
Molecular Detection and Genetic Characterization of Vertically Transmitted Viruses in Ducks


1
College of Veterinary Medicine, Southwest University, Chongqing 400715, China
2
Harbin Veterinary Research Institute, Chinese Academy of Agricultural Sciences, Harbin 150068, China
3
Chongqing Academy of Animal Science, Chongqing 408599, China
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Animals 2024, 14(1), 6; https://doi.org/10.3390/ani14010006
Submission received: 13 October 2023
/
Revised: 4 December 2023
/
Accepted: 11 December 2023
/
Published: 19 December 2023
(This article belongs to the Section Poultry)
Abstract
:Simple Summary
Vertically transmitted duck viruses are viruses that are transmitted from a female duck to its offspring when it is an egg, which seriously threatens production in the duck breeding industry. In this study, we evaluated the distribution and genetic variation in four vertically transmitted duck pathogens, including DHBV, DuCV, DHAV-3, and ARV. This study found that DHBV was the most prevalent virus, followed by DuCV, and then ARV and DHAV-3. The genetic analysis results showed that all the identified duck viruses here had complex qualities. These findings will improve our knowledge of the evolution of DuCV, DHAV-3, and ARV and help choose suitable strains for vaccination.
Abstract
To investigate the distribution and genetic variation in four vertically transmitted duck pathogens, including duck hepatitis B virus (DHBV), duck circovirus (DuCV), duck hepatitis A virus 3 (DHAV-3), and avian reoviruses (ARV), we conducted an epidemiology study using PCR and RT-PCR assays on a duck population. We found that DHBV was the most prevalent virus (69.74%), followed by DuCV (39.48%), and then ARV (19.92%) and DHAV-3 (8.49%). Among the 271 duck samples, two, three or four viruses were detected in the same samples, indicating that the coinfection of vertical transmission agents is common in ducks. The genetic analysis results showed that all four identified DuCV strains belonged to genotype 1, the DHAV-3 strain was closely clustered with previously identified strains from China, and the ARV stain was clustered under genotype 1. These indicate that different viral strains are circulating among the ducks. Our findings will improve the knowledge of the evolution of DuCV, DHAV-3, and ARV, and help choose suitable strains for vaccination.
1. Introduction
Duck viral diseases pose a serious threat to the health and production performance of duck flocks and are prevalent worldwide. The highly pathogenic avian influenza, Newcastle disease virus, duck viral enteritis, and duck viral hepatitis are among the main culprits causing significant economic losses to duck flocks. Numerous studies have been conducted to address these diseases [1]. However, there are also several duck vertical transmission viral diseases that are often overlooked, including duck hepatitis B virus (DHBV), duck circovirus (DuCV), duck hepatitis A virus (DHAV), and avian reoviruses (ARV). These diseases can pose health risks to the duck industry, leading to problems such as duckling deformities, increased mortality rates, and poor growth and development [2,3,4,5,6].
DHBV, a member of the Avihepadnavirus genus in the Hepadnaviridae family, is a small DNA virus with a diameter of 40–45 nm. The DHBV model is widely utilized as an animal infection model for studying human HBV infections [7]. The previous reports have shown that ducks that are persistently infected with DHBV have reduced egg hatching rates during breeding and their ducklings have limited growth [8,9]. Although DHBV infections usually do not show serious symptoms or liver damage in ducks, a virus infection can affect the glucose metabolism by disrupting the glucose tolerance in the liver and other organs [10,11].
DuCV, a non-enveloped virus, belongs to the Circovirus genus of the Circovirus family and has a diameter of approximately 15–16 nm. Its viral genome is a single-stranded circular DNA of around 2.0 kilobases (kb) and contains two major open reading frames (ORFs) that encode the Rep gene and Cap proteins [12,13]. DuCVs are currently classified into two genotypes, DuCV-1 and DuCV-2, based on the complete genome and Cap gene sequence [14,15,16]. The prevalence of DuCV has been observed in various duck breeds, including Cherry Valley ducks, Pekin ducks, Muscovy ducks, mule ducks, and wild ducks [17,18,19,20,21,22,23]. The published data have demonstrated that a DuCV infection primarily affects the immune system and leads to immunosuppression, making the ducks more susceptible to other infections.
DHAV, a non-enveloped, single positive-stranded RNA virus, belongs to the Avihepatovirus genus in the Picornaviridae family [24,25]. The genome of DHAV contains an open reading frame (ORF) that encodes an inactive precursor protein. This precursor protein is subsequently cleaved into multiple viral proteins, including the structural protein P1 region (capsid proteins VP0, VP1, and VP3) and the non-structural proteins P2 region (2A, 2B, and 2C proteins) and P3 region (3A, 3B, 3C, and 3D proteins) [26,27,28,29,30,31]. DHAV is classified into three serotypes, DHAV-1, DHAV-2, and DHAV-3, based on the VP1 gene sequence. Currently, the most prevalent serotypes in China are DHAV-1 and DHAV-3 [32,33,34,35,36]. DHAV-1 is the classical serotype found worldwide. In China, the use of an officially approved live-attenuated vaccine for breeder ducks since 2013 has effectively controlled DHAV-1 infections. However, a high proportion of DHAV infections caused by DHAV-3 has been observed in China, which can cause liver damage, immune suppression, and death in some duck breeds [35,37,38].
ARV, a non-enveloped virus with a particle size from 70 to 80 nm, belongs to the Orthoreovirus genus of the Reoviridae family [39,40,41]. The genomes of ARV contain ten double-stranded RNA segments separated into three size classes, large (L1, L2, and L3), medium (M1, M2, and M3), and small (S1, S2, S3, and S4) according to their electrophoretic mobility, which encode twelve proteins, large (λA, λB, and λC), medium (μA, μB, and μC), and small (σC, P10, P17, σA, σB, and σNS) [42,43,44,45]. The σC is the most variable protein of ARV, which contains specific epitopes and is used as a genetic marker [46,47,48,49]. The phylogenetic analysis of the σC gene has showed that ARVs are divided into six genotypic clusters [50,51]. The ARVs are divergent in their pathogenicity and can infect a wide variety of avian species [51].
Despite the previous reports on these viruses, the epidemiological characteristics and co-infection rate of each virus in ducks have not been reported. Here, to understand the epidemiology and distribution of DHBV, DuCV, DHAV-3, and ARV in ducks, a surveillance study was conducted involving the PCR- and RT-PCR-based screening of anal swab samples collected from a duck population. To explore the genetic diversity of these detected viruses, the nucleotide sequences were analyzed and compared with some previously reported strains.
2. Materials and Methods
2.1. Specimen Collection
Between September 2020 and January 2022, a total of 271 fresh duck anal swabs were collected. The swabs were from two breeds of ducks from the same duck factory, including 152 Jinding duck swabs and 119 Shaoxing duck swabs. The fresh swab samples were stored in RNase-free containers at −80 °C.
2.2. Viral DNA/RNA Extraction and Reverse Transcription
Viral DNA/RNA was extracted from swabs following the procedure of a viral genome DNA/RNA extraction kit (Tiangen Biotech, Beijing, China). To determine the presence or absence of two RNA viruses, reverse transcription (RT) was initially performed to obtain cDNAs using a reverse transcription kit (TakaRa, Dalian, China).
2.3. PCR
To detect DNA viruses (DuCV and DHBV), viral DNA was extracted and used as the template. For RNA viruses (DHAV-1, DHAV-3, and ARV), viral RNA was extracted and used as the template to obtain cDNA using a reverse transcription kit. Following this, PCR was performed using the primers listed in Table 1. The positive PCR products were then recovered and ligated to a pMD18T vector for sequencing. To conduct a more detailed analysis of the molecular evolvement of these viruses, the full-length sequences of DuCV, the DHAV-3 VP1 gene fragment, and the ARV σC gene fragment were amplified using the primers provided in Table 2 and sequenced with some positive samples.
2.4. DNA Sequences and Phylogenetic Analysis
BLASTN was used to compare the nucleotide sequences with those available in the NCBI nucleotide database. MegAlign was used to compare the sequence homologies. Phylogenetic trees of three viruses were constructed using the neighbor joining and the Kimura 2-parameter model with the bootstrap analysis (1000 replicates) method in MEGA7.0. MegAlign 7.1.0 software was used to compare and analyze some amino acid sequences. Subsequently, ProtParam tool on the ExPASy website (Available from: http://www.expasy.org, accessed on 6 March 2023) was used to analyze the physicochemical properties of the strains obtained in this study and those previously popular strains (only marked with different amino acid sites), including the theoretical pI, instability index, aliphatic index, and grand average of hydropathicity (GRAVY).
3. Results
3.1. Prevalence of DHBV, DuCV, DHAV-3, and ARV in Ducks
In this study, a total of 271 anal swabs collected from Jinding and Shaoxing ducklings were analyzed using PCR and RT-PCR techniques to detect the presence of DHBV, DuCV, DHAV-3, and ARV (Figure 1). The positive samples were randomly selected and further confirmed by sequencing before conducting large-scale screening. Out of the total samples tested, 94.10% (255/271) were positive for at least one of the target viruses. The positive rates for DuCV, DHAV-3, DHBV, and ARV were 39.48%, 8.49%, 69.74%, and 19.92%, respectively (Table 3).
3.2. Coinfection of DHBV, DuCV, DHAV-3, and ARV
Furthermore, we conducted the analysis of the co-infection rates of the four viruses in ducks. Among the ducks that tested positive for at least one target virus, a co-infection rate of 38.04% (97/255) was observed. In addition, co-infections with three or even four viruses were also detected in the ducks (Figure 2). Out of the 255 ducks that were infected with at least one target virus, 22.75% (58/255) were found to be co-infected with DuCV and DHBV, 10.98% (28/255) were found to be co-infected with DuCV and ARV, 10.20% (26/255) were found to be co-infected with ARV and DHBV, and 1.96% (5/255) were found to be co-infected with DuCV, DHAV-3, and DHBV (Table 4).
3.3. Sequencing and Phylogenetic Analysis
To better understand the prevalent strains in the ducks, some positive samples were randomly selected for sequencing. Thus, no significant sequence variation was observed between the DHBV samples that tested positive in this study and the reference DHBV strains from GenBank. Therefore, further sequence analysis for DHBV was not conducted in the subsequent experiment.
The whole-genome sequences of four DuCV strains were obtained in this study (GenBank accession number: OQ658500-OQ658503). A phylogenetic tree of DuCVs was constructed by using the whole-genome sequences of these four newly DuCVs strains and forty-two reference strains from GenBank. The results showed that the four DuCV strains were clustered under the DuCV-1 genotype evolutionary branch with reference strains from China, the United States, Korea, and Brazil (Figure 3a). The nucleotide identity between them ranged from 82.4% to 99.8% (Table 5). Further sequence analysis showed that these four newly identified DuCV strains had 99.8% or higher nucleotide sequence similarity to the reference strain GD/ZQ/144 (ON227548) from Guangdong province in China. Amino acid sequence analysis revealed that the Cap proteins from these four DuCV-1 strain were identical (Table 6). However, several amino acid residues in the Cap protein of the four strains differed significantly from those in the reference strains (Figure 3b).
Five DHAV-3 positive samples were tested for VP1 gene amplification, and the sequence analysis revealed that these samples had identical sequences. The only GenBank accession number available for these samples is OQ658504. A phylogenetic tree was constructed based on the VP1 gene from the DHAV-3 strain identified here, along with 48 reference strains. The analysis results showed that the DHAV-3 strain identified here was closely clustered with the strains from Heihongjiang, Sichuan, and Shandong provinces in China, and this stain shared the most of its nucleotide identity, 99.72%, with one of the DHAV-3 strains from Heilongjiang province (Figure 4a). The homology in the VP1 gene sequence between the DHAV-3 strain here and the reference strains ranged from 66.9% to 100% (Table 7). Subsequent analysis revealed variations in multiple amino acid residues (Figure 4b), which aligned with the patterns observed in recent epidemic strains.
One σC gene was obtained from a positive ARV sample (GenBank accession number: OR046324) and used to conduct phylogenetic classification. The analysis was based on the σC gene alignment of the identified ARV strain and 49 reference strains. The results showed that the ARV strain detected in this study was clustered with the strains from China, Taiwan, the United States, Korea, and Brazil. The genetic evolution of the ARV σC gene showed that the identified ARV strain was distributed in branch I, which contains the largest number of reference strains in genotype I (Figure 5a). The nucleotide sequence homology of the σC gene showed that the ARV strain obtained here was the most homologous with other genotypes I strains, ranging from 75.7% to 98.3%. This was followed by genotype II, with a range from 54.2% to 60.5%; genotype III, with a range from 56.9% to 57.8%; genotype IV, with a range from 53.4% to 54.2%; genotype V, with a range from 51.1% to 54.0%; and genotype VI, with a range from 49.5% to 49.7% (Table 8). The nucleotide homologies of the ARV strain here with the strains from Canada (L39002), the United States (EF122836), Taiwan (AF204947), Brazil (DQ868789), and India (EU681254) were 98.2%, 98.8%, 98.0%, 99.5%, and 98.9%, respectively. These findings indicate that the ARV strain is widely distributed worldwide and has a significant global impact. The results OF amino acid sequence analysis showed that the observed changes in these amino acid residues are likely associated with the virus’s prevalence and spread, taking into account the year and region information (Figure 5b). These changes are consistent with those observed in recent epidemic strains.
In this study, the physicochemical properties of the obtained sequences were determined. The instability coefficients of these proteins were analyzed, taking into account the influence of different amino acid sites on their physicochemical properties (Table 9). The results showed that the instability coefficients of these proteins were significantly different, indicating variations in their structural stability. The stability of the viral proteins is crucial for their proper folding, assembly, and function. Changes in the physicochemical properties of viral proteins can affect their stability, potentially leading to alterations in viral replication, infectivity, and host interactions. Therefore, the observed differences in the instability coefficients of these proteins may contribute to the variations in the prevalence of different virus strains.
4. Discussion
China holds the distinction of being the largest producer and consumer of waterfowl globally. In fact, China’s duck production accounts for over 80% of the total worldwide quantity, making duck the third most consumed meat in the country, following pork and chicken [52]. As the duck breeding industry continues to intensify, the incidence of viral infectious diseases in ducks is also on the rise. In this study, we gathered evidence supporting the presence of four vertically transmitted viruses in duck populations, including DuCV, DHAV-3, DHBV, and ARV. We also observed the co-infections of these four viruses and evaluated the genotypes of DuCV, DHAV-3, and ARV. Additionally, the amino acid sequence and physicochemical properties of the proteins were analyzed.
DuCV, a novel virus, has garnered significant attention from the veterinary industry and has been reported in several countries, including Germany, Hungary, Taiwan in China, South Korea, and more [20,53,54,55]. Currently, DuCV is widely prevalent among the domestic duck populations [17,56]. DuCV infections in domestic ducks are typically subclinical and primarily affect the host’s immune system, leading to various secondary infections [14,53,57,58,59,60]. The high prevalence of DcCV has been reported in asymptomatic duck populations in China, with ducks of various age groups being susceptible to the virus. The DuCV-positive ducks exhibited a higher rate of infection by DHV-I, Riemerella anatipestifer, and E. coli compared to that of the DuCV-negative ducks [61,62]. In this study, we have identified four DuCV strains in the duck populations. Through sequence analysis, we determined that these viruses belong to genotype 1 and are closely clustered with the strains from Shandong, Hebei, and Guangdong provinces in China. Among the two genotypes of the virus, DuCV-1 and DuCV-2, there have been more reported cases of DuCV-1 in countries such as Germany, Hungary, the United States, China, South Korea, and Poland. However, phylogenetic analysis indicates that there is a significant variation in the genotype of DuCV in Mainland China, where both genotype 1 and genotype 2 are widely distributed and prevalent [60,63]. And previous epidemiological investigations have shown DuCV-1 to be more widespread and more pathogenic than DuCV-2 is [60]. Moreover, DuCV has been linked to emerging diseases, such as “beak atrophy and dwarfism syndrome (BADS)” and primary sclerosing cholangitis, which can cause severe clinical symptoms [12,18]. It is worth noting that gene recombination has consistently played a crucial role in the virus’ evolution. Several recombination events have been observed in circoviruses, such as recombination between porcine circovirus 2, porcine circovirus 3, goose circovirus, and DuCV [64,65,66,67,68]. However, our study did not yield sufficient evidence of recombination among the four DuCV isolates sequenced in this study. This limited evidence could be attributed to the limited number of DuCV sequences included in the analysis. Despite this, our findings indicate that these sequences align with significant changes in the amino acid sequence of the Cap protein observed in recent years. Additionally, our analysis based on the regional and yearly data suggest that 2020 may represent a crucial time period for significant changes in multiple amino acid sites, as depicted in Figure 3b. The pathobiological characteristics of DuCV-1 encompass systemic infection, persistent infection, as well as horizontal and vertical transmission [60]. Regrettably, despite advancements in the vaccine research, the absence of a robust culturing system has hampered the development of effective prevention methods and satisfactory antiviral drugs for DuCV. This limitation has posed challenges in both DuCV vaccine development and antiviral research [69,70].
DHAV is responsible for a severe and fast-spreading viral infections among young ducklings, characterized by high fatality rates reaching 95% and posing a significant economic risk to the duck industry [71]. On the other hand, adult ducks typically exhibit resistance to DHAV and do not show apparent clinical symptoms upon infection. However, the infected adult ducks can still shed the virus, serving as a potential source of infection for others [6]. However, a recent study has reported that a DHAV-1 infection can potentially trigger egg drop syndrome [72,73]. Furthermore, epidemiological investigations have indicated that following the widespread use of the officially approved DHAV-1 live vaccine in 2013, an increased incidence of DHAV infections caused by DHAV-3 was observed. In fact, the DHAV-3 strain has emerged as the predominant viral type in China [35,37,38,74,75]. In the present investigation, there were no instances of DHAV-1 detection, and therefore, the focus was solely on the genetic evolution analysis of the identified DHAV-3 strain. The identified DHAV-3 strain exhibited a significant relationship with the Sichuan and Shandong strains (MN912703, KX523286, and KX523289) from China, as they formed a distinct cluster within the evolutionary branch associated with the DHAV-3 genotype. Our analysis determined that this particular strain exhibited a close genetic relationship with the MN164467.1 strain, which was isolated from Heilongjiang, China. The nucleotide homology between them reached 99.72%. Notably, our findings indicated that the sequence displayed notable alterations in the amino acid sequence of the VP1 protein, reflecting the significant changes that have occurred in recent years. Despite these modifications, our analysis did not reveal any significant impact on the physicochemical properties of the proteins. However, further investigations are required to assess whether these alterations influence viral adsorption and the other related functions.
The σC protein of ARV encoded by the third open reading frame of the S1 genome fragment is a minor component of the virion coat and plays a crucial role in cell attachment and the induction of neutralizing antibodies [76]. Moreover, the ARV σC protein has a variety of functions, such as inducing apoptosis, an anti-tumor effect, and immune enhancement [77]. Notably, mutations in the σC protein are responsible for segment rearrangement, leading to genetic diversity within the reovirus population [72,76]. Mutations in the amino acid sites of this protein can lead to changes in pathogen infectivity, resulting in antigenic variation. Other studies have shown that when the amino acid sequence difference between the vaccine strain and the circulating strain is equal to or greater than 5%, the cross-protection effect of the vaccine is weakened [77]. The nucleotide and amino acid homologies between the σC sequence obtained in this study and the σC sequence of the S1133 vaccine strain were 98.8% and 97.9%, respectively. The amino acid sequence encoded by the major protective antigen σC gene of the current commercial vaccine strain S1133 is highly homologous with that of the ARV strain identified in this study, so we speculate that the existing vaccine can produce a protective effect against this variant strain. Other studies have shown significant genetic changes in certain regions of the ARVs genome and the emergence of ARVs genetic variation prior to 1990 [78]. Consequently, it is crucial to promptly comprehend the epidemic situation and pathogenic characteristics of ARV, and effectively manage the outbreak and prevalence of the disease by enhancing the prevention and control measures, bolstering the research and development of novel vaccines, and implementing a series of measures.
The DHAV-3 VP1 and ARV σC sequences obtained in this study were closely related to those isolated and identified in Heilongjiang province. We hypothesize that virus transmission may have occurred due to trade within the province. On the other hand, the DuCV strains were closely clustered with the strains from Shandong, Hebei, and Guangdong. The similarity of DuCV strains in these areas may be related to the migration of wild ducks. In addition, these sequences are consistent with the changes in the amino acid sequences of major immunogenic proteins of epidemic strains in recent years. Furthermore, the amino acid sequences of the major immunogenic proteins of these epidemic strains have shown changes in recent years. This suggests ongoing viral evolution and adaptation, which may have implications for the effectiveness of existing vaccines and the development of new preventive measures. In order to gain a more comprehensive understanding of the vertical transmission of related viruses in ducks in mainland China, enhance biosafety breeding, and reinforce the prevention and control programs, it is necessary to broaden the geographical scope of the survey and contribute to the overall understanding and management of viral infectious diseases in the ducks in mainland China.
5. Conclusions
This study revealed the prevalence of DuCV, DHAV-3, DHBV, and ARV in ducks. It also demonstrates the coexistence of these viruses in healthy adult ducks. The detection of DuCV, DHAV-3, DHBV, and ARV in healthy adult ducks suggests a high likelihood of vertical transmission, leading to infections among ducklings. Hence, further research is required to gain a deeper understanding of these viruses’ role in ducks and their potential health implications.
Author Contributions
Conceptualization, X.W. and H.Y.; methodology, X.W. and H.Y.; software, X.W. and H.Y.; validation, X.W., H.Y. and W.Z.; formal analysis, X.W., H.Y. and W.Z.; investigation, X.W., H.Y. and W.Z.; resources, L.F. and Y.W.; data curation, X.W., H.Y. and W.Z.; writing—original draft preparation, X.W., H.Y. and Y.W.; writing—review and editing, L.F. and Y.W.; visualization, H.Y. and W.Z.; supervision, W.Z. and Y.W.; project administration, L.F. and Y.W.; funding acquisition, Y.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Fundamental Research Funds for the Central Universities; grant number: SWU-KR22036.
Institutional Review Board Statement
Ethical review and approval were waived for this study, as the anal samples were collected from ducks that were raised for commercial purposes and were not subjected to any experimental procedures.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The virus strains obtained in this study are available in GenBank under accession nos. OQ658500 for DB2022-1/Heilongjiang/2022, OQ658501 for DB2022-2/Heilongjiang/2022, OQ658502 for DB2022-3/Heilongjiang/2022, OQ658503 for DB2022-4/Heilongjiang/2022, QQ659504 for DHAV3/DB2022-5/Heilongjiang/2022, and OR046324 for DB2022-6/Heilongjiang/2022.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Patil, S.S.; Shinduja, R.; Suresh, K.P.; Phukan, S.; Kumar, S.; Sengupta, P.P.; Amachawadi, R.G.; Raut, A.; Roy, P.; Syed, A.; et al. A systematic review and meta-analysis on the prevalence of infectious diseases of Duck: A world perspective. Saudi J. Biol. Sci. 2021, 28, 5131–5144. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Wang, X.; Zhang, R.; Chen, J.; Xia, L.; Lin, S.; Xie, Z.; Jiang, S. Evidence of possible vertical transmission of duck circovirus. Vet. Microbiol. 2014, 174, 229–232. [Google Scholar] [CrossRef] [PubMed]
- Tagawa, M.; Robinson, W.S.; Marion, P.L. Duck hepatitis B virus replicates in the yolk sac of developing embryos. J. Virol. 1987, 61, 2273–2279. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, Y.; Gao, B.; Zhang, S.; Diao, Y.; Tang, Y. Evidence of vertical transmission of novel duck orthoreovirus in ducks. Vet. Microbiol. 2020, 251, 108861. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Yang, Y.; Lan, J.; Xie, Z.; Zhang, X.; Jiang, S. Evidence of possible vertical transmission of duck hepatitis A virus type 1 in ducks. Transbound. Emerg. Dis. 2021, 68, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wu, S.; Liu, W.; Hu, Z. Current status and future direction of duck hepatitis A virus vaccines. Avian Pathol. 2023, 52, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Mason, W.S.; Seal, G.; Summers, J. Virus of Pekin ducks with structural and biological relatedness to human hepatitis B virus. J. Virol. 1980, 36, 829–836. [Google Scholar] [CrossRef] [PubMed]
- Uchida, T.; Suzuki, K.; Esumi, M.; Arii, M.; Shikata, T. Influence of aflatoxin B1 intoxication on duck livers with duck hepatitis B virus infection. Cancer Res. 1988, 48, 1559–1565. [Google Scholar]
- Zhang, D.; Wang, Y.; He, Y.; Ji, L.; Zhao, K.; Yang, S.; Zhang, W. Identification of avihepadnaviruses and circoviruses in an unexplained death event in farmed ducks. Arch. Virol. 2023, 168, 85. [Google Scholar] [CrossRef]
- Liu, Q.; Jia, R.; Wang, M.; Huang, J.; Zhu, D.; Chen, S.; Yin, Z.; Wang, Y.; Chen, X.; Cheng, A. Cloning, expression and purification of duck hepatitis B virus (DHBV) core protein and its use in the development of an indirect ELISA for serologic detection of DHBV infection. Arch. Virol. 2014, 159, 897–904. [Google Scholar] [CrossRef]
- Tan, Y.; Liu, J.; Qin, Y.; Liang, B.; Gu, Y.; Liang, L.; Liu, L.; Liu, Y.; Su, H. Glucose Homeostasis Is Dysregulated in Ducks Infected with Duck Hepatitis B Virus. Intervirology 2021, 64, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Zhou, D.; Liu, J.; Hao, X.; Cheng, Z. Duck circovirus induces a new pathogenetic characteristic, primary sclerosing cholangitis. Comp. Immunol. Microbiol. Infect. Dis. 2019, 63, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Liu, N.; Zhang, L.; Jiang, W.; Fan, X.; Wang, X.; Miao, R.; Zhai, X.; Wei, L.; Jiang, S.; et al. Research Note: Complete genome cloning and genetic evolution analysis of four Cherry Valley duck circovirus strains in China in 2022. Poult. Sci. 2023, 102, 102920. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Jia, R.; Lu, Y.; Wang, M.; Zhu, D.; Chen, S.; Yin, Z.; Chen, X.; Cheng, A. Identification, genotyping, and molecular evolution analysis of duck circovirus. Gene 2013, 529, 288–295. [Google Scholar] [CrossRef] [PubMed]
- Wen, H.; Wu, Y.; Yang, C.; Zhang, X.; Lian, C.; Chen, H.; Han, L. Comments on duck circovirus (DuCV) genotype definition. Gene 2014, 538, 207–208. [Google Scholar] [CrossRef]
- Shen, M.; Gao, P.; Wang, C.; Li, N.; Zhang, S.; Jiang, Y.; Liu, D.; Jia, B.; Xu, L.; Huang, B.; et al. Pathogenicity of duck circovirus and fowl adenovirus serotype 4 co-infection in Cherry Valley ducks. Vet. Microbiol. 2023, 279, 109662. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Li, L.X.; Sun, W.C.; Shi, N.; Sun, X.T.; Jin, N.Y.; Si, X.K. Molecular survey of duck circovirus infection in poultry in southern and southwestern China during 2018 and 2019. BMC Vet. Res. 2020, 16, 80. [Google Scholar] [CrossRef]
- Li, P.; Li, J.; Zhang, R.; Chen, J.; Wang, W.; Lan, J.; Xie, Z.; Jiang, S. Duck “beak atrophy and dwarfism syndrome” disease complex: Interplay of novel goose parvovirus-related virus and duck circovirus? Transbound. Emerg. Dis. 2018, 65, 345–351. [Google Scholar] [CrossRef]
- Banda, A.; Galloway-Haskins, R.I.; Sandhu, T.S.; Schat, K.A. Genetic analysis of a duck circovirus detected in commercial Pekin ducks in New York. Avian Dis. 2007, 51, 90–95. [Google Scholar] [CrossRef]
- Chen, C.L.; Wang, P.X.; Lee, M.S.; Shien, J.H.; Shien, H.K.; Ou, S.J.; Chen, C.H.; Chang, P.C. Development of a polymerase chain reaction procedure for detection and differentiation of duck and goose circovirus. Avian Dis. 2006, 50, 92–95. [Google Scholar] [CrossRef]
- Matczuk, A.K.; Krawiec, M.; Wieliczko, A. A new duck circovirus sequence, detected in velvet scoter (Melanitta fusca) supports great diversity among this species of virus. Virol. J. 2015, 12, 121. [Google Scholar] [CrossRef] [PubMed]
- Liao, J.Y.; Xiong, W.J.; Tang, H.; Xiao, C.T. Identification and characterization of a novel circovirus species in domestic laying ducks designated as duck circovirus 3 (DuCV3) from Hunan province, China. Vet. Microbiol. 2022, 275, 109598. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Zhang, Y.; Cheng, A.; Wang, M.; Liu, M.; Zhu, D.; Chen, S.; Zhao, X.; Yang, Q.; Wu, Y.; et al. Duck Circovirus genotype 2 ORF3 protein induces apoptosis through the mitochondrial pathway. Poult. Sci. 2023, 102, 102533. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.; Cheng, A.; Wang, M.; Ou, X.; Sun, D.; Zhang, S.; Mao, S.; Yang, Q.; Tian, B.; Wu, Y.; et al. DHAV 3CD targets IRF7 and RIG-I proteins to block the type I interferon upstream signaling pathway. Vet. Res. 2023, 54, 5. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, M.; Zhou, S.; Cheng, A.; Ou, X.; Sun, D.; Wu, Y.; Yang, Q.; Gao, Q.; Huang, J.; et al. The DHAV-1 protein VP1 interacts with PI3KC3 to induce autophagy through the PI3KC3 complex. Vet. Res. 2022, 53, 64. [Google Scholar] [CrossRef] [PubMed]
- Wen, X.; Cheng, A.; Wang, M.; Jia, R.; Zhu, D.; Chen, S.; Liu, M.; Sun, K.; Yang, Q.; Wu, Y.; et al. Recent advances from studies on the role of structural proteins in enterovirus infection. Future Microbiol. 2015, 10, 1529–1542. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Ou, X.; Zhu, D.; Ma, G.; Cheng, A.; Wang, M.; Chen, S.; Jia, R.; Liu, M.; Sun, K.; et al. The 2A2 protein of Duck hepatitis A virus type 1 induces apoptosis in primary cell culture. Virus Genes 2016, 52, 780–788. [Google Scholar] [CrossRef]
- Sun, D.; Chen, S.; Cheng, A.; Wang, M. Roles of the Picornaviral 3C Proteinase in the Viral Life Cycle and Host Cells. Viruses 2016, 8, 82. [Google Scholar] [CrossRef]
- Sun, D.; Wang, M.; Wen, X.; Cheng, A.; Jia, R.; Sun, K.; Yang, Q.; Wu, Y.; Zhu, D.; Chen, S.; et al. Cleavage of poly(A)-binding protein by duck hepatitis A virus 3C protease. Sci. Rep. 2017, 7, 16261. [Google Scholar] [CrossRef]
- Yang, C.; Shah, P.T.; Bahoussi, A.N.; Wu, C.; Wang, L.; Xing, L. Duck hepatitis a virus: Full-length genome-based phylogenetic and phylogeographic view during 1986–2020. Virus Res. 2023, 336, 199216. [Google Scholar] [CrossRef]
- Zhou, S.; Li, S.; Wang, Y.; Li, X.; Zhang, T. Duck hepatitis A virus prevalence in mainland China between 2009 and 2021: A systematic review and meta-analysis. Prev. Vet. Med. 2022, 208, 105730. [Google Scholar] [CrossRef] [PubMed]
- Tseng, C.H.; Tsai, H.J. Molecular characterization of a new serotype of duck hepatitis virus. Virus Res. 2007, 126, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.C.; Kwon, Y.K.; Joh, S.J.; Kim, S.J.; Tolf, C.; Kim, J.H.; Sung, H.W.; Lindberg, A.M.; Kwon, J.H. Recent Korean isolates of duck hepatitis virus reveal the presence of a new geno- and serotype when compared to duck hepatitis virus type 1 type strains. Arch. Virol. 2007, 152, 2059–2072. [Google Scholar] [CrossRef] [PubMed]
- Wen, X.J.; Cheng, A.C.; Wang, M.S.; Jia, R.Y.; Zhu, D.K.; Chen, S.; Liu, M.F.; Liu, F.; Chen, X.Y. Detection, differentiation, and VP1 sequencing of duck hepatitis A virus type 1 and type 3 by a 1-step duplex reverse-transcription PCR assay. Poult. Sci. 2014, 93, 2184–2192. [Google Scholar] [CrossRef] [PubMed]
- Wen, X.; Zhu, D.; Cheng, A.; Wang, M.; Chen, S.; Jia, R.; Liu, M.; Sun, K.; Zhao, X.; Yang, Q.; et al. Molecular epidemiology of duck hepatitis a virus types 1 and 3 in China, 2010–2015. Transbound. Emerg. Dis. 2018, 65, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.; Xia, X.; Cheng, A.; Wang, M.; Ou, X.; Mao, S.; Sun, D.; Zhang, S.; Yang, Q.; Wu, Y.; et al. DHAV-1 Blocks the Signaling Pathway Upstream of Type I Interferon by Inhibiting the Interferon Regulatory Factor 7 Protein. Front. Microbiol. 2021, 12, 700434. [Google Scholar] [CrossRef]
- Wu, F.; Lu, F.; Fan, X.; Chao, J.; Liu, C.; Pan, Q.; Sun, H.; Zhang, X. Immune-related miRNA-mRNA regulation network in the livers of DHAV-3-infected ducklings. BMC Genom. 2020, 21, 123. [Google Scholar] [CrossRef]
- Cao, J.; Zhang, Y.; Chen, Y.; Liang, S.; Liu, D.; Fan, W.; Xu, Y.; Liu, H.; Zhou, Z.; Liu, X.; et al. Dynamic Transcriptome Reveals the Mechanism of Liver Injury Caused by DHAV-3 Infection in Pekin Duck. Front. Immunol. 2020, 11, 568565. [Google Scholar] [CrossRef]
- Jones, R.C. Avian reovirus infections. Rev. Sci. Tech. 2000, 19, 614–625. [Google Scholar] [CrossRef]
- Varela, R.; Benavente, J. Protein coding assignment of avian reovirus strain S1133. J. Virol. 1994, 68, 6775–6777. [Google Scholar] [CrossRef]
- Farnoushi, Y.; Heller, D.; Lublin, A. Development of a wide-range real-time RT-PCR assay for detection of Avian reovirus (ARV). J. Virol. Methods 2022, 310, 114613. [Google Scholar] [CrossRef] [PubMed]
- Benavente, J.; Martínez-Costas, J. Avian reovirus: Structure and biology. Virus Res. 2007, 123, 105–119. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Zou, Z.; Song, S.; Liu, H.; Gong, X.; Li, B.; Liu, P.; Wang, Q.; Liu, F.; Luan, D.; et al. Epidemiological Analysis of Avian Reovirus in China and Research on the Immune Protection of Different Genotype Strains from 2019 to 2020. Vaccines 2023, 11, 485. [Google Scholar] [CrossRef] [PubMed]
- Egana-Labrin, S.; Broadbent, A.J. Avian reovirus: A furious and fast evolving pathogen. J. Med. Microbiol. 2023, 72, 001761. [Google Scholar] [CrossRef] [PubMed]
- Goldenberg, D. Avian Reovirus in Israel, Variants and Vaccines-A Review. Avian Dis. 2022, 66, 447–451. [Google Scholar] [CrossRef]
- Goldenberg, D.; Pasmanik-Chor, M.; Pirak, M.; Kass, N.; Lublin, A.; Yeheskel, A.; Heller, D.; Pitcovski, J. Genetic and antigenic characterization of sigma C protein from avian reovirus. Avian Pathol. 2010, 39, 189–199. [Google Scholar] [CrossRef]
- Kant, A.; Balk, F.; Born, L.; van Roozelaar, D.; Heijmans, J.; Gielkens, A.; ter Huurne, A. Classification of Dutch and German avian reoviruses by sequencing the sigma C protein. Vet. Res. 2003, 34, 203–212. [Google Scholar] [CrossRef]
- Liu, H.J.; Lee, L.H.; Shih, W.L.; Li, Y.J.; Su, H.Y. Rapid characterization of avian reoviruses using phylogenetic analysis, reverse transcription-polymerase chain reaction and restriction enzyme fragment length polymorphism. Avian Pathol. 2004, 33, 171–180. [Google Scholar] [CrossRef]
- Troxler, S.; Rigomier, P.; Bilic, I.; Liebhart, D.; Prokofieva, I.; Robineau, B.; Hess, M. Identification of a new reovirus causing substantial losses in broiler production in France, despite routine vaccination of breeders. Vet. Rec. 2013, 172, 556. [Google Scholar] [CrossRef]
- Lu, H.; Tang, Y.; Dunn, P.A.; Wallner-Pendleton, E.A.; Lin, L.; Knoll, E.A. Isolation and molecular characterization of newly emerging avian reovirus variants and novel strains in Pennsylvania, USA, 2011–2014. Sci. Rep. 2015, 5, 14727. [Google Scholar] [CrossRef]
- Ayalew, L.E.; Gupta, A.; Fricke, J.; Ahmed, K.A.; Popowich, S.; Lockerbie, B.; Tikoo, S.K.; Ojkic, D.; Gomis, S. Phenotypic, genotypic and antigenic characterization of emerging avian reoviruses isolated from clinical cases of arthritis in broilers in Saskatchewan, Canada. Sci. Rep. 2017, 7, 3565. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, L.; Shang, H.; Zhou, F.; Wang, C.; Zhang, S.; Gao, P.; Guo, P.; Zhu, R.; Sun, Z.; et al. Effects of duck circovirus on immune function and secondary infection of Avian Pathogenic Escherichia coli. Poult. Sci. 2022, 101, 101799. [Google Scholar] [CrossRef]
- Hattermann, K.; Schmitt, C.; Soike, D.; Mankertz, A. Cloning and sequencing of Duck circovirus (DuCV). Arch. Virol. 2003, 148, 2471–2480. [Google Scholar] [CrossRef] [PubMed]
- Fringuelli, E.; Scott, A.N.; Beckett, A.; McKillen, J.; Smyth, J.A.; Palya, V.; Glavits, R.; Ivanics, E.; Mankertz, A.; Franciosini, M.P.; et al. Diagnosis of duck circovirus infections by conventional and real-time polymerase chain reaction tests. Avian Pathol. 2005, 34, 495–500. [Google Scholar] [CrossRef] [PubMed]
- Cha, S.Y.; Kang, M.; Cho, J.G.; Jang, H.K. Genetic analysis of duck circovirus in Pekin ducks from South Korea. Poult. Sci. 2013, 92, 2886–2891. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Xie, X.; Zhang, D.; Ma, G.; Wang, X.; Zhang, D. Detection of duck circovirus in China: A proposal on genotype classification. Vet. Microbiol. 2011, 147, 410–415. [Google Scholar] [CrossRef]
- Soike, D.; Albrecht, K.; Hattermann, K.; Schmitt, C.; Mankertz, A. Novel circovirus in mulard ducks with developmental and feathering disorders. Vet. Rec. 2004, 154, 792–793. [Google Scholar] [CrossRef]
- Zhang, L.; Jiang, W.; Zhang, F.; Li, Y.; Li, J.; Liang, S.; Yu, X.; Peng, C.; Liu, S.; Wang, J.; et al. Development of a dual-labeled, hydrolysis probe-based, real-time quantitative PCR assay for detection of both genotypes of duck circovirus-1 (DuCV-1) and DuCV-2. Virus Genes 2021, 57, 453–458. [Google Scholar] [CrossRef]
- Wan, C.H.; Fu, G.H.; Shi, S.H.; Cheng, L.F.; Chen, H.M.; Peng, C.X.; Lin, S.; Huang, Y. Epidemiological investigation and genome analysis of duck circovirus in Southern China. Virol. Sin. 2011, 26, 289–296. [Google Scholar] [CrossRef]
- Hong, Y.T.; Kang, M.; Jang, H.K. Pathogenesis of duck circovirus genotype 1 in experimentally infected Pekin ducks. Poult. Sci. 2018, 97, 3050–3057. [Google Scholar] [CrossRef]
- Liu, S.N.; Zhang, X.X.; Zou, J.F.; Xie, Z.J.; Zhu, Y.L.; Zhao, Q.; Zhou, E.M.; Jiang, S.J. Development of an indirect ELISA for the detection of duck circovirus infection in duck flocks. Vet. Microbiol. 2010, 145, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Jiang, S.; Wu, J.; Zhao, Q.; Sun, Y.; Kong, Y.; Li, X.; Yao, M.; Chai, T. An investigation of duck circovirus and co-infection in Cherry Valley ducks in Shandong Province, China. Vet. Microbiol. 2009, 133, 252–256. [Google Scholar] [CrossRef] [PubMed]
- Niu, X.; Liu, L.; Han, C.; Li, J.; Zeng, X. First findings of duck circovirus in migrating wild ducks in China. Vet. Microbiol. 2018, 216, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Zheng, M.; Cao, H.; Lu, H.; Wei, X.; Pan, Y.; Zhang, H.; Su, J.; Li, J.; Jin, N. Genome Sequences of a Novel Recombinant Duck Circovirus in China. Genome Announc. 2016, 4, e01181-16. [Google Scholar] [CrossRef] [PubMed]
- Saraiva, G.L.; Vidigal, P.M.P.; Fietto, J.L.R.; Bressan, G.C.; Silva Júnior, A.; de Almeida, M.R. Evolutionary analysis of Porcine circovirus 3 (PCV3) indicates an ancient origin for its current strains and a worldwide dispersion. Virus Genes 2018, 54, 376–384. [Google Scholar] [CrossRef] [PubMed]
- Stenzel, T.; Dziewulska, D.; Muhire, B.M.; Hartnady, P.; Kraberger, S.; Martin, D.P.; Varsani, A. Recombinant Goose Circoviruses Circulating in Domesticated and Wild Geese in Poland. Viruses 2018, 10, 107. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.; Lin, Z.; Dai, A.; Chen, H.; Ma, Y.; Li, N.; Wu, Y.; Yang, X.; Luo, M.; Liu, J. Emergence of a novel recombinant porcine circovirus type 2 in China: PCV2c and PCV2d recombinant. Transbound. Emerg. Dis. 2019, 66, 2496–2506. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, D.; Bai, C.X.; Guo, X.; Gao, W.H.; Li, M.L.; Wang, J.; Li, Y.D. Molecular characteristics of a novel duck circovirus subtype 1d emerging in Anhui, China. Virus Res. 2021, 295, 198216. [Google Scholar] [CrossRef]
- Li, Z.; Fu, G.; Feng, Z.; Chen, J.; Shi, S.; Liu, R.; Cheng, L.; Chen, H.; Wan, C.; Yu, H. Evaluation of a novel inactivated vaccine against duck circovirus in muscovy ducks. Vet. Microbiol. 2020, 241, 108574. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, S.; Shang, H.; Wang, C.; Zhou, F.; Liu, Y.; Jiang, Y.; Gao, P.; Li, N.; Liu, D.; et al. Evaluation of the antiviral effect of four plant polysaccharides against duck circovirus. Res. Vet. Sci. 2022, 152, 446–457. [Google Scholar] [CrossRef]
- Zhang, X.; Cao, C.; Qu, Z.; Zhang, W.; Liu, Y.; Qi, H.; Hao, C.; Zhang, W.; Gao, M.; Wang, J.; et al. Pathogenicity of duck hepatitis A virus type 3 and innate immune responses of the ducklings to virulent DHAV-3. Mol. Immunol. 2018, 95, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Saikia, D.P.; Yadav, K.; Pathak, D.C.; Ramamurthy, N.; D’Silva, A.L.; Marriappan, A.K.; Ramakrishnan, S.; Vakharia, V.N.; Chellappa, M.M.; Dey, S. Recombinant Newcastle Disease Virus (NDV) Expressing Sigma C Protein of Avian Reovirus (ARV) Protects against Both ARV and NDV in Chickens. Pathogens 2019, 8, 145. [Google Scholar] [CrossRef] [PubMed]
- Shih, W.L.; Hsu, H.W.; Liao, M.H.; Lee, L.H.; Liu, H.J. Avian reovirus sigmaC protein induces apoptosis in cultured cells. Virology 2004, 321, 65–74. [Google Scholar] [CrossRef]
- Fehér, E.; Jakab, S.; Bali, K.; Kaszab, E.; Nagy, B.; Ihász, K.; Bálint, Á.; Palya, V.; Bányai, K. Genomic Epidemiology and Evolution of Duck Hepatitis A Virus. Viruses 2021, 13, 1592. [Google Scholar] [CrossRef] [PubMed]
- Mansour, S.M.G.; Mohamed, F.F.; ElBakrey, R.M.; Eid, A.A.M.; Mor, S.K.; Goyal, S.M. Outbreaks of Duck Hepatitis A Virus in Egyptian Duckling Flocks. Avian Dis. 2019, 63, 68–74. [Google Scholar] [CrossRef]
- Liu, H.J.; Lee, L.H.; Hsu, H.W.; Kuo, L.C.; Liao, M.H. Molecular evolution of avian reovirus: Evidence for genetic diversity and reassortment of the S-class genome segments and multiple cocirculating lineages. Virology 2003, 314, 336–349. [Google Scholar] [CrossRef]
- Cavanagh, D. Coronavirus avian infectious bronchitis virus. Vet. Res. 2007, 38, 281–297. [Google Scholar] [CrossRef]
- Egaña-Labrin, S.; Hauck, R.; Figueroa, A.; Stoute, S.; Shivaprasad, H.L.; Crispo, M.; Corsiglia, C.; Zhou, H.; Kern, C.; Crossley, B.; et al. Genotypic Characterization of Emerging Avian Reovirus Genetic Variants in California. Sci. Rep. 2019, 9, 9351. [Google Scholar] [CrossRef]
Figure 1.
Part of PCR or RT-PCR results of DuCV (a), DHBV (b), DHAV-3 (c), and ARV (d).
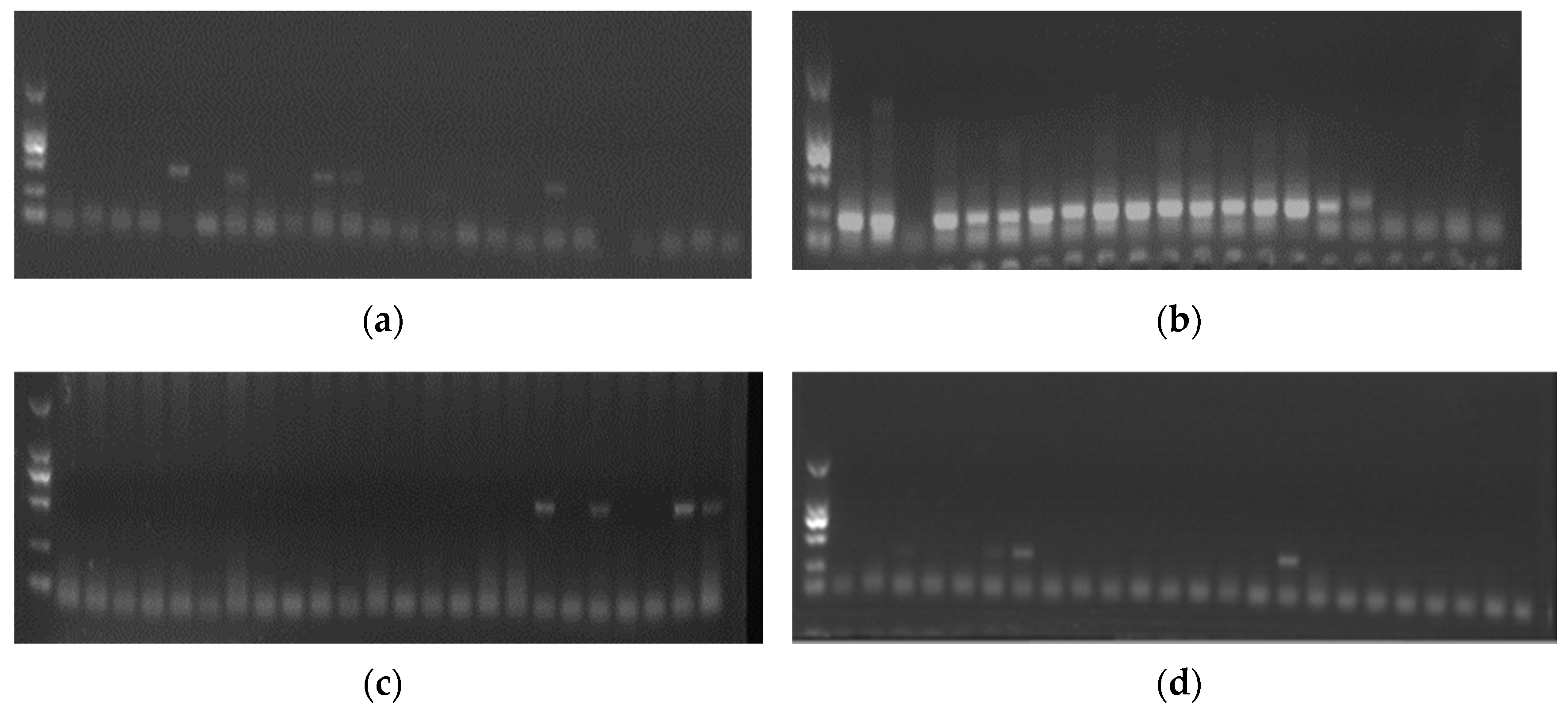
Figure 2.
Coinfection rates of DHBV, DuCV, DHAV-3, and ARV.

Figure 3.
Sequence analysis of DuCV. (a): Phylogenetic tree of DuCV. Strains detected in this study are marked with “⦁”. (b): Amino acid sequence analysis of DuCV-Cap. Strains detected in this study are marked with red color.
Figure 3.
Sequence analysis of DuCV. (a): Phylogenetic tree of DuCV. Strains detected in this study are marked with “⦁”. (b): Amino acid sequence analysis of DuCV-Cap. Strains detected in this study are marked with red color.
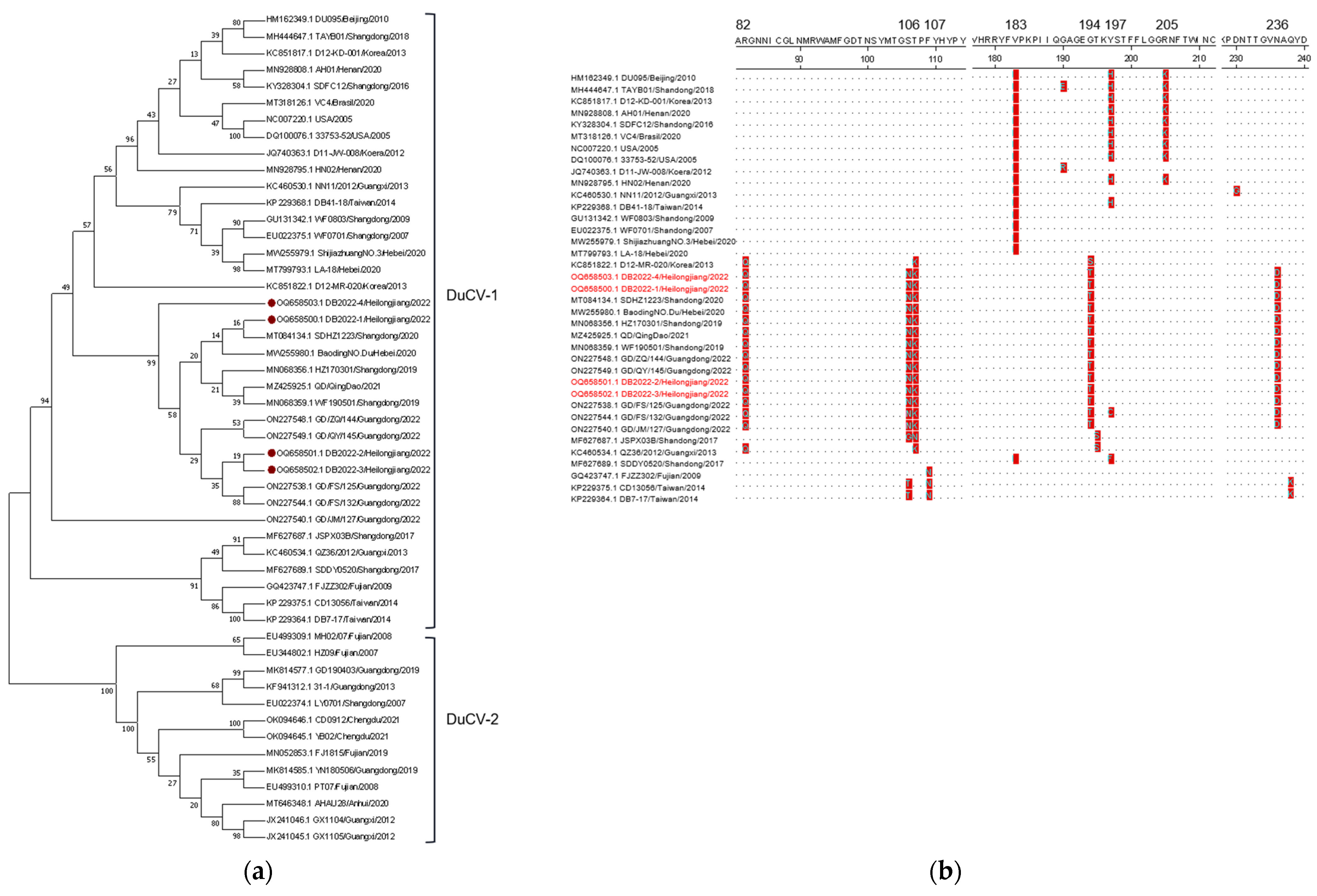
Figure 4.
Sequence analysis of DHAV-VP1. (a): Phylogenetic tree of DHAV. (b): Amino acid sequence analysis of DHAV3-VP1. The DHAV strain identified in this study are marked with “⦁”. The strains detected in this study are marked in red.
Figure 4.
Sequence analysis of DHAV-VP1. (a): Phylogenetic tree of DHAV. (b): Amino acid sequence analysis of DHAV3-VP1. The DHAV strain identified in this study are marked with “⦁”. The strains detected in this study are marked in red.
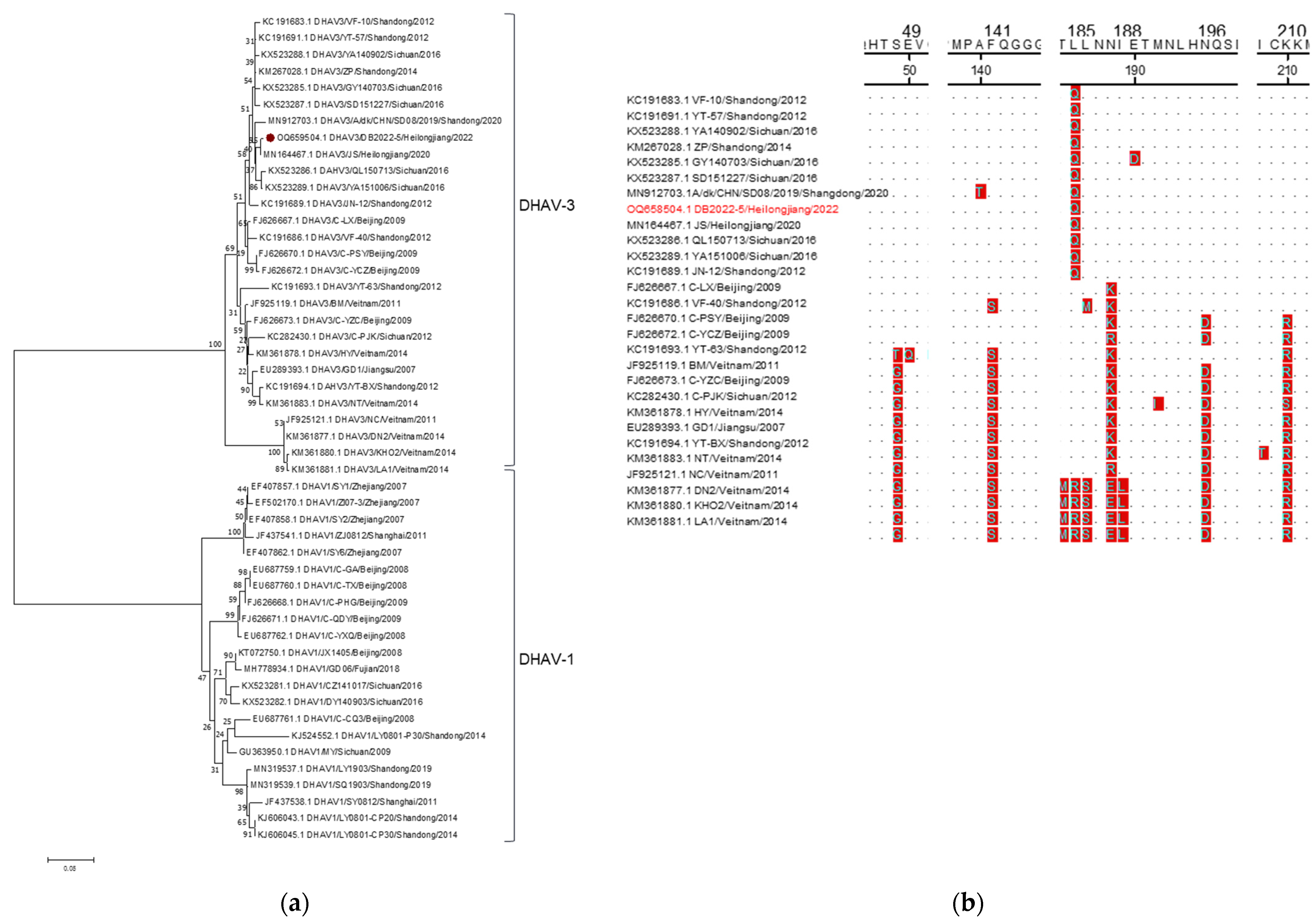
Figure 5.
Sequence analysis of ARV. (a): Phylogenetic tree of ARV. The ARV strain identified in this study are marked with “⦁”. (b) B: Amino acid sequence analysis of ARV-σC. The strain detected in this study is marked in red.
Figure 5.
Sequence analysis of ARV. (a): Phylogenetic tree of ARV. The ARV strain identified in this study are marked with “⦁”. (b) B: Amino acid sequence analysis of ARV-σC. The strain detected in this study is marked in red.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Primers sequences used for detection.
| Virus | Forward Primer (5′-3′) | Reverse Primer (5′-3′) | Fragment Size (bp) |
|---|---|---|---|
| DuCV | gcacgctcgacaattgcaagt | gccacgcccaaagattacataag | 338 |
| DHBV | gggctaggagattgctttg | ggttcgagtccacgaggtt | 217 |
| DHAV-1 | agcttaaggcccggtgccccg | ggtagggtagggaatagtaaagt | 399 |
| DHAV-3 | aacccctttgatccacactg | gataaggcatccacaccatc | 544 |
| ARV | taatttagacggtttgagga | cgttgagaacagaagtaggg | 324 |
Table 2.
Primers sequences used for amplification.
| Virus | Forward Primer (5′-3′) | Reverse Primer (5′-3′) | Fragment Size (bp) |
|---|---|---|---|
| DuCV | accggcgcttgtactccgtactcc | aataatataacggcgcttgtgcggt | 1995 |
| DHAV-3-VP1 | ggtgattccaatcagcttggcga | ttcaatttctagatggagctcaaag | 720 |
| ARV-σC | atggcgggtctcaatccatcgca | ttaggtgtcgatgccggtacgcacg | 981 |
Table 3.
The prevalence of DHBV, DuCV, DHAV-3, and ARV.
| Duck Breed | Number of Samples | Positive Sample (%) | |||
|---|---|---|---|---|---|
| DHBV | DuCV | DHAV-3 | ARV | ||
| Jinding | 152 | 63.15 (96/152) | 53.95 (82/152) | 8.55 (13/152) | 18.42 (28/152) |
| Shaoxing | 119 | 78.15 (93/119) | 21.01 (25/119) | 8.40 (10/119) | 21.85 (26/119) |
| Overall | 271 | 69.74 (189/271) | 39.48 (107/271) | 8.49 (23/271) | 19.92 (54/271) |
Table 4.
Coinfection rates of DuCV, DHAV-3, DHBV, and ARV.
| Virus | Coinfection Rate (%) | Virus | Coinfection Rate (%) |
|---|---|---|---|
| DHBV and DuCV | 22.75 | DHBV and ARV | 10.20 |
| DHBV and DHAV-3 | 5.49 | DHBV, DHAV-3 and ARV | 0.78 |
| DuCV and ARV | 10.98 | DHBV, DuCV and DHAV-3 | 1.96 |
| DuCV and DHAV-3 | 3.92 | DuCV, DHAV-3 and ARV | 0.78 |
| DHAV-3 and ARV | 1.96 | DHBV, DuCV, DHAV-3 and ARV | 0.78 |
Table 5.
Homology comparison of the whole genome of the identified DuCV with reference strains.
| No | Strain | Year | Region | Accession No. | Cap Gene Sequence Identity (%) | Rep Gene Sequence Identity (%) | Full Genome Identity (%) | ||
|---|---|---|---|---|---|---|---|---|---|
| nt | aa | nt | aa | nt | |||||
| 1 | DU095 | 2010 | Beijing/CN | HM162349.1 | 94.1% | 96.5% | 98.2% | 97.6% | 96.2% |
| 2 | TAYB01 | 2018 | Shandong/CN | MH444647.1 | 93.9% | 96.5% | 98.4% | 96.9% | 96.4% |
| 3 | D12-KD-001 | 2013 | Korea | KC851817.1 | 94.4% | 96.9% | 98.2% | 98.0% | 96.4% |
| 4 | AH01 | 2020 | Henan/CN | MN928808.1 | 94.4% | 96.5% | 99.1% | 97.6% | 97.3% |
| 5 | SDFC12 | 2016 | Shandong/CN | KY328304.1 | 94.2% | 96.5% | 98.5% | 97.3% | 97.1% |
| 6 | VC4 | 2020 | Brazil | MT318126.1 | 93.5% | 96.5% | 98.2% | 97.3% | 96.4% |
| 7 | - | 2005 | USA | NC007220.1 | 93.5% | 96.5% | 98.1% | 96.9% | 96.2% |
| 8 | 33753-52 | 2005 | USA | DQ100076.1 | 93.5% | 96.5% | 98.1% | 96.9% | 96.2% |
| 9 | D11-JW-008 | 2012 | Korea | JQ740363.1 | 94.7% | 96.9% | 98.2% | 97.6% | 97.0% |
| 10 | HN02 | 2020 | Henan/CN | MN928795.1 | 94.4% | 96.5% | 99.2% | 99.0% | 97.4% |
| 11 | NN11/2012 | 2013 | Guangxi/CN | KC460530.1 | 96.5% | 96.1% | 99.0% | 99.0% | 98.0% |
| 12 | DB41-18 | 2014 | Taiwan/CN | KP229368.1 | 96.4% | 97.3% | 98.3% | 98.0% | 97.4% |
| 13 | WF0803 | 2009 | Shandong/CN | GU131342.1 | 97.2% | 97.7% | 99.2% | 99.0% | 97.8% |
| 14 | WF0701 | 2007 | Shandong/CN | EU022375.1 | 96.9% | 97.3% | 98.9% | 99.0% | 97.6% |
| 15 | ShijiazhuangNo. 3 | 2020 | Hebei/CN | MW255979.1 | 97.2% | 97.3% | 98.8% | 98.6% | 97.9% |
| 16 | LA-18 | 2020 | Hebei/CN | MT799793.1 | 96.6% | 97.3% | 98.5% | 98.6% | 97.6% |
| 17 | D12-MR-020 | 2013 | Korea | KC851822.1 | 97.3% | 98.8% | 98.8% | 99.0% | 98.0% |
| 18 | SDHZ1223 | 2020 | Shandong/CN | MT084134.1 | 99.9% | 100.0% | 99.9% | 99.7% | 99.7% |
| 19 | BaodingNO.Du | 2020 | Hebei/CN | MW255980.1 | 99.7% | 100.0% | 99.7% | 99.7% | 99.7% |
| 20 | HZ170301 | 2019 | Shandong/CN | MN068356.1 | 99.9% | 100.0% | 99.8% | 99.7% | 99.8% |
| 21 | QD | 2021 | QingDao/CN | MZ425925.1 | 99.6% | 99.6% | 99.3% | 98.6% | 99.5% |
| 22 | WF190501 | 2019 | Shandong/CN | MN068359.1 | 99.9% | 100.0% | 99.3% | 99.0% | 99.4% |
| 23 | GD/ZQ/144 | 2022 | Guangdong/CN | ON227548.1 | 100.0% | 100.0% | 99.9% | 99.7% | 99.8% |
| 24 | GD/QY/145 | 2022 | Guangdong/CN | ON227549.1 | 100.0% | 100.0% | 99.5% | 99.3% | 99.7% |
| 25 | GD/FS/125 | 2022 | Guangdong/CN | ON227538.1 | 99.6% | 99.6% | 99.2% | 98.3% | 99.4% |
| 26 | GD/FS/132 | 2022 | Guangdong/CN | ON227544.1 | 99.5% | 98.8% | 99.5% | 98.6% | 99.6% |
| 27 | GD/JM/127 | 2022 | Guangdong/CN | ON227540.1 | 99.4% | 100.0% | 97.0% | 95.9% | 97.6% |
| 28 | JSPX03B | 2017 | Shandong/CN | MF627687.1 | 92.2% | 97.3% | 97.6% | 97.6% | 95.5% |
| 29 | QZ36/2012 | 2013 | Guangxi/CN | KC460534.1 | 91.0% | 97.7% | 96.7% | 95.9% | 94.3% |
| 30 | SDDY0520 | 2017 | Shandong/CN | MF627689.1 | 93.2% | 96.9% | 96.8% | 96.9% | 95.1% |
| 31 | FJZZ302 | 2009 | Fujian/CN | GQ423747.1 | 92.2% | 96.9% | 97.8% | 98.0% | 95.2% |
| 32 | CD13056 | 2014 | Taiwan/CN | KP229375.1 | 90.2% | 96.5% | 97.4% | 98.0% | 94.3% |
| 33 | DB7-17 | 2014 | Taiwan/CN | KP229364.1 | 89.5% | 95.7% | 97.4% | 98.0% | 94.0% |
| 34 | MH02/07 | 2008 | Fujian/CN | EU499309.1 | 79.1% | 89.1% | 92.4% | 85.7% | 84.8% |
| 35 | HZ09 | 2007 | Fujian/CN | EU344802.1 | 78.9% | 89.1% | 92.3% | 85.7% | 85.2% |
| 36 | GD190403 | 2019 | Guangdong/CN | MK814577.1 | 78.9% | 88.4% | 88.2% | 82.9% | 82.9% |
| 37 | 31-1 | 2013 | Guangdong/CN | KF941312.1 | 79.1% | 88.4% | 88.2% | 82.9% | 82.9% |
| 38 | LY0701 | 2007 | Shandong/CN | EU022374.1 | 78.7% | 87.6% | 87.8% | 81.2% | 82.7% |
| 39 | CD0912 | 2021 | Chengdu/CN | OK094646.1 | 79.1% | 88.4% | 87.8% | 83.3% | 82.8% |
| 40 | YB02 | 2021 | Chengdu/CN | OK094645.1 | 79.1% | 88.4% | 87.5% | 82.3% | 82.7% |
| 41 | FJ1815 | 2019 | Fujian/CN | MN052853.1 | 78.9% | 88.0% | 87.5% | 81.6% | 82.7% |
| 42 | YN180506 | 2019 | Guangdong/CN | MK814585.1 | 78.4% | 87.2% | 87.6% | 82.3% | 82.4% |
| 43 | PT07 | 2008 | Fujian/CN | EU499310.1 | 79.1% | 88.4% | 87.5% | 80.9% | 82.8% |
| 44 | AHAU28 | 2020 | Anhui/CN | MT646348.1 | 78.8% | 88.0% | 87.6% | 81.6% | 82.7% |
| 45 | GX1104 | 2012 | Guangxi/CN | JX241046.1 | 78.9% | 88.0% | 87.5% | 81.9% | 82.6% |
| 46 | GX1105 | 2012 | Guangxi/CN | JX241045.1 | 78.9% | 88.0% | 87.7% | 82.3% | 82.7% |
Table 6.
Complete genome sequence details of isolated DuCV strains.
| Strain | Nucleotide and Deduced Amino Acid Lengths of Region in Isolated DuCVs | Identities of Nucleotide and Deduced Amino Acid (%) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Cap nt (aa) | Rep nt (aa) | Full Genome (nt) | DB2022-1 | DB2022-2 | DB2022-3 | DB2022-4 | |||||
| Cap nt (aa) | Rep nt (aa) | Cap nt (aa) | Rep nt (aa) | Cap nt (aa) | Rep nt (aa) | Cap nt (aa) | Rep nt (aa) | ||||
| DB2022-1 | 774 (257) | 879 (292) | 1993 | - | - | ||||||
| DB2022-2 | 774 (257) | 879 (292) | 1993 | 99.7 (100) | 100 (100) | ||||||
| DB2022-3 | 774 (257) | 879 (292) | 1993 | 99.7 (100) | 100 (100) | 100 (100) | 100 (100) | ||||
| DB2022-4 | 774 (257) | 879 (292) | 1994 | 99.7 (100) | 100 (100) | 100 (100) | 100 (100) | 100 (100) | 100 (100) | - | - |
Table 7.
Homology comparison of the VP1 gene of the identified DHAV with reference strains.
| No | Strain | Year | Region | Accession No. | Genotype | Identities of Full Genome Sequence (%) | |
|---|---|---|---|---|---|---|---|
| nt | aa | ||||||
| 1 | VF-10 | 2012 | Shandong/CN | KC191683.1 | DHAV-3 | 98.2% | 99.6% |
| 2 | YT-57 | 2012 | Shandong/CN | KC191691.1 | DHAV3 | 98.6% | 100.0% |
| 3 | YA140902 | 2016 | Sichuan/CN | KX523288.1 | DHAV3 | 98.3% | 100.0% |
| 4 | ZP | 2014 | Shandong/CN | KM267028.1 | DHAV3 | 98.5% | 100.0% |
| 5 | GY140703 | 2016 | Sichuan/CN | KX523285.1 | DHAV3 | 98.2% | 99.6% |
| 6 | SD151227 | 2016 | Sichuan/CN | KX523287.1 | DHAV3 | 98.1% | 100.0% |
| 7 | A/dk/CHN/SD08/2019 | 2020 | Shandong/CN | MN912703.1 | DHAV3 | 97.9% | 99.6% |
| 8 | JS | 2020 | Heilongjiang/CN | MN164467.1 | DHAV3 | 99.7% | 100.0% |
| 9 | QL150713 | 2016 | Sichuan/CN | KX523286.1 | DHAV3 | 98.3% | 100.0% |
| 10 | YA151006 | 2016 | Sichuan/CN | KX523289.1 | DHAV3 | 98.5% | 100.0% |
| 11 | JN-12 | 2012 | Shandong/CN | KC191689.1 | DHAV3 | 97.2% | 99.6% |
| 12 | C-LX | 2009 | Beijing/CN | FJ626667.1 | DHAV3 | 97.4% | 99.2% |
| 13 | VF-40 | 2012 | Shandong/CN | KC191686.1 | DHAV3 | 96.7% | 98.3% |
| 14 | C-PSY | 2009 | Beijing/CN | FJ626670.1 | DHAV3 | 96.8% | 98.3% |
| 15 | C-YCZ | 2009 | Beijing/CN | FJ626672.1 | DHAV3 | 96.5% | 97.9% |
| 16 | YT-63 | 2012 | Shandong/CN | KC191693.1 | DHAV3 | 94.0% | 95.4% |
| 17 | BM | 2011 | Vietnam | JF925119.1 | DHAV3 | 95.8% | 97.5% |
| 18 | C-YZC | 2009 | Beijing/CN | FJ626673.1 | DHAV3 | 95.7% | 97.1% |
| 19 | C-PJK | 2012 | Sichuan/CN | KC282430.1 | DHAV3 | 94.0% | 96.2% |
| 20 | HY | 2014 | Vietnam | KM361878.1 | DHAV3 | 95.1% | 97.5% |
| 21 | GD1 | 2007 | Jiangsu/CN | EU289393.1 | DHAV3 | 94.7% | 96.7% |
| 22 | YT-BX | 2012 | Shandong/CN | KC191694.1 | DHAV3 | 94.2% | 95.4% |
| 23 | NT | 2014 | Vietnam | KM361883.1 | DHAV3 | 94.2% | 95.8% |
| 24 | NC | 2011 | Vietnam | JF925121.1 | DHAV3 | 91.0% | 92.1% |
| 25 | DN2 | 2014 | Vietnam | KM361877.1 | DHAV3 | 91.0% | 92.1% |
| 26 | KHO2 | 2014 | Vietnam | KM361880.1 | DHAV3 | 90.6% | 90.8% |
| 27 | LA1 | 2014 | Vietnam | KM361881.1 | DHAV3 | 90.6% | 90.4% |
| 28 | SY1 | 2007 | Zhejiang/CN | EF407857.1 | DHAV1 | 70.0% | 76.7% |
| 29 | ZI07-3 | 2007 | Zhejiang/CN | EF502170.1 | DHAV1 | 69.9% | 75.8% |
| 30 | SY2 | 2007 | Zhejiang/CN | EF407858.1 | DHAV1 | 70.2% | 76.7% |
| 31 | ZJ0812 | 2011 | Shanghai/CN | JF437541.1 | DHAV1 | 69.6% | 75.8% |
| 32 | SY6 | 2007 | Zhejiang/CN | EF407862.1 | DHAV1 | 70.4% | 76.7% |
| 33 | C-GA | 2008 | Beijing/CN | EU687759.1 | DHAV1 | 70.2% | 75.8% |
| 34 | C-TX | 2008 | Beijing/CN | EU687760.1 | DHAV1 | 70.0% | 75.8% |
| 35 | C-PHG | 2009 | Beijing/CN | FJ626668.1 | DHAV1 | 70.4% | 75.8% |
| 36 | C-QDY | 2009 | Beijing/CN | FJ626671.1 | DHAV1 | 70.7% | 75.8% |
| 37 | C-YXQ | 2008 | Beijing/CN | EU687762.1 | DHAV1 | 70.6% | 75.8% |
| 38 | JX140 | 2008 | Beijing/CN | KT072750.1 | DHAV1 | 71.1% | 76.7% |
| 39 | GD06 | 2018 | Fujian/CN | MH778934.1 | DHAV1 | 71.0%% | 76.7% |
| 40 | CZ141017 | 2016 | Sichuan/CN | KX523281.1 | DHAV1 | 70.4% | 76.7% |
| 41 | DY140903 | 2016 | Sichuan/CN | KX523282.1 | DHAV1 | 71.0% | 76.2% |
| 42 | C-CQ3 | 2008 | Beijing/CN | EU687761.1 | DHAV1 | 70.4% | 76.7% |
| 43 | LY0801-P30 | 2014 | Shandong/CN | KJ524552.1 | DHAV1 | 69.0% | 77.5% |
| 44 | MY | 2009 | Sichuan/CN | GU363950.1 | DHAV1 | 71.3% | 77.1% |
| 45 | LY1903 | 2019 | Shandong/CN | MN319537.1 | DHAV1 | 70.2% | 75.4% |
| 46 | SQ1903 | 2019 | Shandong/CN | MN319539.1 | DHAV1 | 70.6% | 75.8% |
| 47 | SY0812 | 2011 | Shanghai/CN | JF437538.1 | DHAV1 | 70.0% | 75.0% |
| 48 | LY0801-CP20 | 2014 | Shandong/CN | KJ606043.1 | DHAV1 | 70.3% | 75.4% |
| 49 | LY0801-CP30 | 2014 | Shandong/CN | KJ606045.1 | DHAV1 | 70.3% | 75.4% |
Table 8.
Homology comparison of the σC gene of the isolated ARV with reference strains.
| No | Strain | Year | Region | Accession No. | Genotype | Whole-Genome Sequence Identity (%) | |
|---|---|---|---|---|---|---|---|
| nt | aa | ||||||
| 1 | HB06 | 2008 | Beijing/CN | EU526387.1 | I | 98.3% | 96.3% |
| 2 | XJ-2011-0315 | 2013 | Heilongjiang/CN | KC963045.1 | I | 98.5% | 97.2% |
| 3 | JR1 | 2006 | USA | EF122836.1 | I | 98.8% | 97.9% |
| 4 | LN09-1 | 2014 | Beijing/CN | KP288843.1 | I | 98.8% | 97.9% |
| 5 | HB10-1 | 2014 | Beijing/CN | KP288833.1 | I | 98.8% | 97.6% |
| 6 | BJ10-1 | 2014 | Beijing/CN | KP288867.1 | I | 98.3% | 97.9% |
| 7 | S113 | 1995 | Canada | L39002.1 | I | 98.2% | 96.3% |
| 8 | 601SI | 1999 | Taiwan/CN | AF204947.1 | I | 98.0% | 96.3% |
| 9 | 1733 | 1997 | USA | AF004857.1 | I | 99.4% | 98.8% |
| 10 | GuangxiR1 | 2012 | Guangxi/CN | KC183744.1 | I | 99.3% | 98.5% |
| 11 | GuangxiR2 | 2013 | Guangxi/CN | KF741732.1 | I | 99.3% | 98.5% |
| 12 | Fahey-Crawley | 2006 | Brazil | DQ868789.1 | I | 99.5% | 99.1% |
| 13 | 176 | 1999 | Canada | AF218358.1 | I | 99.5% | 99.1% |
| 14 | G-98 | 2006 | Huhhot/CN | DQ643975.1 | I | 99.7% | 99.7% |
| 15 | 2012-0129 | 2013 | Heilongjiang/CN | KC963052.1 | I | 99.7% | 99.7% |
| 16 | ARV-HeB01 | 2016 | Heilongjiang/CN | KX451231.1 | I | 99.7% | 99.7% |
| 17 | ARV-XY01 | 2016 | Heilongjiang/CN | KX451229.1 | I | 99.5% | 99.1% |
| 18 | SD-2010-0085 | 2013 | Heilongjiang/CN | KC963039.1 | I | 99.4% | 98.8% |
| 19 | LN05 | 2016 | Heilongjiang/CN | KX451225.1 | I | 99.7% | 99.7% |
| 20 | C-98 | 2006 | Huhhot/CN | EF057397.1 | I | 99.6% | 99.4% |
| 21 | T-98 | 2006 | Huhhot/CN | EF057398.1 | I | 99.5% | 99.4% |
| 22 | ARV-LN01 | 2016 | Heilongjiang/CN | KX451221.1 | I | 99.6% | 99.4% |
| 23 | ARV-LN06 | 2016 | Heilongjiang/CN | KX451226.1 | I | 99.6% | 99.4% |
| 24 | B-98 | 2006 | Huhhot/CN | DQ643974.1 | I | 99.5% | 99.1% |
| 25 | VA-1 | 2008 | India | EU681254.1 | I | 98.9% | 97.2% |
| 26 | GEL12 98M | 2001 | Netherlands | AF354225.1 | I | 77.3% | 77.6% |
| 27 | Reo/Broiler/YTLY/161024a | 2018 | Shandong/CN | MK189468.1 | I | 73.7% | 73.9% |
| 28 | 916 | 2000 | Taiwan/CN | AF297214.1 | II | 61.2% | 58.1% |
| 29 | ARV/Crow/Kagawa/P3/2012 | 2016 | Japan | LC164026.1 | II | 61.0% | 56.6% |
| 30 | GEL13a98M | 2001 | Netherlands | AF354226.1 | II | 51.3% | 54.4% |
| 31 | NC/98 | 2006 | USA | DQ995806.1 | II | 61.0% | 55.7% |
| 32 | TARV-MN3 | 2013 | USA | KF872234.1 | II | 53.3% | 56.7% |
| 33 | TARV-ONEIL | 2013 | USA | KF872231.1 | II | 52.0% | 56.2% |
| 34 | TARV-Crestview | 2013 | USA | KF872238.1 | II | 52.5% | 55.5% |
| 35 | T1781 | 2013 | Hungary | KC865792.1 | III | 59.7% | 50.8% |
| 36 | SD18 | 2020 | Shandong/CN | MT747423.1 | III | 59.3% | 50.5% |
| 37 | 42563-4 | 2006 | USA | DQ872801.1 | III | 58.9% | 54.7% |
| 38 | GEL13b98M | 2001 | Netherlands | AF354227.1 | III | 58.1% | 53.5% |
| 39 | D6 | 2019 | Canada | MN879650.1 | IV | 45.6% | 48.9% |
| 40 | D12 | 2019 | Canada | MN879710.1 | IV | 55.5% | 49.8% |
| 41 | Reo/USA/Broiler/1057NY/18 | 2021 | USA | MW854823.1 | IV | 56.2% | 49.5% |
| 42 | AVS-B | 2010 | Hungary | FR694197.1 | IV | 54.8% | 48.6% |
| 43 | K1600657 | 2019 | USA | MK583337.1 | IV | 54.7% | 49.5% |
| 44 | NLI12 96M | 2001 | Netherlands | AF354230.1 | IV | 54.1% | 50.2% |
| 45 | 918 | 2000 | Taiwan/CN | AF297215.1 | V | 57.0% | 48.0% |
| 46 | 1017-1 | 2000 | Taiwan/CN | AF297216.1 | V | 56.5% | 48.3% |
| 47 | NGN20 7-1 2b | 2021 | Japan | LC604650.1 | V | 55.9% | 47.0% |
| 48 | Reo/PA/Broiler/19981/13 | 2015 | USA | KR856993.1 | VI | 55.5% | 49.3% |
| 49 | Reo/PA/Broiler/03476/12 | 2015 | USA | KP727784.1 | VI | 55.8% | 48.0% |
| 50 | Reo/PA/Broiler/03200/12 | 2015 | USA | KP727785.1 | VI | 55.8% | 48.0% |
Table 9.
Physicochemical properties analysis of DuCV-Cap, DHAV-3-VP1, and ARV-σC.
| Sequence | pI | Instability Index | Aliphatic Index | GRAVY |
|---|---|---|---|---|
| DuCV-KC851817 (2013) | 10.74 | 58.9 | 50.47 | −0.748 |
| DuCV-OQ658501.1 (2022) | 10.61 | 60.94 | 50.08 | −0.765 |
| DHAV-3-FJ626673 (2009) | 6.62 | 51.32 | 83.67 | −0.149 |
| DHAV-3-OQ658504.1 (2022) | 7.29 | 52.39 | 82.04 | −0.163 |
| ARV-EF122836 (2006) | 5.0 | 41.68 | 89.72 | −0.057 |
| ARV-OR046324.1 (2022) | 4.81 | 36.12 | 89.11 | −0.036 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, X.; Yu, H.; Zhang, W.; Fu, L.; Wang, Y. Molecular Detection and Genetic Characterization of Vertically Transmitted Viruses in Ducks. Animals 2024, 14, 6. https://doi.org/10.3390/ani14010006
AMA Style
Wang X, Yu H, Zhang W, Fu L, Wang Y. Molecular Detection and Genetic Characterization of Vertically Transmitted Viruses in Ducks. Animals. 2024; 14(1):6. https://doi.org/10.3390/ani14010006
Chicago/Turabian StyleWang, Xinrong, Haidong Yu, Wenli Zhang, Lizhi Fu, and Yue Wang. 2024. "Molecular Detection and Genetic Characterization of Vertically Transmitted Viruses in Ducks" Animals 14, no. 1: 6. https://doi.org/10.3390/ani14010006
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.