Fork length ranged from 10.65 to 20.1 cm (mean value: 14.18, SD: 2.49); the weight ranged from 18.9 to 151.3 g (mean value: 57.18, SD: 29). Among the 22 sample units assembled, 11 units consisted of only female trout and 11 of only male ones; 45% of female sample units and 27% of male sample units were sexually mature. The sample mean fat content was 0.033 g g−1 muscle (range 0.080–0.130 g). GI parasites infested 54% of sample units.
3.1. OCs in Trout Muscle
PCBs were the most plentiful OCs followed by DDTs, HCB, and dieldrin (
Table 1 and
Table 2). Among PCBs, the mean concentration was highest for the Σ15ndl-PCBs group, followed by Σ6ndl-PCBs, Σ5dl-PCBs, and Σ3dl-PCBs (
Table 1). The Σ6ndl-PCBs amounted to roughly 72% of Σ15ndl-PCBs.
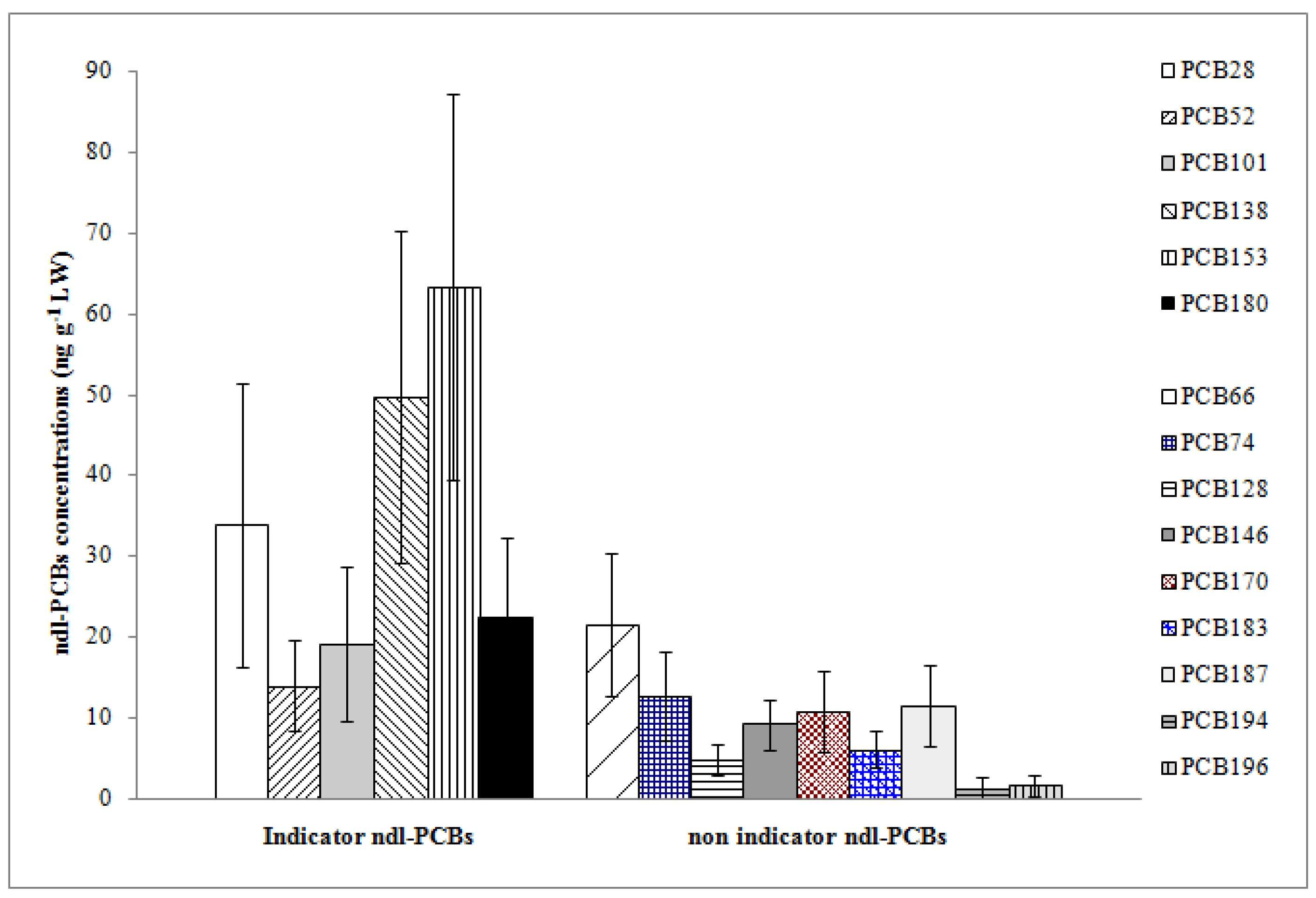
Each indicator ndl-PCB congener was detected in all sample units (
Figure 2). The indicator hexa-chlorinated PCBs IUPAC Nos. 153 and 138 showed the highest mean concentrations among all OCs, with values of 63.25 and 49.74 ng g
−1 LW, respectively. Adding up these two congeners, we accounted for 56% and about 40% of Σ6ndl-PCBs and Σ15ndl-PCBs, respectively. The tri-chlorinated PCB 28 followed with a mean concentration of 33.77 ng g
−1 LW. The indicator tetra-chlorinated PCB 52 was detected at the lowest concentration, contributing to 7% of Σ6ndl-PCBs (
Figure 2).
Regarding the non-indicator ndl-PCBs, the tetra-chlorinated PCB 66 was detected with the highest mean concentration (21.56 ng g−1 LW), accounting for 7.6% of Σ15ndl-PCBs, while the PCBs Nos. 194 and 196 were detected with the lowest ones (1.30 and 1.56 ng g−1 LW, respectively). The latter two congeners were found in 65% of sample units; other non-indicator ndl congeners were in all sample units.
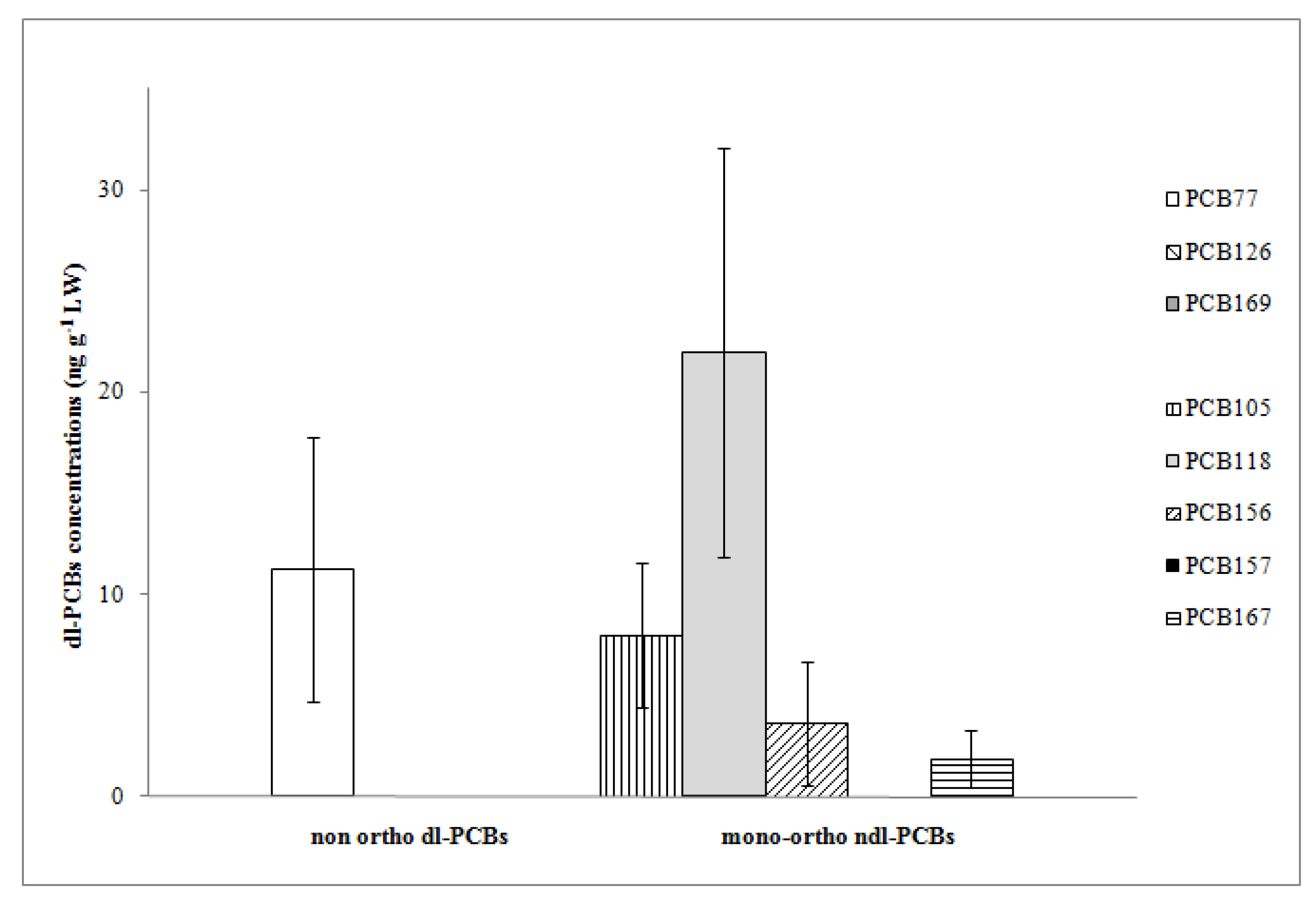
Regarding dl-PCBs, Σ5dl-PCBs were higher than Σ3dl-PCBs (mean value of 35.44 ng g
−1 LW vs. 11.30 ng g
−1 LW) (
Table 1). Among non-ortho congeners, PCBs Nos. 126 and 169 were never detected (
Figure 3); therefore, the concentration of Σ3dl-PCBs is only due to PCB 77 (mean value of 11.28 ng g
−1 LW and detection in 83% of sample units). Four out of five mono-ortho PCBs were detected; in particular, in all sample units, we found PCB 105 and PCB 118 (mean values 7.99 and 21.96 ng g
−1 LW, respectively). The latter congener showed the highest concentration among dl-PCBs and contributed 62% to Σ5dl-PCBs. PCB 156 and PCB 167 had relatively low mean concentration levels (3.63 and 1.86 ng g
−1 LW) (
Figure 3); they were detected in 69% and 78% of the sample units, respectively. The congeners PCB 118, PCB 77, and PCB 105, recognized as more toxic and usually detected at lower concentrations than ndl ones [
45], were instead found at mean concentrations roughly comparable to those of some indicator and non-indicator ndl-PCBs (i.e., PCB 52, 101, 180, 66, 74).
The finding of relatively high concentrations of PCBs fairly reflects our expectations. In fact, a National Interest Site and three sites contaminated with OCs undergoing remediation procedures are located in the same province of the study area.
Regarding OCPs,
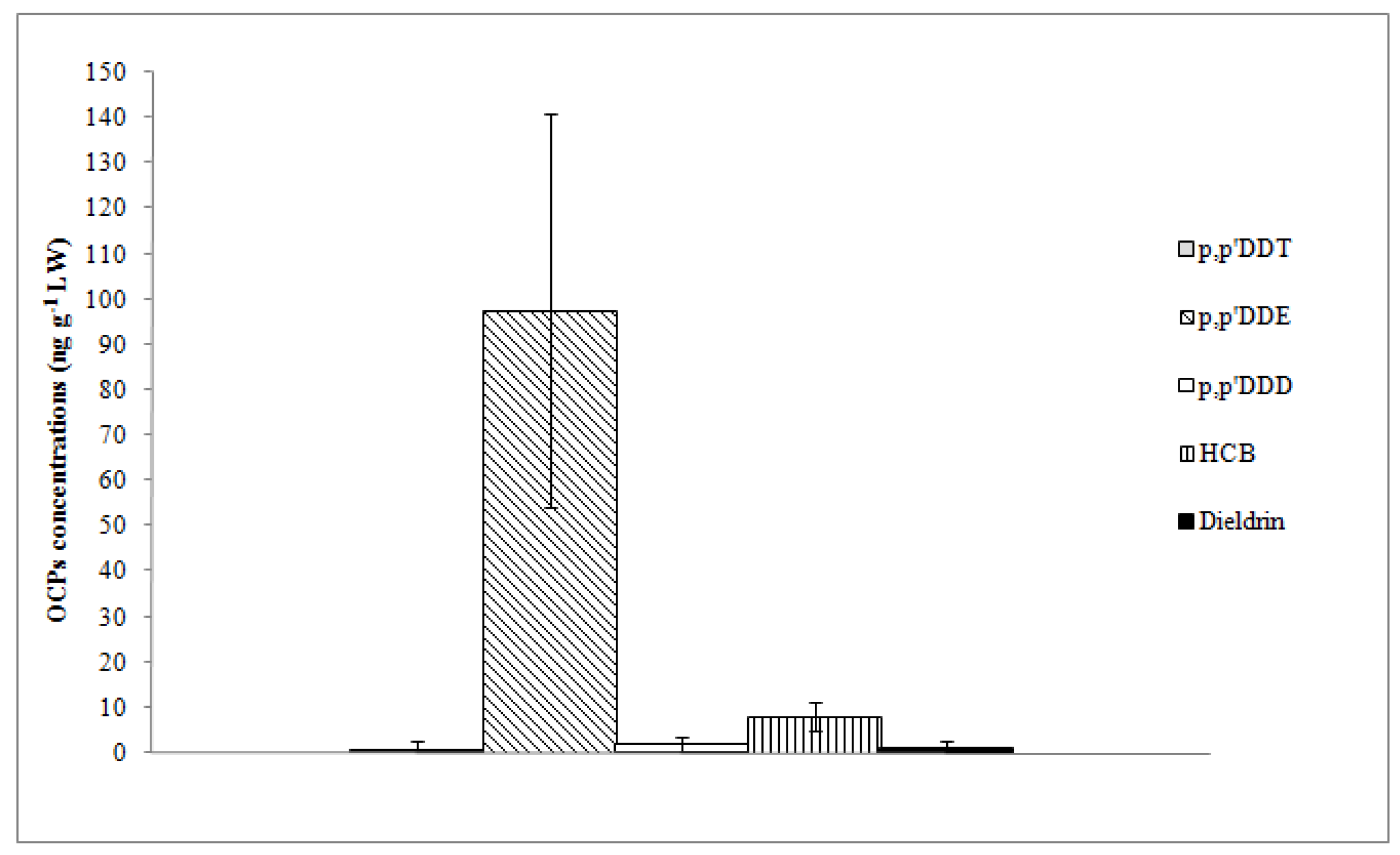
p,p’-DDE and HCB were found in all sample units with the highest mean concentrations (97.57 and 8.09 ng g
−1 LW, respectively);
p,p’-DDE was the dominant compound in terms of contribution to ΣDDTs (97%) (
Table 2).
p,p’-DDD,
p,p’-DDT, and dieldrin were detected in 83%, 26%, and 43% of sample units, respectively, with quite low concentrations (
Figure 4).
As it is well known, DDT is quickly metabolized to DDE that persists for a long time in the environment. Indeed, the relatively high
p,p’-DDE concentration suggests that the parent compound was strongly used in the past. This hypothesis is supported by the value of the DDE/DDT ratio, based on the average concentration of the compounds, which is usually used to investigate the timing of DDT entering the environment [
46]. In our case, the ratio is much higher than 1 (amounting to 123). The reasons behind the presence of HCB can be identified both with its past use as a fungicide and because it is a by-product of several chlorine-containing chemicals [
47].
Trout feeding could explain the presence of OCs. Crustaceans, insects, and small fish are the main components of trout’s diet. These organisms live on the seabed in direct contact with sediments that accumulate OCs.
3.2. Regression Analysis of Parasites Infection and OCs
A total of 233 helminth parasites belonging to three taxa were found in the GI tract of 41 brown trout. The most prevalent (42.5%) and abundant taxon (mean abundance: 5.4 per specimen ranging from 1 to 35) was the trematode
Crepidostomum metoecus, with the other trophically transmitted parasites found in 5.7% (the acanthocephalan
Echinorhynchus sp.) and 2.3% (unidentified nematode larva) trout, respectively (
Table 3).
Crepidostomum metoecus has been previously reported as a common parasite of brown trout in European countries [
31]. However, its presence has been rarely investigated in Italian rivers [
48].
The results of the regression analysis are shown in
Table 4,
Table 5 and
Table 6. When the outcome variable identified the presence or not of parasite infection, that is, the outcome variable was a dummy, and OC concentration levels were expressed on LW basis, the relationship between OCs and parasite presence was (i) negative and statistically significant in the case of Σ6ndl-PCBs, Σ15ndl-PCBs, Σ5dl-PCBs, and ΣDDTs; and (ii) negative but insignificant in the case of Σ3dl-PCBs, HCB, and dieldrin (see
Table 4). These results suggest that relatively high OC concentrations might reduce the probability of parasite infection.
The use of the two control variables enabled us to verify if previous results were biased by potential confounding factors.
Table 5 shows that the negative and statistically significant relationship between parasite infection and OC concentration still emerged for Σ6ndl-PCBs, Σ15ndl-PCBs, and Σ5dl-PCBs. Moreover, we also notice that while the variable sex does not have any statistical impact, the coefficient attached to the stage of sexual development (immature versus mature) was estimated negative for all regressions, and statistically significant in several cases, consistent with the idea that immature younger fish had less time to accumulate parasite.
Our main evidence was also confirmed by exploiting the Poisson regression analysis where the outcome variable was the number of parasites (see
Table 6). A negative and statistically significant relationship between parasite infection and OC concentration was estimated for Σ6ndl-PCBs, Σ15ndl-PCBs, and Σ5dl-PCBs.
Our evidence regarding the negative relationship between chemical pollutants and parasites accords with that of other authors. By relying on a meta-analysis of the effects of pollution on parasitism in aquatic animals, Blanar et al. [
26] conclude with a strong, significant negative effect for Digenea and Monogenea, especially in response to metal pollution. For many other parasite/contaminant interactions, effect sizes were instead not significantly different from zero. However, few studies focus on OCs and do not reach a consensus [
25,
49].
The digenetic class of trematodes, to which
Crepidostomum metoecus belongs, has been reported as more vulnerable than other parasite species and a negative correlation with OCs has been noticed [
26]; for freshwater taxa, the correlation was not determined due to lack of data relative to OC concentration levels. Vidal-Martinez et al. [
50] evidenced a significant negative correlation between the degree of parasitism by larval trematode
Mesostephanus appendiculatoides and DDT concentrations in the Mayan catfish collected in Chetumal Bay, Mexico. The authors reported that, although the catfish were highly contaminated, they seemed to be barely susceptible to the OCPs toxicity, which instead heavily affected parasites. A negative correlation was also documented with respect to the digenetic trematode
Steringophorus furciger in the GI tract of flatfish species, which is a significantly greater presence of parasites in specimens poorly contaminated by PCBs [
51]. Similar evidence was also reported by Carreras-Aubets et al. [
25], who observed a lower abundance of adult digenean endoparasite
Opecoeloides furcatus in
Mullus barbatus fish related to higher concentrations of the sum of indicator PCBs plus PCB 118 in Western Mediterranean sediments. To the best of our knowledge, a positive relationship has never been recorded for the digenetic class of trematodes. With data from German Bight, Schmidt et al. [
52] evidenced a negative relationship between the infection degree to some parasite species and OC concentrations measured in the muscle of European flounder, sediments, and blue mussels. In particular, they documented a lower abundance of parasites with higher concentrations of HCB, DDD, DDE, and the sum of indicator PCBs. In the liver and muscle of acanthocephalan-infected perch, Brázová and co-authors [
53] detected PCB concentrations several times lower than in not parasitized perc. Similar findings, observed only for some PCB congeners, were recently reported by the same authors for another host-parasite system (GI cestode–freshwater bream) [
54].
In addition to aquatic species, a negative correlation was also evidenced in dogs [
55] and African immigrants [
24]. The authors showed that serum concentration levels of some indicator PCBs (PCB 52, PCB 138, PCB 153) were significantly lower in subjects infected with GI parasites than in non-parasitized ones. The same results were found examining several OCs, including those that we analyzed in the present study, in dogs positive for earthworm
Dirofilaria immitis [
55]. Since OC serum concentrations remained low after antiparasitic treatment, the authors hypothesized that nematode parasites may metabolize OCs in addition to accumulating them.
Some other studies reported, instead, an increase in parasitism because of animal exposure to OCs. Parasitic infection prevalently by nematodes of the lung (but also intestine and uterus) was significantly and positively correlated to
p,p’-DDT,
p,p’-DDD, and
o,p’-DDD concentrations measured in the blubber of finless porpoises (
Neophocaena phocaenoides) [
56]. In blubber of the same species, Isobe et al. [
49] evidenced significantly higher concentrations of PCBs (expressed as the sum of 62 congeners including indicator congeners and dl-PCBs) in animals infected with liver trematodes, relative to non-infected ones. A similar correlation was also reported by Kannan and co-authors [
57] for PCBs and infectious agents including intestinal acanthocephalans, protozoa, bacteria, and fungi in another marine mammal, the Southern Sea otter. Bamidele and co-authors [
58,
59] observed a significant positive correlation between the protozoa
Myxosoma sp infection found in blackchin tilapia and silver catfish from Lagos lagoon (Nigeria) and
p,p’-DDT concentrations in freshwaters; however, no correlation was determined between intestinal helminth parasites and OCP congeners [
58].
Few potential explanations have been raised for the results obtained. Some authors related the findings of a positive relationship to the frequent immunosuppressive effect of the OCs reported in
in vivo and
in vitro studies [
6,
60,
61,
62]. Indeed, exposure to chemical xenobiotics can disrupt the host’s immune response, so that adverse effects may depend on the indirect outcome of the pollutant on the host’s capacity to cope with a pathogen [
13], making the host itself more susceptible to parasitosis [
63,
64]. Instead, regarding the negative correlation, some authors suggest that high environmental OC concentrations may disrupt the parasite life cycle, impairing the ability of the free-life form to survive and reach the host [
25,
51,
65]. To protect themselves from chemical contaminants, parasites may rely on their host for detoxification mechanisms, even though the parasitic infection itself could cause a decrease in host detoxification enzymes [
65]. In the case of trematodes, which have a complex life cycle with a first intermediate host, usually a mollusk, several free-living forms, and, finally, one or multiple vertebrate hosts, the toxic effects of the pollutants may impair their ability to assimilate essential nutrients; as a consequence of this condition, also their reproductive potential results to be affected and some mutagenic damages could occur to developing embryos, leading to a reduced transmission [
65].
The negative relationship has been also interpreted suggesting that parasites determine a decrease in OC concentrations. The explanation would be that, because of their lipid content, GI parasites, and thus also trematodes, are able to bioaccumulate OCs introduced by the animal host through contaminated feed consumption [
21,
53,
54,
66,
67]. This makes OCs less bioavailable for the host and therefore may increase the tolerance of the host to contaminants-induced toxicity. For parasites other than helminths, another explanation may result from their metabolizing ability through cytochrome P-450 activity leading to the reduction of OCs body burden in the host [
68]. Indeed, helminths perform limited OC detoxification because of the lack of cytochrome P450 mono-oxygenases [
69,
70]. Moreover, according to Brázová and co-authors [
54], since a higher Fulton’s condition factor was determined in infected breams, some parasites may even positively impact their hosts co-exposed to PCBs.
Overall, assessing the potential outcomes for parasites exploiting contaminated hosts is a crucial but neglected issue, since toxic effects on parasites may alter interspecific relationships. However, joint effects of parasites and chemical pollutants on host performance are extremely intricate depending on the level of parasitism, as well as the chemical pollutant, its mode of action, and the exposure levels. Contaminant accumulation by parasite results in positive effects among which is a reduction of both oxidative stress and histological modifications. This scenario indicates the possibility that a shift from parasitism to mutualism occurs. If concentration levels of the contaminant are high, they could induce damage to both host and parasites changing their relationships (see [
71] and references therein).
3.3. Risk Assessment
Human exposure to OCs is due to a large extent to the consumption of foods of animal origin, mainly dairy and fishery products [
72]. For consumer health risk assessment, the European Union (EU) established maximum residue limits (MRLs) for some OCs, namely dioxins, dl-PCBs, and six indicator ndl-PCBs in foodstuffs of animal origin (European Commission Regulation no. 1259/2011). The regulation fixed for “Muscle meat of wild caught freshwater fish, with the exception of diadromous fish species caught in freshwater, and products thereof” a MRL of 125 ng g
−1 WW for ∑6ndl-PCBs. All the samples analyzed showed concentrations below this threshold.
For dioxins and dl-PCBs, MRLs are expressed as Toxic Equivalent Quantity (WHO-TEQ). To evaluate the compliance of any sample units with the thresholds set by the EU, we summed up the concentration of each congener of interest multiplied by the respective Toxic Equivalency Factor adopted by the World Health Organization (WHO-TEFs) and then compared the sum with the corresponding MRL. TEFs are the results of the comparison of the relative different toxicity of individual dioxins/furans and dl-PCBs congeners to that of 2,3,7,8-TCDD [
73]. Since there is no specific limit for dl-PCBs, we considered a value of 3 pg g
−1 fat arising from the difference between the MRL referred to as the sum of all dioxin-related compounds (PCDDs/Fs and dl-PCBs) and the value referred to PCDDs/Fs alone. Also, in this case, the measured concentrations were lower than the above-extrapolated value.
With respect to the considered pesticides, the EU has not set a specific MRL for fish and fish products to date. As far as concern Italian national legislation, tolerance limits for residues of chloro-organic substances in products of animal origin have been established by Decreto Ministeriale on 13 May 2005 (annex 4), which states limits differentiated according to the different fat content. All sample units in the present study showed a lipid percentage below 5%, thus falling into the so-called group 1, with tolerance limits for ∑DDTs, HCB, and Dieldrin, respectively, of 0.050, 0.010, and 0.005 µg g−1 WW. No sample units exceeded the limits of the above-mentioned pesticides established by the Italian legislation.

 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}