
Taurine Supplementation to Plant-Based Diets Improves Lipid Metabolism in Senegalese Sole

 ,
,  ,
,  , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Diets
2.2. Rearing Trial
2.3. Metabolic Trial
2.4. Chemical and Biochemical Analysis
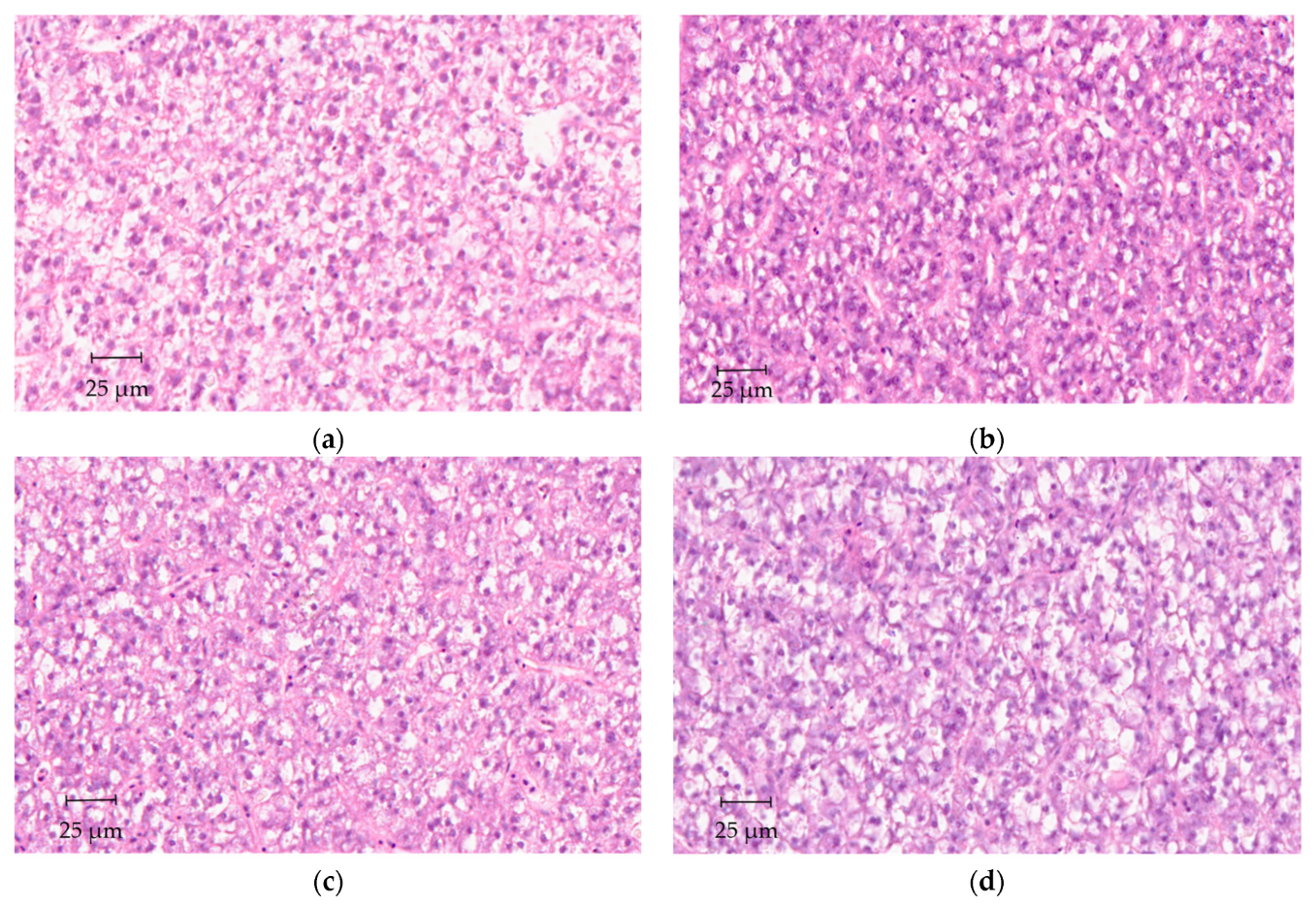
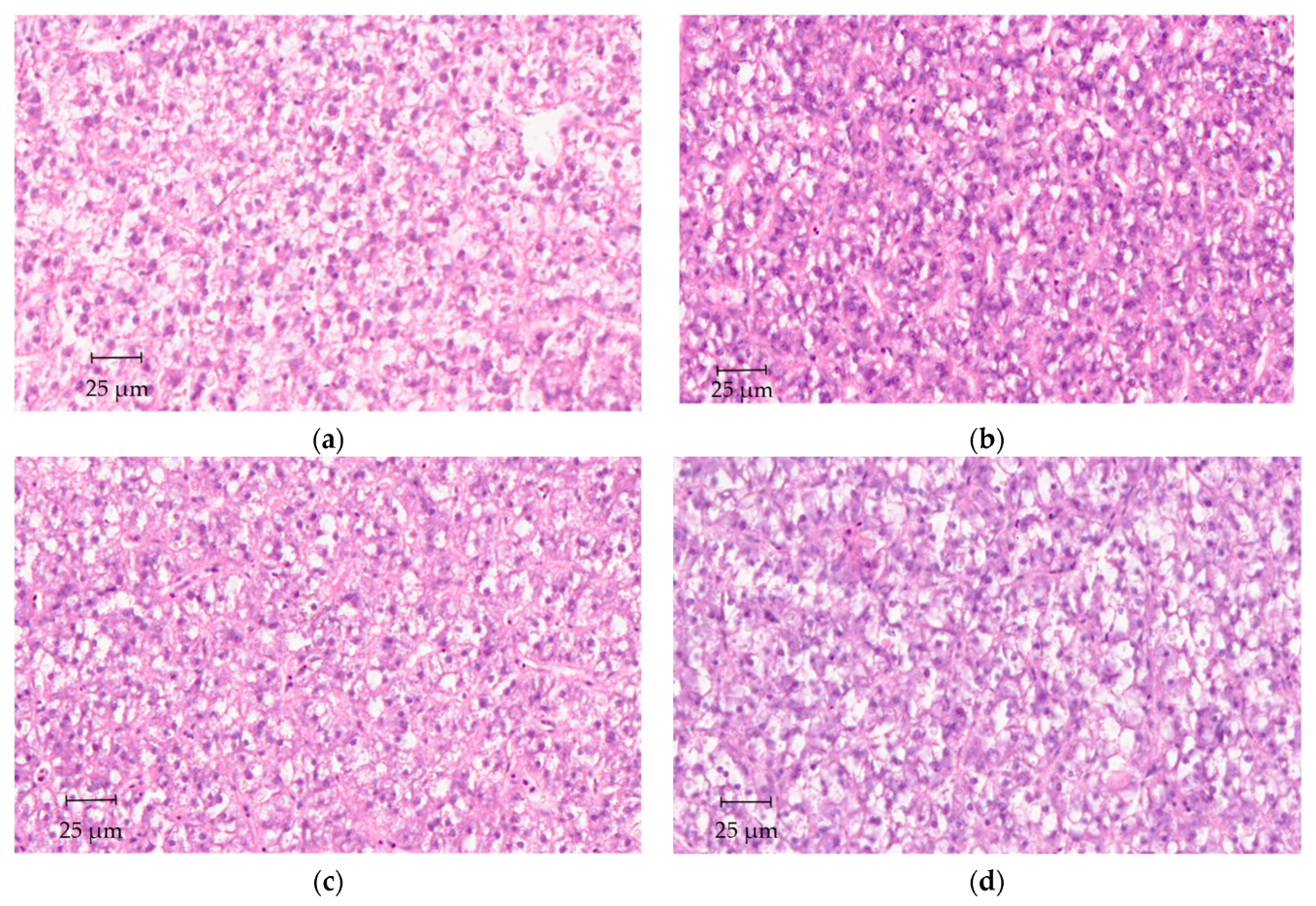
2.5. Histological Analyses
2.6. Gene Expression
2.6.1. RNA Extraction and cDNA Synthesis
2.6.2. Real-Time PCR (RT-PCR)
2.7. Statistical and Data Analysis
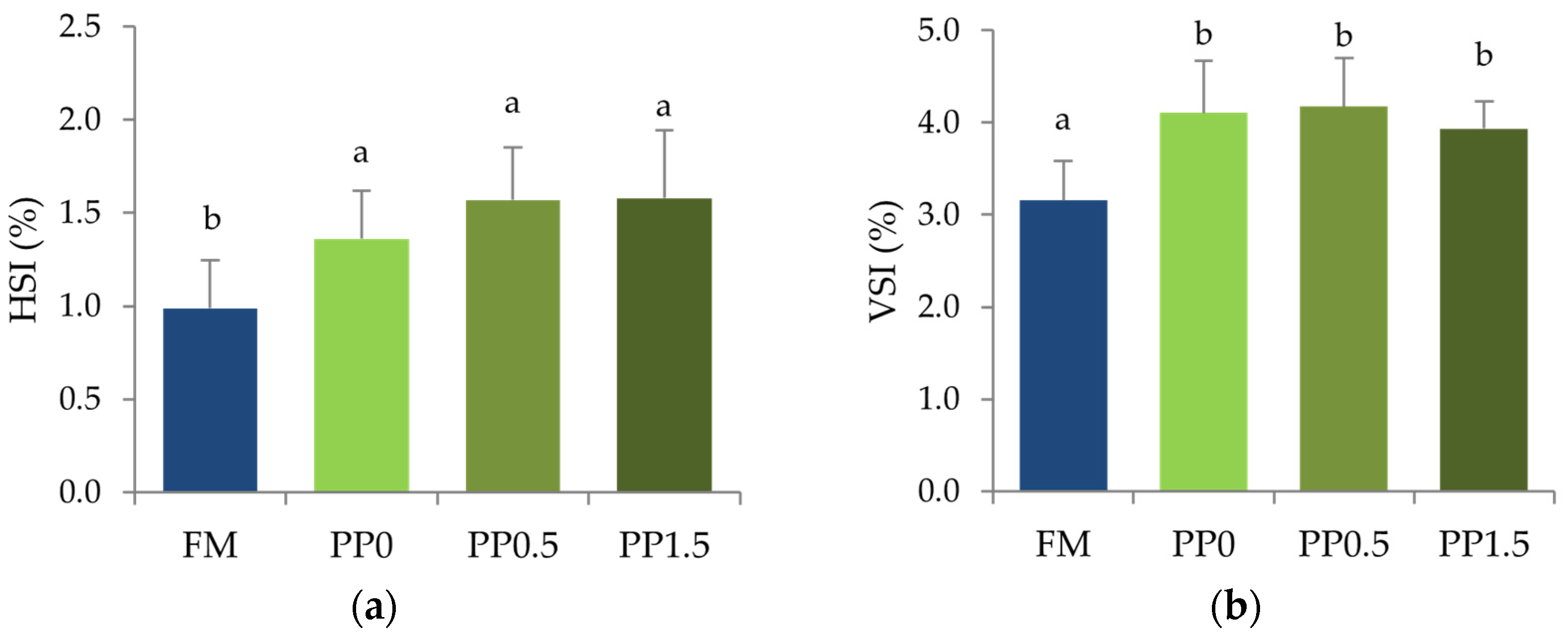
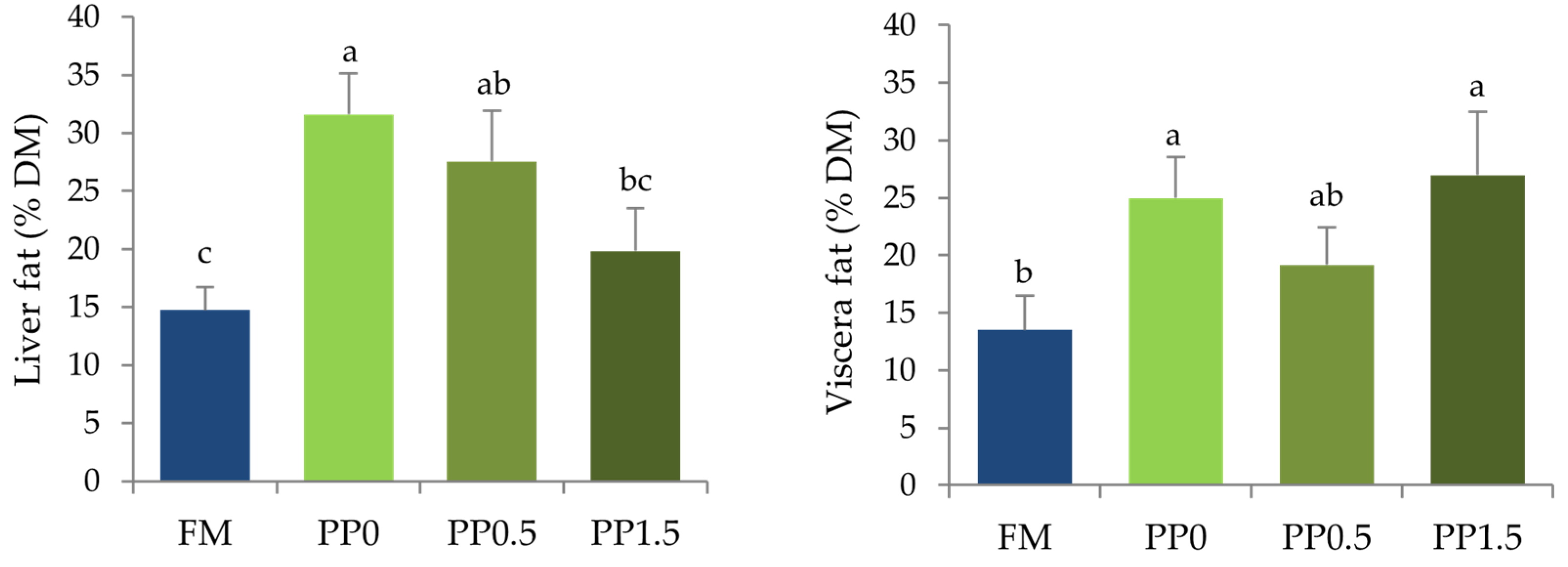
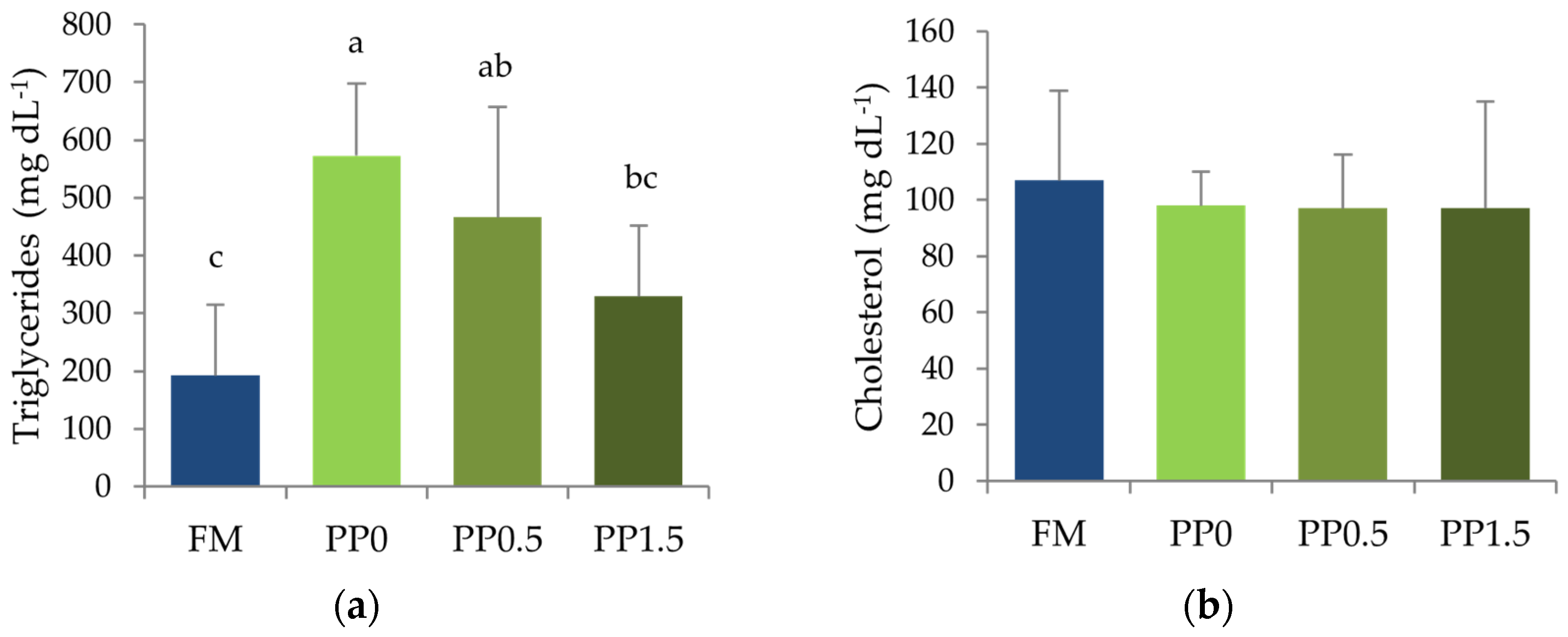
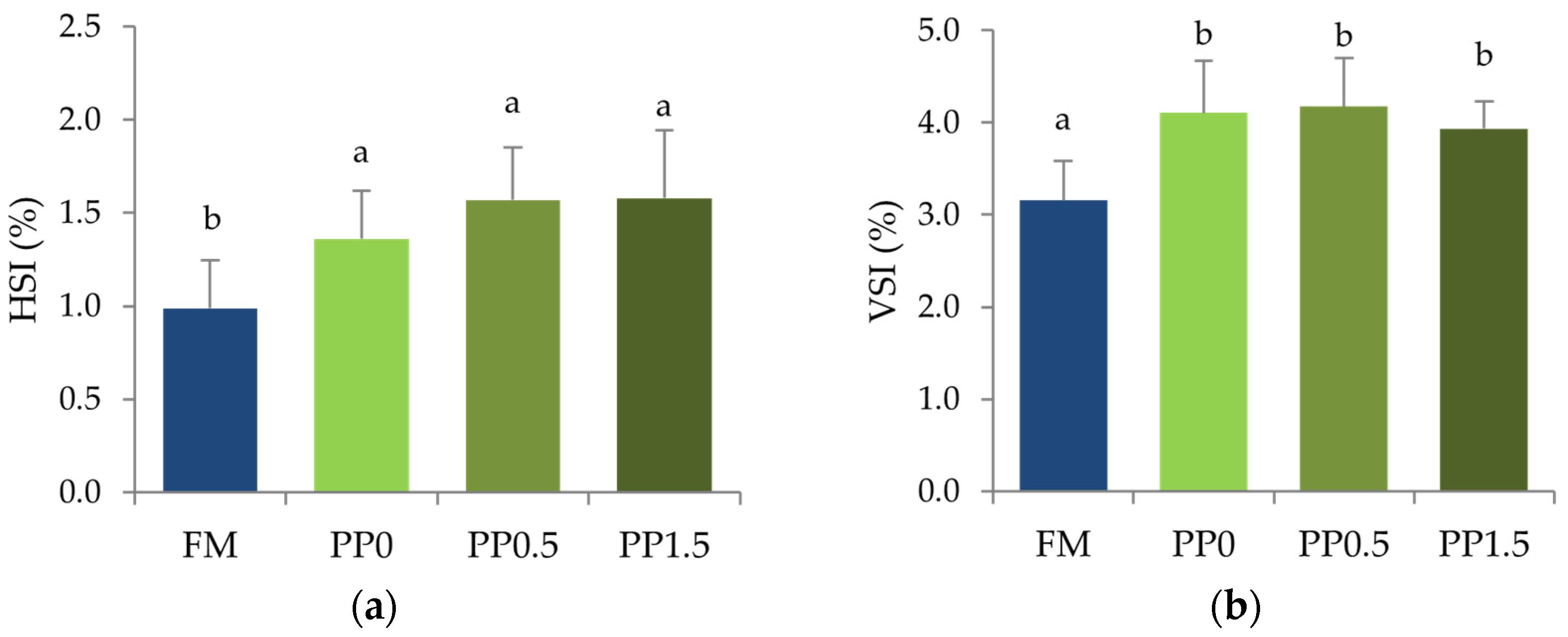
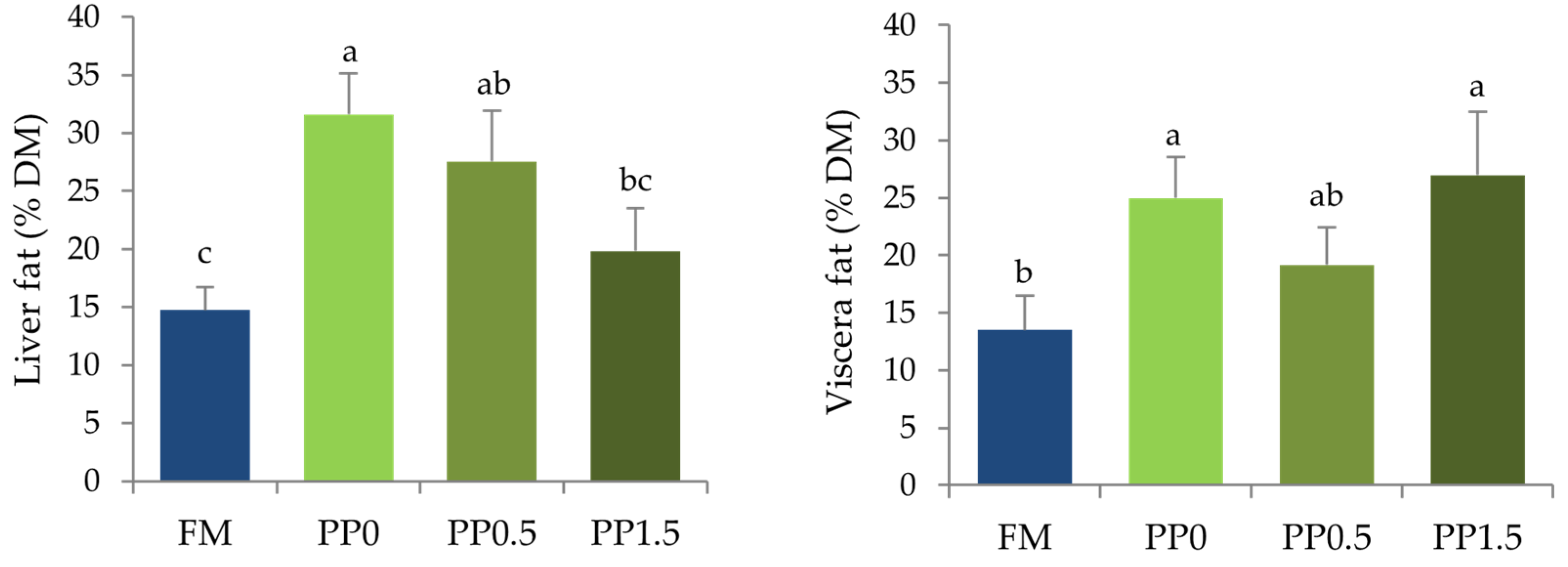
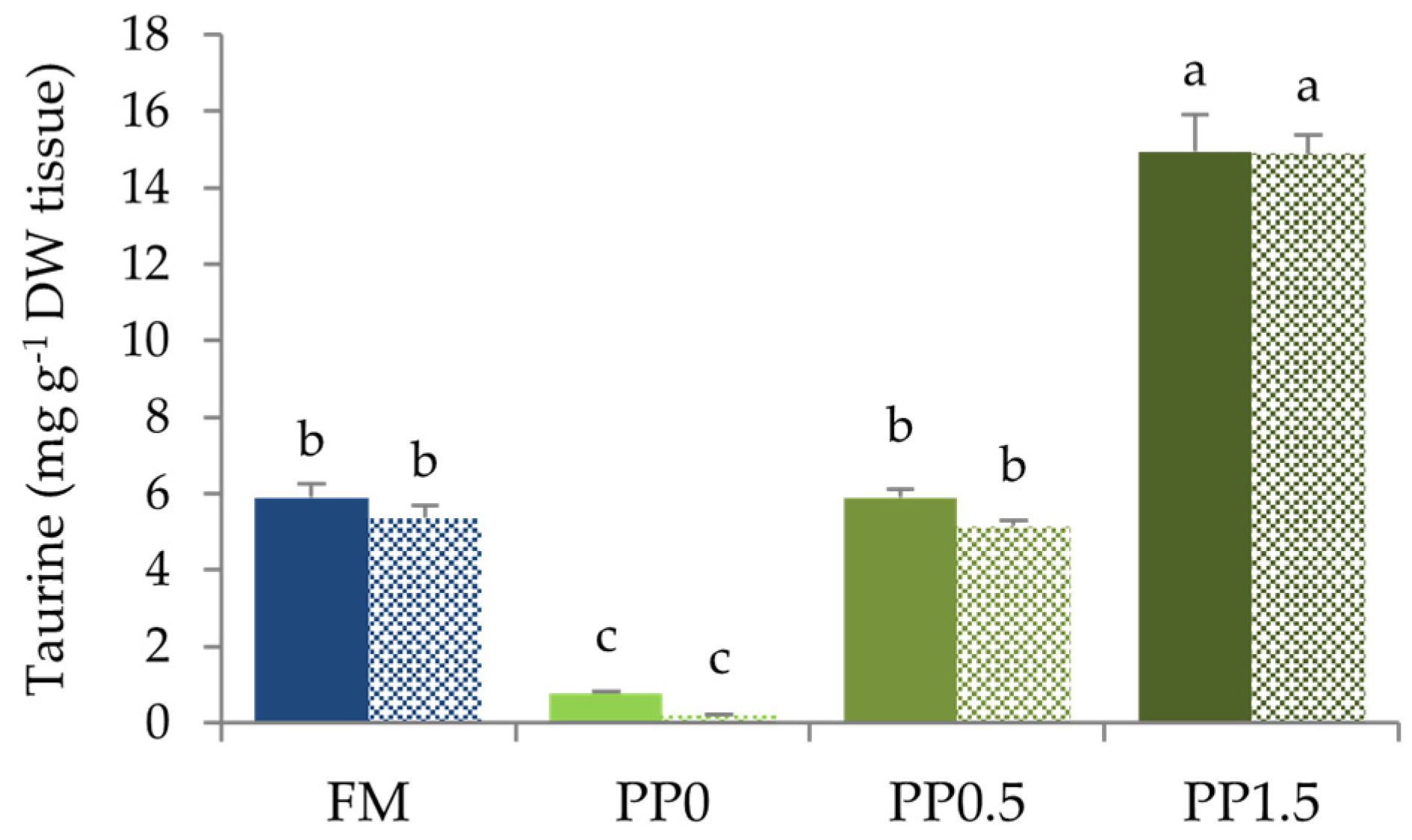
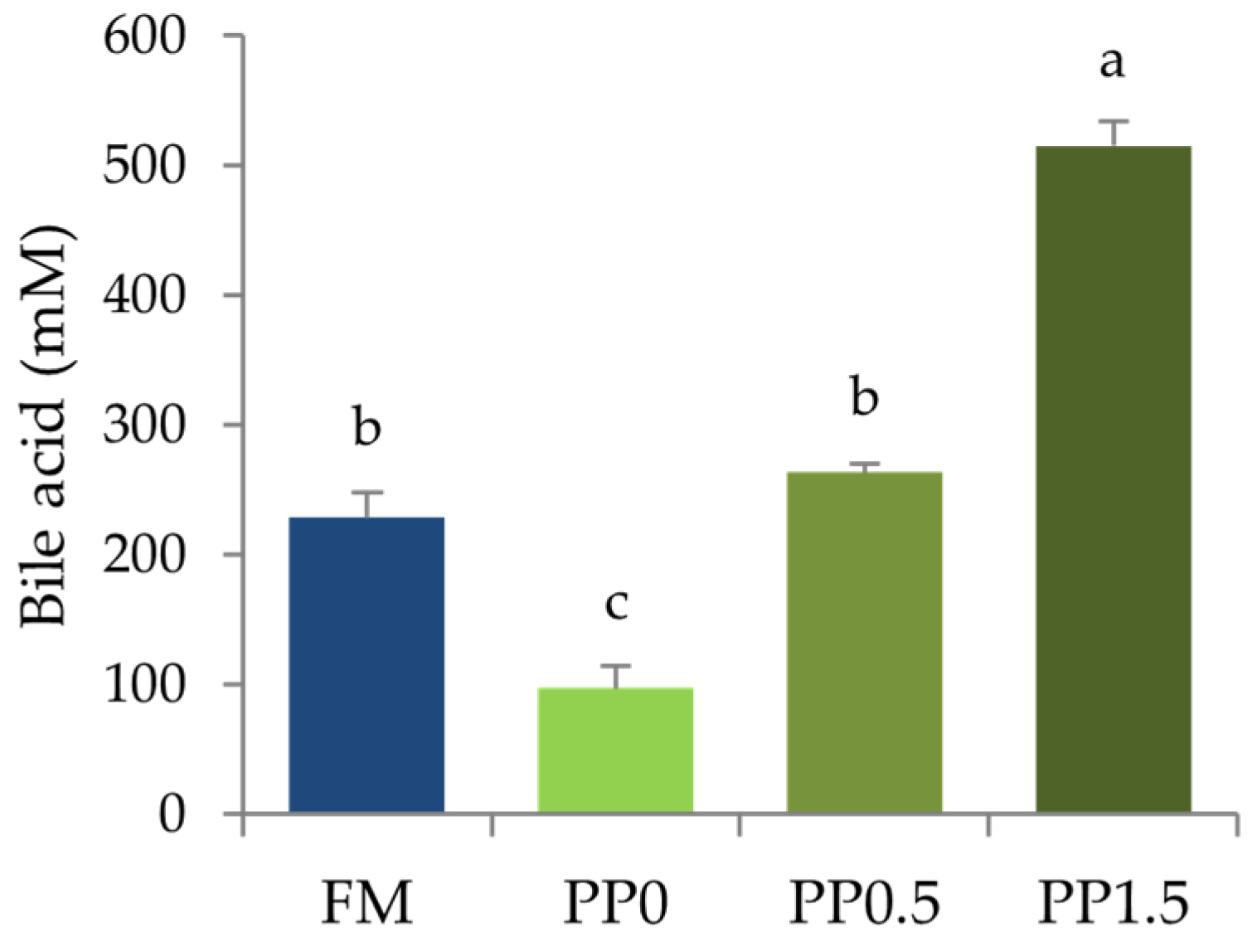
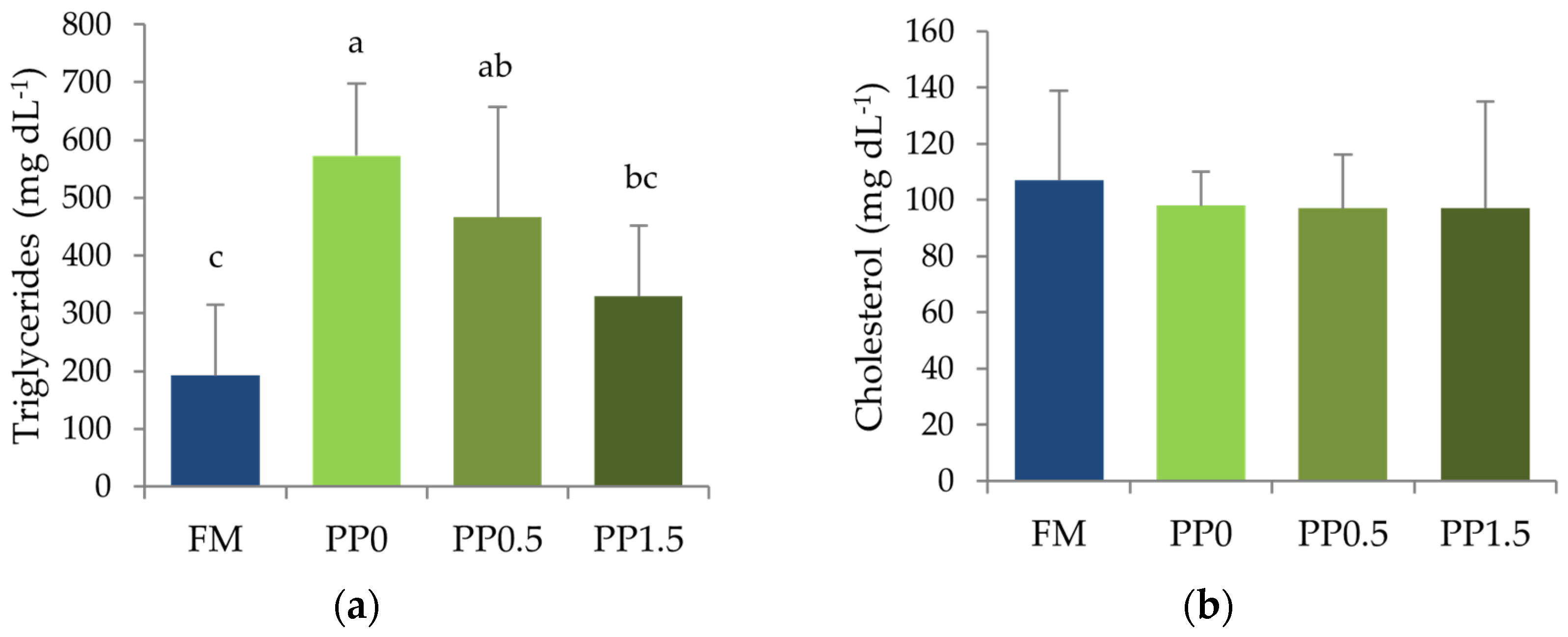
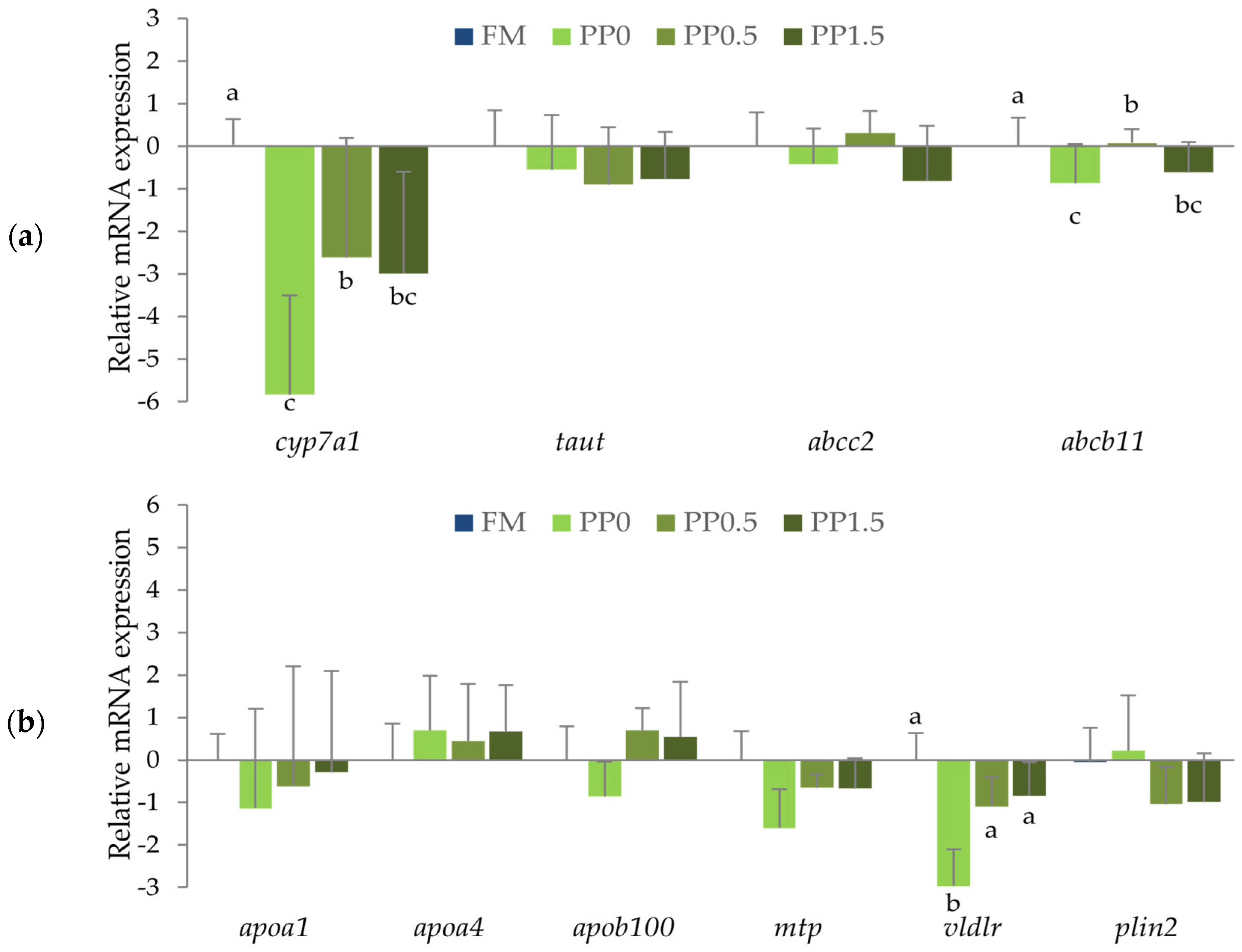
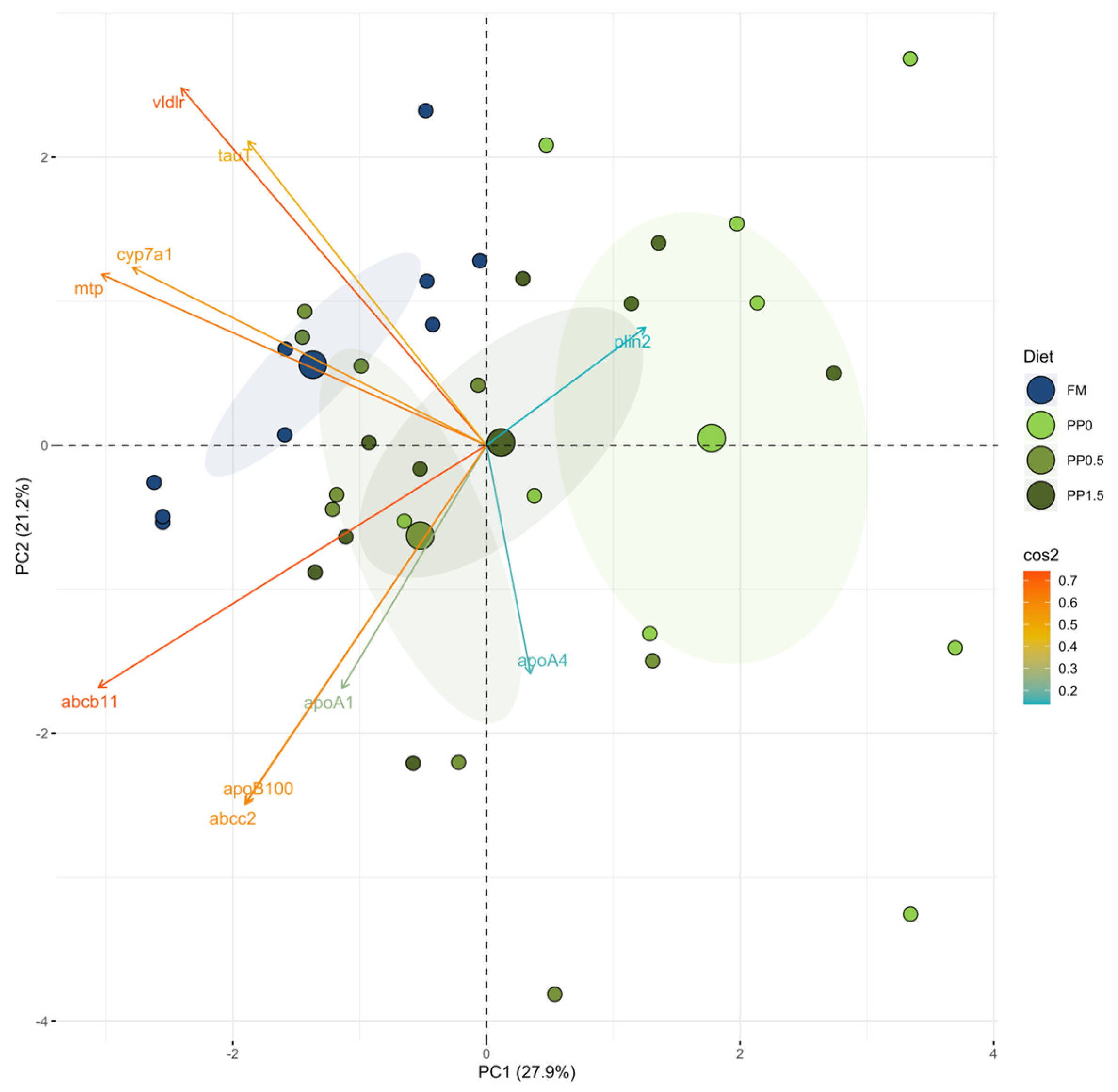
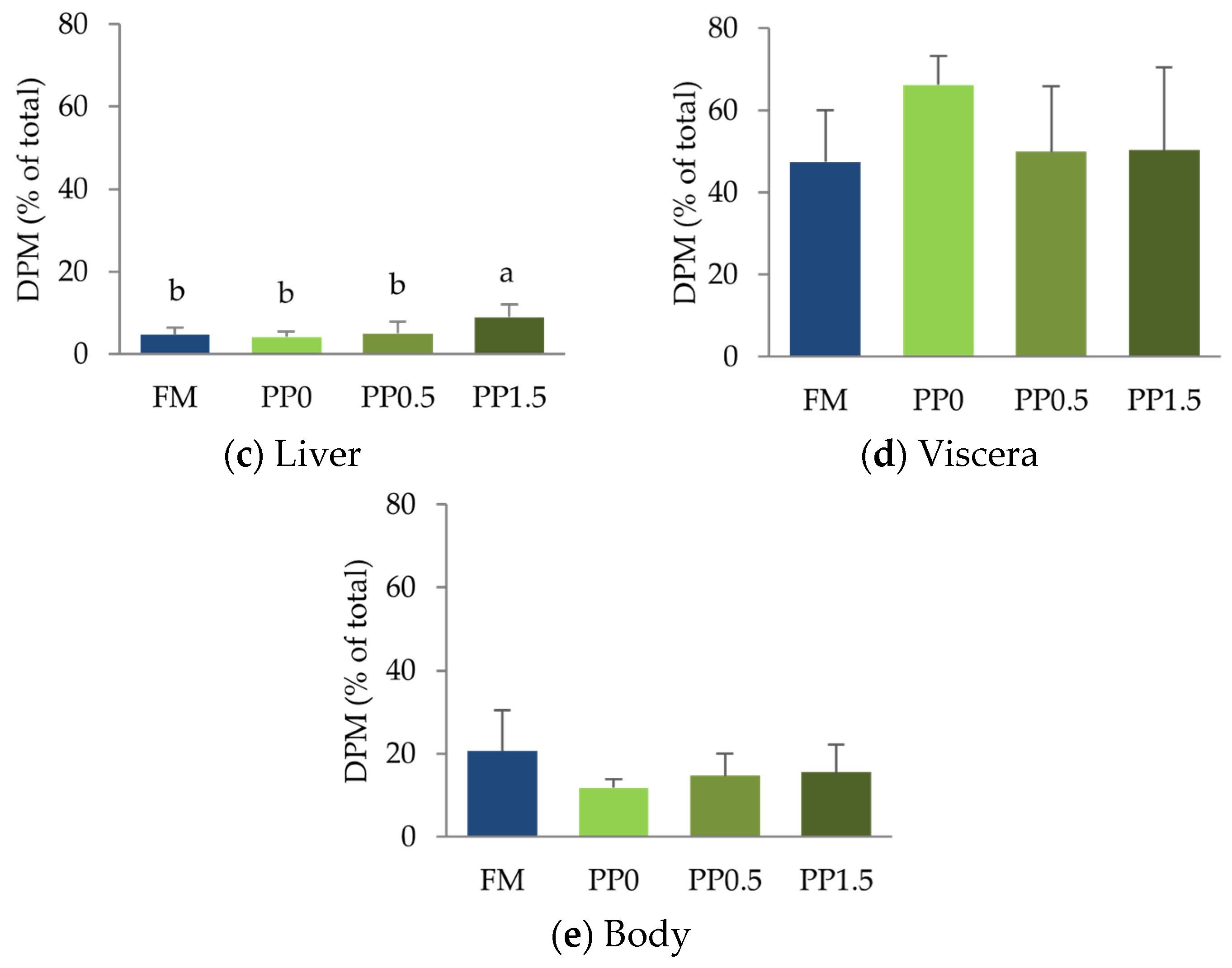
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Naylor, R.L.; Hardy, R.W.; Buschmann, A.H.; Bush, S.R.; Cao, L.; Klinger, D.H.; Little, D.C.; Lubchenco, J.; Shumway, S.E.; Troell, M. A 20-year retrospective review of global aquaculture. Nature 2021, 591, 551–563. [Google Scholar] [CrossRef] [PubMed]
- Salze, G.P.; Davis, D.A. Taurine: A critical nutrient for future fish feeds. Aquaculture 2015, 437, 215–229. [Google Scholar] [CrossRef]
- El-Sayed, A.-F.M. Is dietary taurine supplementation beneficial for farmed fish and shrimp? A comprehensive review. Rev. Aquac. 2014, 6, 241–255. [Google Scholar] [CrossRef]
- NRC. Nutrient Requirements of Fish and Shrimp; National Research Council: Washington, DC, USA, 2011. [Google Scholar] [CrossRef]
- Hundt, M.; Basit, H.; John, S. Physiology, Bile Secretion; SUNY Upstate Medical University; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Hofmann, A.F.; Hagey, L.R.; Krasowski, M.D. Bile salts of vertebrates: Structural variation and possible evolutionary significance. J. Lipid. Res. 2010, 51, 226–246. [Google Scholar] [CrossRef]
- Ben Saad, A.; Bruneau, A.; Mareux, E.; Lapalus, M.; Delaunay, J.-L.; Gonzales, E.; Jacquemin, E.; Aït-Slimane, T.; Falguières, T. Molecular regulation of canalicular ABC transporters. Int. J. Mol. Sci. 2021, 22, 2113. [Google Scholar] [CrossRef]
- Gaylord, T.G.; Teague, A.M.; Barrows, F.T. Taurine supplementation of all-plant protein diets for rainbow trout (Oncorhynchus mykiss). J. World Aquac. Soc. 2006, 37, 509–517. [Google Scholar] [CrossRef]
- Lim, S.-J.; Oh, D.-H.; Khosravi, S.; Cha, J.-H.; Park, S.-H.; Kim, K.-W.; Lee, K.-J. Taurine is an essential nutrient for juvenile parrot fish Oplegnathus fasciatus. Aquaculture 2013, 414–415, 274–279. [Google Scholar] [CrossRef]
- Chatzifotis, S.; Polemitou, I.; Divanach, P.; Antonopoulou, E. Effect of dietary taurine supplementation on growth performance and bile salt activated lipase activity of common dentex, Dentex dentex, fed a fish meal/soy protein concentrate-based diet. Aquaculture 2008, 275, 201–208. [Google Scholar] [CrossRef]
- Kim, S.-K.; Matsunari, H.; Takeuchi, T.; Yokoyama, M.; Furuita, H.; Murata, Y.; Goto, T. Comparison of taurine biosynthesis ability between juveniles of Japanese flounder and common carp. Amino Acids 2008, 35, 161–168. [Google Scholar] [CrossRef]
- Adeshina, I.; Abdel-Tawwab, M. Dietary taurine incorporation to high plant protein-based diets improved growth, biochemical, immunity, and antioxidants biomarkers of African catfish, Clarias gariepinus (B.). Fish Physiol. Biochem. 2020, 46, 1323–1335. [Google Scholar] [CrossRef]
- Zhang, J.; Hu, Y.; Ai, Q.; Mao, P.; Tian, Q.; Zhong, L.; Xiao, T.; Chu, W. Effect of dietary taurine supplementation on growth performance, digestive enzyme activities and antioxidant status of juvenile black carp (Mylopharyngodon piceus) fed with low fish meal diet. Aquac. Res. 2018, 49, 3187–3195. [Google Scholar] [CrossRef]
- Li, M.; Lai, H.; Li, Q.; Gong, S.; Wang, R. Effects of dietary taurine on growth, immunity and hyperammonemia in juvenile yellow catfish Pelteobagrus fulvidraco fed all-plant protein diets. Aquaculture 2016, 450, 349–355. [Google Scholar] [CrossRef]
- Yan, L.-C.; Feng, L.; Jiang, W.-D.; Wu, P.; Liu, Y.; Jiang, J.; Tang, L.; Tang, W.-N.; Zhang, Y.-A.; Yang, J.; et al. Dietary taurine supplementation to a plant protein source-based diet improved the growth and intestinal immune function of young grass carp (Ctenopharyngodon idella). Aquac. Nutr. 2019, 25, 873–896. [Google Scholar] [CrossRef]
- Michelato, M.; Furuya, W.M.; Gatlin, D.M. Metabolic responses of Nile tilapia Oreochromis niloticus to methionine and taurine supplementation. Aquaculture 2018, 485, 66–72. [Google Scholar] [CrossRef]
- Guimarães, I.G.; Skjaerven, K.; Moren, M.; Espe, M.; Hamre, K. Dietary taurine supplementation in plant protein based diets do not affect growth and reproductive performance of zebrafish. Aquac. Res. 2018, 49, 2013–2022. [Google Scholar] [CrossRef]
- Espe, M.; Ruohonen, K.; El-Mowafi, A. Effect of taurine supplementation on the metabolism and body lipid-to-protein ratio in juvenile Atlantic salmon (Salmo salar). Aquac. Res. 2012, 43, 349–360. [Google Scholar] [CrossRef]
- Yun, B.; Ai, Q.; Mai, K.; Xu, W.; Qi, G.; Luo, Y. Synergistic effects of dietary cholesterol and taurine on growth performance and cholesterol metabolism in juvenile turbot (Scophthalmus maximus L.) fed high plant protein diets. Aquaculture 2012, 324–325, 85–91. [Google Scholar] [CrossRef]
- Magalhães, R.; Martins, N.; Martins, S.; Lopes, T.; Diáz-Rosales, P.; Pousão-Ferreira, P.; Oliva-Teles, A.; Peres, H. Is dietary taurine required for white seabream (Diplodus sargus) juveniles? Aquaculture 2019, 502, 296–302. [Google Scholar] [CrossRef]
- López, L.M.; Flores-Ibarra, M.; Bañuelos-Vargas, I.; Galaviz, M.A.; True, C.D. Effect of fishmeal replacement by soy protein concentrate with taurine supplementation on growth performance, hematological and biochemical status, and liver histology of totoaba juveniles (Totoaba macdonaldi). Fish Physiol. Biochem. 2015, 41, 921–936. [Google Scholar] [CrossRef]
- Takagi, S.; Murata, H.; Goto, T.; Ichiki, T.; Munasinghe, D.M.S.; Endo, M.; Matsumoto, T.; Sakurai, A.; Hatate, H.; Yoshida, T.; et al. The green liver syndrome is caused by taurine deficiency in yellowtail, Seriola quinqueradiata fed diets without fishmeal. Aquac. Sci. 2005, 53, 279–290. [Google Scholar] [CrossRef]
- Takagi, S.; Murata, H.; Goto, T.; Hatate, H.; Endo, M.; Yamashita, H.; Miyatake, H.; Ukawa, M. Role of taurine deficiency in inducing green liver symptom and effect of dietary taurine supplementation in improving growth in juvenile red sea bream Pagrus major fed non-fishmeal diets based on soy protein concentrate. Fish Sci. 2011, 77, 235–244. [Google Scholar] [CrossRef]
- Koven, W.; Peduel, A.; Gada, M.; Nixon, O.; Ucko, M. Taurine improves the performance of white grouper juveniles (Epinephelus aeneus) fed a reduced fish meal diet. Aquaculture 2016, 460, 8–14. [Google Scholar] [CrossRef]
- Matsunari, H.; Furuita, H.; Yamamoto, T.; Kim, S.-K.; Sakakura, Y.; Takeuchi, T. Effect of dietary taurine and cystine on growth performance of juvenile red sea bream Pagrus major. Aquaculture 2008, 274, 142–147. [Google Scholar] [CrossRef]
- Tong, S.; Wang, L.; Kalhoro, H.; Volatiana, J.A.; Shao, Q. Effects of supplementing taurine in all-plant protein diets on growth performance, serum parameters, and cholesterol 7α-hydroxylase gene expression in black sea bream, Acanthopagrus schlegelii. J. World Aquac. Soc. 2020, 51, 990–1001. [Google Scholar] [CrossRef]
- Bañuelos-Vargas, I.; López, L.M.; Pérez-Jiménez, A.; Peres, H. Effect of fishmeal replacement by soy protein concentrate with taurine supplementation on hepatic intermediary metabolism and antioxidant status of totoaba juveniles (Totoaba macdonaldi). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2014, 170, 18–25. [Google Scholar] [CrossRef]
- Martins, N.; Diógenes, A.F.; Magalhães, R.; Matas, I.; Oliva-Teles, A.; Peres, H. Dietary taurine supplementation affects lipid metabolism and improves the oxidative status of European seabass (Dicentrarchus labrax) juveniles. Aquaculture 2021, 531, 735820. [Google Scholar] [CrossRef]
- Martins, N.; Magalhães, R.; Castro, C.; Couto, A.; Díaz-Rosales, P.; Oliva-Teles, A.; Peres, H. Taurine modulates hepatic oxidative status and gut inflammatory markers of European seabass (Dicentrarchus labrax) fed plant feedstuffs-based diets. Amino Acids 2019, 51, 1307–1321. [Google Scholar] [CrossRef]
- Hoseini, S.M.; Hosseini, S.A.; Eskandari, S.; Amirahmadi, M.; Soudagar, M. The effect of dietary taurine on growth performance and liver histopathology in Persian sturgeon, Acipenser persicus (Borodin, 1897) fed plant-based diet. Aquac. Res. 2017, 48, 4184–4196. [Google Scholar] [CrossRef]
- Hoseini, S.M.; Hosseini, S.A.; Eskandari, S.; Amirahmadi, M. Effect of dietary taurine and methionine supplementation on growth performance, body composition, taurine retention and lipid status of Persian sturgeon, Acipenser persicus (Borodin, 1897), fed with plant-based diet. Aquac. Nutr. 2018, 24, 324–331. [Google Scholar] [CrossRef]
- Takagi, S.; Murata, H.; Goto, T.; Hatate, H.; Endo, M.; Yamashita, H.; Miyatake, H.; Ukawa, M. Necessity of dietary taurine supplementation for preventing green liver symptom and improving growth performance in yearling red sea bream Pagrus major fed nonfishmeal diets based on soy protein concentrate. Fish. Sci. 2010, 76, 119–130. [Google Scholar] [CrossRef]
- Militante, J.D.; Lombardini, J.B. Dietary taurine supplementation: Hypolipidemic and antiatherogenic effects. Nutr. Res. 2004, 24, 787–801. [Google Scholar] [CrossRef]
- Xu, H.; Zhang, Q.; Kim, S.K.; Liao, Z.; Wei, Y.; Sun, B.; Jia, L.; Chi, S.; Liang, M. Dietary taurine stimulates the hepatic biosynthesis of both bile acids and cholesterol in the marine teleost, tiger puffer (Takifugu rubripes). Br. J. Nutr. 2020, 123, 1345–1356. [Google Scholar] [CrossRef]
- Kim, S.-K.; Kim, K.-G.; Kim, K.-D.; Kim, K.-W.; Son, M.-H.; Rust, M.; Johnson, R. Effect of dietary taurine levels on the conjugated bile acid composition and growth of juvenile Korean rockfish Sebastes schlegeli (Hilgendorf). Aquac. Res. 2015, 46, 2768–2775. [Google Scholar] [CrossRef]
- de Moura, L.B.; Diógenes, A.F.; Campelo, D.A.V.; de Almeida, F.L.A.; Pousão-Ferreira, P.M.; Furuya, W.M.; Peres, H.; Oliva-Teles, A. Nutrient digestibility, digestive enzymes activity, bile drainage alterations and plasma metabolites of meagre (Argyrosomus regius) feed high plant protein diets supplemented with taurine and methionine. Aquaculture 2019, 511, 734231. [Google Scholar] [CrossRef]
- Watson, A.M.; Barrows, F.T.; Place, A.R. Effects of graded taurine levels on juvenile cobia. N. Am. J. Aquac. 2014, 76, 190–200. [Google Scholar] [CrossRef]
- Pinto, W.; Engrola, S.; Conceição, L.E.C. Towards an early weaning in Senegalese sole: A historical review. Aquaculture 2018, 496, 1–9. [Google Scholar] [CrossRef]
- Silva, J.M.G.; Espe, M.; Conceição, L.E.C.; Dias, J.; Valente, L.M.P. Senegalese sole juveniles (Solea senegalensis Kaup, 1858) grow equally well on diets devoid of fish meal provided the dietary amino acids are balanced. Aquaculture 2009, 296, 309–317. [Google Scholar] [CrossRef]
- Silva, J.M.G.; Espe, M.; Conceição, L.E.C.; Dias, J.; Costas, B.; Valente, L.M.P. Feed intake and growth performance of Senegalese sole (Solea senegalensis Kaup, 1858) fed diets with partial replacement of fish meal with plant proteins. Aquac. Res. 2010, 41, e20–e30. [Google Scholar] [CrossRef]
- Cabral, E.M.; Bacelar, M.; Batista, S.; Castro-Cunha, M.; Ozório, R.O.A.; Valente, L.M.P. Replacement of fishmeal by increasing levels of plant protein blends in diets for Senegalese sole (Solea senegalensis) juveniles. Aquaculture 2011, 322–323, 74–81. [Google Scholar] [CrossRef]
- Cabral, E.M.; Fernandes, T.J.R.; Campos, S.D.; Castro-Cunha, M.; Oliveira, M.B.P.P.; Cunha, L.M.; Valente, L.M.P. Replacement of fish meal by plant protein sources up to 75% induces good growth performance without affecting flesh quality in ongrowing Senegalese sole. Aquaculture 2013, 380–383, 130–138. [Google Scholar] [CrossRef]
- Valente, L.M.P.; Linares, F.; Villanueva, J.L.R.; Silva, J.M.G.; Espe, M.; Escórcio, C.; Pires, M.A.; Saavedra, M.J.; Borges, P.; Medale, F.; et al. Dietary protein source or energy levels have no major impact on growth performance, nutrient utilisation or flesh fatty acids composition of market-sized Senegalese sole. Aquaculture 2011, 318, 128–137. [Google Scholar] [CrossRef]
- Borges, P.; Oliveira, B.; Casal, S.; Dias, J.; Conceição, L.; Valente, L.M.P. Dietary lipid level affects growth performance and nutrient utilisation of Senegalese sole (Solea senegalensis) juveniles. Br. J. Nutr. 2009, 102, 1007–1014. [Google Scholar] [CrossRef] [PubMed]
- Aragão, C.; Colen, R.; Ferreira, S.; Pinto, W.; Conceição, L.E.C.; Dias, J. Microencapsulation of taurine in Senegalese sole diets improves its metabolic availability. Aquaculture 2014, 431, 53–58. [Google Scholar] [CrossRef]
- Richard, N.; Colen, R.; Aragão, C. Supplementing taurine to plant-based diets improves lipid digestive capacity and amino acid retention of Senegalese sole (Solea senegalensis) juveniles. Aquaculture 2017, 468, 94–101. [Google Scholar] [CrossRef]
- Colen, R.; Ramalho, A.; Rocha, F.; Dinis, M.T. Solea solea. In FAO Cultured Aquatic Species Information Programme; Fisheries and Aquaculture Division: Rome, Italy, 2015. [Google Scholar]
- Rønnestad, I.; Rojas-García, C.R.; Tonheim, S.K.; Conceição, L.E.C. In vivo studies of digestion and nutrient assimilation in marine fish larvae. Aquaculture 2001, 201, 161–175. [Google Scholar] [CrossRef]
- Costas, B. Stress Mitigation in Sole (Solea senegalensis) through Improved Nitrogen Nutrition: Amino Acid Utilization, Disease Resistance and Immune Status. Ph.D. Thesis, University of Porto, Porto, Portugal, 2011. [Google Scholar]
- Teodósio, R.; Engrola, S.; Cabano, M.; Colen, R.; Masagounder, K.; Aragão, C. Metabolic and nutritional responses of Nile tilapia juveniles to dietary methionine sources. Br. J. Nutr. 2022, 127, 202–213. [Google Scholar] [CrossRef]
- Teodósio, R.; Aragão, C.; Conceição, L.E.C.; Dias, J.; Engrola, S. Metabolic fate is defined by amino acid nature in gilthead seabream fed different diet formulations. Animals 2022, 12, 1713. [Google Scholar] [CrossRef]
- Aragão, C.; Cabano, M.; Colen, R.; Teodósio, R.; Gisbert, E.; Dias, J.; Engrola, S. Modulation of dietary protein to lipid ratios for gilthead seabream on-growing during summer temperature conditions. Aquac. Rep. 2022, 25, 101262. [Google Scholar] [CrossRef]
- Ribeiro, L.; Moura, J.; Santos, M.; Colen, R.; Rodrigues, V.; Bandarra, N.; Soares, F.; Ramalho, P.; Barata, M.; Moura, P.; et al. Effect of vegetable based diets on growth, intestinal morphology, activity of intestinal enzymes and haematological stress indicators in meagre (Argyrosomus regius). Aquaculture 2015, 447, 116–128. [Google Scholar] [CrossRef]
- Infante, C.; Matsuoka, M.; Asensio, E.; Canavate, J.; Reith, M.; Manchado, M. Selection of housekeeping genes for gene expression studies in larvae from flatfish using real-time PCR. BMC Mol. Biol. 2008, 9, 28. [Google Scholar] [CrossRef]
- Pinto, W.; Rønnestad, I.; Jordal, A.E.O.; Gomes, A.S.; Dinis, M.T.; Aragão, C. Cloning, tissue and ontogenetic expression of the taurine transporter in the flatfish Senegalese sole (Solea senegalensis). Amino Acids 2012, 42, 1317–1327. [Google Scholar] [CrossRef] [PubMed]
- Borges, P.; Medale, F.; Veron, V.; dos Pires, M.A.; Dias, J.; Valente, L.M.P. Lipid digestion, absorption and uptake in Solea senegalensis. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2013, 166, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Xie, F.; Xiao, P.; Chen, D.; Xu, L.; Zhang, B. MiRDeepFinder: A miRNA analysis tool for deep sequencing of plant small RNAs. Plant Mol. Biol. 2012, 80, 75–84. [Google Scholar] [CrossRef]
- Ennos, R. Statistical and Data Handling Skills in Biology, 2nd ed.; Pearson Prentice Hall: Harlow, UK, 2007. [Google Scholar]
- de Moura, L.B.; Diógenes, A.F.; Campelo, D.A.V.V.; de Almeida, F.L.; Pousão-Ferreira, P.M.; Furuya, W.M.; Oliva-Teles, A.; Peres, H. Taurine and methionine supplementation as a nutritional strategy for growth promotion of meagre (Argyrosomus regius) fed high plant protein diets. Aquaculture 2018, 497, 389–395. [Google Scholar] [CrossRef]
- Pinto, W.; Figueira, L.; Santos, A.; Barr, Y.; Helland, S.; Dinis, M.T.; Aragão, C. Is dietary taurine supplementation beneficial for gilthead seabream (Sparus aurata) larvae? Aquaculture 2013, 384–387, 1–5. [Google Scholar] [CrossRef]
- ARRAINA. Feed Ingredients in Aquaculture, Technical Booklet. 2015. Available online: https://www.sparos.pt/wp-content/uploads/2019/04/BOOKLET-ARRAIANA.pdf (accessed on 8 January 2023).
- Schöttler, U.; Daniels, D.; Zapf, K. Influence of anoxia on adaptation of euryhaline polychaetes to hyposmotic conditions. Mar. Biol. 1990, 104, 443–451. [Google Scholar] [CrossRef]
- Qi, G.; Ai, Q.; Mai, K.; Xu, W.; Liufu, Z.; Yun, B.; Zhou, H. Effects of dietary taurine supplementation to a casein-based diet on growth performance and taurine distribution in two sizes of juvenile turbot (Scophthalmus maximus L.). Aquaculture 2012, 358–359, 122–128. [Google Scholar] [CrossRef]
- Kim, S.-K.; Takeuchi, T.; Yokoyama, M.; Murata, Y. Effect of dietary supplementation with taurine, beta-alanine and GABA on the growth of juvenile and fingerling Japanese flounder Paralichthys olivaceus. Fish Sci. 2003, 69, 242–248. [Google Scholar] [CrossRef]
- Coutinho, F.; Simões, R.; Monge-Ortiz, R.; Furuya, W.M.; Pousão-Ferreira, P.; Kaushik, S.; Oliva-Teles, A.; Peres, H. Effects of dietary methionine and taurine supplementation to low-fish meal diets on growth performance and oxidative status of European sea bass (Dicentrarchus labrax) juveniles. Aquaculture 2017, 479, 447–454. [Google Scholar] [CrossRef]
- Khaoian, P.; Nguyen, H.P.; Ogita, Y.; Fukada, H.; Masumoto, T. Taurine supplementation and palm oil substitution in low-fish meal diets for young yellowtail Seriola quinqueradiata. Aquaculture 2014, 420–421, 219–224. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diets | ||||
|---|---|---|---|---|
| Ingredients | FM | PP0 | PP0.5 | PP1.5 |
| Fishmeal Super Prime 1 | 37.00 | 3.00 | 3.00 | 3.00 |
| Fishmeal 60 2 | 15.00 | 0.00 | 0.00 | 0.00 |
| Fish soluble protein concentrate 3 | 7.50 | 3.00 | 3.00 | 3.00 |
| Squid meal 4 | 7.50 | 3.00 | 3.00 | 3.00 |
| Fish gelatine 5 | 2.00 | 2.00 | 2.00 | 2.00 |
| Pea protein concentrate 6 | 0.00 | 20.00 | 20.00 | 20.00 |
| Potato protein concentrate 7 | 0.00 | 12.00 | 12.00 | 12.00 |
| Wheat gluten 8 | 0.00 | 10.00 | 9.45 | 7.70 |
| Corn gluten meal 9 | 0.00 | 8.00 | 8.00 | 8.00 |
| Soybean meal 10 | 5.00 | 5.00 | 5.00 | 5.00 |
| Soy protein concentrate 11 | 5.00 | 5.00 | 5.00 | 5.00 |
| Wheat meal 12 | 6.40 | 5.10 | 5.10 | 5.10 |
| Pea starch 13 | 7.00 | 7.00 | 7.00 | 7.00 |
| Fish oil 14 | 2.50 | 6.80 | 6.80 | 6.80 |
| Vitamin and mineral premix 15 | 0.20 | 0.20 | 0.20 | 0.20 |
| Lutavit C35 16 | 0.10 | 0.10 | 0.10 | 0.10 |
| Lutavit E50 17 | 0.05 | 0.05 | 0.05 | 0.05 |
| Monocalcium phosphate 18 | 0.00 | 4.00 | 4.00 | 4.00 |
| Glycerol 19 | 2.50 | 2.50 | 2.50 | 2.50 |
| Binder 20 | 2.00 | 2.00 | 2.00 | 2.00 |
| Antioxidant 21 | 0.25 | 0.25 | 0.25 | 0.25 |
| Lysine 22,* | 0.00 | 0.50 | 0.50 | 0.50 |
| Methionine 23,* | 0.00 | 0.50 | 0.50 | 0.50 |
| Taurine 24,* | 0.00 | 0.00 | 0.55 | 2.30 |
| Proximate composition (% as fed) | ||||
| Dry matter | 92.7 | 92.4 | 92.6 | 92.5 |
| Ash | 9.2 | 9.2 | 9.3 | 9.1 |
| Crude protein | 55.2 | 54.9 | 55.1 | 55.0 |
| Crude fat | 8.6 | 8.6 | 8.6 | 8.6 |
| Taurine | 0.40 | 0.08 | 0.43 | 1.40 |
| Amino Acids | Diets | |||
|---|---|---|---|---|
| (% as Fed) | FM | PP0 | PP0.5 | PP1.5 |
| Arginine | 3.9 | 3.6 | 3.7 | 3.7 |
| Histidine | 1.4 | 1.1 | 1.2 | 1.2 |
| Lysine | 4.2 | 4.1 | 4.1 | 4.1 |
| Threonine | 2.1 | 1.8 | 1.9 | 1.9 |
| Isoleucine | 2.3 | 2.2 | 2.3 | 2.3 |
| Leucine | 3.5 | 4.2 | 4.2 | 4.2 |
| Valine | 2.6 | 2.6 | 2.6 | 2.5 |
| Methionine | 1.7 | 1.3 | 1.2 | 1.2 |
| Phenylalanine | 2.2 | 2.6 | 2.5 | 2.6 |
| Cystine | 0.2 | 0.3 | 0.3 | 0.3 |
| Tyrosine | 1.8 | 1.9 | 1.9 | 2.0 |
| Aspartic acid | 4.3 | 4.0 | 4.1 | 4.1 |
| Glutamic acid | 6.8 | 10.5 | 10.4 | 10.6 |
| Alanine | 2.7 | 2.2 | 2.2 | 2.2 |
| Glycine | 3.3 | 2.1 | 2.1 | 2.1 |
| Proline | 2.3 | 3.5 | 3.5 | 3.5 |
| Serine | 2.2 | 2.5 | 2.5 | 2.5 |
| Gene | Genbank/Unigene * Accession Number | Forward Primer (5′-3′) | Reverse Primer (5′-3′) | AT 1 (°C) |
|---|---|---|---|---|
| gapdh | AB291587 | AGCCACCGTGTCGCCGACCT | AAAAGAGGAGATGGTGGGGGGTGGT | 64 |
| cyp7a1 | 416791 * | GCCTACAGTGCCAGAGAGAAC | GCGAAGCCCAAAGCAGTG | 64 |
| taut | HQ148721.1 | CCGAAAGCTGTGTCCATGATG | CAATAGAGGTGATCTGTCCTTCCA | 63 |
| abbc2 | XM_044046365.1 | GCTTACATCCACGACTGCTTCCAA | ACATCCTGACTGACGCCTTCCTT | 60 |
| abcb11 | 29210 * | AAGCAGAACAACCAGCCATCAGG | CCACCACCATCATCAGCACATCTT | 60 |
| apoa1 | FF283994 | TTGAGGCTAATCGTGCCAAA | CCTGCGTGCTTGTCCTTGTA | 62 |
| apoa4 | KP842775.1 | AGGAACTCCAGCAGAACCTG | CCTGCGTGCTTGTCCTTGTA | 60 |
| apob100 | 14427 * | CCGCTGAGATGGAGAGATA | CTGGGTCATCTTGGAGAAGG | 64 |
| mtp | KC888960 | TGGCACGTTACTGTGGACAT | CCAGGGCAGAGATGATTC | 63 |
| vldlr | AJ_879619.2 | CTGTGTTTGAGGACCGAGTGTT | GACCTGCGTCTTCTTGCTCT | 64 |
| plin2 | 185823 * | CTGTCTGGTCCTTGTCTC | GCCTTGCTGAAGTTAGTG | 56 |
| Dietary Treatments | FM | PP0 | PP0.5 | PP1.5 |
|---|---|---|---|---|
| IBW (g) 1 | 13.7 ± 5.0 | 13.7 ± 5.7 | 13.6 ± 4.6 | 13.6 ± 4.9 |
| FBW (g) 2 | 27.6 ± 6.8 | 24.2 ± 7.0 | 24.2 ± 5.3 | 27.1 ± 7.3 |
| WG (%) 3 | 102.6 ± 10.9 a | 77.2 ± 4.8 b | 77.5 ± 4.5 b | 98.9 ± 5.2 a |
| VFI (% day−1) 4 | 1.2 ± 0.01 | 1.2 ± 0.04 | 1.2 ± 0.03 | 1.2 ± 0.01 |
| FCR 5 | 1.7 ± 0.2 | 2.3 ± 0.4 | 1.7 ± 0.3 | 2.0 ± 0.1 |
| Survival (%) | 83.8 ± 7.2 | 85.7 ± 7.6 | 83.8 ± 3.3 | 75.2 ± 5.9 |
| Dietary Treatments | FM | PP0 | PP0.5 | PP1.5 |
|---|---|---|---|---|
| Dry matter (%) | 74.8 ± 1.1 | 75.0 ± 0.4 | 75.5 ± 0.7 | 76.0 ± 1.3 |
| Ash (% DM) | 7.0 ± 0.2 b | 9.3 ± 1.2 a | 7.9 ± 0.4 ab | 8.2 ± 0.6 ab |
| Protein (% DM) | 67.2 ± 1.4 | 68.2 ± 0.7 | 69.3 ± 1.0 | 69.4 ± 0.7 |
| Fat (% DM) | 19.9 ± 1.9 a | 16.3 ± 1.6 b | 15.3 ± 0.7 b | 13.7 ± 1.0 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aragão, C.; Teodósio, R.; Colen, R.; Richard, N.; Rønnestad, I.; Dias, J.; Conceição, L.E.C.; Ribeiro, L. Taurine Supplementation to Plant-Based Diets Improves Lipid Metabolism in Senegalese Sole. Animals 2023, 13, 1501. https://doi.org/10.3390/ani13091501
Aragão C, Teodósio R, Colen R, Richard N, Rønnestad I, Dias J, Conceição LEC, Ribeiro L. Taurine Supplementation to Plant-Based Diets Improves Lipid Metabolism in Senegalese Sole. Animals. 2023; 13(9):1501. https://doi.org/10.3390/ani13091501
Chicago/Turabian StyleAragão, Cláudia, Rita Teodósio, Rita Colen, Nadège Richard, Ivar Rønnestad, Jorge Dias, Luís E. C. Conceição, and Laura Ribeiro. 2023. "Taurine Supplementation to Plant-Based Diets Improves Lipid Metabolism in Senegalese Sole" Animals 13, no. 9: 1501. https://doi.org/10.3390/ani13091501