
Cultivable Gut Microbiota in Synanthropic Bats: Shifts of Its Composition and Diversity Associated with Hibernation

 , ,
, ,  , and
, and 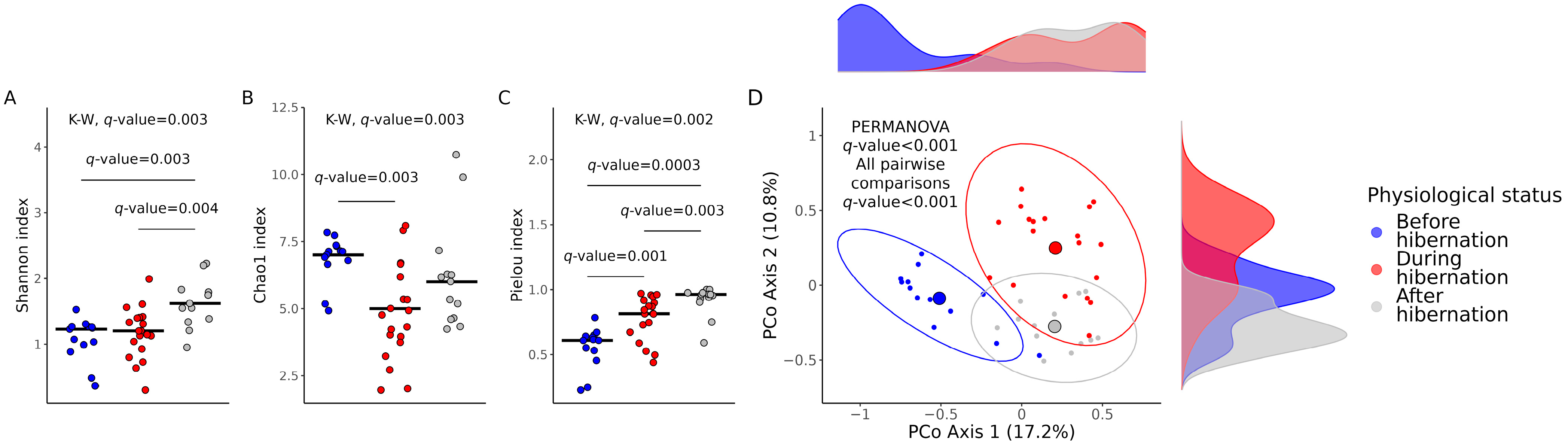{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Gut Bacteria Isolation, Counting, and Identification
2.3. Statistical Analyses
3. Results
3.1. Cultivable Gut Microbiota Diversity
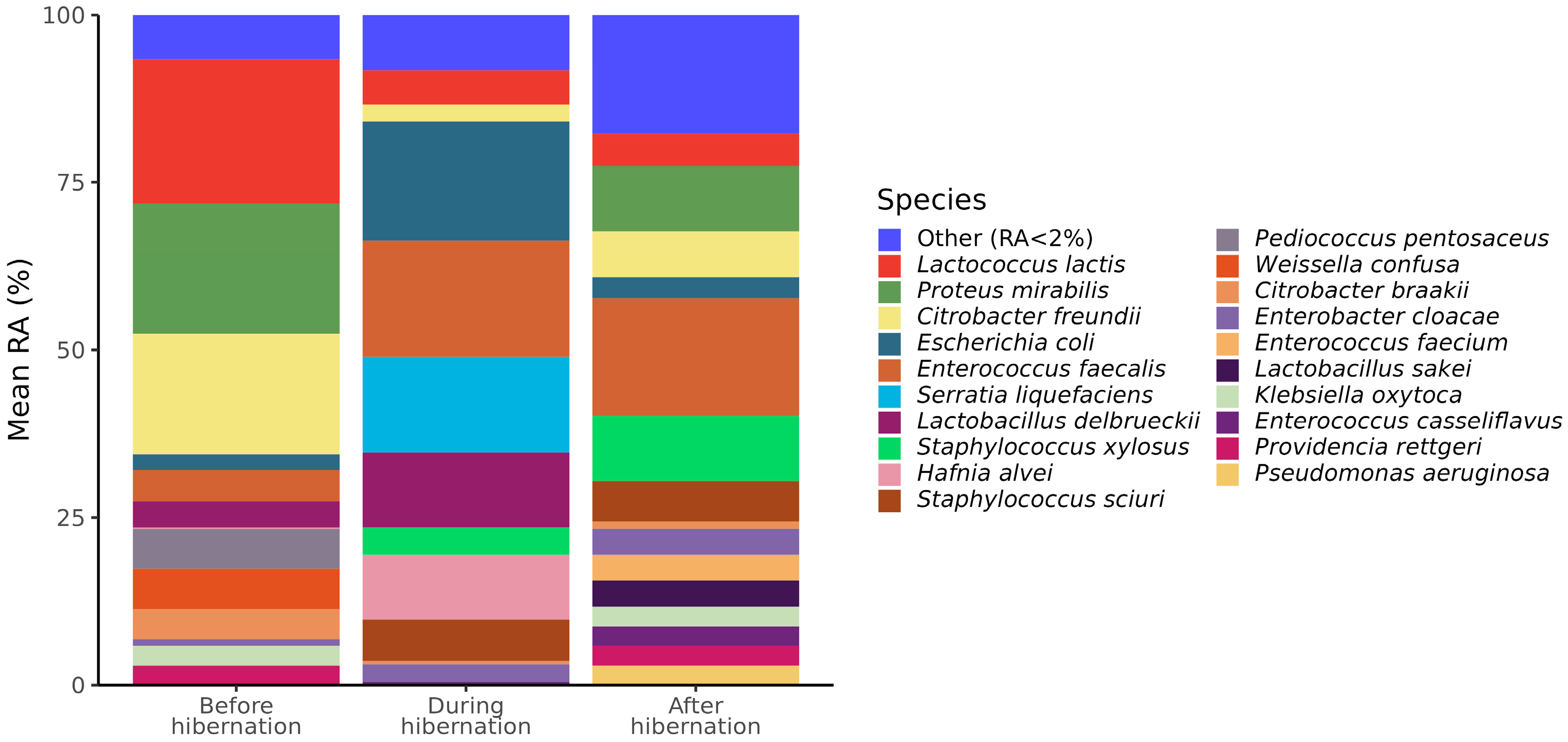
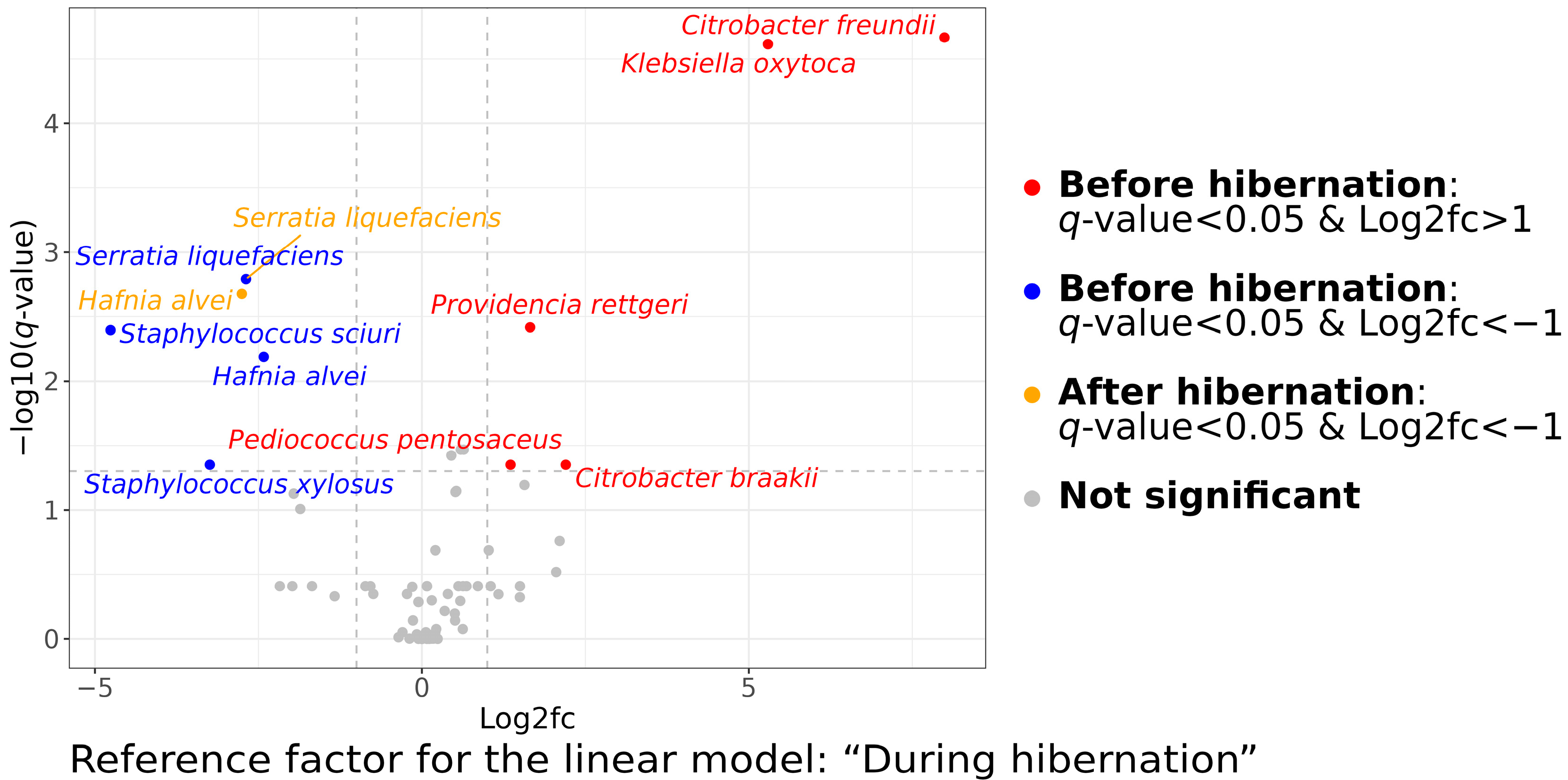
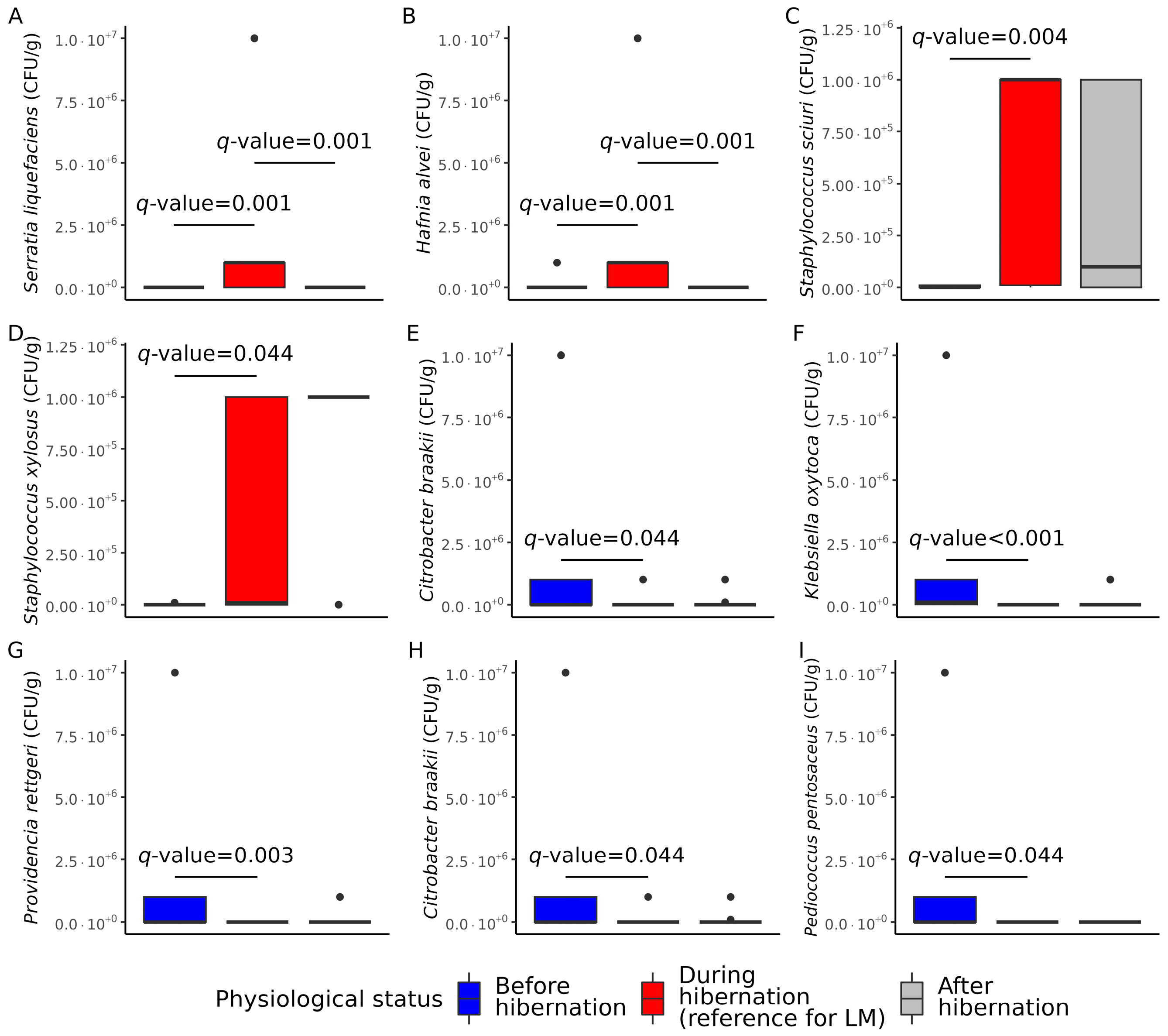
3.2. Cultivable Gut Microbiota Composition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bai, Y.; Urushadze, L.; Osikowicz, L.; McKee, C.; Kuzmin, I.; Kandaurov, A.; Babuadze, G.; Natradze, I.; Imnadze, P.; Kosoy, M. Molecular Survey of Bacterial Zoonotic Agents in Bats from the Country of Georgia (Caucasus). PLoS ONE 2017, 12, e0171175. [Google Scholar] [CrossRef] [PubMed]
- Subudhi, S.; Rapin, N.; Misra, V. Immune System Modulation and Viral Persistence in Bats: Understanding Viral Spillover. Viruses 2019, 11, 192. [Google Scholar] [CrossRef]
- Jebb, D.; Huang, Z.; Pippel, M.; Hughes, G.M.; Lavrichenko, K.; Devanna, P.; Winkler, S.; Jermiin, L.S.; Skirmuntt, E.C.; Katzourakis, A.; et al. Six reference-quality genomes reveal evolution of bat adaptations. Nature 2020, 583, 578–584. [Google Scholar] [CrossRef]
- Scheben, A.; Mendivil Ramos, O.; Kramer, M.; Goodwin, S.; Oppenheim, S.; Becker, D.J.; Schatz, M.C.; Simmons, N.B.; Siepel, A.; McCombie, W.R. Long-Read Sequencing Reveals Rapid Evolution of Immunity- and Cancer-Related Genes in Bats. Genome Biol. Evol. 2023, 15, evad148. [Google Scholar] [CrossRef] [PubMed]
- Popov, I.V.; Mazanko, M.S.; Kulaeva, E.D.; Golovin, S.N.; Malinovkin, A.V.; Aleshukina, I.S.; Aleshukina, A.V.; Prazdnova, E.V.; Tverdokhlebova, T.I.; Chikindas, M.L.; et al. Gut microbiota of bats: Pro-mutagenic properties and possible frontiers in preventing emerging disease. Sci. Rep. 2021, 11, 21075. [Google Scholar] [CrossRef]
- Bonilla-Aldana, D.K.; Jimenez-Diaz, S.D.; Arango-Duque, J.S.; Aguirre-Florez, M.; Balbin-Ramon, G.J.; Paniz-Mondolfi, A.; Suarez, J.A.; Pachar, M.R.; Perez-Garcia, L.A.; Delgado-Noguera, L.A.; et al. Bats in ecosystems and their Wide spectrum of viral infectious potential threats: SARS-CoV-2 and other emerging viruses. Int. J. Infect. Dis. 2021, 102, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Larson, P.A.; Bartlett, M.L.; Garcia, K.; Chitty, J.; Balkema-Buschmann, A.; Towner, J.; Kugelman, J.; Palacios, G.; Sanchez-Lockhart, M. Genomic features of humoral immunity support tolerance model in Egyptian rousette bats. Cell Rep. 2021, 35, 109140. [Google Scholar] [CrossRef] [PubMed]
- McGuire, L.P.; Fuller, N.W.; Dzal, Y.A.; Haase, C.G.; Silas, K.A.; Willis, C.K.R.; Olson, S.H.; Lausen, C.L. Similar hibernation physiology in bats across broad geographic ranges. J. Comp. Physiol. B 2022, 192, 171–181. [Google Scholar] [CrossRef]
- Geiser, F. Hibernation. Curr. Biol. 2013, 23, R188–R193. [Google Scholar] [CrossRef]
- Carey, H.V.; Assadi-Porter, F.M. The Hibernator Microbiome: Host-Bacterial Interactions in an Extreme Nutritional Symbiosis. Annu. Rev. Nutr. 2017, 37, 477–500. [Google Scholar] [CrossRef]
- Sommer, F.; Stahlman, M.; Ilkayeva, O.; Arnemo, J.M.; Kindberg, J.; Josefsson, J.; Newgard, C.B.; Frobert, O.; Backhed, F. The Gut Microbiota Modulates Energy Metabolism in the Hibernating Brown Bear Ursus arctos. Cell Rep. 2016, 14, 1655–1661. [Google Scholar] [CrossRef] [PubMed]
- Greene, L.K.; Andriambeloson, J.B.; Rasoanaivo, H.A.; Yoder, A.D.; Blanco, M.B. Variation in gut microbiome structure across the annual hibernation cycle in a wild primate. FEMS Microbiol. Ecol. 2022, 98, fiac070. [Google Scholar] [CrossRef] [PubMed]
- Dill-McFarland, K.A.; Neil, K.L.; Zeng, A.; Sprenger, R.J.; Kurtz, C.C.; Suen, G.; Carey, H.V. Hibernation alters the diversity and composition of mucosa-associated bacteria while enhancing antimicrobial defence in the gut of 13-lined ground squirrels. Mol. Ecol. 2014, 23, 4658–4669. [Google Scholar] [CrossRef]
- Russo, D.; Ancillotto, L. Sensitivity of bats to urbanization: A review. Mamm. Biol. 2015, 80, 205–212. [Google Scholar] [CrossRef]
- Nunes, H.; Rocha, F.L.; Cordeiro-Estrela, P. Bats in urban areas of Brazil: Roosts, food resources and parasites in disturbed environments. Urban. Ecosyst. 2017, 20, 953–969. [Google Scholar] [CrossRef] [PubMed]
- Ayivor, J.S.; Ohemeng, F.; Tweneboah Lawson, E.; Waldman, L.; Leach, M.; Ntiamoa-Baidu, Y. Living with Bats: The Case of Ve Golokuati Township in the Volta Region of Ghana. J. Environ. Public. Health 2017, 2017, 5938934. [Google Scholar] [CrossRef] [PubMed]
- Brito, J.E.C.; de Mello, B.G.V.; Gaeta, N.C.; Batista, J.M.N.; Brito, T.R.; Agostinho, W.C.; Brandao, P.E.; Heinemann, M.B.; Dias, R.A. Bats (Mammalia: Chiroptera) in urban-rural interfaces: Community structure associated with pathogen screening in Sao Paulo-the largest metropolitan region in Brazil. Vet. Res. Commun. 2023, 47, 1575–1600. [Google Scholar] [CrossRef]
- Xiao, G.; Liu, S.; Xiao, Y.; Zhu, Y.; Zhao, H.; Li, A.; Li, Z.; Feng, J. Seasonal Changes in Gut Microbiota Diversity and Composition in the Greater Horseshoe Bat. Front. Microbiol. 2019, 10, 2247. [Google Scholar] [CrossRef]
- Liu, S.; Xiao, Y.; Wang, X.; Guo, D.; Wang, Y.; Wang, Y. Effects of Microhabitat Temperature Variations on the Gut Microbiotas of Free-Living Hibernating Animals. Microbiol. Spectr. 2023, 11, e0043323. [Google Scholar] [CrossRef]
- Oksanen, J.; Simpson, G.; Blanchet, F.; Kindt, R.; Legendre, P.; Minchin, P.; O’Hara, R.; Solymos, P.; Stevens, M.; Szoecs, E.; et al. Vegan: Community Ecology Package, R package Version 2.6-5. 2023. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 23 November 2023).
- Mallick, H.; Rahnavard, A.; McIver, L.J.; Ma, S.; Zhang, Y.; Nguyen, L.H.; Tickle, T.L.; Weingart, G.; Ren, B.; Schwager, E.H.; et al. Multivariable association discovery in population-scale meta-omics studies. PLoS Comput. Biol. 2021, 17, e1009442. [Google Scholar] [CrossRef]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer-Verlag: New York, NY, USA, 2016. [Google Scholar]
- Kubista, C.E.; Bruckner, A. Importance of urban trees and buildings as daytime roosts for bats. Biologia 2015, 70, 1545–1552. [Google Scholar] [CrossRef]
- Voigt, C.C.; Phelps, K.L.; Aguirre, L.F.; Corrie Schoeman, M.; Vanitharani, J.; Zubaid, A. Bats and Buildings: The Conservation of Synanthropic Bats. In Bats in the Anthropocene: Conservation of Bats in a Changing World; Springer: Berlin/Heidelberg, Germany, 2015; pp. 427–462. [Google Scholar] [CrossRef]
- Salinas-Ramos, V.B.; Tomassini, A.; Ferrari, F.; Boga, R.; Russo, D. Admittance to Wildlife Rehabilitation Centres Points to Adverse Effects of Climate Change on Insectivorous Bats. Biology 2023, 12, 543. [Google Scholar] [CrossRef]
- Minor, R.L.; Dopfer, D.; Lemley, E.M.; Thurber, M.I. Factors Impacting Successful Rehabilitation of Big Brown Bats (Eptesicus fuscus) in a Wisconsin Wildlife Rehabilitation Center: A 5-Year Retrospective. J. Zoo. Wildl. Med. 2023, 54, 32–39. [Google Scholar] [CrossRef]
- Camacho, M.; Hernandez, J.M.; Lima-Barbero, J.F.; Hofle, U. Use of wildlife rehabilitation centres in pathogen surveillance: A case study in white storks (Ciconia ciconia). Prev. Vet. Med. 2016, 130, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.; Bell, T.; Coleman, C.M.; Harris, D.; Woodward, G.; Worledge, L.; Roberts, H.; McElhinney, L.; Aegerter, J.; Ransome, E.; et al. Testing bats in rehabilitation for SARS-CoV-2 before release into the wild. Conserv. Sci. Pract. 2022, 4, e12707. [Google Scholar] [CrossRef] [PubMed]
- Popov, I.V.; Ohlopkova, O.V.; Donnik, I.M.; Zolotukhin, P.V.; Umanets, A.; Golovin, S.N.; Malinovkin, A.V.; Belanova, A.A.; Lipilkin, P.V.; Lipilkina, T.A.; et al. Detection of coronaviruses in insectivorous bats of Fore-Caucasus, 2021. Sci. Rep. 2023, 13, 2306. [Google Scholar] [CrossRef]
- Alkhovsky, S.; Lenshin, S.; Romashin, A.; Vishnevskaya, T.; Vyshemirsky, O.; Bulycheva, Y.; Lvov, D.; Gitelman, A. SARS-like Coronaviruses in Horseshoe Bats (Rhinolophus spp.) in Russia, 2020. Viruses 2022, 14, 113. [Google Scholar] [CrossRef]
- Lenshin, S.V.; Romashin, A.V.; Vyshemirsky, O.I.; Lvov, D.K.; Alkhovsky, S.V. Bats of the subtropical climate zone of the Krasnodar Territory of Russia as a possible reservoir of zoonotic viral infections. Vopr. Virusol. 2021, 66, 112–122. [Google Scholar] [CrossRef]
- Speranskaya, A.S.; Artiushin, I.V.; Samoilov, A.E.; Korneenko, E.V.; Khabudaev, K.V.; Ilina, E.N.; Yusefovich, A.P.; Safonova, M.V.; Dolgova, A.S.; Gladkikh, A.S.; et al. Identification and Genetic Characterization of MERS-Related Coronavirus Isolated from Nathusius’ Pipistrelle (Pipistrellus nathusii) near Zvenigorod (Moscow Region, Russia). Int. J. Environ. Res. Public. Health 2023, 20, 3702. [Google Scholar] [CrossRef] [PubMed]
- Letko, M.; Seifert, S.N.; Olival, K.J.; Plowright, R.K.; Munster, V.J. Bat-borne virus diversity, spillover and emergence. Nat. Rev. Microbiol. 2020, 18, 461–471. [Google Scholar] [CrossRef]
- Luo, J.; Liang, S.; Jin, F. Gut microbiota in antiviral strategy from bats to humans: A missing link in COVID-19. Sci. China Life Sci. 2021, 64, 942–956. [Google Scholar] [CrossRef]
- Hoyt, J.R.; Kilpatrick, A.M.; Langwig, K.E. Ecology and impacts of white-nose syndrome on bats. Nat. Rev. Microbiol. 2021, 19, 196–210. [Google Scholar] [CrossRef]
- Garzoli, L.; Riccucci, M.; Patriarca, E.; Debernardi, P.; Boggero, A.; Pecoraro, L.; Picco, A.M. First Isolation of Pseudogymnoascus destructans, the Fungal Causative Agent of White-Nose Disease, in Bats from Italy. Mycopathologia 2019, 184, 637–644. [Google Scholar] [CrossRef]
- Garzoli, L.; Bozzetta, E.; Varello, K.; Cappelleri, A.; Patriarca, E.; Debernardi, P.; Riccucci, M.; Boggero, A.; Girometta, C.; Picco, A.M. White-Nose Syndrome Confirmed in Italy: A Preliminary Assessment of Its Occurrence in Bat Species. J. Fungi 2021, 7, 192. [Google Scholar] [CrossRef]
- Lemieux-Labonte, V.; Simard, A.; Willis, C.K.R.; Lapointe, F.J. Enrichment of beneficial bacteria in the skin microbiota of bats persisting with white-nose syndrome. Microbiome 2017, 5, 115. [Google Scholar] [CrossRef]
- Lemieux-Labonte, V.; Dorville, N.A.S.; Willis, C.K.R.; Lapointe, F.J. Antifungal Potential of the Skin Microbiota of Hibernating Big Brown Bats (Eptesicus fuscus) Infected With the Causal Agent of White-Nose Syndrome. Front. Microbiol. 2020, 11, 1776. [Google Scholar] [CrossRef] [PubMed]
- Gerbacova, K.; Malinicova, L.; Kiskova, J.; Maslisova, V.; Uhrin, M.; Pristas, P. The Faecal Microbiome of Building-Dwelling Insectivorous Bats (Myotis myotis and Rhinolophus hipposideros) also Contains Antibiotic-Resistant Bacterial Representatives. Curr. Microbiol. 2020, 77, 2333–2344. [Google Scholar] [CrossRef]
- Huang, Y.Y.; Sun, Y.M.; Huang, Q.N.; Lv, X.L.; Pu, J.; Zhu, W.T.; Lu, S.; Jin, D.; Liu, L.Y.; Shi, Z.L.; et al. The Threat of Potentially Pathogenic Bacteria in the Feces of Bats. Microbiol. Spectr. 2022, 10, e01802-22. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Vivas, J. Microbiology of Hafnia alvei. Enferm. Infecc. Microbiol. Clin. 2020, 38 (Suppl. 1), 1–6. [Google Scholar] [CrossRef] [PubMed]
- Stepanovic, S.; Dimitrijevic, V.; Vukovic, D.; Dakic, I.; Savic, B.; Svabic-Vlahovic, M. Staphylococcus sciuri as a part of skin, nasal and oral flora in healthy dogs. Vet. Microbiol. 2001, 82, 177–185. [Google Scholar] [CrossRef]
- Nemeghaire, S.; Argudin, M.A.; Fessler, A.T.; Hauschild, T.; Schwarz, S.; Butaye, P. The ecological importance of the Staphylococcus sciuri species group as a reservoir for resistance and virulence genes. Vet. Microbiol. 2014, 171, 342–356. [Google Scholar] [CrossRef] [PubMed]
- Vela, J.; Hildebrandt, K.; Metcalfe, A.; Rempel, H.; Bittman, S.; Topp, E.; Diarra, M. Characterization of Staphylococcus xylosus isolated from broiler chicken barn bioaerosol. Poult. Sci. 2012, 91, 3003–3012. [Google Scholar] [CrossRef]
- Liu, L.; Chen, D.; Liu, L.; Lan, R.; Hao, S.; Jin, W.; Sun, H.; Wang, Y.; Liang, Y.; Xu, J. Genetic Diversity, Multidrug Resistance, and Virulence of Citrobacter freundii From Diarrheal Patients and Healthy Individuals. Front. Cell Infect. Microbiol. 2018, 8, 233. [Google Scholar] [CrossRef] [PubMed]
- Singh, L.; Cariappa, M.P.; Kaur, M. Klebsiella oxytoca: An emerging pathogen? Med. J. Armed Forces India 2016, 72, S59–S61. [Google Scholar] [CrossRef] [PubMed]
- Sagar, S.; Narasimhaswamy, N.; D’Souza, J. Providencia Rettgeri: An Emerging Nosocomial Uropathogen in an Indwelling Urinary Catheterised Patient. J. Clin. Diagn. Res. 2017, 11, DD01–DD02. [Google Scholar] [CrossRef] [PubMed]
- Oyeka, M.; Antony, S. Citrobacter braakii Bacteremia: Case Report and Review of the Literature. Infect. Disord. Drug Targets 2017, 17, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Ni, L.; Xu, Y.; Chen, L. First Experimental Evidence for the Presence of Potentially Virulent Klebsiella oxytoca in 14 Species of Commonly Consumed Aquatic Animals, and Phenotyping and Genotyping of K. oxytoca Isolates. Antibiotics 2021, 10, 1235. [Google Scholar] [CrossRef]
- Wie, S.H. Clinical significance of Providencia bacteremia or bacteriuria. Korean J. Intern. Med. 2015, 30, 167–169. [Google Scholar] [CrossRef]
- Liu, L.; Lan, R.; Liu, L.; Wang, Y.; Zhang, Y.; Wang, Y.; Xu, J. Antimicrobial Resistance and Cytotoxicity of Citrobacter spp. in Maanshan Anhui Province, China. Front. Microbiol. 2017, 8, 1357. [Google Scholar] [CrossRef]
- Aminharati, F.; Ehrampoush, M.H.; Soltan Dallal, M.M.; Yaseri, M.; Dehghani Tafti, A.A.; Rajabi, Z. Citrobacter freundii Foodborne Disease Outbreaks Related to Environmental Conditions in Yazd Province, Iran. Iran. J. Public. Health 2019, 48, 1099–1105. [Google Scholar] [CrossRef]
- Gadelha-Alves, R.; Rozensztranch, A.M.D.; Rocha-Barbosa, O. Comparative Intestinal Histomorphology of Five Species of Phyllostomid Bats (Phyllostomidae, Microchiroptera): Ecomorphological Relations with Alimentary Habits. Int. J. Morphol. 2008, 26, 591–602. [Google Scholar] [CrossRef]
- Donaldson, G.P.; Lee, S.M.; Mazmanian, S.K. Gut biogeography of the bacterial microbiota. Nat. Rev. Microbiol. 2016, 14, 20–32. [Google Scholar] [CrossRef] [PubMed]
- Caviedes-Vidal, E.; McWhorter, T.J.; Lavin, S.R.; Chediack, J.G.; Tracy, C.R.; Karasov, W.H. The digestive adaptation of flying vertebrates: High intestinal paracellular absorption compensates for smaller guts. Proc. Natl. Acad. Sci. USA 2007, 104, 19132–19137. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Cai, L.; Lv, L.; Li, L. Pediococcus pentosaceus, a future additive or probiotic candidate. Microb. Cell Fact. 2021, 20, 45. [Google Scholar] [CrossRef] [PubMed]
- Demin, K.A.; Refeld, A.G.; Bogdanova, A.A.; Prazdnova, E.V.; Popov, I.V.; Kutsevalova, O.Y.; Ermakov, A.M.; Bren, A.B.; Rudoy, D.V.; Chistyakov, V.A.; et al. Mechanisms of Candida Resistance to Antimycotics and Promising Ways to Overcome It: The Role of Probiotics. Probiotics Antimicrob. Proteins 2021, 13, 926–948. [Google Scholar] [CrossRef]
- Durazzi, F.; Sala, C.; Castellani, G.; Manfreda, G.; Remondini, D.; De Cesare, A. Comparison between 16S rRNA and shotgun sequencing data for the taxonomic characterization of the gut microbiota. Sci. Rep. 2021, 11, 3030. [Google Scholar] [CrossRef]
- Nearing, J.T.; Douglas, G.M.; Hayes, M.G.; MacDonald, J.; Desai, D.K.; Allward, N.; Jones, C.M.A.; Wright, R.J.; Dhanani, A.S.; Comeau, A.M.; et al. Microbiome differential abundance methods produce different results across 38 datasets. Nat. Commun. 2022, 13, 342. [Google Scholar] [CrossRef]
- Yang, L.; Chen, J. A comprehensive evaluation of microbial differential abundance analysis methods: Current status and potential solutions. Microbiome 2022, 10, 130. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Popov, I.V.; Berezinskaia, I.S.; Popov, I.V.; Martiusheva, I.B.; Tkacheva, E.V.; Gorobets, V.E.; Tikhmeneva, I.A.; Aleshukina, A.V.; Tverdokhlebova, T.I.; Chikindas, M.L.; et al. Cultivable Gut Microbiota in Synanthropic Bats: Shifts of Its Composition and Diversity Associated with Hibernation. Animals 2023, 13, 3658. https://doi.org/10.3390/ani13233658
Popov IV, Berezinskaia IS, Popov IV, Martiusheva IB, Tkacheva EV, Gorobets VE, Tikhmeneva IA, Aleshukina AV, Tverdokhlebova TI, Chikindas ML, et al. Cultivable Gut Microbiota in Synanthropic Bats: Shifts of Its Composition and Diversity Associated with Hibernation. Animals. 2023; 13(23):3658. https://doi.org/10.3390/ani13233658
Chicago/Turabian StylePopov, Igor V., Iraida S. Berezinskaia, Ilia V. Popov, Irina B. Martiusheva, Elizaveta V. Tkacheva, Vladislav E. Gorobets, Iuliia A. Tikhmeneva, Anna V. Aleshukina, Tatiana I. Tverdokhlebova, Michael L. Chikindas, and et al. 2023. "Cultivable Gut Microbiota in Synanthropic Bats: Shifts of Its Composition and Diversity Associated with Hibernation" Animals 13, no. 23: 3658. https://doi.org/10.3390/ani13233658
APA StylePopov, I. V., Berezinskaia, I. S., Popov, I. V., Martiusheva, I. B., Tkacheva, E. V., Gorobets, V. E., Tikhmeneva, I. A., Aleshukina, A. V., Tverdokhlebova, T. I., Chikindas, M. L., Venema, K., & Ermakov, A. M. (2023). Cultivable Gut Microbiota in Synanthropic Bats: Shifts of Its Composition and Diversity Associated with Hibernation. Animals, 13(23), 3658. https://doi.org/10.3390/ani13233658