
Identification of SWI/SNF Subcomplex GBAF Presence, Intra-Complex Interactions, and Transcriptional Dynamics during Early Porcine Development
 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Oocyte Collection and In Vitro Embryo Culture
2.2. Cell Lines and Culture Conditions
2.3. Nuclear Lysate Extraction
2.4. Antibodies
2.5. Immunoprecipitation
2.6. Immunoblot Analysis
2.7. Immunocytochemistry
2.8. RNA-Seq Analysis
2.9. Statistical Analysis
3. Results
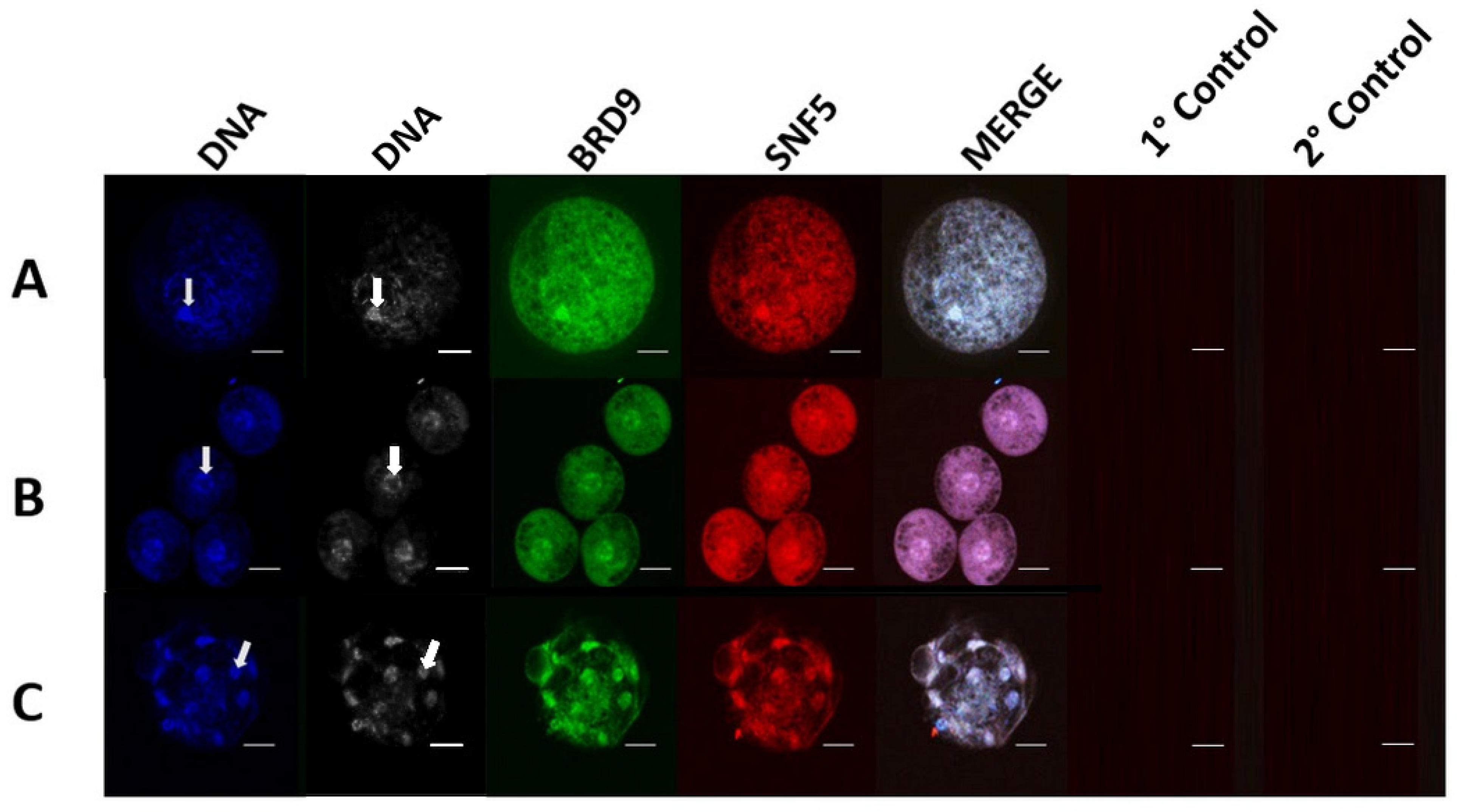
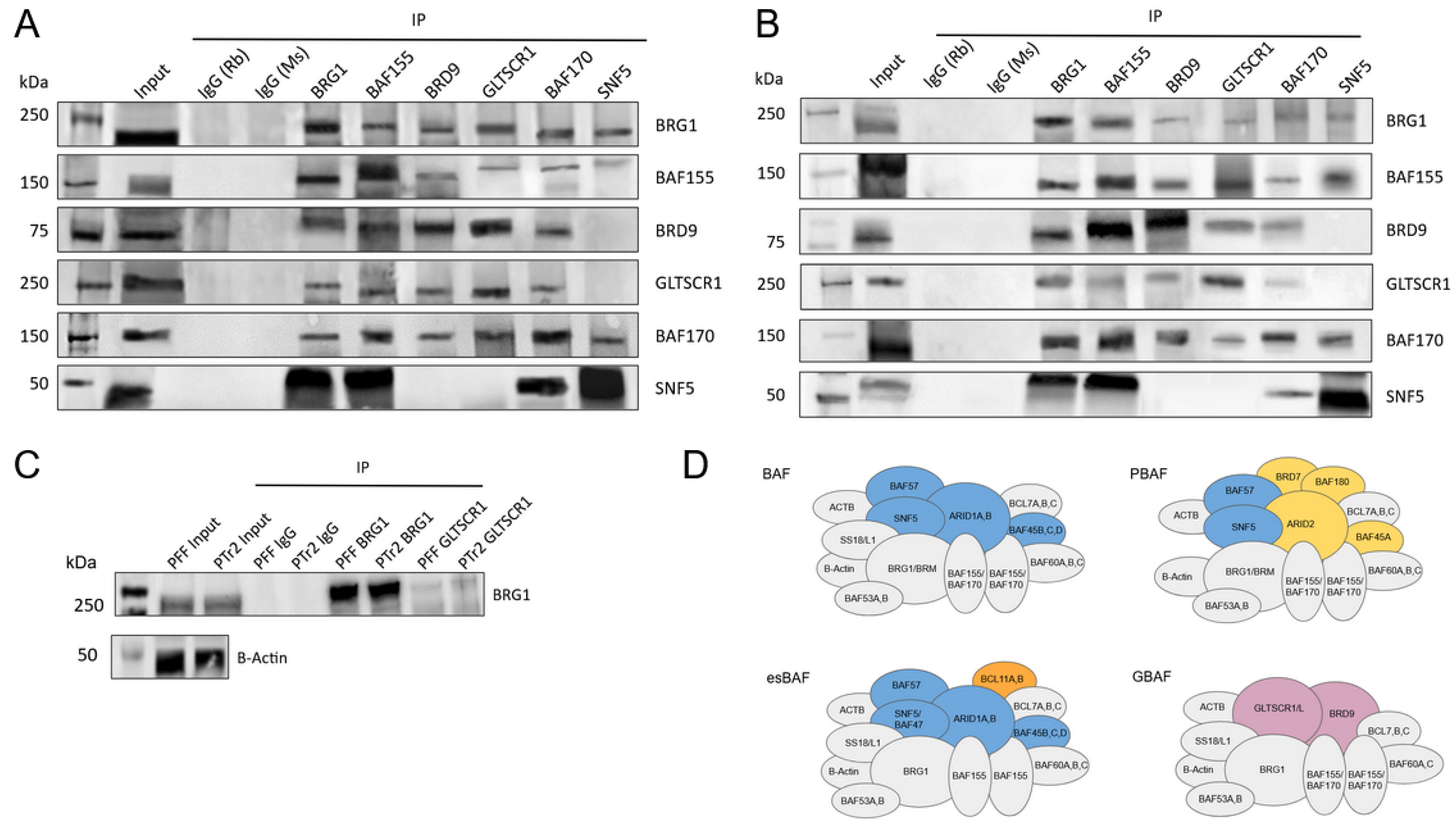
3.1. GBAF Characteristic Subunits BRD9 and GLTSCR1 Are Present in Porcine Oocytes, Embryos, and Cell Lines
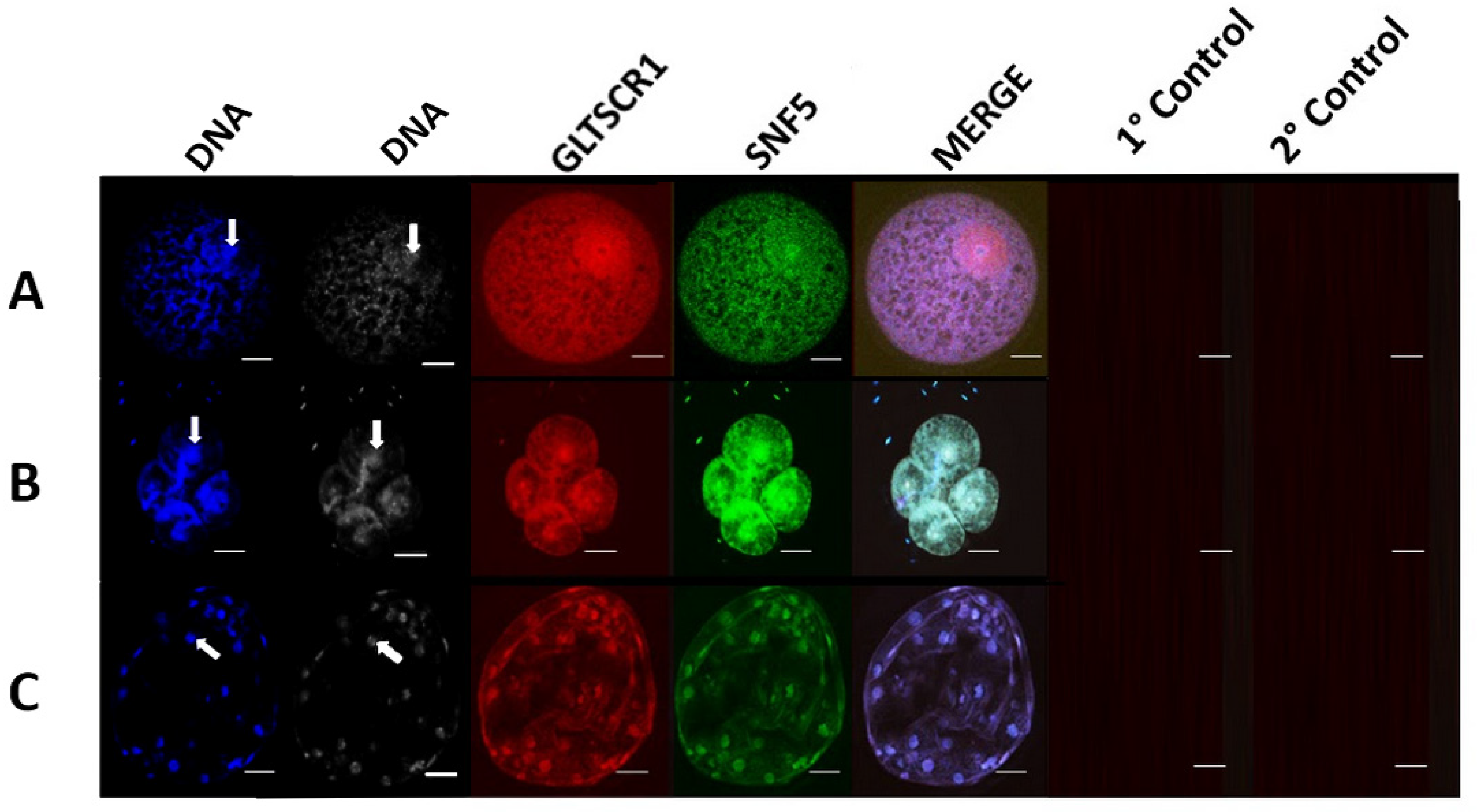
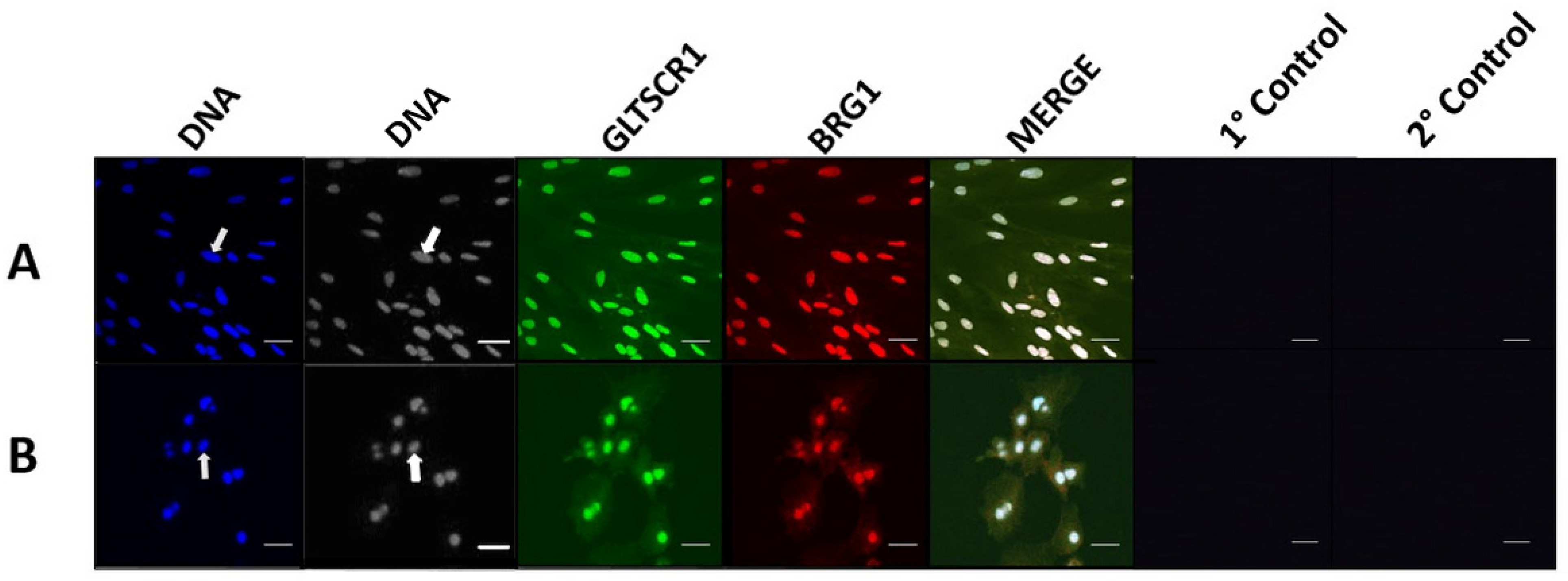
3.2. GLTSCR1 Is Present in Porcine Pre-Implantation and Early Development Cell Lines and Interacts with BAF170
3.3. GBAF Subunit Expression Varies across Developmental Stages
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gluckman, P.D.; Hanson, M.A.; Low, F.M. The Role of Developmental Plasticity and Epigenetics in Human Health. Birth Defects Res. C Embryo Today 2011, 93, 12–18. [Google Scholar] [CrossRef]
- Tiffon, C. The Impact of Nutrition and Environmental Epigenetics on Human Health and Disease. Int. J. Mol. Sci. 2018, 19, 3425. [Google Scholar] [CrossRef]
- Stover, P.J.; James, W.P.T.; Krook, A.; Garza, C. Emerging Concepts on the Role of Epigenetics in the Relationships between Nutrition and Health. J. Intern. Med. 2018, 284, 37–49. [Google Scholar] [CrossRef]
- Hargreaves, D.C.; Crabtree, G.R. ATP-Dependent Chromatin Remodeling: Genetics, Genomics and Mechanisms. Cell Res. 2011, 21, 396–420. [Google Scholar] [CrossRef]
- Alfert, A.; Moreno, N.; Kerl, K. The BAF Complex in Development and Disease. Epigenetics Chromatin 2019, 12, 19. [Google Scholar] [CrossRef]
- Marmorstein, R.; Trievel, R.C. Histone Modifying Enzymes: Structures, Mechanisms, and Specificities. Biochim. Biophys. Acta 2009, 1789, 58–68. [Google Scholar] [CrossRef]
- Rippe, K.; Schrader, A.; Riede, P.; Strohner, R.; Lehmann, E.; Längst, G. DNA Sequence- and Conformation-Directed Positioning of Nucleosomes by Chromatin-Remodeling Complexes. Proc. Natl. Acad. Sci. USA 2007, 104, 15635–15640. [Google Scholar] [CrossRef]
- Tyagi, M.; Imam, N.; Verma, K.; Patel, A.K. Chromatin Remodelers: We Are the Drivers!! Nucleus 2016, 7, 388–404. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Côté, J.; Xue, Y.; Zhou, S.; Khavari, P.A.; Biggar, S.R.; Muchardt, C.; Kalpana, G.V.; Goff, S.P.; Yaniv, M.; et al. Purification and Biochemical Heterogeneity of the Mammalian SWI-SNF Complex. EMBO J. 1996, 15, 5370–5382. [Google Scholar] [CrossRef] [PubMed]
- Phelan, M.L.; Sif, S.; Narlikar, G.J.; Kingston, R.E. Reconstitution of a Core Chromatin Remodeling Complex from SWI/SNF Subunits. Mol. Cell 1999, 3, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Alpsoy, A.; Dykhuizen, E.C. Glioma Tumor Suppressor Candidate Region Gene 1 (GLTSCR1) and Its Paralog GLTSCR1-like Form SWI/SNF Chromatin Remodeling Subcomplexes. J. Biol. Chem. 2018, 293, 3892–3903. [Google Scholar] [CrossRef]
- Ho, L.; Ronan, J.L.; Wu, J.; Staahl, B.T.; Chen, L.; Kuo, A.; Lessard, J.; Nesvizhskii, A.I.; Ranish, J.; Crabtree, G.R. An Embryonic Stem Cell Chromatin Remodeling Complex, EsBAF, Is Essential for Embryonic Stem Cell Self-Renewal and Pluripotency. Proc. Natl. Acad. Sci. USA 2009, 106, 5181–5186. [Google Scholar] [CrossRef]
- King, H.W.; Klose, R.J. The Pioneer Factor OCT4 Requires the Chromatin Remodeller BRG1 to Support Gene Regulatory Element Function in Mouse Embryonic Stem Cells. eLife 2017, 6, e22631. [Google Scholar] [CrossRef]
- Gatchalian, J.; Malik, S.; Ho, J.; Lee, D.-S.; Kelso, T.W.R.; Shokhirev, M.N.; Dixon, J.R.; Hargreaves, D.C. A Non-Canonical BRD9-Containing BAF Chromatin Remodeling Complex Regulates Naive Pluripotency in Mouse Embryonic Stem Cells. Nat. Commun. 2018, 9, 5139. [Google Scholar] [CrossRef]
- Hohmann, A.F.; Martin, L.J.; Minder, J.L.; Roe, J.-S.; Shi, J.; Steurer, S.; Bader, G.; McConnell, D.; Pearson, M.; Gerstberger, T.; et al. Sensitivity and Engineered Resistance of Myeloid Leukemia Cells to BRD9 Inhibition. Nat. Chem. Biol. 2016, 12, 672–679. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Wang, Y.; Li, Q.; Fei, X.; Ma, H.; Hu, R. MiR-140-3p Functions as a Tumor Suppressor in Squamous Cell Lung Cancer by Regulating BRD9. Cancer Lett. 2019, 446, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, S.; Troisi, E.C.; Howard, T.P.; Haswell, J.R.; Wolf, B.K.; Hawk, W.H.; Ramos, P.; Oberlick, E.M.; Tzvetkov, E.P.; et al. BRD9 Defines a SWI/SNF Sub-Complex and Constitutes a Specific Vulnerability in Malignant Rhabdoid Tumors. Nat. Commun. 2019, 10, 1881. [Google Scholar] [CrossRef] [PubMed]
- Michel, B.C.; D’Avino, A.R.; Cassel, S.H.; Mashtalir, N.; McKenzie, Z.M.; McBride, M.J.; Valencia, A.M.; Zhou, Q.; Bocker, M.; Soares, L.M.M.; et al. A Non-Canonical SWI/SNF Complex Is a Synthetic Lethal Target in Cancers Driven by BAF Complex Perturbation. Nat. Cell Biol. 2018, 20, 1410–1420. [Google Scholar] [CrossRef] [PubMed]
- Brien, G.L.; Remillard, D.; Shi, J.; Hemming, M.L.; Chabon, J.; Wynne, K.; Dillon, E.T.; Cagney, G.; Van Mierlo, G.; Baltissen, M.P.; et al. Targeted Degradation of BRD9 Reverses Oncogenic Gene Expression in Synovial Sarcoma. eLife 2018, 7, e41305. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Kollmeyer, T.M.; Buckner, K.; Bamlet, W.R.; Ballman, K.V.; Jenkins, R.B. Polymorphisms InGLTSCR1 AndERCC2 Are Associated with the Development of Oligodendrogliomas. Cancer 2005, 103, 2363–2372. [Google Scholar] [CrossRef]
- Ma, X.; Du, T.; Zhu, D.; Chen, X.; Lai, Y.; Wu, W.; Wang, Q.; Lin, C.-C.; Li, Z.; Liu, L.; et al. High Levels of Glioma Tumor Suppressor Candidate Region Gene 1 Predicts a Poor Prognosis for Prostate Cancer. Oncol. Lett. 2018, 16, 6749–6755. [Google Scholar] [CrossRef] [PubMed]
- Han, F.; Zhang, L.; Chen, C.; Wang, Y.; Zhang, Y.; Qian, L.; Sun, W.; Zhou, D.; Yang, B.; Zhang, H.; et al. GLTSCR1 Negatively Regulates BRD4-Dependent Transcription Elongation and Inhibits CRC Metastasis. Adv. Sci. 2019, 6, 1901114. [Google Scholar] [CrossRef] [PubMed]
- Reyes, J.C.; Barra, J.; Muchardt, C.; Camus, A.; Babinet, C.; Yaniv, M. Altered Control of Cellular Proliferation in the Absence of Mammalian Brahma (SNF2α). EMBO J. 1998, 17, 6979–6991. [Google Scholar] [CrossRef] [PubMed]
- Bultman, S.J. Maternal BRG1 Regulates Zygotic Genome Activation in the Mouse. Genes Dev. 2006, 20, 1744–1754. [Google Scholar] [CrossRef] [PubMed]
- Bultman, S.; Gebuhr, T.; Yee, D.; La Mantia, C.; Nicholson, J.; Gilliam, A.; Randazzo, F.; Metzger, D.; Chambon, P.; Crabtree, G.; et al. A Brg1 Null Mutation in the Mouse Reveals Functional Differences among Mammalian SWI/SNF Complexes. Mol. Cell 2000, 6, 1287–1295. [Google Scholar] [CrossRef] [PubMed]
- Kaeser, M.D.; Aslanian, A.; Dong, M.-Q.; Yates, J.R.; Emerson, B.M. BRD7, a Novel PBAF-Specific SWI/SNF Subunit, Is Required for Target Gene Activation and Repression in Embryonic Stem Cells. J. Biol. Chem. 2008, 283, 32254–32263. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Tate, P.; Hu, P.; Tjian, R.; Skarnes, W.C.; Wang, Z. ES Cell Pluripotency and Germ-Layer Formation Require the SWI/SNF Chromatin Remodeling Component BAF250a. Proc. Natl. Acad. Sci. USA 2008, 105, 6656–6661. [Google Scholar] [CrossRef] [PubMed]
- Magnani, L.; Cabot, R.A. Manipulation of SMARCA2 and SMARCA4 Transcript Levels in Porcine Embryos Differentially Alters Development and Expression of SMARCA1, SOX2, NANOG, and EIF1. Reproduction 2009, 137, 23–33. [Google Scholar] [CrossRef]
- Panamarova, M.; Cox, A.; Zernicka-Goetz, M.; Butler, R.J.; Bulgakova, N.A.; Jeon, S.T.; Rosen, B.; Seong, R.H.; Skarnes, W.C.; Crabtree, R.H.; et al. BAF Chromatin Remodelling Complex Is an Epigenetic Regulator of Lineage Specification in the Early Mouse Embryo. Development 2016, 143, 1271–1283. [Google Scholar] [CrossRef]
- Cabot, B.; Tseng, Y.-C.; Crodian, J.; Cabot, R. Differential Expression of Key Subunits of SWI/SNF Chromatin Remodeling Complexes in Porcine Embryos Derived In Vitro or In Vivo. Mol. Reprod. Dev. 2017, 84, 1238–1249. [Google Scholar] [CrossRef]
- Tseng, Y.-C.; Cabot, B.; Cabot, R. ARID1A, a Component of SWI/SNF Chromatin Remodeling Complexes, Is Required for Porcine Embryo Development. Mol. Reprod. Dev. 2017, 84, 1250–1256. [Google Scholar] [CrossRef] [PubMed]
- Abeydeera, L.R.; Wang, W.; Prather, R.S.; Day, B.N. Maturation in Vitro of Pig Oocytes in Protein-Free Culture Media: Fertilization and Subsequent Embryo Development in Vitro1. Biol. Reprod. 1998, 58, 1316–1320. [Google Scholar] [CrossRef]
- Kühholzer, B.; Hawley, R.J.; Lai, L.; Kolber-Simonds, D.; Prather, R.S. Clonal Lines of Transgenic Fibroblast Cells Derived from the Same Fetus Result in Different Development When Used for Nuclear Transfer in Pigs1. Biol. Reprod. 2001, 64, 1695–1698. [Google Scholar] [CrossRef] [PubMed]
- Ka, H.; Jaeger, L.A.; Johnson, G.A.; Spencer, T.J.; Bazer, F.W. Keratinocyte Growth Factor Is Up-Regulated by Estrogen in the Porcine Uterine Endometrium and Functions in Trophectoderm Cell Proliferation and Differentiation. Endocrinology 2001, 142, 2303–2310. [Google Scholar] [CrossRef] [PubMed]
- Krueger, F. Trim Galore. GitHub. Available online: https://github.com/FelixKrueger/TrimGalore (accessed on 3 December 2023).
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A Fast Spliced Aligner with Low Memory Requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python Framework to Work with High-Throughput Sequencing Data. Bioinformatics 2014, 31, 166–169. [Google Scholar] [CrossRef]
- Smyth, G.K. Limma: Linear Models for Microarray Data. In Bioinformatics and Computational Biology Solutions Using R and Bioconductor; Springer: New York, NY, USA, 2005; pp. 397–420. [Google Scholar] [CrossRef]
- Law, C.W.; Chen, Y.; Shi, W.; Smyth, G.K. Voom: Precision Weights Unlock Linear Model Analysis Tools for RNA-Seq Read Counts. Genome Biol. 2014, 15, R29. [Google Scholar] [CrossRef]
- Liu, R.; Holik, A.Z.; Su, S.; Jansz, N.; Chen, K.; Leong, H.S.; Blewitt, M.E.; Asselin-Labat, M.-L.; Smyth, G.K.; Ritchie, M.E. Why Weight? Modelling Sample and Observational Level Variability Improves Power in RNA-Seq Analyses. Nucleic Acids Res. 2015, 43, e97. [Google Scholar] [CrossRef]
- Robinson, M.D.; Oshlack, A. A Scaling Normalization Method for Differential Expression Analysis of RNA-Seq Data. Genome Biol. 2010, 11, R25. [Google Scholar] [CrossRef]
- Wang, M.; Ibeagha-Awemu, E.M. Impacts of Epigenetic Processes on the Health and Productivity of Livestock. Front. Genet. 2021, 11, 613636. [Google Scholar] [CrossRef]
- Kropp, J.; Carrillo, J.A.; Namous, H.; Daniels, A.; Salih, S.M.; Song, J.; Khatib, H. Male Fertility Status Is Associated with DNA Methylation Signatures in Sperm and Transcriptomic Profiles of Bovine Preimplantation Embryos. BMC Genom. 2017, 18, 280. [Google Scholar] [CrossRef]
- Fang, L.; Zhou, Y.; Liu, S.; Jiang, J.; Bickhart, D.M.; Null, D.J.; Li, B.; Schroeder, S.G.; Rosen, B.D.; Cole, J.B.; et al. Integrating Signals from Sperm Methylome Analysis and Genome-Wide Association Study for a Better Understanding of Male Fertility in Cattle. Epigenomes 2019, 3, 10. [Google Scholar] [CrossRef]
- Doherty, R.; Whiston, R.; Cormican, P.; Finlay, E.K.; Couldrey, C.; Brady, C.; O’Farrelly, C.; Meade, K.G. The CD4+ T Cell Methylome Contributes to a Distinct CD4+ T Cell Transcriptional Signature in Mycobacterium Bovis-Infected Cattle. Sci. Rep. 2016, 6, 31014. [Google Scholar] [CrossRef] [PubMed]
- Ross, J.W.; Hale, B.J.; Gabler, N.K.; Rhoads, R.P.; Keating, A.F.; Baumgard, L.H. Physiological Consequences of Heat Stress in Pigs. Anim. Prod. Sci. 2015, 55, 1381. [Google Scholar] [CrossRef]
- Serviento, A.M.; Lebret, B.; Renaudeau, D. Chronic Prenatal Heat Stress Alters Growth, Carcass Composition, and Physiological Response of Growing Pigs Subjected to Postnatal Heat Stress. J. Anim. Sci. 2020, 98, skaa161. [Google Scholar] [CrossRef] [PubMed]
- Salces-Ortiz, J.; González, C.; Bolado-Carrancio, A.; Rodríguez-Rey, J.C.; Calvo, J.H.; Muñoz, R.; Serrano, M.M. Ovine HSP90AA1 Gene Promoter: Functional Study and Epigenetic Modifications. PLoS ONE 2015, 20, 1001–1012. [Google Scholar] [CrossRef]
- Lu, Z.; Ma, Y.; Li, Q.; Liu, E.; Jin, M.; Zhang, L.; Wei, C. The Role of N6-Methyladenosine RNA Methylation in the Heat Stress Response of Sheep (Ovis aries). Cell Stress Chaperones 2019, 24, 333–342. [Google Scholar] [CrossRef]
- Zhang, F.; Chen, S.; Yang, T.; Ao, H.; Zhai, L.; Li, Q.; Xing, K.; Liu, Y.; Liu, H.; Yu, Y.; et al. Novel DNA Methylation Markers of PRRSV-Specific Antibodies and Their Intergenerational Transmission from Pregnant Sows to Piglets. Gene 2021, 801, 145831. [Google Scholar] [CrossRef]
- Elolimy, A.; Vailati-Riboni, M.; Liang, Y.; Loor, J.J. Cellular Mechanisms and Epigenetic Changes. Vet. Clin. N. Am. Food Anim. Prac. 2019, 35, 249–263. [Google Scholar] [CrossRef] [PubMed]
- Ouellet, V.; Boucher, A.; Dahl, G.E.; Laporta, J. Consequences of Maternal Heat Stress at Different Stages of Embryonic and Fetal Development on Dairy Cows’ Progeny. Anim. Front. 2021, 11, 48–56. [Google Scholar] [CrossRef]
- Costa, T.C.; Gionbelli, M.P.; Duarte, M. de S. Fetal Programming in Ruminant Animals: Understanding the Skeletal Muscle Development to Improve Meat Quality. Anim. Front. 2021, 11, 66–73. [Google Scholar] [CrossRef]
- Clarke, S.; Caulton, A.; McRae, K.; Brauning, R.; Couldrey, C.; Dodds, K. Beyond the Genome: A Perspective on the Use of DNA Methylation Profiles as a Tool for the Livestock Industry. Anim. Front. 2021, 11, 90–94. [Google Scholar] [CrossRef]
- Laporta, J.; Dado-Senn, B.; Skibiel, A.L. Late Gestation Hyperthermia: Epigenetic Programming of Daughter’s Mammary Development and Function. Domest. Anim. Endocrinol. 2022, 78, 106681. [Google Scholar] [CrossRef]
- Chen, J.; Archer, T.K. Regulating SWI/SNF Subunit Levels via Protein-Protein Interactions and Proteasomal Degradation: BAF155 and BAF170 Limit Expression of BAF57. Mol. Cell Biol. 2005, 25, 9016–9027. [Google Scholar] [CrossRef]
- Zhang, X.; Li, B.; Li, W.; Ma, L.; Zheng, D.; Li, L.; Yang, W.; Chu, M.; Chen, W.; Mailman, R.B.; et al. Transcriptional Repression by the BRG1-SWI/SNF Complex Affects the Pluripotency of Human Embryonic Stem Cells. Stem Cell Rep. 2014, 3, 460–474. [Google Scholar] [CrossRef]
- Matalon, O.; Horovitz, A.; Levy, E.D. Different Subunits Belonging to the Same Protein Complex Often Exhibit Discordant Expression Levels and Evolutionary Properties. Curr. Opin. Struct. Biol. 2014, 26, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Chen, Z.; Yin, Q.; Fu, X.; Li, Y.; Stopka, T.; Skoultchi, A.I.; Zhang, Y. The Chromatin Remodeler Snf2h Is Essential for Oocyte Meiotic Cell Cycle Progression. Genes Dev. 2020, 34, 166–178. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Huang, B.; Chen, H.; Yin, Q.; Liu, Y.; Xiang, Y.; Zhang, B.; Liu, B.; Wang, Q.; Xia, W.; et al. The Landscape of Accessible Chromatin in Mammalian Preimplantation Embryos. Nature 2016, 534, 652–657. [Google Scholar] [CrossRef] [PubMed]
- Torres-Padilla, M.E.; Zernicka-Goetz, M. Role of TIF1α as a Modulator of Embryonic Transcription in the Mouse Zygote. J. Cell Biol. 2006, 174, 329–338. [Google Scholar] [CrossRef]
- Cabot, B.; Cabot, R.A. Chromatin Remodeling in Mammalian Embryos. Reproduction 2018, 155, R147–R158. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Kruskal-Wallis Rank-Sum Test | Wilcoxon Rank-Sum Exact Test | ||||||
|---|---|---|---|---|---|---|---|
| Subunit | Mean | SD | Χ2 | df | p-Value | Comparison | padj |
| BRD9 | |||||||
| Oocytes | 2.84 | 1.47 | 0.29 | 2 | 0.863 | Oocytes—4-cells | 0.93 |
| 4-Cell embryos | 2.56 | 1.69 | Oocytes—blastocysts | 0.93 | |||
| Blastocysts | 2.96 | 1.70 | 4-cells-blastocysts | 0.93 | |||
| GLTSCR1 | |||||||
| Oocytes | 0.23 | 1.82 | 2.43 | 2 | 0.297 | Oocytes—4-cells | 0.420 |
| 4-Cell embryos | −0.76 | 1.32 | Oocytes—blastocysts | 0.420 | |||
| Blastocysts | −1.96 | 2.23 | 4-cells-blastocysts | 0.420 | |||
| GLTSCR1L | |||||||
| Oocytes | 1.29 | 1.39 | 6.28 | 2 | 0.044 | Oocytes—4-cells | 0.690 |
| 4-Cell embryos | 0.99 | 0.62 | Oocytes—blastocysts | 0.266 | |||
| Blastocysts | 2.40 | 0.80 | 4-cells-blastocysts | 0.026 | |||
| BRG1 | |||||||
| Oocytes | 3.64 | 0.21 | 12.53 | 2 | 0.002 | Oocytes—4-cells | 0.032 |
| 4-Cell embryos | 2.74 | 0.77 | Oocytes—blastocysts | 0.007 | |||
| Blastocysts | 4.76 | 0.95 | 4-cells-blastocysts | 0.007 | |||
| BAF155 | |||||||
| Oocytes | 5.76 | 0.50 | 5.09 | 2 | 0.079 | Oocytes—4-cells | 0.690 |
| 4-Cell embryos | 5.73 | 1.46 | Oocytes—blastocysts | 0.013 | |||
| Blastocysts | 6.62 | 0.22 | 4-cells-blastocysts | 0.690 | |||
| BAF170 | |||||||
| Oocytes | 4.64 | 0.45 | 7.99 | 2 | 0.018 | Oocytes—4-cells | 0.421 |
| 4-Cell embryos | 4.21 | 0.84 | Oocytes—blastocysts | 0.026 | |||
| Blastocysts | 2.92 | 0.76 | 4-cells-blastocysts | 0.078 | |||
| BAF53A | |||||||
| Oocytes | 4.75 | 0.37 | 2.31 | 2 | 0.315 | Oocytes—4-cells | 0.840 |
| 4-Cell embryos | 4.49 | 1.51 | Oocytes—blastocysts | 0.490 | |||
| Blastocysts | 4.30 | 0.56 | 4-cells-blastocysts | 0.490 | |||
| BAF60A | |||||||
| Oocytes | 4.97 | 1.05 | 5.76 | 2 | 0.056 | Oocytes—4-cells | 0.032 |
| 4-Cell embryos | 3.72 | 1.78 | Oocytes—blastocysts | 0.007 | |||
| Blastocysts | 2.94 | 0.38 | 4-cells-blastocysts | 0.007 | |||
| SS18 | |||||||
| Oocytes | 5.82 | 0.97 | 1.32 | 2 | 0.516 | Oocytes—4-cells | 0.790 |
| 4-Cell embryos | 4.86 | 1.31 | Oocytes—blastocysts | 0.790 | |||
| Blastocysts | 5.46 | 1.23 | 4-cells-blastocysts | 0.790 | |||
| BCL7A | |||||||
| Oocytes | 4.06 | 1.33 | 1.58 | 2 | 0.453 | Oocytes—4-cells | 0.840 |
| 4-Cell embryos | 3.68 | 1.02 | Oocytes—blastocysts | 0.740 | |||
| Blastocysts | 2.85 | 1.54 | 4-cells-blastocysts | 0.810 | |||
| BCL7B | |||||||
| Oocytes | −1.02 | 1.84 | 0.26 | 2 | 0.877 | Oocytes—4-cells | 0.930 |
| 4-Cell embryos | −0.45 | 2.35 | Oocytes—blastocysts | 0.930 | |||
| Blastocysts | −0.51 | 1.64 | 4-cells-blastocysts | 0.930 | |||
| BCL7C | |||||||
| Oocytes | 1.57 | 1.15 | 3.51 | 2 | 0.173 | Oocytes—4-cells | 0.330 |
| 4-Cell embryos | −0.39 | 2.08 | Oocytes—blastocysts | 0.790 | |||
| Blastocysts | 1.35 | 0.80 | 4-cells-blastocysts | 0.250 | |||
| Comparison (PTr2-PFF) | MeanPTr2 | SDPTr2 | MeanPFF | SDPFF | t | df | 95% Confidence Interval | p-Value | |
|---|---|---|---|---|---|---|---|---|---|
| Lower | Upper | ||||||||
| BRD9 | 5.29 | 0.02 | 6.15 | 0.02 | −52.74 | 3.65 | −0.91 | −0.81 | 2.168 × 10−6 |
| GLTSCR1 | 1.41 | 0.03 | 4.42 | 0.11 | −45.61 | 2.35 | −3.25 | −2.76 | 1.621 × 10−4 |
| GLTSCR1L | 2.49 | 0.33 | 3.74 | 0.06 | −6.47 | 2.12 | −2.04 | −0.46 | 0.02 |
| BRG1 | 5.42 | 0.09 | 7.78 | 0.01 | −45.80 | 2.01 | −2.59 | −2.14 | 4.575 × 10−4 |
| BAF155 | 5.32 | 0.06 | 6.93 | 0.04 | −39.99 | 3.74 | −1.73 | −1.50 | 4.609 × 10−6 |
| BAF170 | 4.40 | 0.09 | 6.98 | 0.02 | −50.17 | 2.20 | −2.78 | −2.38 | 2.017 × 10−4 |
| BAF60A | 2.08 | 0.20 | 5.61 | 0.05 | −21.21 | 2.21 | −3.01 | −2.06 | 0.001 |
| BAF53A | 3.18 | 0.07 | 4.97 | 0.09 | −27.77 | 3.84 | −1.97 | −1.61 | 1.425 × 10−5 |
| SS18 | 6.53 | 0.04 | 6.24 | 0.05 | 7.52 | 3.98 | 0.18 | 0.38 | 0.002 |
| BCL7A | 3.96 | 0.08 | 3.99 | 0.06 | −0.44 | 3.63 | −0.20 | 0.15 | 0.682 |
| BCL7B | −0.52 | 0.87 | 2.61 | 0.08 | −6.19 | 2.04 | −5.26 | −0.99 | 0.024 |
| BCL7C | 5.16 | 0.06 | 4.40 | 0.20 | 6.27 | 2.31 | 0.30 | 1.23 | 0.017 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Innis, S.; Alpsoy, A.; Crodian, J.; Tseng, Y.-C.; Dykhuizen, E.; Cabot, B.; Cabot, R. Identification of SWI/SNF Subcomplex GBAF Presence, Intra-Complex Interactions, and Transcriptional Dynamics during Early Porcine Development. Animals 2024, 14, 773. https://doi.org/10.3390/ani14050773
Innis S, Alpsoy A, Crodian J, Tseng Y-C, Dykhuizen E, Cabot B, Cabot R. Identification of SWI/SNF Subcomplex GBAF Presence, Intra-Complex Interactions, and Transcriptional Dynamics during Early Porcine Development. Animals. 2024; 14(5):773. https://doi.org/10.3390/ani14050773
Chicago/Turabian StyleInnis, Sarah, Aktan Alpsoy, Jennifer Crodian, Yu-Chun Tseng, Emily Dykhuizen, Birgit Cabot, and Ryan Cabot. 2024. "Identification of SWI/SNF Subcomplex GBAF Presence, Intra-Complex Interactions, and Transcriptional Dynamics during Early Porcine Development" Animals 14, no. 5: 773. https://doi.org/10.3390/ani14050773