapež, U.; Vergles Rataj, A.; Zorman Rojs, O.; Pulko, B.; et al. Health status and stress in different categories of racing pigeons. Animals 2021, 11, 2686. [Google Scholar] [CrossRef] [PubMed]
- Ganswindt, A.; Brown, J.L.; Freeman, E.W.; Kouba, A.J.; Penfold, L.M.; Santymire, R.M.; Vick, M.M.; Wielebnowski, N.; Willis, E.L.; Milnes, M.R. International society for wildlife endocrinology: The future of endocrine measures for reproductive science, animal welfare and conservation biology. Biol. Lett. 2012, 8, 695–697. [Google Scholar] [CrossRef] [Green Version]
- Stewart, M.; Webster, J.; Schaefer, A.; Cook, N.; Scott, S. Infrared thermography as a non-invasive tool to study animal welfare. Anim. Welf. 2005, 14, 319–325. [Google Scholar] [CrossRef]
- Schaub, G.A.; Lawrenz, A.; Stadler, A. “Living Syringes”: Use of hematophagous bugs as blood samplers from small and wild animals. In Nature Helps… Parasitology Research Monographs; Springer: Berlin/Heidelberg, Germany, 2012; Volume 1, pp. 243–271. [Google Scholar] [CrossRef]
- Kruszewicz, A.; Grothmann, P.; Czujkowska, A.; Stadler, A.; Lawrenz, A.; Schaub, G. Wykorzystanie krwiopijnych pluskwiaków do pobierania próbek krwi od zwierząt egzotycznych—Use of kissing bugs for blood sampling of exotic animals. Życie Wet. 2009, 84, 405–407. [Google Scholar]
- Kvapil, P.; Tomašek, O.; Bartova, E.; Harej, M.; Kastelic, M.; Primožič, T.; Kašparkova, N.; Račnik, J. Validation of Medicinal leeches (Hirudo medicinalis) as a non-invasive blood sampling tool for hematology and biochemistry profiling in mammals. Front. Vet. Sci. 2022, 9, 831836. [Google Scholar] [CrossRef] [PubMed]
- Wagner, R.; Fieseler, H.; Kaiser, M.; Muller, H.; Mielenz, N.; Spilke, J.; Gottschalk, J.; Einspanier, A.; Palme, R.; Rizk, A.; et al. Cortisol concentrations in sheep before, during and after sham foot trimming on a tilt table—The suitability of different matrices. Schweiz. Arch. Fur. Tierheilkd. 2021, 164, 753–766. [Google Scholar] [CrossRef] [PubMed]
- Palme, R.; Rettenbacher, S.; Touma, C.; El-Bahr, S.M.; Mostl, E. Stress hormones in mammals and birds: Comparative aspects regarding metabolism, excretion, and noninvasive measurement in fecal samples. Ann. N. Y. Acad. Sci. 2005, 1040, 162–171. [Google Scholar] [CrossRef]
- Prandi, A.; Peric, T.; Corazzin, M.; Comin, A.; Colitti, M. A first survey on hair cortisol of an Alpine ibex (Capra ibex ibex) population. Anim. Sci. Pap. Rep. 2018, 36, 57–74. [Google Scholar]
- Touma, C.; Palme, R. Measuring fecal glucocorticoid metabolites in mammals and birds: The importance of validation. Ann. N. Y. Acad. Sci. 2005, 1046, 54–74. [Google Scholar] [CrossRef] [Green Version]
- Cook, N.J. Review: Minimally invasive sampling media and the measurement of corticosteroids as biomarkers of stress in animals. Can. J. Anim. Sci. 2012, 92, 227–259. [Google Scholar] [CrossRef]
- Jewgenow, K.; Azevedo, A.; Albrecht, M.; Kirschbaum, C.; Dehnhard, M. Hair cortisol analyses in different mammal species: Choosing the wrong assay may lead to erroneous results. Conserv. Physiol. 2020, 8, coaa009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mostl, E.; Maggs, J.L.; Schrotter, G.; Besenfelder, U.; Palme, R. Measurement of cortisol metabolites in faeces of ruminants. Vet. Res. Commun. 2002, 26, 127–139. [Google Scholar] [CrossRef]
- Sgorlon, S.; Fanzago, M.; Guiatti, D.; Gabai, G.; Stradaioli, G.; Stefanon, B. Factors affecting milk cortisol in mid lactating dairy cows. BMC Vet. Res. 2015, 11, 259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verkerk, G.; Phipps, A.; Carragher, J.; Matthews, L.; Stelwagen, K. Characterization of milk cortisol concentrations as a measure of short-term stress responses in lactating dairy cows. Anim. Welf. 1998, 7, 77–86. [Google Scholar] [CrossRef]
- Monk, C.S.; Hart, K.A.; Berghaus, R.D.; Norton, N.A.; Moore, P.A.; Myrna, K.E. Detection of endogenous cortisol in equine tears and blood at rest and after simulated stress. Vet. Ophthalmol. 2014, 17, 53–60. [Google Scholar] [CrossRef]
- Vincent, I.C.; Michell, A.R. Comparison of cortisol concentrations in saliva and plasma of dogs. Res. Vet. Sci. 1992, 53, 342–345. [Google Scholar] [CrossRef] [PubMed]
- Weaver, S.J.; Hynd, P.I.; Ralph, C.R.; Hocking Edwards, J.E.; Burnard, C.L.; Narayan, E.; Tilbrook, A.J. Chronic elevation of plasma cortisol causes differential expression of predominating glucocorticoid in plasma, saliva, fecal, and wool matrices in sheep. Domest. Anim. Endocrinol. 2021, 74, 106503. [Google Scholar] [CrossRef] [PubMed]
- Brivio, F.; Grignolio, S.; Sica, N.; Cerise, S.; Bassano, B. Assessing the Impact of Capture on Wild Animals: The Case Study of Chemical Immobilisation on Alpine Ibex. PLoS ONE 2015, 10, e0130957. [Google Scholar] [CrossRef] [PubMed]
- Kvapil, P.; Kastelic, M.; Jež, N.; Sedlák, K.; Kašpárková, N.; Jelovšek, M.; Avšič-Županc, T.; Bártová, E.; Račnik, J. Detection of antibodies against tick-borne encephalitis virus in zoo animals using non-invasive blood sampling with Medicinal leeches (Hirudo medicinalis). Pathogens 2021, 10, 952. [Google Scholar] [CrossRef]
- Schwarzenberger, F.; Möstl, E.; Palme, R.; Bamberg, E. Faecal steroid analysis for non-invasive monitoring of reproductive status in farm, wild and zoo animals. Anim. Reprod. Sci. 1996, 42, 515–526. [Google Scholar] [CrossRef]
- Colding-Jorgensen, P.; Hestehave, S.; Abelson, K.S.P.; Kalliokoski, O. Hair glucocorticoids are not a historical marker of stress—Exploring the time-scale of corticosterone incorporation into hairs in a rat model. Gen. Comp. Endocrinol. 2023, 341, 114335. [Google Scholar] [CrossRef]
- Plut, J.; Snoj, T.; Golinar Oven, I.; Štukelj, M. The Combination of Serum and Oral Fluid Cortisol Levels and Welfare Quality Protocol® for Assessment of Pig Welfare on Intensive Farms. Agriculture 2023, 13, 351. [Google Scholar]
- Cansu Karaer, M.; Čebulj-Kadunc, N.; Snoj, T. Stress in wildlife: Comparison of the stress response among domestic, captive, and free-ranging animals. Front. Vet. Sci. 2023, 10, 1167016. [Google Scholar] [CrossRef] [PubMed]
- Umeda, T.; Hiramatsu, R.; Iwaoka, T.; Shimada, T.; Miura, F.; Sato, T. Use of saliva for monitoring unbound free cortisol levels in serum. Clin. Chim. Acta. 1981, 110, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Perogamvros, I.; Keevil, B.G.; Ray, D.W.; Trainer, P.J. Salivary Cortisone Is a Potential Biomarker for Serum Free Cortisol. J. Clin. Endocrinol. Metab. 2010, 95, 4951–4958. [Google Scholar] [CrossRef]
- Gundlach, N.H.; Piechotta, M.; Siebert, U. Is lachrymal fluid a potential method for cortisol measurement in wild harbor seals? A pilot study. J. Vet. Med. Res. 2014, 2016, 967043. [Google Scholar] [CrossRef] [Green Version]
- Kvapil, P.; Pirš, T.; Slavec, B.; Luštrik, R.; Zemljič, T.; Bartova, E.; Stranjac, B.; Kastelic, M. Tear production, intraocular pressure and conjunctival bacterial flora in selected captive wild ruminants. Vet. Ophthalmol. 2018, 21, 52–57. [Google Scholar] [CrossRef] [Green Version]
- Hill, S.P.; Broom, D.M. Measuring zoo animal welfare: Theory and practice. Zoo. Biol. 2009, 28, 531–544. [Google Scholar] [CrossRef]
- Melfi, V.A.; Ward, S.J. Welfare Implications of Zoo Animal Training. In Zoo Animal Learning and Training; John Wiley & Sons Ltd.: Chichester, UK, 2020; pp. 271–288. [Google Scholar] [CrossRef]
- Markvardsen, S.N.; Kjelgaard-Hansen, M.; Ritz, C.; Sorensen, D.B. Less invasive blood sampling in the animal laboratory: Clinical chemistry and haematology of blood obtained by the Triatominae bug Dipetalogaster maximus. Lab. Anim. 2012, 46, 136–141. [Google Scholar] [CrossRef] [Green Version]
- Legislation. Council Directive 92/65/EEC of 13 July 1992 Laying down Animal Health Requirements Governing Trade in and Imports into the Community of Animals, Semen, ova and Embryos Not Subject to Animal Health Requirements Laid down in Specific Community Rules Referred to in Annex A (I) to Directive 90/425/EEC (BALAI). 1992. Available online: https://eur-lex.europa.eu/eli/dir/1992/65/oj (accessed on 31 July 2023).
- Legislation. Regulation (EU) 2016/429 of the European Parliament and of the Council of 9 March 2016 on Transmissible Animal Diseases and Amending and Repealing Certain Acts in the Area of Animal Health (‘Animal Health Law’) (Text with EEA Relevance). 2016. Available online: https://eur-lex.europa.eu/eli/reg/2016/429/oj (accessed on 31 July 2023).
- Čebulj-Kadunc, N.; Frangež, R.; Kruljc, P. Fluctuations of physiological variables during conditioning of Lipizzan fillies before starting under saddle. Animals 2022, 12, 836. [Google Scholar] [CrossRef] [PubMed]
- Nedić, S.; Pantelić, M.; Vranjes-Durić, S.; Nedic, D.; Jovanović, L.; Čebulj-Kadunc, N.; Kobal, S.; Snoj, T.; Kirovski, D. Cortisol concentrations in hair, blood and milk of Holstein and Busha cattle. Slov. Vet. Res. 2017, 54, 163–172. [Google Scholar] [CrossRef] [Green Version]
- Kvapil, P.; Kastelic, M.; Dovč, A.; Štrus, Š.; Bartova, E. Five-year survey of the intestinal parasites in reptiles, ratites, hoofed mammals, carnivores and primates in ZOO Ljubljana. In Proceedings of the International Conference on Diseases of Zoo and Wild Animals 2015, Barcelona, Spain, 13–16 May 2015; pp. 146–149. [Google Scholar]
- Cook, N.J.; Schaefer, A.L.; Lepage, P.; Morgan Jones, S.D. Radioimmunoassay for Cortisol in Pig Saliva and Serum. J. Agric. Food Chem. 1997, 45, 395–399. [Google Scholar] [CrossRef]
- Gold, A.J.; Langlois, D.K.; Refsal, K.R. Evaluation of Basal Serum or Plasma Cortisol Concentrations for the Diagnosis of Hypoadrenocorticism in Dogs. J. Vet. Intern. Med. 2016, 30, 1798–1805. [Google Scholar] [CrossRef]
- Seth, J.; Brown, L.M. A simple radioimmunoassay for plasma cortisol. Clinica Chimica Acta 1978, 86, 109–120. [Google Scholar] [CrossRef]
- Sica, N. Dealing with Stress: Responses for a Good Fitness on Alpine Ibex. Ph.D. Thesis, University of Sassarit, School in Natural Sciences, Sassari, Italy, 2014. [Google Scholar]
- Posautz, C. Measurement of Glucocorticoid Metabolites in Feces of Capricorns (Alpine ibex); University of Veterinary Medicine: Vienna, Austria, 2012. [Google Scholar]
- Sartorelli, P.; Agnes, F.; Lanfranchi, P. Pathophysiological significance of hematochemical parameters of Capra ibex. Hystrix Ital. J. Mammal. 1997, 9, 39–44. [Google Scholar] [CrossRef]
- Gómez-Guillamón, F.; Caballero-Gómez, J.; Agüero, M.; Camacho-Sillero, L.; Risalde, M.A.; Zorrilla, I.; Villalba, R.; Rivero-Juárez, A.; García-Bocanegra, I. Re-emergence of bluetongue virus serotype 4 in Iberian ibex (Capra pyrenaica) and sympatric livestock in Spain, 2018–2019. Transbound Emerg. Dis. 2021, 68, 458–466. [Google Scholar] [CrossRef]
- Aurich, J.; Wulf, M.; Ille, N.; Erber, R.; von Lewinski, M.; Palme, R.; Aurich, C. Effects of season, age, sex, and housing on salivary cortisol concentrations in horses. Domest. Anim. Endocrinol. 2015, 52, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Shutt, K.; Heistermann, M.; Kasim, A.; Todd, A.; Kalousova, B.; Profosouva, I.; Petrzelkova, K.; Fuh, T.; Dicky, J.-F.; Bopalanzognako, J.-B.; et al. Effects of habituation, research and ecotourism on faecal glucocorticoid metabolites in wild western lowland gorillas: Implications for conservation management. Biol. Conserv. 2014, 172, 72–79. [Google Scholar] [CrossRef] [Green Version]
- Palme, R. Non-invasive measurement of glucocorticoids: Advances and problems. Physiol. Behav. 2019, 199, 229–243. [Google Scholar] [CrossRef]
- Russell, E.; Koren, G.; Rieder, M.; Van Uum, S. Hair cortisol as a biological marker of chronic stress: Current status, future directions and unanswered questions. Psychoneuroendocrinology 2012, 37, 589–601. [Google Scholar] [CrossRef]
- Dulude-de Broin, F.; Côté, S.D.; Whiteside, D.P.; Mastromonaco, G.F. Faecal metabolites and hair cortisol as biological markers of HPA-axis activity in the Rocky mountain goat. Gen. Comp. Endocrinol. 2019, 280, 147–157. [Google Scholar] [CrossRef] [PubMed]
- El-Farhan, N.; Rees, D.A.; Evans, C. Measuring cortisol in serum, urine and saliva—Are our assays good enough? Ann. Clin. Biochem. 2017, 54, 308–322. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
| Year of Sampling | 2020 | 2020 | 2021 | 2022 |
|---|---|---|---|---|
| July August | September October | July August | July August | |
| Serum (V) | 10 | 22 | 12 | 12 |
| Serum (K) | 8 | 10 | 1 | 10 |
| Serum (L) | 1 | 10 | 4 | 8 |
| Saliva | / | 18 | 11 | / |
| Hair | 10 | 21 | 12 | / |
| Faeces | 19 | 19 | 5 | / |
| Urine | / | / | 6 | / |
| Tears | / | / | / | 2 |
| Abbreviation | |
|---|---|
| Minimum value | min |
| Maximum value | max |
| Mean value | M |
| Standard error | SE |
| Standard deviation | SD |
| Kruskal–Wallis test | Hi2 |
| Standardised score of Z-test | Z |
| Statistical significance | p |
| Degrees of freedom | df |
| Effect size | r |
| Number of tested samples | N |
| Total | Serum—V (ng/mL) | Serum—K (ng/mL) | Serum—L (ng/mL) | Saliva (ng/mL) | Faeces (ng/g) | Hair (ng/g) |
|---|---|---|---|---|---|---|
| Mean | 93.49 | 52.86 | 77.04 | 12.63 | 22.59 | 27.40 |
| SE | 3.45 | 4.29 | 5.50 | 1.14 | 1.53 | 1.92 |
| SD | 25.85 | 23.08 | 26.37 | 6.16 | 10.04 | 12.59 |
| N | 56 | 29 | 23 | 29 | 43 | 43 |
| Matrices | Sex | Mann–Whitney | Effect Size | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Males | Females | ||||||||
| M | SD | N | M | SD | N | Z | p | r | |
| Serum (V) | 89.71 | 24.03 | 25 | 96.53 | 27.23 | 31 | −1.39 | 0.164 | −0.19 |
| Saliva | 10.12 | 6.31 | 15 | 15.32 | 4.86 | 14 | −2.27 | 0.023 | −0.42 |
| Faeces | 23.66 | 9.88 | 22 | 21.47 | 10.33 | 21 | −0.91 | 0.362 | −0.14 |
| Hair | 25.98 | 11.05 | 20 | 28.64 | 13.92 | 23 | −0.44 | 0.661 | −0.07 |
| Matrices | Age | Kruskal–Wallis Test | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ten Months and Younger | Ten Months to Ten Years | Older than Ten Years | ||||||||||
| M | SD | N | M | SD | N | M | SD | N | Hi2 | df | p | |
| Serum (V) | 78.08 | 19.23 | 16 | 94.26 | 24.32 | 27 | 110.85 | 25.96 | 13 | 12.94 | 2 | 0.002 |
| Saliva | 15.14 | 6.01 | 10 | 11.71 | 5.17 | 14 | 10.20 | 8.43 | 5 | 2.96 | 2 | 0.228 |
| Faeces | 17.56 | 7.96 | 11 | 23.09 | 9.30 | 20 | 26.38 | 11.69 | 12 | 4.88 | 2 | 0.087 |
| Hair | 31.05 | 15.18 | 13 | 25.64 | 11.95 | 19 | 26.14 | 10.37 | 11 | 1.21 | 2 | 0.546 |
| Matrices | Period | Mann–Whitney | Effect Size | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Summer | Autumn | ||||||||
| M | SD | N | M | SD | N | Z | p | r | |
| Serum (V) | 88.43 | 19.55 | 34 | 101.30 | 32.30 | 22 | −1.80 | 0.073 | −0.24 |
| Saliva | 12.61 | 6.82 | 11 | 12.64 | 5.92 | 18 | 0.02 | 0.982 | <0.01 |
| Faeces | 23.18 | 9.53 | 24 | 21.86 | 10.88 | 19 | −0.43 | 0.668 | −0.07 |
| Hair | 19.24 | 7.90 | 22 | 35.95 | 10.83 | 21 | −4.52 | <0.001 | −0.69 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kastelic, M.; Gregurić Gračner, G.; Tomažič, I.; Kvapil, P.; Harej, M.; Dovč, A. Comparison of Cortisol Concentrations in Different Matrices in Alpine Ibex (Capra ibex) at the Zoo. Animals 2023, 13, 2491. https://doi.org/10.3390/ani13152491
Kastelic M, Gregurić Gračner G, Tomažič I, Kvapil P, Harej M, Dovč A. Comparison of Cortisol Concentrations in Different Matrices in Alpine Ibex (Capra ibex) at the Zoo. Animals. 2023; 13(15):2491. https://doi.org/10.3390/ani13152491
Chicago/Turabian StyleKastelic, Marjan, Gordana Gregurić Gračner, Iztok Tomažič, Pavel Kvapil, Mojca Harej, and Alenka Dovč. 2023. "Comparison of Cortisol Concentrations in Different Matrices in Alpine Ibex (Capra ibex) at the Zoo" Animals 13, no. 15: 2491. https://doi.org/10.3390/ani13152491