
The Challenge of Global Warming in Water Buffalo Farming: Physiological and Behavioral Aspects and Strategies to Face Heat Stress

 , , , ,
, , , ,  , ,
, ,  ,
,  ,
,  , ,
, ,  ,
,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
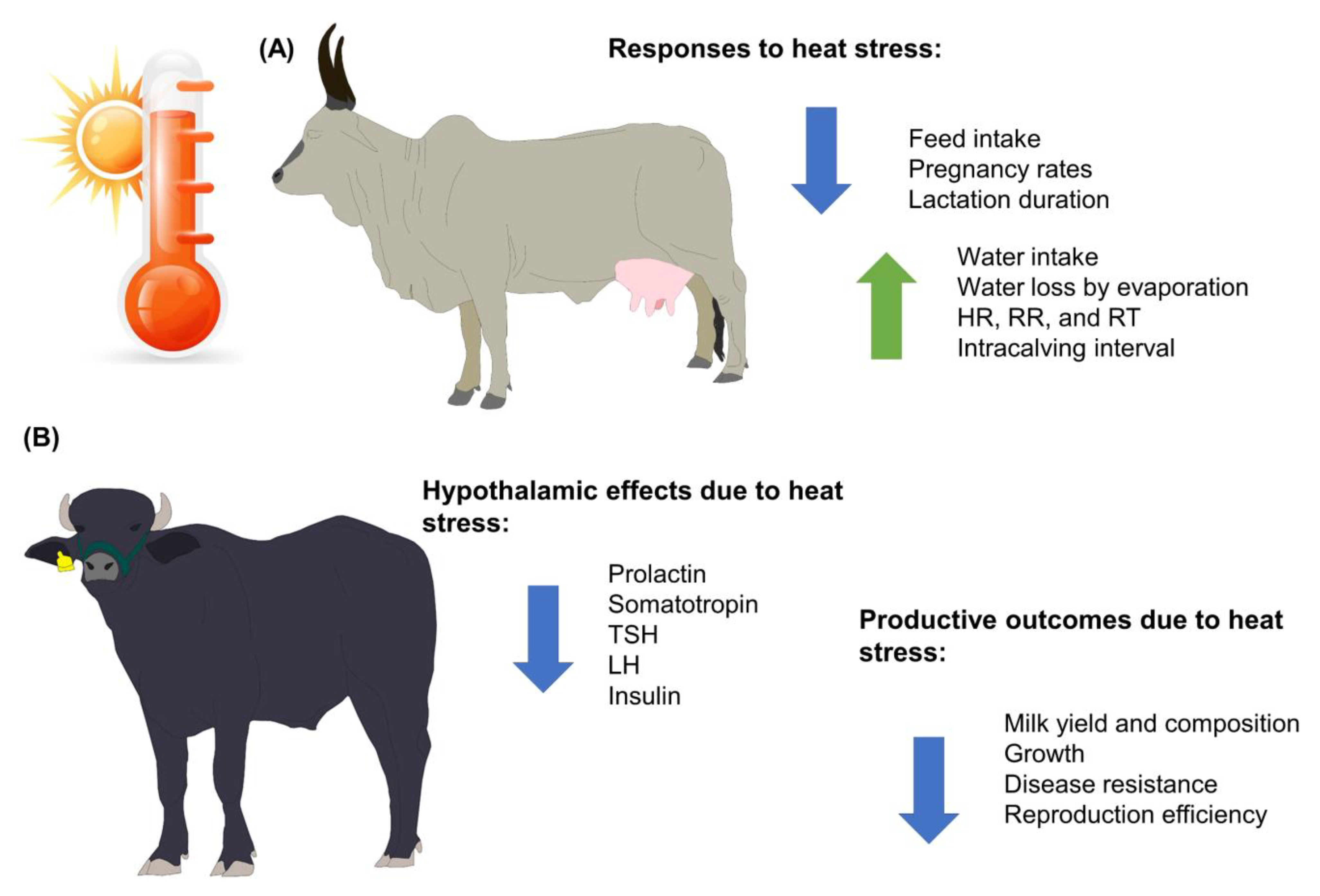
2. Adverse Effects of Global Warming on Livestock Farming: An Approach for Meat and Dairy Products
3. Thermoregulatory Physiology of Water Buffaloes Facing Heat Stress
3.1. Physiological Aspects
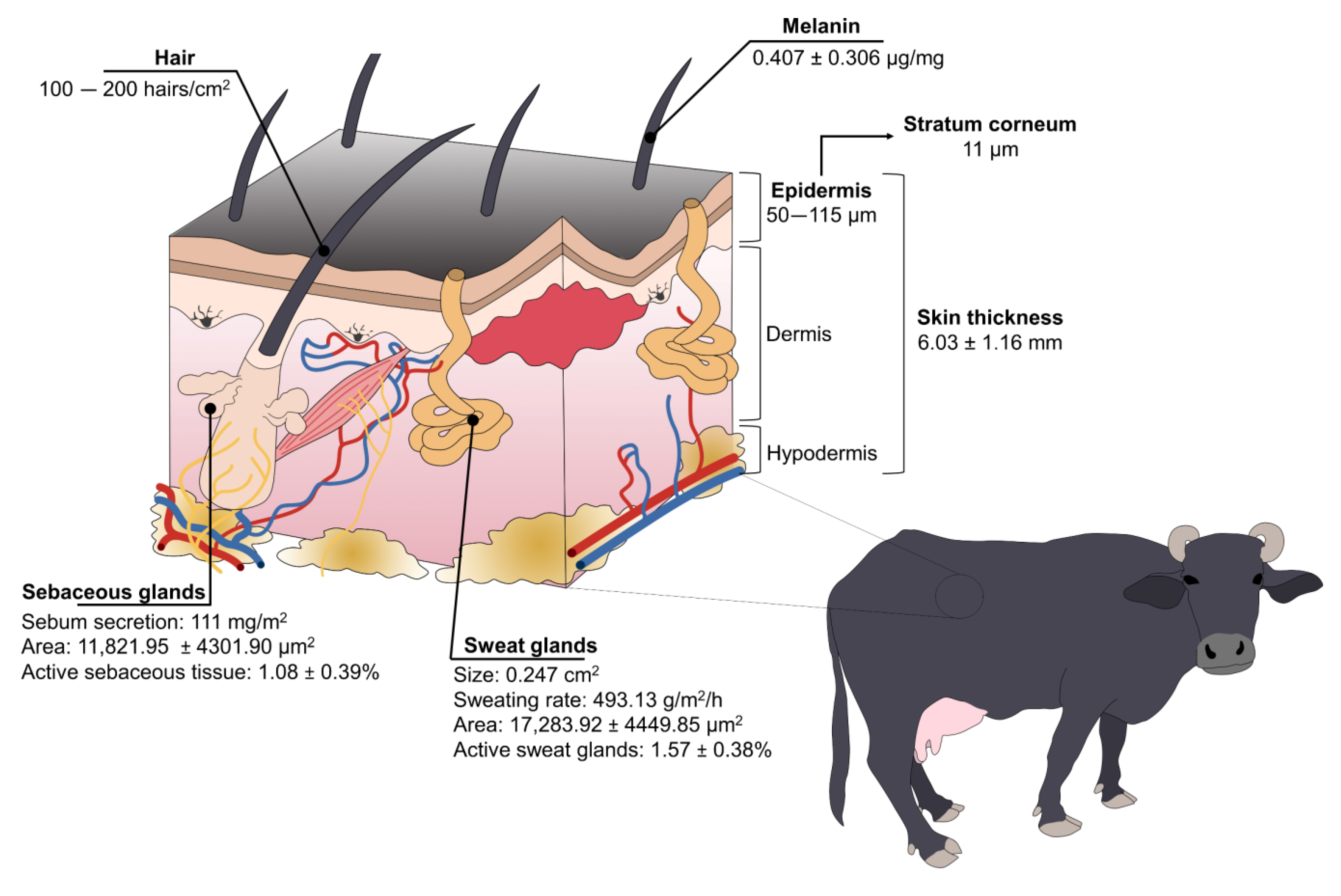
3.2. Anatomical Aspects
4. Behavioral Responses of Water Buffaloes to Diminish Heat Stress
4.1. Water Immersion
4.2. Seeking Shade
5. Influence of Natural/Artificial Shade on Buffaloes’ Thermoregulation
6. Thermal Imaging Applied to Evaluate Buffaloes’ Thermal States
7. Future Directions
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wisconsin Department of Natural Resources. The Science of Climate Change. Available online: https://dnr.wisconsin.gov/climatechange/science (accessed on 13 August 2023).
- Shraddha, R.; Nain, A.S. Impact of weather variables on Milk Production of Buffaloes. J. Agrometeorol. 2014, 16, 90–94. [Google Scholar]
- Bernabucci, U. Climate change: Impact on livestock and how can we adapt. Anim. Front. 2019, 9, 3–5. [Google Scholar] [CrossRef] [PubMed]
- Wolfenson, D.; Roth, Z.; Meidan, R. Impaired reproduction in heat-stressed cattle: Basic and applied aspects. Anim. Reprod. Sci. 2000, 60–61, 535–547. [Google Scholar] [CrossRef] [PubMed]
- Rawat, S.; Nain, A.S.; Roy, S. Biometeorological aspects of conception rate in cattle. J. Agrometeorol. 2014, 16, 116–120. [Google Scholar] [CrossRef]
- Thornton, P.; Nelson, G.; Mayberry, D.; Herrero, M. Impacts of heat stress on global cattle production during the 21st century: A modelling study. Lancet Planet. Health 2022, 6, e192–e201. [Google Scholar] [CrossRef]
- Cartwright, S.L.; Schmied, J.; Karrow, N.; Mallard, B.A. Impact of heat stress on dairy cattle and selection strategies for thermotolerance: A review. Front. Vet. Sci. 2023, 10, 1198697. [Google Scholar] [CrossRef]
- Habimana, V.; Nguluma, A.S.; Nziku, Z.C.; Ekine-Dzivenu, C.C.; Morota, G.; Mrode, R.; Chenyambuga, S.W. Heat stress effects on milk yield traits and metabolites and mitigation strategies for dairy cattle breeds reared in tropical and sub-tropical countries. Front. Vet. Sci. 2023, 10, 1121499. [Google Scholar] [CrossRef] [PubMed]
- Marumo, J.L.; Lusseau, D.; Speakman, J.R.; Mackie, M.; Hambly, C. Influence of environmental factors and parity on milk yield dynamics in barn-housed dairy cattle. J. Dairy Sci. 2022, 105, 1225–1241. [Google Scholar] [CrossRef]
- El Debaky, H.A.; Kutchy, N.A.; Ul-Husna, A.; Indriastuti, R.; Akhter, S.; Purwantara, B.; Memili, E. Review: Potential of water buffalo in world agriculture: Challenges and opportunities. Appl. Anim. Sci. 2019, 35, 255–268. [Google Scholar] [CrossRef]
- Sinha, A. Global Warming: Why India Is Heating Up Slower than the World Average. Available online: https://indianexpress.com/article/explained/explained-climate/climate-change-why-india-is-heating-up-slower-than-the-world-average-8602414/ (accessed on 3 August 2023).
- Imran, Z. The “Press and Pulse” of Climate Change Strains Farmers in Pakistan. Available online: https://thebulletin.org/2023/02/the-press-and-pulse-of-climate-change-strains-farmers-in-pakistan/ (accessed on 3 August 2023).
- Data, W. The Climate in China. Available online: https://www.worlddata.info/asia/china/climate.php (accessed on 3 August 2023).
- Napolitano, F.; Bragaglio, A.; Braghieri, A.; El-Aziz, A.H.A.; Titto, C.G.; Villanueva-García, D.; Mora-Medina, P.; Pereira, A.M.F.; Hernández-Avalos, I.; José-Pérez, N.; et al. The effect of birth weight and time of day on the thermal response of newborn water buffalo calves. Front. Vet. Sci. 2023, 10, 1084092. [Google Scholar] [CrossRef]
- Vilela, R.A.; Lourenço Junior, J.D.B.; Jacintho, M.A.C.; Barbosa, A.V.C.; Pantoja, M.H.D.A.; Oliveira, C.M.C.; Garcia, A.R. Dynamics of Thermolysis and Skin Microstructure in Water Buffaloes Reared in Humid Tropical Climate—A Microscopic and Thermographic Study. Front. Vet. Sci. 2022, 9, 871206. [Google Scholar] [CrossRef] [PubMed]
- Marai, I.F.M.F.M.; Haeeb, A.A.M.A.M. Buffalo’s biological functions as affected by heat stress—A review. Livest. Sci. 2010, 127, 89–109. [Google Scholar] [CrossRef]
- Pereira, A.M.F.; Vilela, R.A.; Titto, C.G.; Leme-dos-Santos, T.M.C.; Geraldo, A.C.M.; Balieiro, J.C.C.; Calviello, R.F.; Birgel Junior, E.H.; Titto, E.A.L. Thermoregulatory Responses of Heat Acclimatized Buffaloes to Simulated Heat Waves. Animals 2020, 10, 756. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Colli, L.; Barker, J.S.F. Asian water buffalo: Domestication, history and genetics. Anim. Genet. 2020, 51, 177–191. [Google Scholar] [CrossRef] [PubMed]
- Berihulay, H.; Abied, A.; He, X.; Jiang, L.; Ma, Y. Adaptation mechanisms of small ruminants to environmental heat stress. Animals 2019, 9, 75. [Google Scholar] [CrossRef]
- Mota-Rojas, D.; Habeeb, A.A.; Napolitano, F.; Sarubbi, J.; Ghezzi, M.D.; Ceriani, M.C.; Cuibus, A.; Martínez-Burnes, J.; Braghieri, A.; Lendez, P.A.; et al. Bienestar del búfalo de agua, bovino europeo y bovino índico: Aspectos medioambientales, fisiológicos y conductuales en respuesta a la sombra natural y artificial. In El Búfalo de Agua en Latinoamérica. Hallazgos Recientes; Napolitano, F., Mota-Rojas, D., Guerrero-Legarreta, I., Orihuela, A., Eds.; B.M. Editores: Mexico City, Mexico, 2020; pp. 959–1015. Available online: https://www.lifescienceglobal.com/journals/journal-of-buffalo-science/97-abstract/jbs/4550-el-bufalo-de-agua-en-latinoamerica-hallazgos-recientes (accessed on 13 September 2023).
- Garcia, A.R.; Silva, L.K.X.; Barros, D.V.; de, B. Lourenço Junior, J.; Martorano, L.G.; Lisboa, L.S.S.; da Silva, J.A.R.; de Sousa, J.S.; da Silva, A.O.A. Key points for the thermal comfort of water buffaloes in Eastern Amazon. Ciênc. Rural 2023, 53, e20210544. [Google Scholar] [CrossRef]
- Meissner, H.H.; Blignaut, J.N.; Smith, H.J.; Toit, C.J.L. The broad-based eco-economic impact of beef and dairy production: A global review. S. Afr. J. Anim. Sci. 2023, 53, 250–275. [Google Scholar]
- Mota-Rojas, D.; Álvarez-Macías, A.; Reyes, B.; Bertoni, A.; Peinado, S.F.; Flores, K.; Galarza, A.; Herrera, Y.; Torres, F.; Legarreta, I.G. La producción animal y el cambio climático. In Ciencia, Tecnología e Innovación en la Industria Cárnica I; Guerrero-Legarreta, I., Ed.; Editorial Académica Española: London, UK, 2020; pp. 9–52. [Google Scholar]
- Da Silva, W.C.; Printes, O.V.N.; Lima, D.O.; da Silva, É.B.R.; dos Santos, M.R.P.; Camargo Júnior, R.N.C.; Barbosa, A.V.C.; da Silva, J.A.R.; e Silva, A.G.M.; Silva, L.K.X.; et al. Evaluation of the temperature and humidity index to support the implementation of a rearing system for ruminants in the Western Amazon. Front. Vet. Sci. 2023, 10, 1198678. [Google Scholar] [CrossRef]
- Mota-Rojas, D.; Pereira, A.M.F.; Wang, D.; Martínez-Burnes, J.; Ghezzi, M.; Hernández-Avalos, I.; Lendez, P.; Mora-Medina, P.; Casas, A.; Olmos-Hernández, A.; et al. Clinical Applications and Factors Involved in Validating Thermal Windows Used in Infrared Thermography in Cattle and River Buffalo to Assess Health and Productivity. Animals 2021, 11, 2247. [Google Scholar] [CrossRef]
- Zucali, M.; Tamburini, A.; Sandrucci, A.; Bava, L. Global warming and mitigation potential of milk and meat production in Lombardy (Italy). J. Clean. Prod. 2017, 153, 474–482. [Google Scholar] [CrossRef]
- Balhara, A.K.; Nayan, V.; Dey, A.; Singh, K.P.; Dahiya, S.S.; Singh, I. Climate change and buffalo farming in major milk producing states of India. Indian J. Anim. Sci. 2017, 87, 403–411. [Google Scholar] [CrossRef]
- ONU. Paz, Dignidad e Igualdad en un Planeta Sano; United Nations: New York, NY, USA, 2022; Volume 1.
- Gerber, P.; Steinfeld, H.; Henderson, B.; Mottet, A.; Opio, C.; Djikman, J.; Falcucci, A.; Tempio, G. Tackling Climate Change through Livestock—A Global Assessment of Emissions and Mitigation Opportunities; Food and Agriculture Organization of the United Nations: Rome, Italy, 2013. [Google Scholar]
- Das, S. Impact of climate change on livestock, various adaptive and mitigative measures for sustainable Livestock Production. Approaches Poult. Dairy Vet. Sci. 2017, 1, 64–70. [Google Scholar] [CrossRef]
- Das, K.S.; Singh, J.K.; Singh, G.; Upadhyay, R.C.; Malik, R.; Oberoi, P.S. Heat stress alleviation in lactating buffaloes: Effect on physiological response, metabolic hormone, milk production and composition. Indian J. Anim. Sci. 2014, 84, 275–280. [Google Scholar]
- De Sousa, K.T.; Deniz, M.; do Vale, M.M.; Dittrich, J.R.; Hötzel, M.J. Influence of microclimate on dairy cows’ behavior in three pasture systems during the winter in south Brazil. J. Therm. Biol. 2021, 97, 102873. [Google Scholar] [CrossRef] [PubMed]
- Kiktev, N.; Lendiel, T.; Vasilenkov, V.; Kapralyuk, O.; Hutsol, T.; Glowacki, S.; Kuboń, M.; Kowalczyk, Z. Automated Microclimate Regulation in Agricultural Facilities Using the Air Curtain System. Sensors 2021, 21, 8182. [Google Scholar] [CrossRef] [PubMed]
- Seerapu, S.R.; Kancharana, A.R.; Chappidi, V.S.; Bandi, E.R. Effect of microclimate alteration on milk production and composition in Murrah buffaloes. Vet. World 2015, 8, 1444–1452. [Google Scholar] [CrossRef]
- Sigdel, A.; Bhattarai, N.; Kolachhapati, M.R. Impacts of climate change on Milk Production of Murrah buffaloes in Kaski, Nepal. In Proceedings of the International Conference on Climate Change Innovation and Resilience for Sustainable Livelihood, Kathmandu, Nepal, 12–14 January 2015; pp. 205–207. [Google Scholar]
- Shahbaz, P.; Boz, I.; ul Haq, S. Adaptation options for small livestock farmers having large ruminants (cattle and buffalo) against climate change in Central Punjab Pakistan. Environ. Sci. Pollut. Res. 2020, 27, 17935–17948. [Google Scholar] [CrossRef]
- Di Stasio, L.; Brugiapaglia, A. Current Knowledge on River Buffalo Meat: A Critical Analysis. Animals 2021, 11, 2111. [Google Scholar] [CrossRef]
- Bragaglio, A.; Maggiolino, A.; Romano, E.; De Palo, P. Role of Corn Silage in the Sustainability of Dairy Buffalo Systems and New Perspective of Allocation Criterion. Agriculture 2022, 12, 828. [Google Scholar] [CrossRef]
- Guerrero-Legarreta, I.; Napolitano, F.; Cruz-Monterrosa, R.; Mota-Rojas, D.; Mora-Medina, P.; Ramírez-Bribiesca, E.; Bertoni, A.; Berdugo-Gutiérrez, J.; Braghieri, A. River buffalo meat production and quality: Sustainability, productivity, nutritional and sensory properties. J. Buffalo Sci. 2020, 9, 159–169. [Google Scholar] [CrossRef]
- Álvarez-Macías, A.; Mota-Rojas, D.; Bertoni, A.; Dávalos-Flores, J.L. Opciones de desarrollo de los sistemas de producción de búfalos de agua de doble propósito en el trópico húmedo latinoamericano. In El búfalo de agua en Latinoamérica, Hallazgos Recientes; Napolitano, F., Mota-Rojas, D., Guerrero-Legarreta, I., Orihuela, A., Eds.; BM Editores: Mexico City, México, 2020; pp. 43–74. Available online: https://www.lifescienceglobal.com/journals/journal-of-buffalo-science/97-abstract/jbs/4550-el-bufalo-de-agua-en-latinoamerica-hallazgos-recientes (accessed on 13 September 2023).
- Kandeepan, G.; Anjaneyulu, A.S.R.; Kondaiah, N.; Mendiratta, S.K.; Lakshmanan, V. Effect of age and gender on the processing characteristics of buffalo meat. Meat Sci. 2009, 83, 10–14. [Google Scholar] [CrossRef] [PubMed]
- Escarcha, J.F.; Lassa, J.A.; Palacpac, E.P.; Zander, K.K. Livelihoods transformation and climate change adaptation: The case of smallholder water buffalo farmers in the Philippines. Environ. Dev. 2020, 33, 100468. [Google Scholar] [CrossRef]
- Collier, R.J.; Baumgard, L.H.; Zimbelman, R.B.; Xiao, Y. Heat stress: Physiology of acclimation and adaptation. Anim. Front. 2019, 9, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Mota-Rojas, D.; Titto, C.G.; Orihuela, A.; Martínez-Burnes, J.; Gómez-Prado, J.; Torres-Bernal, F.; Flores-Padilla, K.; Carvajal-de la Fuente, V.; Wang, D.; la Fuente, V.C.; et al. Physiological and Behavioral Mechanisms of Thermoregulation in Mammals. Animals 2021, 11, 1733. [Google Scholar] [CrossRef]
- Wang, T.A.; Teo, C.F.; Åkerblom, M.; Chen, C.; Tynan-La Fontaine, M.; Greiner, V.J.; Diaz, A.; McManus, M.T.; Jan, Y.N.; Jan, L.Y. Thermoregulation via Temperature-Dependent PGD2 Production in Mouse Preoptic Area. Neuron 2019, 103, 309–322.e7. [Google Scholar] [CrossRef]
- Nakamura, K. Central circuitries for body temperature regulation and fever. Am. J. Physiol. Integr. Comp. Physiol. 2011, 301, R1207–R1228. [Google Scholar] [CrossRef]
- Nazıroğlu, M.; Braidy, N. Thermo-Sensitive TRP Channels: Novel Targets for Treating Chemotherapy-Induced Peripheral Pain. Front. Physiol. 2017, 8, 1040. [Google Scholar] [CrossRef]
- Lezama-García, K.; Mota-Rojas, D.; Pereira, A.M.F.; Martínez-Burnes, J.; Ghezzi, M.; Domínguez, A.; Gómez, J.; de Mira Geraldo, A.; Lendez, P.; Hernández-Ávalos, I.; et al. Transient Receptor Potential (TRP) and Thermoregulation in Animals: Structural Biology and Neurophysiological Aspects. Animals 2022, 12, 106. [Google Scholar] [CrossRef]
- Morgan, M.; Nencini, S.; Thai, J.; Ivanusic, J.J. TRPV1 activation alters the function of Aδ and C fiber sensory neurons that innervate bone. Bone 2019, 123, 168–175. [Google Scholar] [CrossRef]
- Das, S.K.; Upadhyay, R.C.; Madan, M.L. Heat stress in Murrah buffalo calves. Livest. Prod. Sci. 1999, 61, 71–78. [Google Scholar] [CrossRef]
- Da Silva, J.A.R.; de Araújo, A.A.; Lourenço Júnior, J.D.B.; Santos, N.D.F.A.D.; Garcia, A.R.; Nahúm, B.D.S. Conforto térmico de búfalas em sistema silvipastoril na Amazônia Oriental. Pesqui. Agropecuária Bras. 2011, 46, 1364–1371. [Google Scholar] [CrossRef]
- Zhao, Z.-D.; Yang, W.Z.; Gao, C.; Fu, X.; Zhang, W.; Zhou, Q.; Chen, W.; Ni, X.; Lin, J.-K.; Yang, J.; et al. A hypothalamic circuit that controls body temperature. Proc. Natl. Acad. Sci. USA 2017, 114, 2042–2047. [Google Scholar] [CrossRef] [PubMed]
- Morrison, S.F.; Nakamura, K. Central Mechanisms for Thermoregulation. Annu. Rev. Physiol. 2019, 81, 285–308. [Google Scholar] [CrossRef] [PubMed]
- Shafton, A.D.; McAllen, R.M. Location of cat brain stem neurons that drive sweating. Am. J. Physiol. Integr. Comp. Physiol. 2013, 304, R804–R809. [Google Scholar] [CrossRef]
- Rodríguez-González, D.; Guerrero-Legarreta, I.; Cruz Monterrosa, R.G.; Napolitano, F.; Gonçalves-Titto, C.; El-Aziz, A.H.A.; Hernández- Ávalos, I.; Casas-Alvarado, A.; Oliva- Domínguez, A.; Mota-Rojas, D. Assessment of thermal changes in water buffalo mobilized from the paddock and transported by short journeys. Front. Vet. Sci. 2023, 10, 1184577. [Google Scholar] [CrossRef]
- Alam, M.; Gregory, N.; Uddin, M.; Jabbar, M.; Silva-Fletchner, A.; Kempson, A.G.; Saifuddin, A.K. Frequency of heat stress in cattle and water buffalo at livestock markets in Bangladesh. J. Commonw. Vet. Assoc. 2010, 26, 13–17. [Google Scholar]
- Pressicce, G.A. The Buffalo (Bubalus bubalis): Production and Research; Bentham Science Publishers: Sharjah, United Arab Emirates, 2017. [Google Scholar]
- Pi-Hua, C.; Cheng-Yung, L.; Ching-Feng, W. The comparison of histology and melanin contents of hairs and skin between the black and white Taiwan water buffalo. J. Taiwan Livest. Res. 2009, 42, 235–244. [Google Scholar]
- Debbarma, D.; Uppal, V.; Bansal, N.; Gupta, A. Histomorphometrical Study on Regional Variation in Distribution of Sweat Glands in Buffalo Skin. Dermatol. Res. Pract. 2018, 2018, 5345390. [Google Scholar] [CrossRef]
- Hafez, E.S.E.; Badreldin, A.L.; Shafei, M.M. Skin structure of Egyptian buffaloes and cattle with particular reference to sweat glands. J. Agric. Sci. 1955, 46, 19–30. [Google Scholar] [CrossRef]
- Shafie, M.; Abou El-Khair, M.; Shafies, M.M.; El-Khair, M.M.A. Activity of the sebaceous glands of bovines in hot climates. Egypt. J. Anim. Prod. 1970, 10, 81–98. [Google Scholar] [CrossRef]
- El Sabry, M.; Almasri, O. Space allowance: A tool for improving behavior, milk and meat production, and reproduction performance of buffalo in different housing systems—A review. Trop. Anim. Health Prod. 2022, 54, 7–10. [Google Scholar] [CrossRef] [PubMed]
- Galloso-Hernández, M.A.; Rodríguez-Estévez, V.; Alvarez-Díaz, C.A.; Soca-Pérez, M.; Dublin, D.; Iglesias-Gómez, J.; Simon Guelmes, L. Effect of Silvopastoral Systems in the Thermoregulatory and Feeding Behaviors of Water Buffaloes Under Different Conditions of Heat Stress. Front. Vet. Sci. 2020, 7, 393. [Google Scholar] [CrossRef] [PubMed]
- Mota-Rojas, D.; Habeeb, A.; Ghezzi, M.D.; Kanth, P.; Napolitano, F.; Lendez, P.A.; Cuibus, A.; Ceriani, M.C.; Sarubbi, J.; Braghieri, A.; et al. Termorregulación del búfalo de agua: Mecanismos neurobiológicos, cambios microcirculatorios y aplicaciones prácticas de la termografía infrarroja. In El Búfalo de Agua en Latinoamérica, Hallazgos Recientes; Napolitano, F., Mota-Rojas, D., Guerrero-Legarreta, I., Orihuela, A., Eds.; BM Editores: Mexico City, Mexico, 2020; pp. 923–958. [Google Scholar]
- Das Amit, M.; Dash, S.S.; Sahu, S.; Sarangi, A.; Singh, M. Effect of microclimate on feeding, drinking and physiological parameters of buffalo: A review. Pharma Innov. J. 2021, 10, 2416–2419. [Google Scholar]
- Galloso-Hernández, M.A.; Soca-Pérez, M.; Dublin, D.; Alvarez-Díaz, C.A.; Iglesias-Gómez, J.; Díaz-Gaona, C.; Rodríguez-Estévez, V. Thermoregulatory and feeding behavior under different management and heat stress conditions in heifer water buffalo (Bubalus bubalis) in the tropics. Animals 2021, 11, 1162. [Google Scholar] [CrossRef] [PubMed]
- Joksimovic-Todorovic, M.; Davidovic, V.; Hristov, S.; Stankovic, B. Effect of heat stress on milk production in dairy cows. Biotechnol. Anim. Husb. 2011, 27, 1017–1023. [Google Scholar] [CrossRef]
- Napolitano, F.; Pacelli, C.; Grasso, F.; Braghieri, A.; De Rosa, G. The behaviour and welfare of buffaloes (Bubalus bubalis) in modern dairy enterprises. Animal 2013, 7, 1704–1713. [Google Scholar] [CrossRef]
- Mota-Rojas, D.; Napolitano, F.; Braghieri, A.; Guerrero-Legarreta, I.; Bertoni, A.; Martínez-Burnes, J.; Cruz-Monterrosa, R.; Gómez, J.; Ramírez-Bribiesca, E.; Barrios-García, H.; et al. Thermal biology in river buffalo in the humid tropics: Neurophysiological and behavioral responses assessed by infrared thermography. J. Anim. Behav. Biometeorol. 2021, 9, 21003. [Google Scholar] [CrossRef]
- Koga, A.; Chanpongsang, S.; Chaiyabutr, N. Importance of body-water circulation for body-heat dissipation in hot-humid climates: A distinctive body-water circulation in swamp buffaloes. Ital. J. Anim. Sci. 2007, 6, 1219–1222. [Google Scholar] [CrossRef]
- Da Silva, J.A.R.; de, A. Pantoja, M.H.; da Silva, W.C.; de Almeida, J.C.F.; de P.P. Noronha, R.; Barbosa, A.V.C.; de B. Lourenço Júnior, J. Thermoregulatory reactions of female buffaloes raised in the sun and in the shade, in the climatic conditions of the rainy season of the Island of Marajó, Pará, Brazil. Front. Vet. Sci. 2022, 9, 998544. [Google Scholar] [CrossRef]
- Fundora, O.; Sánchez, R.R.R. Datos preliminares de la conducta alimentaria de búfalos de río en pastoreo. Rev. Cuba Cienc. Agrícol. 2001, 35, 15–17. [Google Scholar]
- Galloso, H.M.A. Potencial de los Sistemas Silvopastoriles para la Producción Bufalina en Ambientes Tropicales; UCO Press: Córdoba, Spain, 2021. [Google Scholar]
- Tucker, C.B.; Rogers, A.R.; Schütz, K.E. Effect of solar radiation on dairy cattle behaviour, use of shade and body temperature in a pasture-based system. Appl. Anim. Behav. Sci. 2008, 109, 141–154. [Google Scholar] [CrossRef]
- De, S. Ablas, D.; Titto, E.A.; Pereira, A.M.; da C. Leme, T.M. Comportamento de bubalinos a pasto frente a disponibilidade de sombre e água para imersao. Braz. Anim. Sci. 2007, 8, 167–176. [Google Scholar]
- Tikul, N.; Prachum, S. Passive cooling strategies for cattle housing on small farms: A case study. Maejo Int. J. Sci. Technol. 2022, 16, 25–39. [Google Scholar]
- De Souza, D.; Manuel Franco Pereira, A.; Gonçalves, C.; Da Cunha Leme, M. Comportamento De Bubalinos a Pasto Frente a Disponibilidade De Sombra E Água Para Imersão. Ciênc. Anim. Bras. 2007, 8, 167–175. [Google Scholar]
- Younas, U.; Abdullah, M.; Bhatti, J.A.; Ahmed, N.; Shahzad, F.; Idris, M.; Tehseen, S.; Junaid, M.; Tehseen, S.; Ahmed, S. Biochemical and physiological responses of Nili-Ravi Buffalo (Bubalus bubalis) to heat stress. Turk. J. Vet. Anim. Sci. 2020, 44, 1196–1202. [Google Scholar] [CrossRef]
- Ahmad, M.; Bhatti, J.A.; Abdullah, M.; Ullah, R.; ul Ain, Q.; Hasni, M.S.; Mahboob, A.; Rashid, A.; Qaisar, I.; Rashid, G.; et al. Different ambient management intervention techniques and their effect on milk production and physiological parameters of lactating NiliRavi buffaloes during hot dry summer of subtropical region. Trop. Anim. Health Prod. 2019, 51, 911–918. [Google Scholar] [CrossRef]
- Aggarwal, A.; Upadhyay, R. Shelter management for alleviation of heat stress in cows and buffaloes. In Heat Stress and Animal Productivity; Aggarwal, A., Upadhyay, R., Eds.; Springer: New Delhi, India, 2013; pp. 169–183. [Google Scholar]
- Kumar, A.; Kamboj, M.L.; Chandra, S.; Bharti, P. Effect of modified housing system on physiological parameters of Murrah buffaloes during autumn and winter season. Indian J. Anim. Res. 2017, 52, 829–833. [Google Scholar] [CrossRef]
- Vranda, R.; Satyanarayan, K.; Jagadeeswary, V.; Veeranna, K.C.; Rajeshwari, Y.B.; Sudha, G.; Shree, J.S. A Study on Different Housing Practices of Buffaloes in Bidar District of Karnataka. Int. J. Sci. Environ. Technol. 2017, 6, 295–302. [Google Scholar]
- Patel, N.S.; Patel, J.V.; Parmar, D.V.; Ankuya, K.J.; Patel, V.K.; Madhavatar, M.P. Survey on housing practices of buffaloes owners in Patan district of Gujarat, India. J. Entomol. Zool. Stud. 2019, 7, 635–640. [Google Scholar]
- Mote, M.G.; Dhumal, P.T.; Gaikwad, U.S. Housing management practices followed by Buffalo owners in Purandar Tehsil of Pune district. Indian J. Anim. Prod. Manag. 2021, 36, 36–40. [Google Scholar]
- Barman, R.; Sinha, R.; Prasad, A.; Verma, R.; Jha, D. To Compare the effect of differents shade materials on the physiological and biochemical variables in buffalo calves in summer season. Int. J. Livest. Res. 2018, 8, 292. [Google Scholar] [CrossRef]
- Holt, P.F. Small animals in the study of pathological effects of asbestos. Environ. Health Perspect. 1974, 9, 205–211. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Asbestos in Drinking-Water. Available online: https://iris.who.int/bitstream/handle/10665/350932/WHO-HEP-ECH-WSH-2021.4-eng.pdf (accessed on 28 September 2023).
- Mandal, D.K.; Mandal, A.; Bhakat, C.; Dutta, T.K. Effect of heat stress amelioration through open-ridge ventilated thatched roof housing on production and reproduction performance of crossbred Jersey cows. Trop. Anim. Health Prod. 2021, 53, 144. [Google Scholar] [CrossRef] [PubMed]
- Kamal, R.; Dutt, T.; Patel, M.; Dey, A.; Bharti, P.K.; Chandran, P.C. Heat stress and effect of shade materials on hormonal and behavior response of dairy cattle: A review. Trop. Anim. Health Prod. 2018, 50, 701–706. [Google Scholar] [CrossRef]
- Khongdee, T.; Sripoon, S.; Vajrabukka, C. The effects of high temperature and roof modification on physiological responses of swamp buffalo (Bubalus bubalis) in the tropics. Int. J. Biometeorol. 2013, 57, 349–354. [Google Scholar] [CrossRef]
- Sahu, S.; Saini, R.; Sarangi, A.; Singh, D. Effect on the use of expanded polyethylene sheet and white paint on roof structure as thermal comfort on physiological indices in buffalo heifers. Pharma Innov. J. 2021, 10, 2362–2365. [Google Scholar]
- Sahu, S.; Kumar, R.; Sarangi, A. Effect of roof modifications on microclimatic parameters in alleviating heat stress in dairy shed. Pharma Innov. J. 2021, 10, 2304–2308. [Google Scholar]
- Hazarika, D.; Laskar, S.K.; Amonge, T.K.; Gogoi, A.K.; Hazarika, H. Effect of different roof house on physiological and blood parameters of swamp buffalo calves of Assam. Indian J. Anim. Prod. Manag. 2013, 29, 68–71. [Google Scholar]
- Casas-Alvarado, A.; Mota-Rojas, D.; Hernández-Ávalos, I.; Mora-Medina, P.; Olmos-Hernández, A.; Verduzco-Mendoza, A.; Reyes-Sotelo, B.; Martínez-Burnes, J. Advances in infrared thermography: Surgical aspects, vascular changes, and pain monitoring in veterinary medicine. J. Therm. Biol. 2020, 92, 102664. [Google Scholar] [CrossRef]
- Mota-Rojas, D.; Martínez-Burnes, J.; Casas-Alvarado, A.; Gómez-Prado, J.; Hernández-Ávalos, I.; Domínguez-Oliva, A.; Lezama-García, K.; Jacome-Romero, J.; Rodríguez-González, D.; Pereira, A.M.F. Clinical usefulness of infrared thermography to detect sick animals: Frequent and current cases. CABI Rev. 2022, 17, 1–17. [Google Scholar] [CrossRef]
- José-Pérez, N.; Mota-Rojas, D.; Ghezzi, M.D.; Rosmini, M.R.; Mora-Medina, P.; Bertoni, A.; Rodríguez-González, D.; Domínguez-Oliva, A.; Guerrero-Legarreta, I. Effects of transport on water buffaloes (Bubalus bubalis): Factors associated with the frequency of skin injuries and meat quality. J. Anim. Behav. Biometeorol. 2022, 10, 2216. [Google Scholar] [CrossRef]
- Reyes-Sotelo, B.; Mota-Rojas, D.; Martínez-Burnes, J.; Olmos-Hernández, A.; Hernández-Ávalos, I.; José, N.; Casas-Alvarado, A.; Gómez, J.; Mora-Medina, P. Thermal homeostasis in the newborn puppy: Behavioral and physiological responses. J. Anim. Behav. Biometeorol. 2021, 9, 21012. [Google Scholar] [CrossRef]
- Domínguez-Oliva, A.; Hernández-Ávalos, I.; Olmos-Hernández, A.; Villegas-Juache, J.; Verduzco-Mendoza, A.; Mota-Rojas, D. Thermal response of laboratory rats (Rattus norvegicus) during the application of six methods of euthanasia assessed by infrared thermography. Animals 2023, 13, 2820. [Google Scholar] [CrossRef]
- Da Silva, W.C.; da Silva, J.A.R.; da Silva, É.B.R.; Barbosa, A.V.C.; Sousa, C.E.L.; de Carvalho, K.C.; dos Santos, M.R.P.; Neves, K.A.L.; Martorano, L.G.; Camargo Júnior, R.N.C.; et al. Characterization of Thermal Patterns Using Infrared Thermography and Thermolytic Responses of Cattle Reared in Three Different Systems during the Transition Period in the Eastern Amazon, Brazil. Animals 2023, 13, 2735. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Prado, J.; Pereira, A.M.F.; Wang, D.; Villanueva-García, D.; Domínguez-Oliva, A.; Mora-Medina, P.; Hernández-Avalos, I.; Martínez-Burnes, J.; Casas-Alvarado, A.; Olmos-Hernández, A.; et al. Thermoregulation mechanisms and perspectives for validating thermal windows in pigs with hypothermia and hyperthermia: An overview. Front. Vet. Sci. 2022, 9, 1023294. [Google Scholar] [CrossRef]
- Tattersall, G.J. Infrared thermography: A non-invasive window into thermal physiology. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2016, 202, 78–98. [Google Scholar] [CrossRef]
- Barros, D.V.; Silva, L.K.X.; Kahwage, P.R.; Lourenço Júnior, J.B.; Sousa, J.S.; Silva, A.G.M.; Franco, I.M.; Martorano, L.G.; Garcia, A.R. Assessment of surface temperatures of buffalo bulls (Bubalus bubalis) raised under tropical conditions using infrared thermography. Arq. Bras. Med. Vet. Zootec. 2016, 68, 422–430. [Google Scholar] [CrossRef]
- Brcko, C.C.; da Silva, J.A.R.; Martorano, L.G.; Vilela, R.A.; de, S. Nahúm, B.; Silva, A.G.M.; Barbosa, A.V.C.; Bezerra, A.S.; de B. Lourenço Júnior, J. Infrared Thermography to Assess Thermoregulatory Reactions of Female Buffaloes in a Humid Tropical Environment. Front. Vet. Sci. 2020, 7, 180. [Google Scholar] [CrossRef]
- Sevegnani, K.B.; Fernandes, D.P.B.; da Silva, S.H.M.-G. Evaluation of thermorregulatory capacity of dairy buffaloes using infrared thermography. Eng. Agrícola 2016, 36, 1–12. [Google Scholar] [CrossRef]
- Idris, M.; Uddin, J.; Sullivan, M.; McNeill, D.M.; Phillips, C.J.C. Non-Invasive Physiological Indicators of Heat Stress in Cattle. Animals 2021, 11, 71. [Google Scholar] [CrossRef]
- Nienaber, J.A.; Hahn, G.L. Livestock production system management responses to thermal challenges. Int. J. Biometeorol. 2007, 52, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Leroy Hahn, G. Environmental Management for Improved Livedstock Performance, Health and Well- beging. Jpn. J. Livest. Manag. 1995, 30, 113–127. [Google Scholar]
- Stewart, M.; Wilson, M.T.; Schaefer, A.L.; Huddart, F.; Sutherland, M.A. The use of infrared thermography and accelerometers for remote monitoring of dairy cow health and welfare. J. Dairy Sci. 2017, 100, 3893–3901. [Google Scholar] [CrossRef] [PubMed]
- Lowe, G.; Sutherland, M.; Waas, J.; Schaefer, A.; Cox, N.; Stewart, M. Infrared Thermography—A Non-Invasive Method of Measuring Respiration Rate in Calves. Animals 2019, 9, 535. [Google Scholar] [CrossRef]
- Kim, S.; Hidaka, Y. Breathing Pattern Analysis in Cattle Using Infrared Thermography and Computer Vision. Animals 2021, 11, 207. [Google Scholar] [CrossRef]
- Mota-Rojas, D.; Wang, D.D.-H.; Titto, C.G.; Martínez-Burnes, J.; Villanueva-García, D.; Lezama, K.; Domínguez, A.; Hernández-Avalos, I.; Mora-Medina, P.; Verduzco, A.; et al. Neonatal infrared thermography images in the hypothermic ruminant model: Anatomical-morphological-physiological aspects and mechanisms for thermoregulation. Front. Vet. Sci. 2022, 9, 963205. [Google Scholar] [CrossRef]
- Lezama-García, K.; Martínez-Burnes, J.; Marcet-Rius, M.; Gazzano, A.; Olmos-Hernández, A.; Mora-Medina, P.; Domínguez-Oliva, A.; Pereira, A.M.F.; Hernández-Ávalos, I.; Baqueiro-Espinosa, U.; et al. Is the Weight of the Newborn Puppy Related to Its Thermal Balance? Animals 2022, 12, 3536. [Google Scholar] [CrossRef]
- Reyes-Sotelo, B.; Mota-Rojas, D.; Mora-Medina, P.; Ogi, A.; Mariti, C.; Olmos-Hernández, A.; Martínez-Burnes, J.; Hernández-Ávalos, I.; Sánchez-Millán, J.; Gazzano, A. Blood Biomarker Profile Alterations in Newborn Canines: Effect of the Mother′s Weight. Animals 2021, 11, 2307. [Google Scholar] [CrossRef]
- Taylor, C.R. The vascularity and possible thermoregulatory function of the horns in goats. Physiol. Zool. 1966, 39, 127–139. [Google Scholar] [CrossRef]
- Picard, K.; Thomas, D.W.; Festa-Bianchet, M.; Belleville, F.; Laneville, A. Differences in the thermal conductance of tropical and temperate bovid horns. Ecoscience 1999, 6, 148–158. [Google Scholar] [CrossRef]
- Cain, J.W.; Krausman, P.R.; Rosenstock, S.S.; Turner, J.C. Mechanisms of thermoregulation and water balance in desert ungulates. Wildl. Soc. Bull. 2006, 34, 570–581. [Google Scholar] [CrossRef]
- Zhao, H.; Zhou, Y.; Li, X.; Liu, C.; Chen, X. The influence of wind speed on infrared temperature in impervious surface areas based on in situ measurement data. GIScience Remote Sens. 2019, 56, 843–863. [Google Scholar] [CrossRef]
- Ring, F.J. Skin Thermal Imaging. In Measuring the Skin; Humbert, P., Maibach, H., Fanian, F., Agache, P., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 1–8. [Google Scholar]
- Abayawansa, W.D.; Prabhakar, S.; Singh, A.K.; Brar, P.S. Effect of climatic changes on reproductive performance of Murrah buffaloes in Punjab: A rertrospective analysis. Indian J. Anim. Sci. 2011, 81, 334–339. [Google Scholar]
- Fournel, S.; Rousseau, A.N.; Laberge, B. Rethinking environment control strategy of confined animal housing systems through precision livestock farming. Biosyst. Eng. 2017, 155, 96–123. [Google Scholar] [CrossRef]
- Fernández-Peña, C.; Reimúndez, A.; Viana, F.; Arce, V.M.; Señarís, R. Sex differences in thermoregulation in mammals: Implications for energy homeostasis. Front. Endocrinol. 2023, 14, 1093376. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
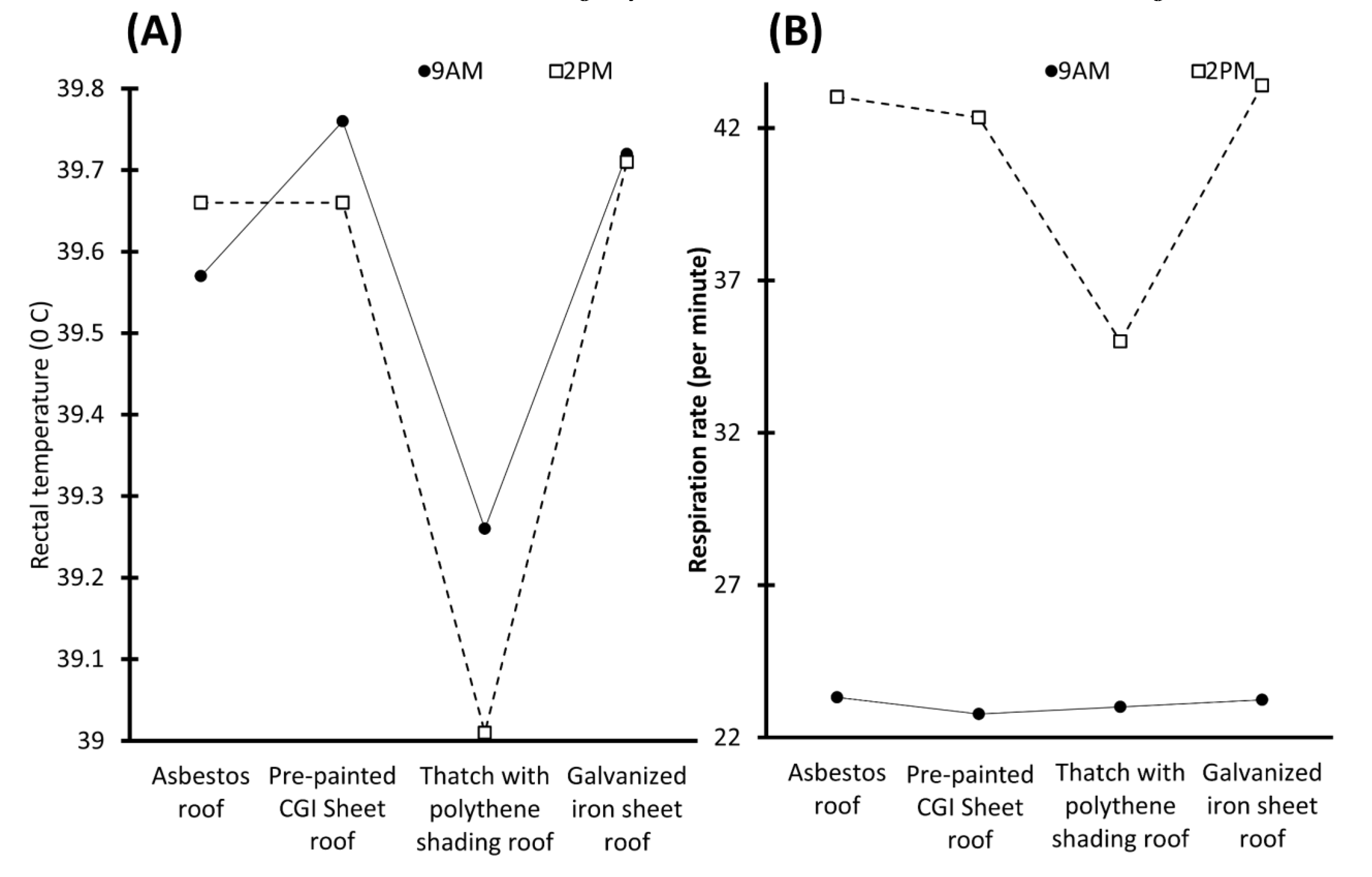
| Evaluated Material | Preferred Material | Physiological Response | THI | Region | Reference |
|---|---|---|---|---|---|
| G1: Corrugated iron roof G2: Corrugated iron roof and shade cloth | G2 | Male swamp buffaloes in G2 had lower RT (39.14 ± 0.07 °C vs. 40.00 ± 0. 10 °C) and cortisol values (2.14 ± 0.24 ng/mL vs. 3.38 ± 0.37 ng/mL). | 91.95 vs. 93.66 | Chainat Province, Thailand | [90] |
| G1: Corrugated asbestos roof G2: Corrugated asbestos roof painted white on upper side G3: Corrugated asbestos roof with expanded polyethylene (EPE) G4: Corrugated asbestos roof painted white on upper side and EPE sheet on lower side | G4 | Murrah buffalo heifers had RR of 26.60 ± 0.34 counts/min, pulse rate of 59.72 ± 0.74 counts/min, and RT of 38.36 ± 17.73 °C. | G1: 83.55 G2: 83.18 G3: 82.73 G4: 82.03 | Hisar, India | [91,92] |
| G1: Asbestos roof G2: Asbestos roof with bamboo ceiling G3: Thatch roof | G2 and G3 | G2 buffaloes had lower pulse rate, glucose, and serum proteins. G3 buffaloes had lower RT and RR. | Not available | Assam, India | [93] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Napolitano, F.; De Rosa, G.; Chay-Canul, A.; Álvarez-Macías, A.; Pereira, A.M.F.; Bragaglio, A.; Mora-Medina, P.; Rodríguez-González, D.; García-Herrera, R.; Hernández-Ávalos, I.; et al. The Challenge of Global Warming in Water Buffalo Farming: Physiological and Behavioral Aspects and Strategies to Face Heat Stress. Animals 2023, 13, 3103. https://doi.org/10.3390/ani13193103
Napolitano F, De Rosa G, Chay-Canul A, Álvarez-Macías A, Pereira AMF, Bragaglio A, Mora-Medina P, Rodríguez-González D, García-Herrera R, Hernández-Ávalos I, et al. The Challenge of Global Warming in Water Buffalo Farming: Physiological and Behavioral Aspects and Strategies to Face Heat Stress. Animals. 2023; 13(19):3103. https://doi.org/10.3390/ani13193103
Chicago/Turabian StyleNapolitano, Fabio, Giuseppe De Rosa, Alfonso Chay-Canul, Adolfo Álvarez-Macías, Alfredo M. F. Pereira, Andrea Bragaglio, Patricia Mora-Medina, Daniela Rodríguez-González, Ricardo García-Herrera, Ismael Hernández-Ávalos, and et al. 2023. "The Challenge of Global Warming in Water Buffalo Farming: Physiological and Behavioral Aspects and Strategies to Face Heat Stress" Animals 13, no. 19: 3103. https://doi.org/10.3390/ani13193103