1. Introduction
Red sea bream iridovirus (RSIV) is a viral disease causing significant economic damage to the aquaculture industry [
1]. RSIV was first discovered in Japanese red sea bream (
Pagrus major) farms in 1990 and is the causative agent of red sea bream iridoviral disease (RSIVD) [
2], one of the reportable diseases currently managed by the World Organization for Animal Health (WOAH) [
1]. RSIV has been primarily reported in East and Southeast Asian countries [
3,
4,
5,
6,
7], and in South Korea, it was first discovered in 1998 in rock bream (
Oplegnathus fasciatus), where it caused substantial mortality every summer in water temperatures of 25 °C or higher [
8,
9]. Since then, RSIV has been detected in more than 30 fish species, and the range of susceptible species is expanding continuously [
10]. The major infection sites of RSIV have been reported to be the spleen and kidney, and histopathological lesions are characterized by abnormally enlarged cells [
10,
11,
12].
Horizontal transmission of RSIV has been demonstrated in previous studies and poses a serious problem for the aquaculture industry [
13]. The primary infection route of RSIV is considered to be horizontal transmission through seawater, and viral shedding following RSIV infection has recently been reported [
14,
15,
16]. Virus replication and shedding dynamics are key factors governing viral infectivity in the environment [
17,
18]. In particular, the waterborne transmission of RSIV infections among fish in net pen aquaculture systems sharing the same seawater has been reported in previous studies [
14]. The optimal temperature range for RSIV replication has been reported to be 20–25 °C in both in vivo and in vitro analyses [
19,
20]. However, RSIV shedding has been reported in rock bream artificially infected with RSIV at the lower susceptibility temperature of 15 °C, and RSIV has been detected in seawater from red sea bream farms at a low water temperature of 11.7 °C [
14,
15]. It has been reported that rock bream surviving 100 days post-RSIV infection still carry the virus [
20]. This suggests that the virus shed from infected fish could be a potential cause of the annual occurrence of RSIVD in the region. Recently, vaccines to prevent RSIV infection have been developed and commercialized; however, additional research is still required on their potential use in various fish species [
21]. In summary, for effective disease control and to prevent the spread of disease, it is important to understand viral transmission between hosts and to detect pathogens at an early stage.
The flathead grey mullet (
Mugil cephalus) is a major aquaculture species, with the third-largest aquaculture production in South Korea. Previous studies have reported the detection of RSIV in both wild and cultured flathead grey mullets [
22,
23,
24]. Although WOAH recognizes the flathead grey mullet as an RSIV-susceptible fish species [
1], no study has reported on the risk to other fish species through horizontal transmission or provided pathogenic information from RSIV infection experiments. In this study, mortality and viral shedding in seawater were confirmed through RSIV immersion infection in flathead grey mullets. In immersion-infected flathead grey mullets, histopathological scoring and infection grade criteria were defined according to RSIV infection kinetics. In addition, a cohabitation challenge model confirmed that horizontal transmission between species occurs in RSIV-infected flathead grey mullets through seawater. Our findings provide novel insights into the risk of RSIV-infected flathead grey mullets and can help in establishing measures to control RSIVD in fish farms.
2. Materials and Methods
2.1. Experimental Fish and Virus
The flathead grey mullets (total length: 10.2 ± 0.9 cm, weight: 9.2 ± 2.7 g), rock bream (total length: 10.4 ± 2.7 cm, weight: 22.7 ± 7.3 g), and red sea bream (total length: 9.7 ± 0.5 cm, weight: 13.7 ± 1.7 g) were purchased from hatcheries in Geoje and Namhae, Gyeongsangnam-do, Korea, where RSIVD was not reported; they were acclimated in a 1600 L tank for 2 weeks. Individual tanks (1600 L) of each fish were placed in a flow-through aquaculture system (500–1000 L/h) and continuously supplied with sand-filtered, 50 μm filter-housed, UV-treated (>30 mW/cm
2) seawater during acclimation. Water temperature was maintained at 25 ± 2 °C and commercial feed was provided twice, daily. Prior to experimentation, 15 fish from each fish species were randomly selected and confirmed to be free of RSIV by polymerase chain reaction (PCR) analysis as described in the Aquatic Animal Diagnostic Test Manual for WOAH and qPCR [
1,
25].
In this study, the RSIV genotype II (accession number: AY532608) was utilized [
15]. The virus was propagated using the
Pagrus major fin (PMF) cell line maintained at 25 °C with L-15 medium (Gibco, Billings, MT, USA) containing 10% fetal bovine serum (Gibco), 1% antibiotic-antimycotic (100 U/mL penicillin, 100 μg/mL streptomycin, and 25 μg/mL amphotericin B, Gibco) [
26]. RSIV was inoculated onto a confluent PMF cell monolayer and incubated at 25 °C for viral replication. Culture supernatants exhibiting complete cytopathic effects (CPE) were harvested and centrifuged at 10,000×
g for 10 min at 4 °C. Subsequently, the supernatant was collected, and the viral copy numbers were determined on the basis of the method described in a previous study [
25]. The virus was stored at −80 °C until further use.
2.2. Experimental Immersion Infection of Flathead Grey Mullets
All experimental protocols followed the guidelines set by the Institutional Animal Care and Use Committee of Gyeongsang National University (approval number: GNU-220526-E0056; GNU-220526-E0057; GNU-220526-E0058). Flathead grey mullets were reared in eight 50 L water tanks at 25 °C and 15 °C (
n = 30 in each group) for two weeks. Fish in the immersion challenge group were exposed to 10
5, 10
3, and 10
1 RSIV copies/mL (final infection concentration in the tank) for 150 min at 25 °C and 15 °C, respectively, after which the seawater was replaced with fresh seawater. The negative control group received no treatment (
n = 30 in each group). Mortality patterns were observed daily for 40 days, and tissues (spleen, gills, kidneys, heart, stomach, eyes, liver, intestines, brain, skin, and muscle) from the dead fish in each group were collected for viral load analysis. Half (50%) of the rearing seawater for each group was replaced daily with sand filtration treated seawater, 1 μm housing filter, and UV (>30 mW/cm
2). To determine the RSIV shedding kinetics, 500 mL of seawater (
n = 3 in each group) was collected from fish tanks used to measure mortality after RSIV immersion at 3, 5, 7, 10, 14, 21, 30, and 40 days post-infection (dpi), virus concentration was determined, and qPCR was performed as described in
Section 2.4.
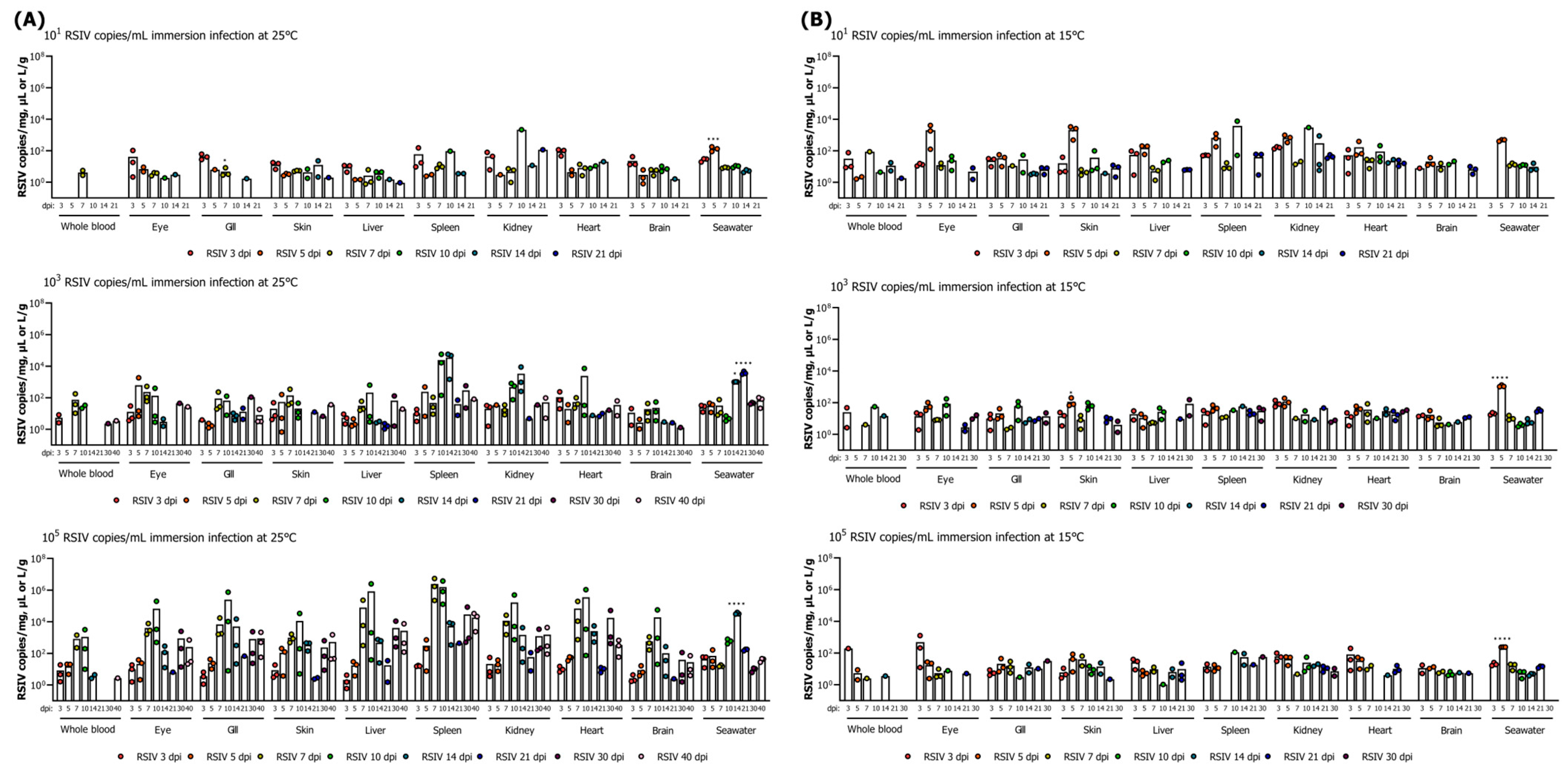
To investigate the dynamics in the tissues of flathead grey mullets after the immersion challenge, another set of experiments was conducted in the manner described above. After immersion challenge, three fish from each tank were sampled at 3, 5, 7, 10, 14, 21, 30, and 40 days and whole blood, eyes, gills, skin, liver, spleen, kidneys, heart, and brain were collected. The collected organs were stored individually at −80 °C for viral load determination and fixed in 10% neutral-buffered formalin for histopathological analysis.
2.3. Cohabitation Challenge
Before the RSIV cohabitation challenge, fish were reared in eight 100 L water tanks at 25 °C and 15 °C (
n = 30 in each group) for two weeks. To confirm the horizontal transmission of the virus in RSIV-infected flathead grey mullets, fish were reared in cohabitation. Flathead grey mullets (donors,
n = 30 in each group) were injected intraperitoneally (IP) with RSIV (10
6 RSIV copies/fish), and naïve rock bream, red sea bream, and flathead grey mullets (recipients,
n = 30 in each group) cohabited with donors. In the negative control group, flathead grey mullets (
n = 30 in each group) injected with L-15 medium and naïve fish (
n = 30 in each group) cohabited. The mortality of each group was observed for 40 days. Donor and recipient fish were distinguishable owing to being reared in two cages that allowed smooth seawater flow within the tank. Half (50%) of the rearing seawater for each group was replaced daily with sand filtration treated seawater, 1 μm housing filter, and UV (>30 mW/cm
2). For the determination of RSIV shedding kinetics, 500 mL of seawater (
n = 3 in each group) was collected from the fish tanks measuring mortality after RSIV cohabitation challenge at 1, 3, 5, 7, 10, 14, 21, 30, and 40 dpi, virus concentration was determined, and qPCR was performed as described in
Section 2.4.
To investigate the dynamics in the tissues of flathead grey mullets after the immersion challenge, another set of experiments was conducted in the manner described above. Spleens and kidneys of fish (donors and recipients,
n = 3 in each group) were sampled as previously described on 1, 3, 5, 7, 10, 14, 21, 30, and 40 days after cohabitant infection to determine the RSIV load in the fish. For histopathological observation, spleen and kidney tissues sampled at 3, 5, 7, 10, 14, 21, 30, and 40 days after RSIV intraperitoneal (IP) injection in fish (donors and recipients,
n = 3 in each group) were fixed in 10% neutral formalin and analyzed as described in
Section 2.6.
2.4. Nucleic Acid Extraction and qPCR
Genomic DNA was extracted from fish tissues (25–50 mg) and blood (100 μL) using an AccuPrep
® Genomic DNA Extraction Kit (Bioneer, Daejeon, Republic of Korea), following the manufacturer’s instructions. The extracted DNA was assessed for RSIV copy number using a previously described TaqMan-based qPCR method [
25]. Briefly, the primer sets included Meg 1041F (5′-CCA CCA GAT GGG AGT AGA C-3′) forward primer, Meg 1139R (5′-GGT TGA TAT TGC CCA TGT CCA-3′) reverse primer, and Meg 1079P (5′-[FAM]-CCT ACT A[i-EBQ]CT TTG CGC CCA GCA TG-[phosphate]-3′) TaqMan probes [
25]. All qPCR assays were performed on a Dice
® Real Time System III (Takara, Kusatsu, Japan), with an initial denaturation of 1 min at 95 °C, followed by 45 cycles of 5 s at 95 °C and 10 s at 60 °C. For each sample, the RSIV qPCR composition consisted of 5 μL DNA, 12.5 μL HS Prime qPCR Premix with UDG (2×) (Genetbio, Daejeon, Republic of Korea), final concentrations of 900 nM forward and reverse primers, and 250 nM TaqMan probes. The cycle threshold (C
t) cut-off value for qPCR was set at 39.75, as described in a previous study [
25]. During qPCR analysis, a negative control (diethyl pyrocarbonate treated water) well was included to confirm the absence of false positives in the PCR reaction. After measuring the weight (mg) of each tissue, the viral copy number was calculated per mg of tissue following the viral quantification analysis [
25].
2.5. Virus Concentration Based on Iron Flocculation in Rearing Seawater
RSIV particles in rearing seawater were concentrated using an iron flocculation assay [
25,
27]. Initially, 500 mL of seawater was filtered through a 1.6 μm pore size glass microfiber filter (GF/A; Whatman, Maidstone, UK) to remove suspended particles. Subsequently, 4.83 g of iron (III) chloride hexahydrate (FeCl
3∙6H
2O) was dissolved in 100 mL of distilled water to form an iron chloride solution, and 50 μL of this solution was dispensed into the 500 mL of seawater to form Fe-virus flocculates. The seawater containing the iron chloride solution was then gently stirred at 20 °C (room temperature) for 1 h at 200 rpm using a magnetic stirrer. The Fe-RSIV flocculates were filtered under reduced pressure through a 0.8 μm pore size polycarbonate filter (Whatman) attached to a filter holder with a receiver (Nalgene, New York, NY, USA). The viruses collected on the filter were transferred to a 2 mL tube, and nucleic acids were extracted using an AccuPrep
® Genomic DNA Extraction Kit (Bioneer). The genomic copy numbers of the virus particles concentrated from seawater were determined using qPCR, as described in
Section 2.4. To determine the viral shedding ratio (RSIV copies/L/g) of RSIV-infected fish, the average number of viral genome copies in seawater was divided by the average weight of fish surviving in the tank. Seawater for the analysis of the viral shedding ratio of all fish was collected from the mortality group tanks.
2.6. Histopathological Analysis
Histopathological analyses were conducted on all dissected organs (brain, eye, gills, heart, kidney, liver, muscle, and spleen) of the fish samples. Each sample was fixed in 10% neutral-buffered formalin for 24 h. Following fixation, samples were collected and refixed in the same solution for another 24 h before undergoing gradual dehydration through an ethanol series (70–100%). Subsequently, the samples were cleared with xylene, embedded in paraffin, and sectioned into 4 μm-thick slices. The sections were then stained with hematoxylin-eosin (H&E) (BBC Biochemical, Washington, DC, USA) following standard protocols.
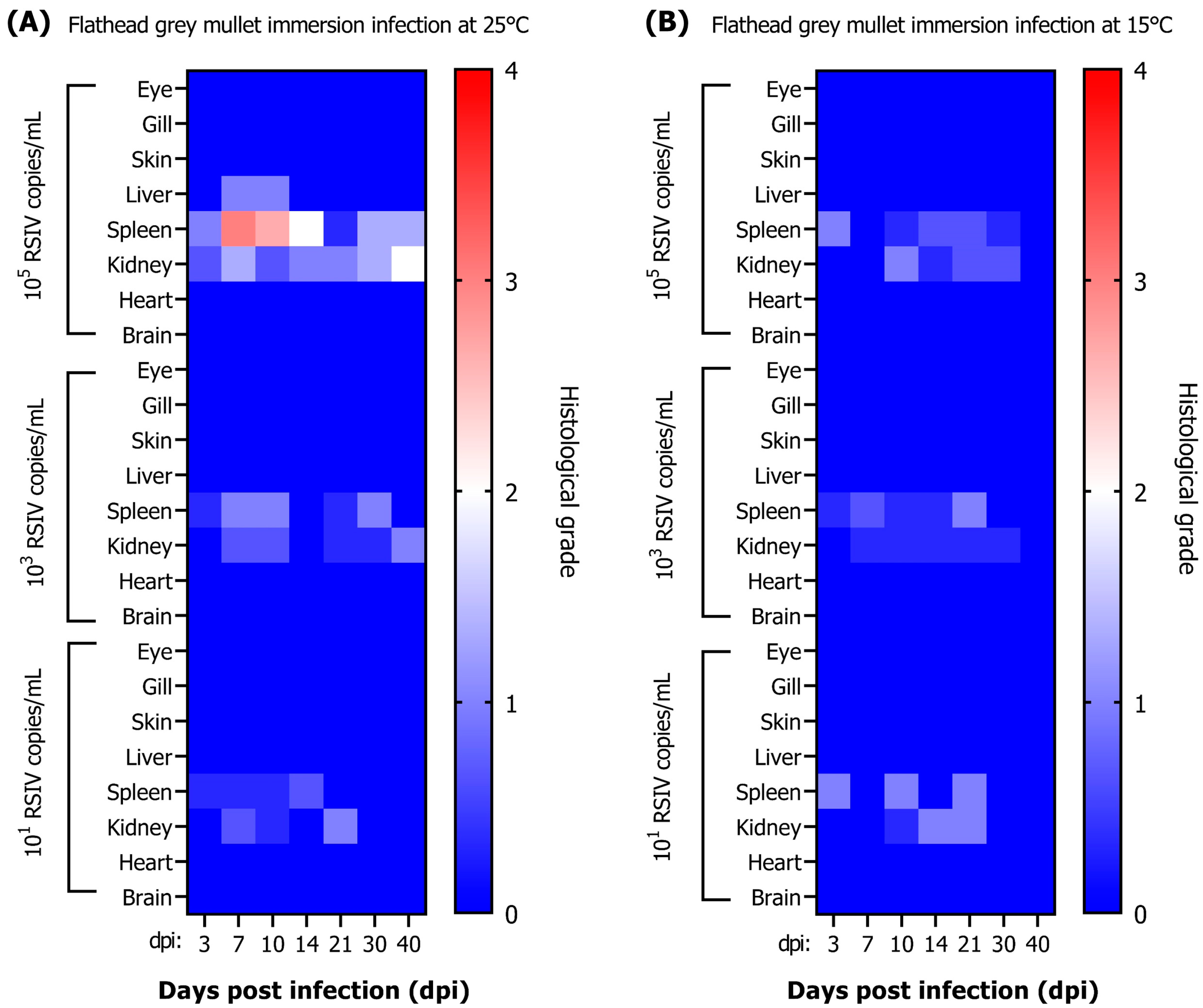
The scoring of RSIV infection was divided into four stages: (1) less, (2) mild, (3) moderate, and (4) severe. Each stage was scored in blind experiments conducted by a fish pathologist. Renal and splenic lesions were assessed in two categories: enlarged cells and necrotic lesions.
Enlarged cells in the spleen were scored from 1 to 4, depending on their frequency of occurrence in the parenchymal tissue. Enlarged cells in the kidney were evaluated based on the following criteria: score 1 (less) if present locally in renal tubules and parenchymal tissues; score 2 (mild) when a small number of cells were observed in the renal tubules, parenchymal tissues, and glomeruli. Necrotic lesions in the spleen were assessed based on these criteria: score 1 (less) for focal lesions involving a small number of cells; score 2 (mild) when numerous cells undergoing necrosis were identified; score 3 (moderate) when necrosis had progressed, cell exudate was observed, and inflammatory cell infiltration had occurred; and score 4 (severe) when necrosis had advanced, resulting in parenchymal tissue atrophy. Necrotic lesions in kidneys were evaluated based on the following criteria: score 1 (less) if localized and present in less than 5% of the renal tubule; score 2 (mild) if present locally and sporadically observed in 5% or more of the renal tubule; score 3 (moderate) if present in parenchymal tissues and glomeruli, in addition to renal tubules; and score 4 (severe) when tubule necrosis occurred due to necrotic lesions and atrophy resulting from parenchymal tissue and glomerular necrosis. The scoring standards are described in
Figure S1.
The grading of RSIV-infected samples was achieved by weighting scores according to lesion severity. In brief, enlarged cells representing general lesions of RSIV infection accounted for 70% of the grading, while RSIV-induced necrotic lesions, which had a lower correlation with viral load, contributed to 30% of the grading. Changes in other organs were recorded, but not graded, due to their insignificant correlation. The grading scale employed was delineated as follows: Grade 0 (G0) included values less than or equal to 0.2; Grade 1 (G1) included values greater than 0.2 and less than or equal to 0.8; Grade 2 (G2) included values greater than 0.8 and less than or equal to 1.5; Grade 3 (G3) included values greater than 1.5 and less than or equal to 2.0; and Grade 4 (G4) included values exceeding 2.0.
2.7. Statistical Analysis
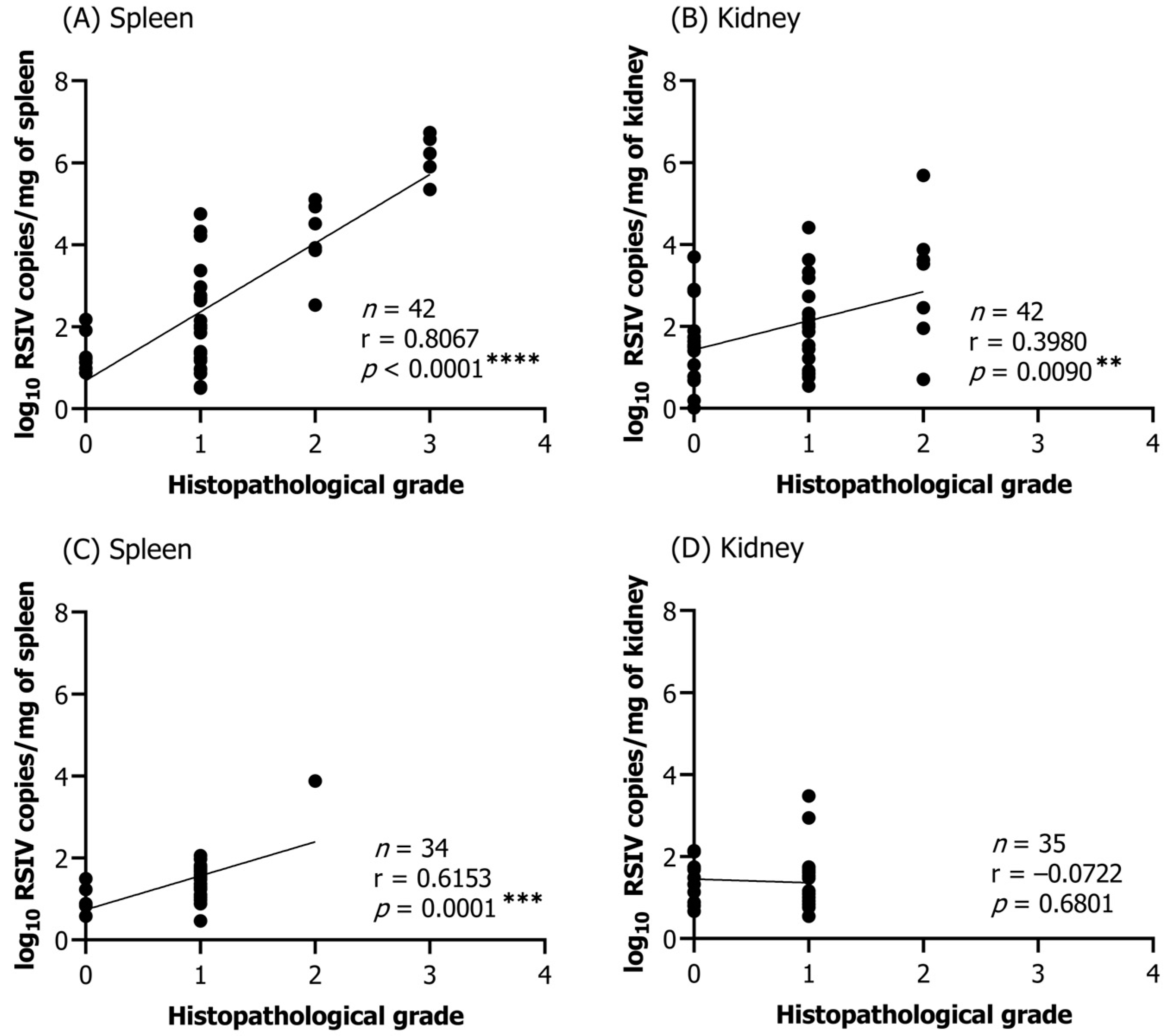
Statistical tests were performed using GraphPad Prism 9.5. An ordinary one-way analysis of variance (ANOVA) with Dunnett’s correction was performed when comparing multiple groups. Significant differences were compared with controls when the virus was first detected within each group. The correlation between histopathological infection grade and viral load was analyzed using Pearson correlation coefficients. Statistical significance was denoted by the following convention: * p < 0.05; ** p < 0.01; *** p < 0.001; **** p < 0.0001.
4. Discussion
The goal of this study was to demonstrate pathogenicity to mullet using immersion infection, similar to the mechanism of natural RSIV infection in fish farms. To understand the mechanisms of viral load, viral shedding, and histopathological lesions in RSIV-infected flathead grey mullets, challenge experiments were performed at various infection concentrations and water temperatures. In addition, a cohabitation challenge model was used to assess the risk of horizontal transmission of RSIV. To the best of our knowledge, this is the first report of RSIV kinetic analysis and cohabitant infection in flathead grey mullets.
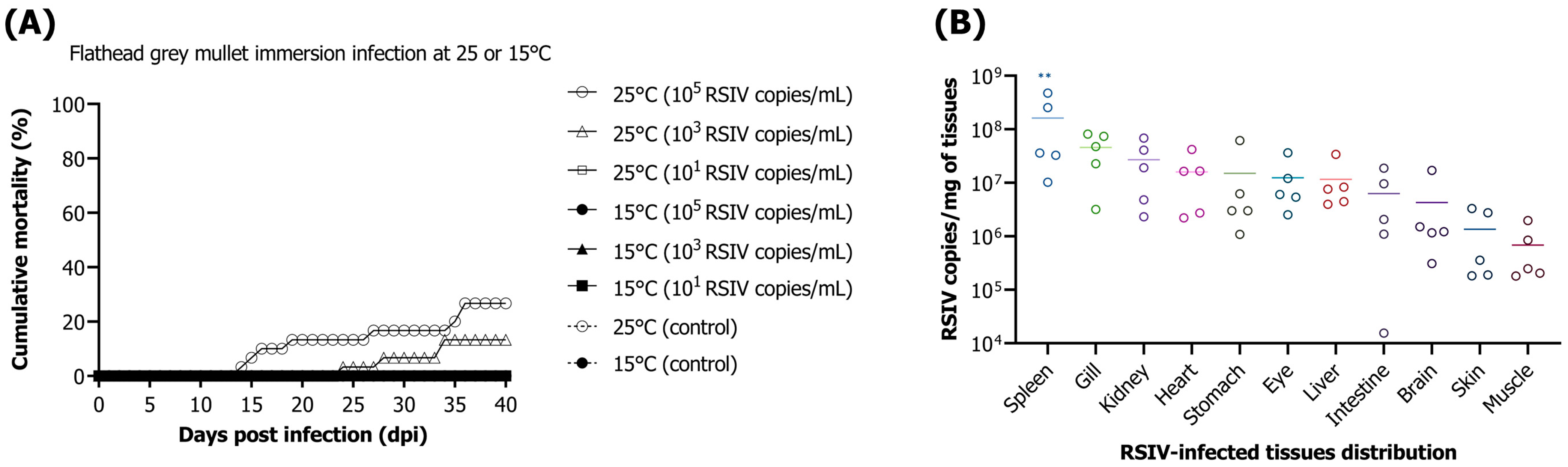
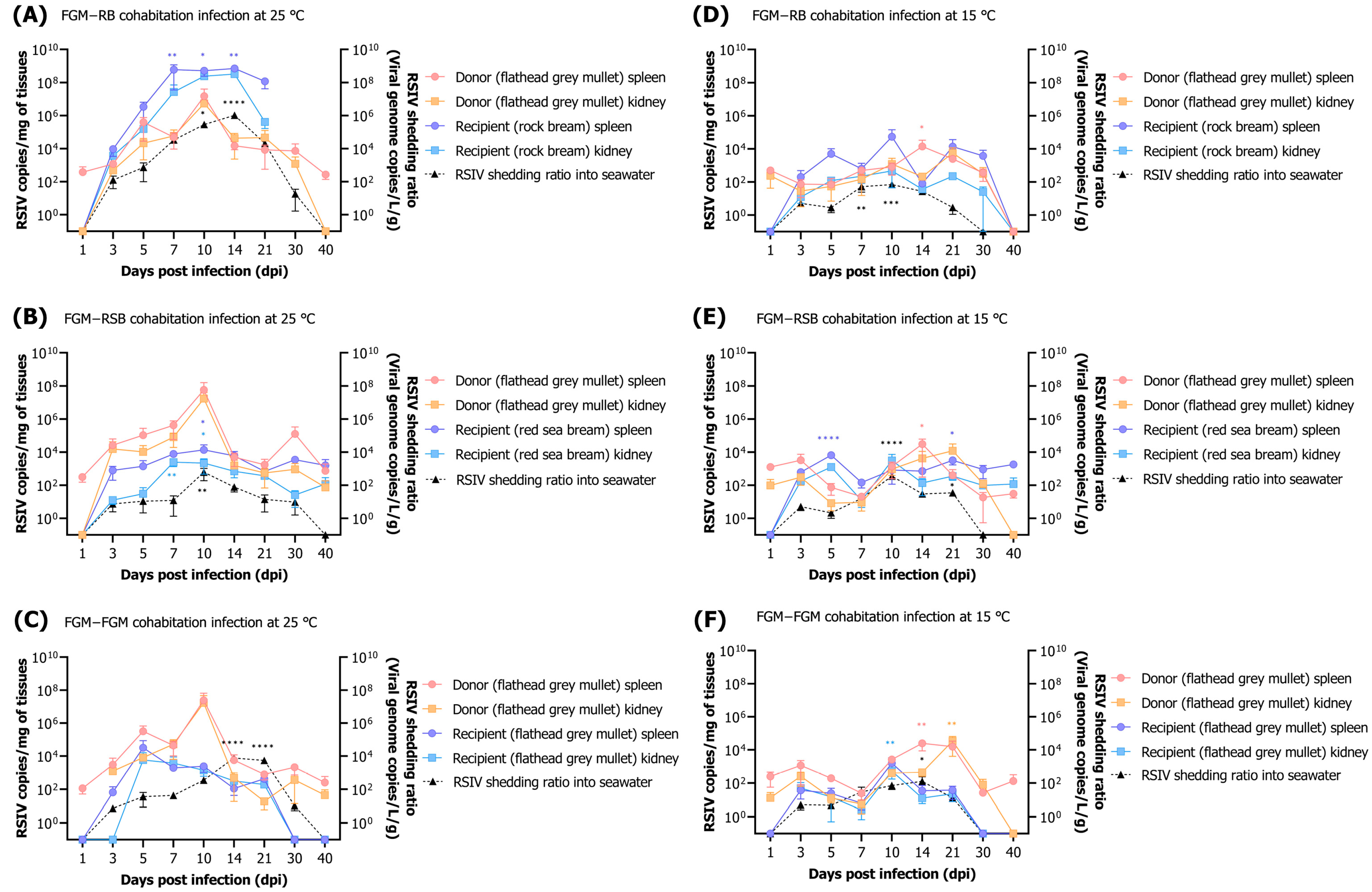
In the current study, the mortality of flathead grey mullets challenged by immersion infection was observed at concentrations of 10
5 and 10
3 RSIV copies/mL. Previous research has shown that Japanese amberjack (
Seriola quinqueradiata) subjected to immersion challenge with RSIV (10
2.7 TCID
50/mL) at 25 °C experienced a 40% mortality rate [
27]. In Pacific bluefin tuna (
Thunnus orientalis), a 44.9% mortality rate was reported when immersion-infected at 25 °C with a concentration of 6.0 × 10
3 RSIV copies/mL [
28]. In rock bream, 100% mortality occurred when challenged by immersion with 10
1, 10
3, 10
5, and 10
7 RSIV copies/mL [
16]. Therefore, mortality in fish challenged by immersion with 10
5 RSIV copies/mL in this study was the lowest at 26.6% compared with those reported in previous research [
16,
27,
28]. Susceptibility to RSIV may vary among species, but the impact of fish species with relatively low susceptibility to disease on other species can be significant in the aquaculture industry. RSIVD is known to occur mainly at water temperatures above 20 °C, with an optimal temperature of approximately 25 °C [
1,
15,
29]. In several studies, the highest viral shedding has been reported to occur just before or during periods of active mortality following exposure to the virus [
15,
16,
17,
27,
30]. In this study, a low mortality rate was observed following the RSIV immersion challenge; however, the peak of virus shedding was observed either during the period of mortality or 3–4 days prior to the occurrence of mortality. These observations suggest that the virus shed from flathead grey mullets may potentially cause horizontal transmission to other fish species. Interestingly, viral shedding was detected from 3 to 5 days in flathead grey mullets that did not exhibit mortality. A similar trend in dynamics was observed in rock bream challenged with RSIV at 15 °C without mortality [
15]. Additionally, in seven-band grouper (
Hyporthodus septemfasciatus) infected with nervous necrosis virus (NNV), viral shedding was detected within 24 h [
31], while in Japanese amberjack infected with RSIV, the virus was found at 3 dpi (10
2.5 RSIV genome copies/L) [
27]. In Atlantic salmon (
Salmo salar L.), excretion of salmonid alphavirus (SAV) through mucus and feces is known to begin immediately after infection and may persist for 3–4 weeks thereafter [
32]. Although this study did not analyze viral loads in feces and mucus, the virus was consistently detected during the initial stages of infection and at low water temperatures for 14–21 days. However, no differences were observed in the viral loads within the fish and the amounts of virus shed into the seawater, even at high RSIV infection concentrations in low water temperatures. While the precise conditions for virus release into seawater cannot be determined, the virus may be shed when an apparent infection is induced within the fish population. However, no RSIV was detected in seawater after 21–30 dpi in water temperatures at which no disease progression was observed. These results are consistent with those of previous studies that showed that viral shedding terminates as viral load declines and RSIV-infected rock bream recover at water temperatures as low as 15 °C [
15,
33]. In the present study, because the challenge experiments were performed in a closed aquaculture system rather than a flow-through aquaculture system, we may not have fully reproduced the disease progression in the aquaculture field. Despite replacing 50% of the tank seawater daily, it cannot be ruled out that the accumulation of virus shed from the fish may have occurred, potentially leading to an overestimation of viral shedding. However, in a previous study, even when a large amount of rearing seawater in the tank was flushed with fresh seawater after virus infection in fish, the time points for the viral load detection in the fish and the peak of viral load in the seawater were consistent [
27]. Our results are the first to demonstrate the establishment of RSIV infection in flathead grey mullets in an immersion infection model that mimics naturally occurring infections. However, conditions in a fish farm environment may differ from those of temporary immersion infection in a laboratory environment, as RSIV present in rearing seawater has the possibility of continuously infecting fish.
The spleen and kidneys are known to be the main target organs for RSIV, but RSIV can also be detected in the gills, heart, and intestines [
1,
25]. In our study, the highest viral load was observed in the spleen in dead flathead grey mullets by RSIV, followed by the gills, kidneys, and heart. In flathead grey mullets challenged by immersion infection at a water temperature of 25 °C (in which RSIV-susceptibility was observed), the RSIV load was the highest in the spleen among all organs tested during most stages from the initial infection to the recovery period. However, in the group without mortality or disease progression (10
1 RSIV copies/mL at 25 °C and all challenge groups at 15 °C), a low viral load was observed in all organs examined regardless of the RSIV target organ. Based on these results, our study strongly suggests that the spleen is the most suitable target organ for the detection of RSIV in flathead grey mullets under disease-progression conditions.
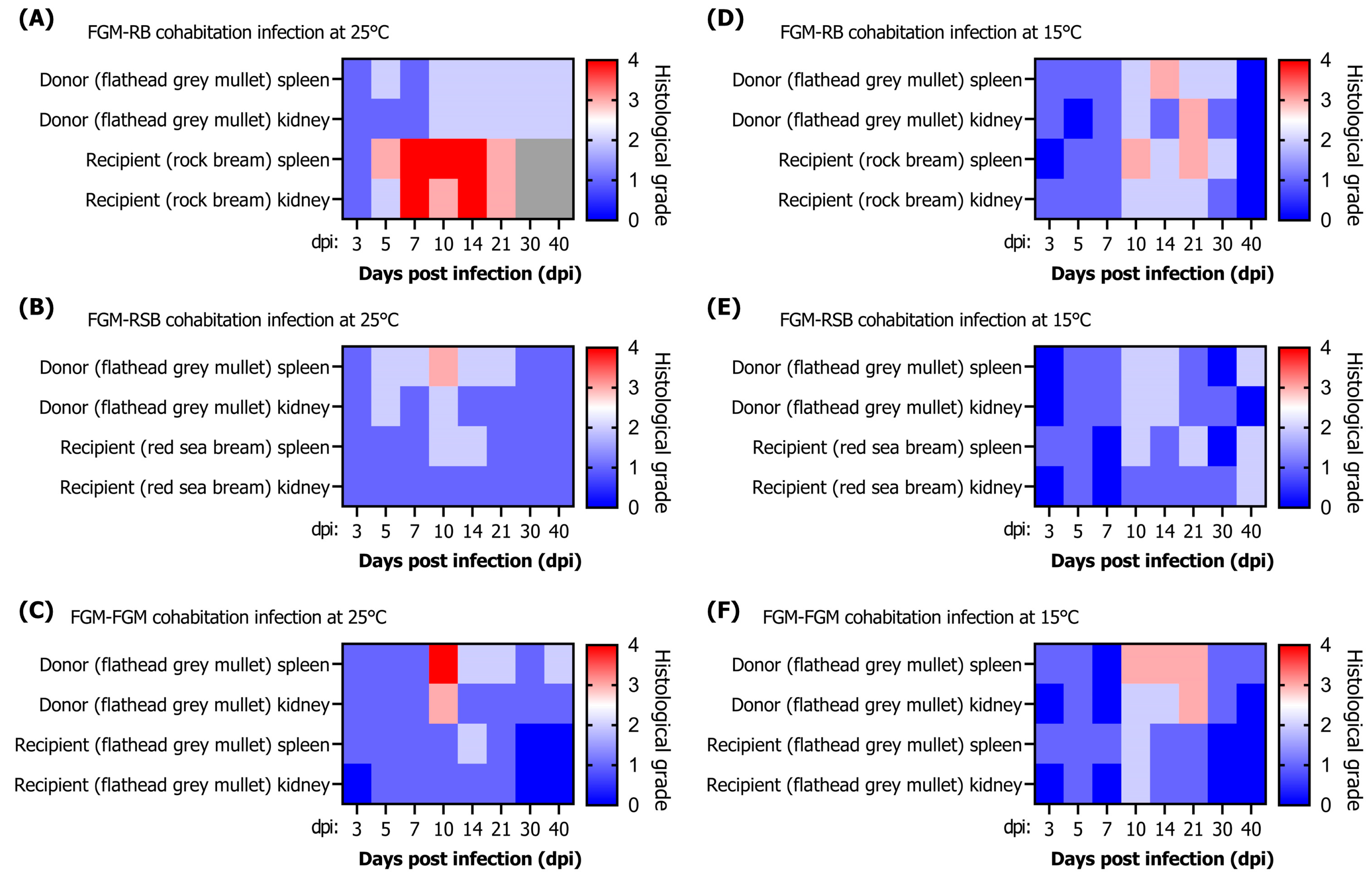
Histopathological lesions in fish can indicate the severity and progression of a disease. Several studies have investigated the correlation between viral loads and histopathological lesions. However, a positive PCR reaction does not necessarily imply ongoing disease progression in fish. In a previous study, RSIV was detected in cultured rockfish (
Sebastes schlegelii) by nested PCR, but no histopathological lesions were observed in the tissues [
34]. In another study, rock bream that survived RSIV infection were reported to still be detectable by PCR even after 100 days [
20]. In our study, histopathological lesions were primarily observed in the spleen and kidney, but some fish tissues with low viral loads did not exhibit enlarged cells or necrotic lesions. This may be due to insufficient initial replication time, inappropriate replication temperature, or inadequate viral infection capacity, resulting in no observable lesions. The formation of abnormally enlarged cells due to RSIV infection is frequently observed in the spleen of infected fish [
35]. In our study, a high correlation between viral load and lesions was confirmed in the spleen of infected flathead grey mullets. The parenchymal tissue of the spleen in fish with high viral loads was replaced by enlarged cells. Necrotic lesions tended to be observed earlier than enlarged cells as a more sensitive primary response at low viral loads. In contrast to the observations for the spleen, a low correlation between lesions in the kidney and viral load was demonstrated. The impact of abnormally enlarged cells on kidney function might be minimal, as these cells only appear on interstitial hematopoietic tissue and did not occupy the renal parenchymal tissue, which is consistent with previous studies [
2]. As there are few reports describing the histopathology of diseased flathead grey mullets, our findings could contribute to future research in fish pathology. Additionally, it is crucial to consider both viral loads and histopathological lesions in order to accurately assess disease status in infected fish.
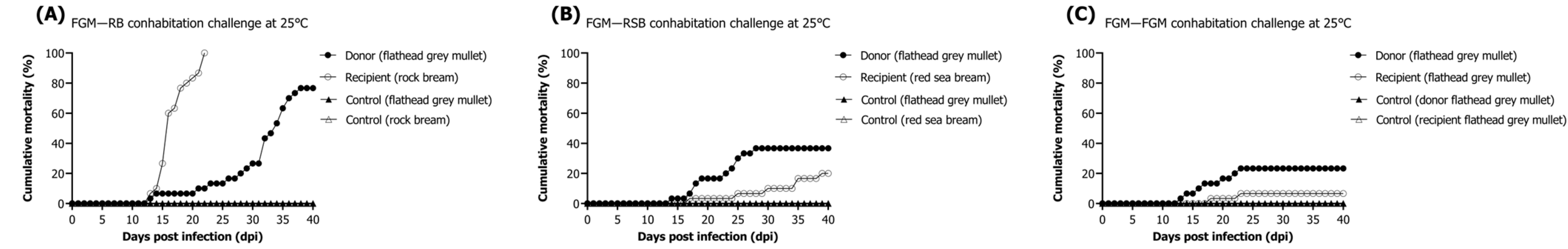
In previous studies, research on RSIV cohabitation challenges between rock bream and rockfish has been reported [
13]; however, there is no literature regarding flathead grey mullets. A cohabitation challenge model was used to assess whether RSIV-infected flathead grey mullets develop horizontal transmission to recipients. Mortality patterns and viral shedding kinetics differed according to the RSIV susceptibility of recipient species cohabiting with flathead grey mullets. The timing of the first mortality of mullet in each challenged group at 25 °C was similar, at 13–14 dpi. The highest cumulative mortality rate was observed in groups cohabitating with rock bream (recipients). These results are thought to be the cause of the high mortality and viral shedding rate, as the rock bream infected with the virus shed by the flathead grey mullet had the highest RSIV susceptibility [
1]. Nevertheless, in our findings, the viral load of flathead grey mullet (donor) was at a similar level in each group within 21–40 days post-infection at 25 °C, but the mortality rates differed. These discrepancies may be due to the tanks used for mortality measurement being different from those sampled for viral load measurement or because the collected live fish were gradually recovering from the disease. The viral load and mortality trends may have differed as a result. After the cohabitation challenge, all deceased fish exhibited an RSIV load higher than approximately 10
7 RSIV copies/mg (data not shown). Lower mortality and viral shedding ratios were observed in the red sea bream and flathead grey mullet groups (recipients). This is consistent with previous research findings suggesting that red sea bream has a lower susceptibility to RSIV compared with rock bream [
1]. Additionally, this aligns with our experience that, in Korean fish farms, mortality due to RSIVD is less frequently observed in these two species than in rock bream. Histopathological lesions were also observed in the cohabitation challenge experiment. Although the histopathological grading of donors and recipients did not show a perfect correlation with viral load, the infection grade followed as the viral load increased. In particular, flathead grey mullets infected at lower water temperatures shed the virus into the seawater, and horizontal transmission was confirmed as the virus was detected in healthy fish. Additionally, the progression of the disease was demonstrated through the observation of lesions within recipient fish tissues. These results suggest that the virus shed from RSIV-infected flathead grey mullets can move through seawater and damage nearby farmed fish such as rock bream, red sea bream, and flathead grey mullets, which are all farmed in open-net pens in Korea.

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






