


Effect of Melatonin on Chemoresistance Exhibited by Spheres Derived from Canine Mammary Carcinoma Cells
 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Methods
2.1. Cell Culture
2.2. Flow Cytometry
2.3. Cell Viability Assays
2.4. qPCR
2.5. Data Analysis
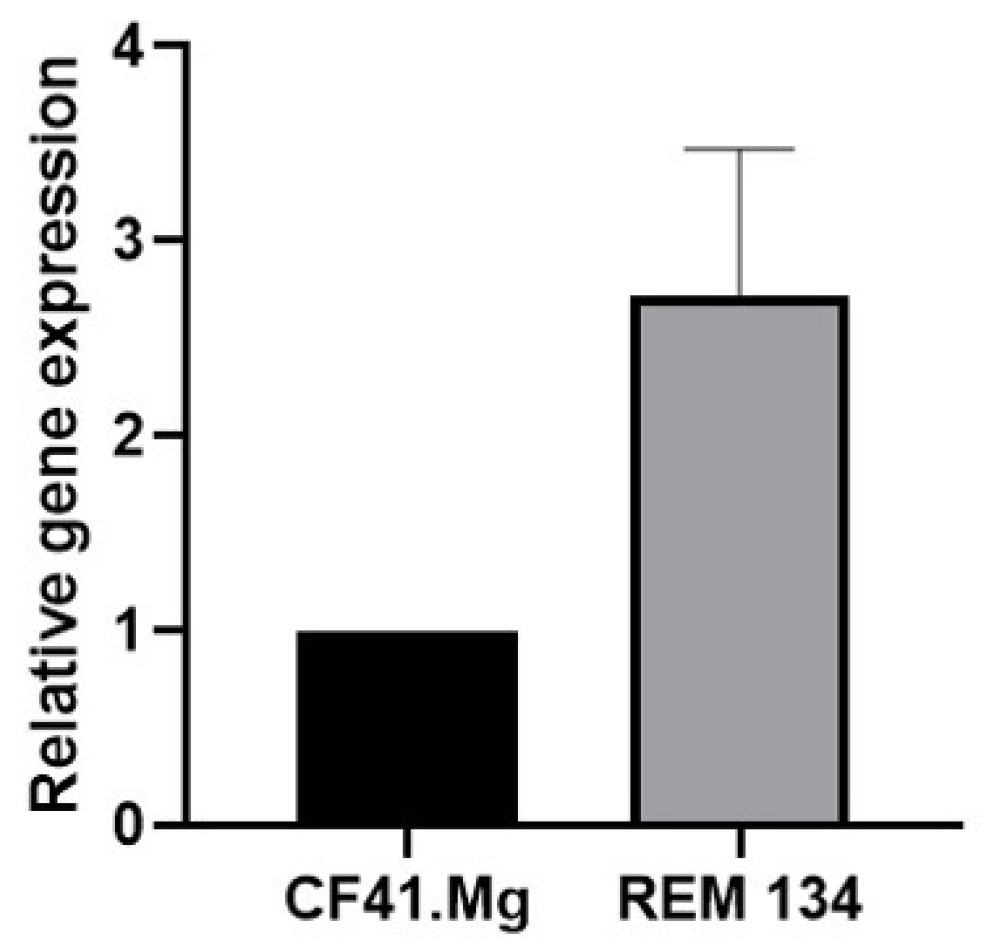
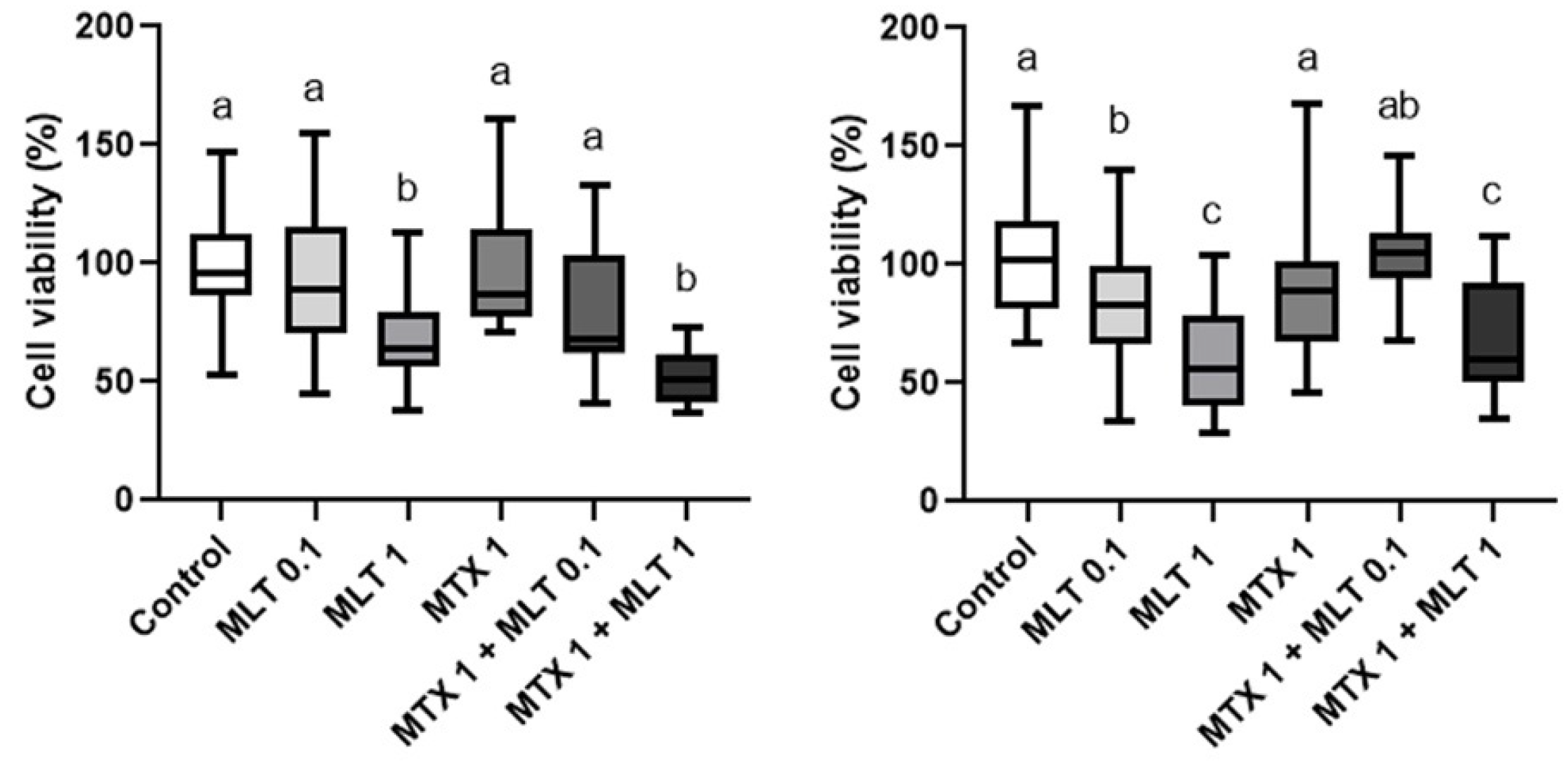
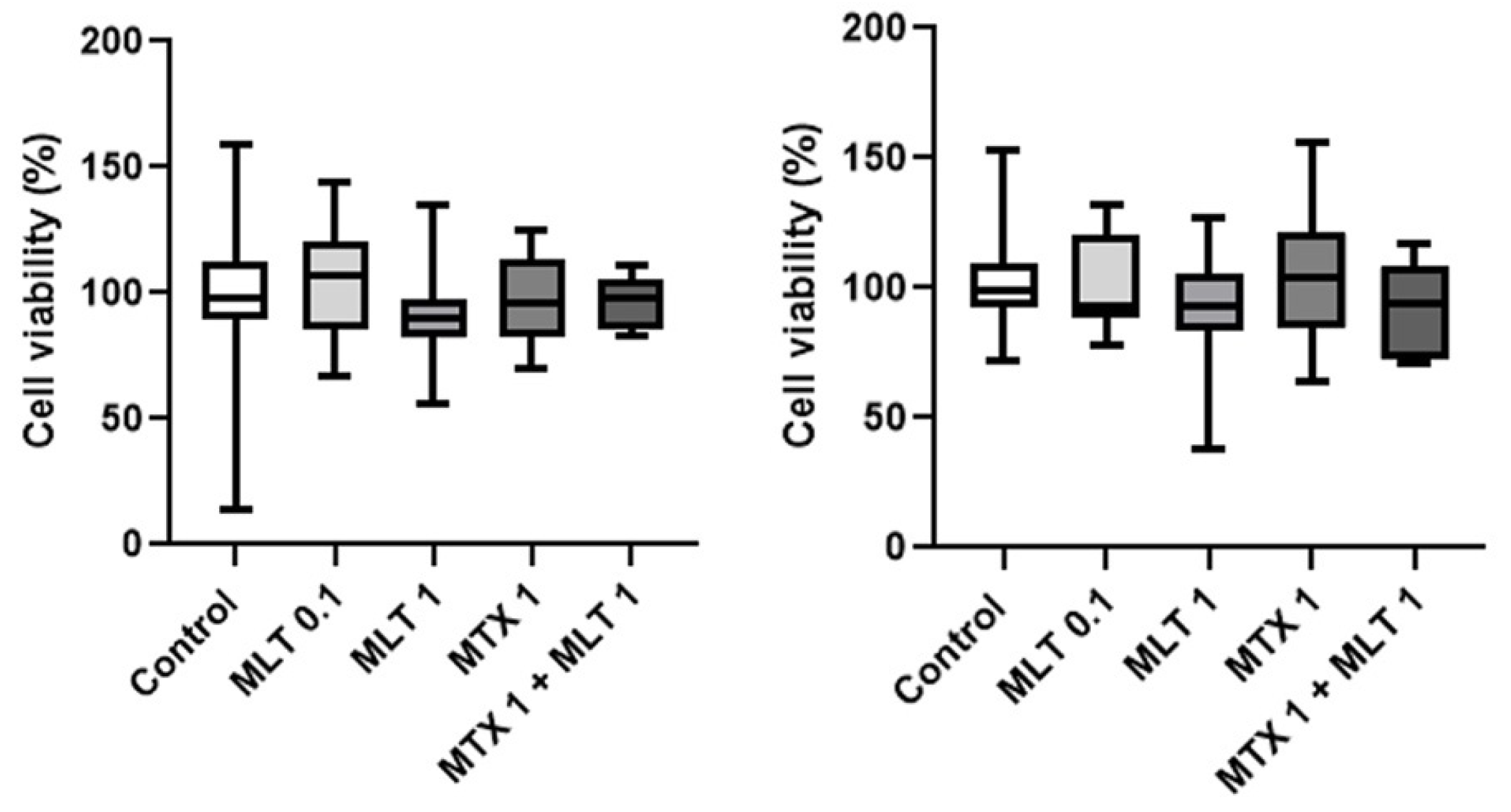
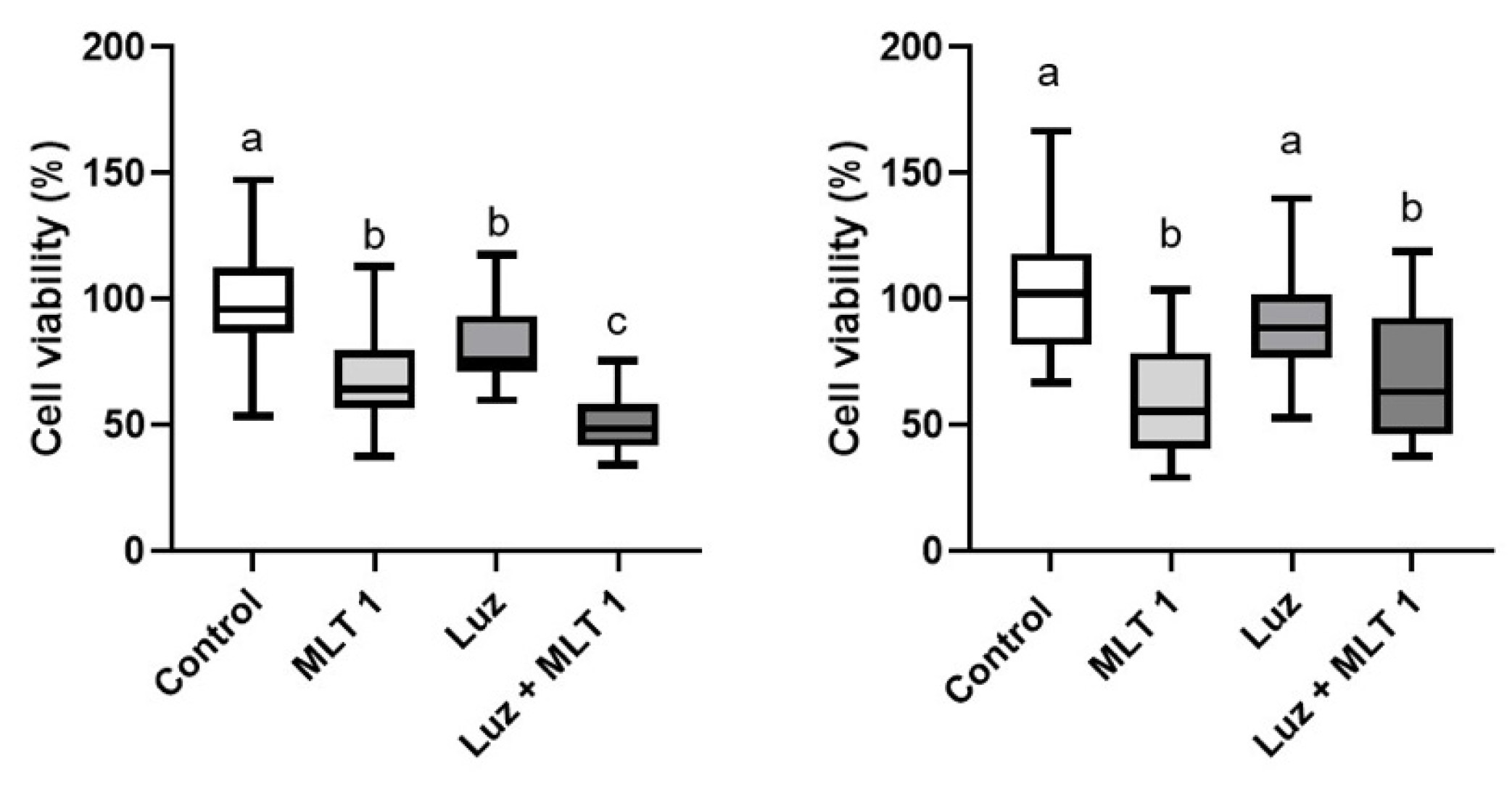
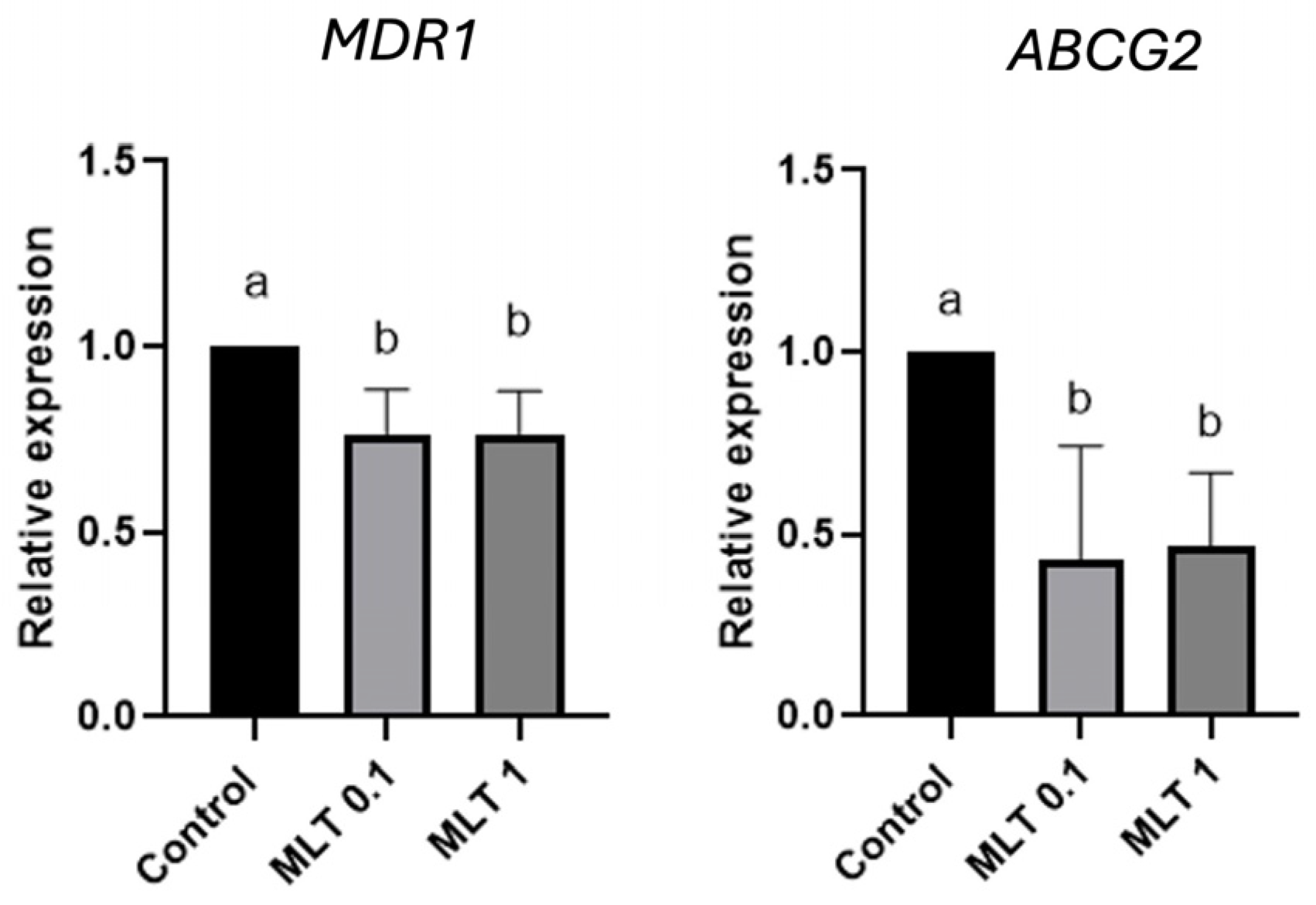
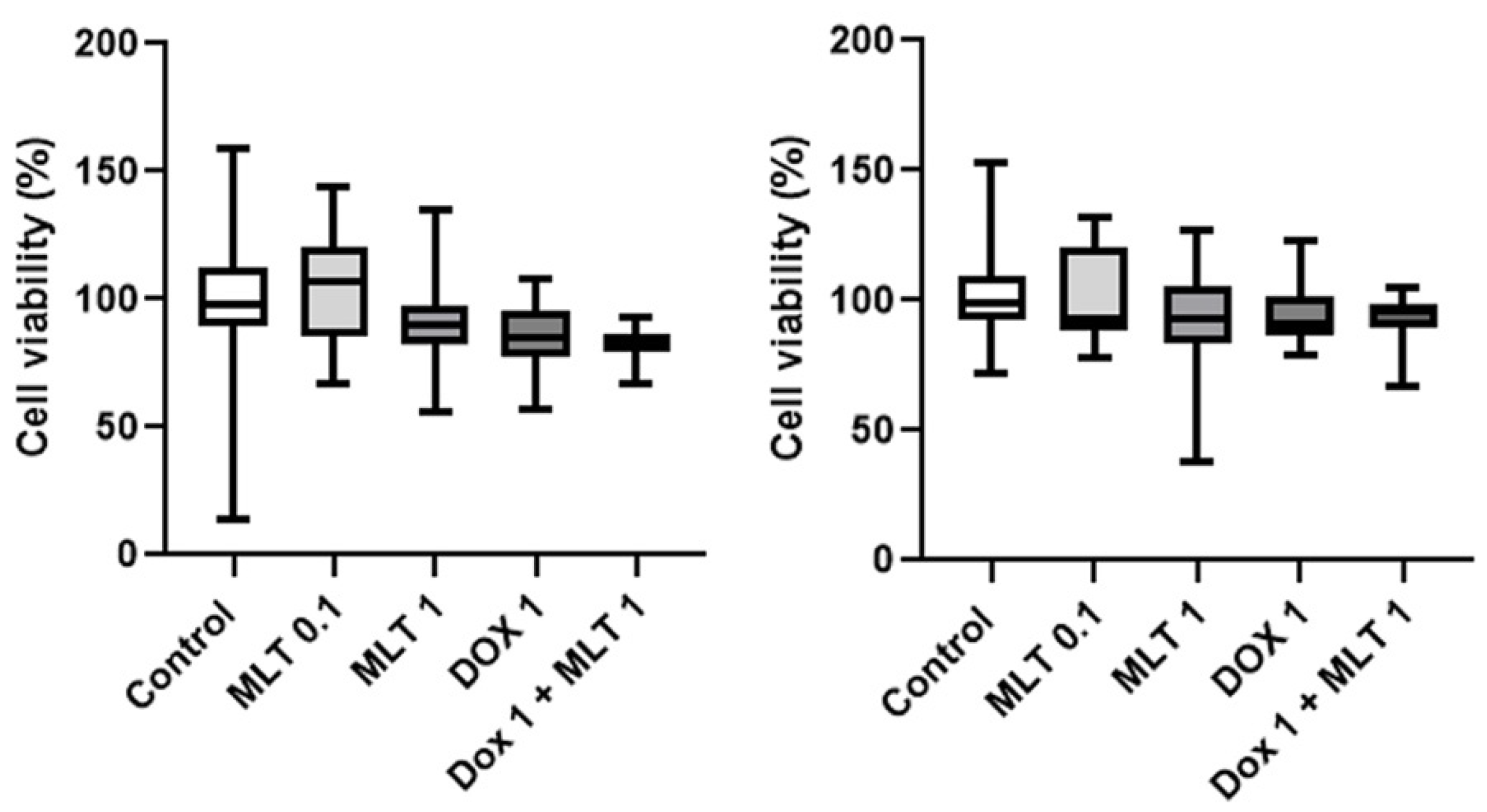
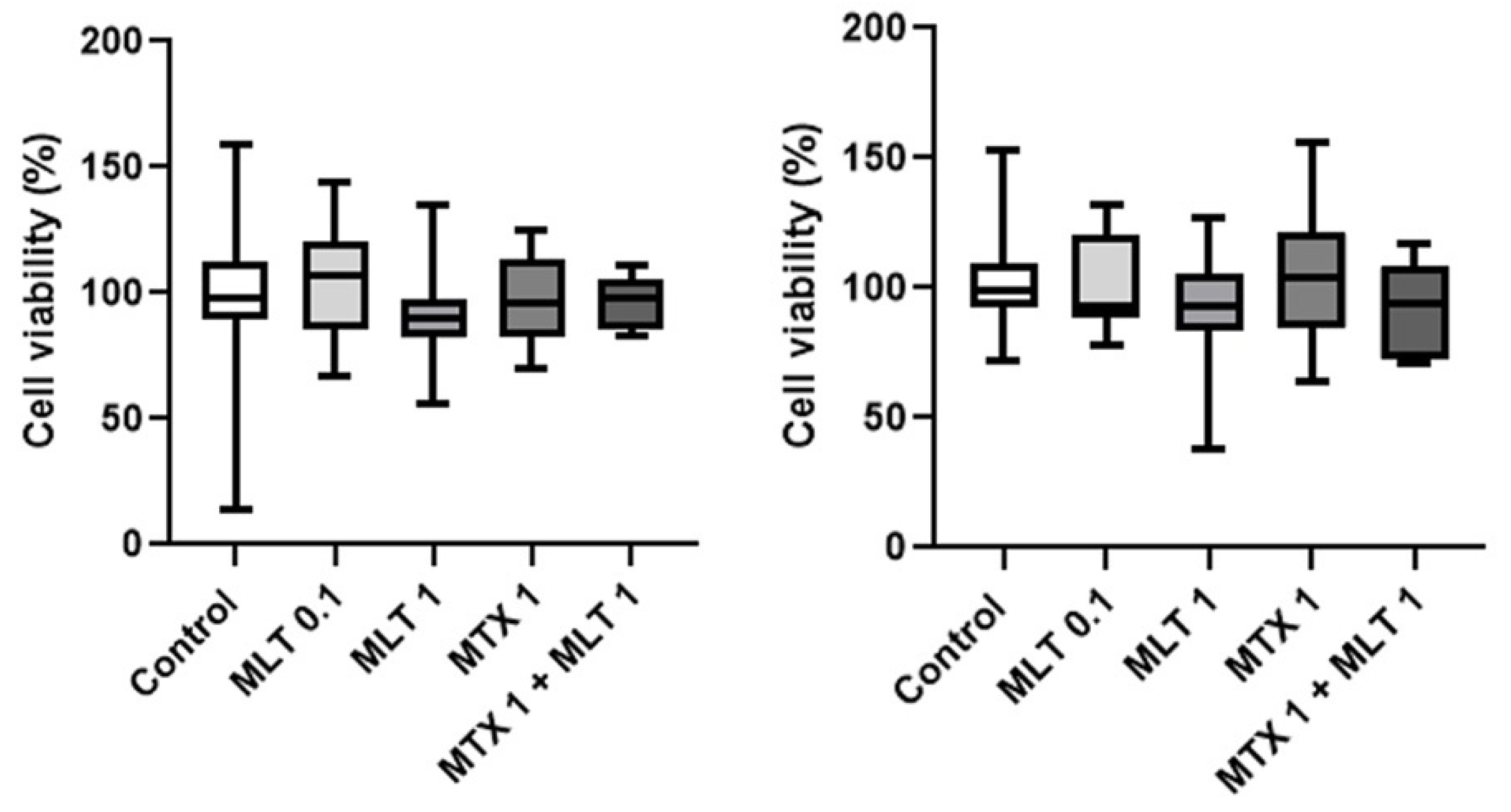
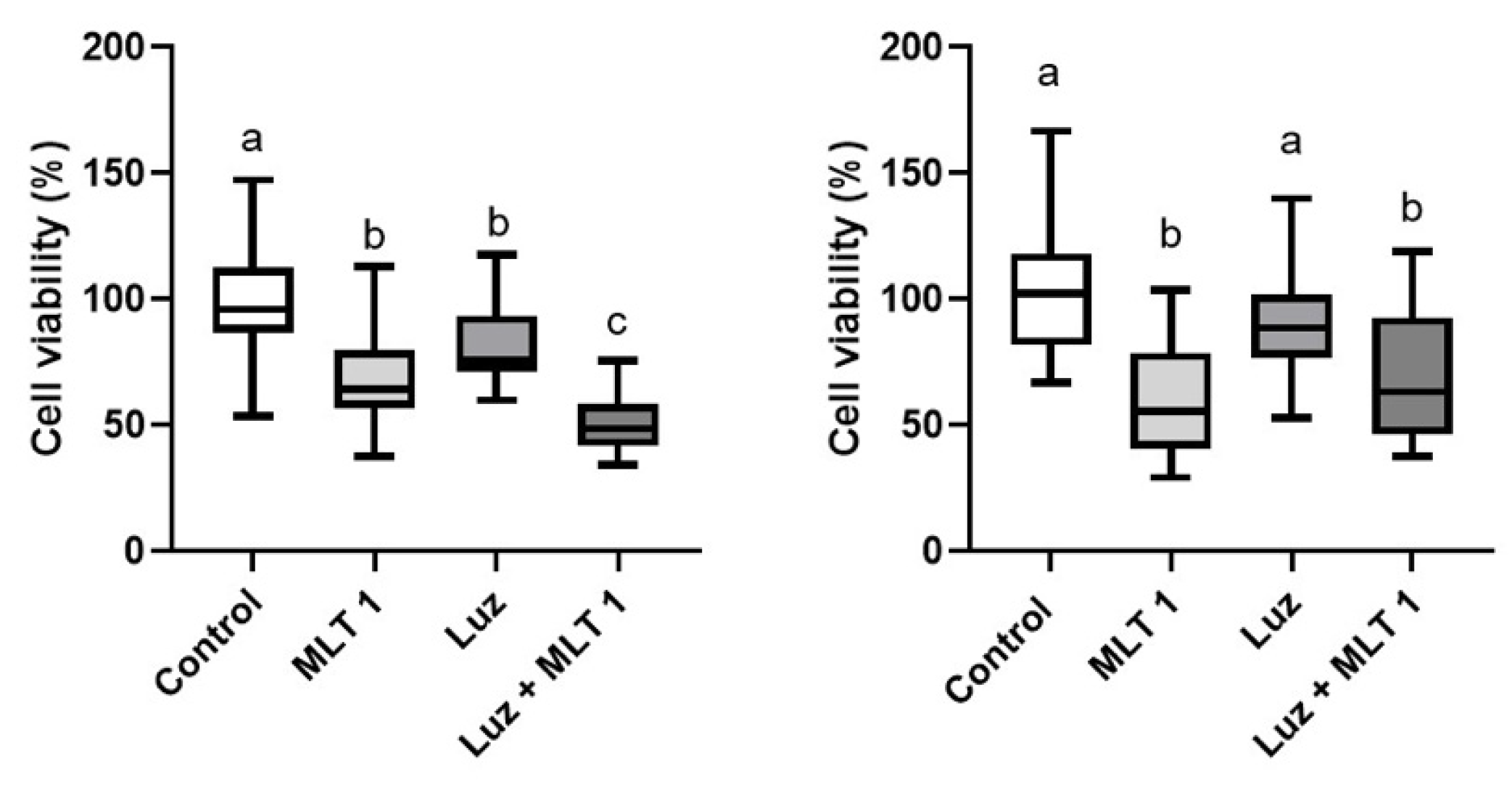
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Salas, Y.; Marquez, A.; Diaz, D.; Romero, L. Epidemiological study of mammary tumors in female dogs diagnosed during the period 2002–2012: A growing animal health problem. PLoS ONE 2015, 10, e0127381. [Google Scholar] [CrossRef] [PubMed]
- Valdivia, G.; Alonso, A.; Perez-Alenza, D.; Peña, L. From conventional to precision therapy in canine mammary cancer: A comprehensive review. Front. Vet. Sci. 2021, 17, 623800. [Google Scholar] [CrossRef] [PubMed]
- Klopfleisch, R.; Von Euler, H.; Sarli, G.; Punho, S.; Gartner, F.; Gruber, A. Molecular carcinogenesis of canine mammary tumors: News from an old disease. Vet. Pathol. 2011, 48, 98–116. [Google Scholar] [CrossRef] [PubMed]
- Michishita, M. Understanding of tumourigenesis in canine mammary tumours based on cancer stem cell research. Vet. J. 2020, 265, 105560. [Google Scholar] [CrossRef] [PubMed]
- Goldschmidt, M.; Peña, L.; Rasotto, R.; Zappulli, V. Classification and grading of canine mammary tumors. Vet. Pathol. 2011, 48, 117–131. [Google Scholar] [CrossRef] [PubMed]
- Goldschmidt, M.; Peña, L.; Zappulli, V. Tumors of the Mammary Gland. In Meuten’s Tumor in Domestic Animals, 5th ed.; John Wiley & Sons: Ames, IA, USA, 2017; pp. 723–765. [Google Scholar]
- Arenas, C.; Peña, L.; Granados-Soler, J.; Perez-Alenza, M. Adjuvant therapy for highly malignant canine mammary tumours: Cox-2 inhibitor versus chemotherapy: A case-control prospective study. Vet. Rec. 2016, 179, 125. [Google Scholar] [CrossRef] [PubMed]
- Klopfleisch, R.; Kohn, B.; Gruber, A. Mechanisms of tumour resistance against chemotherapeutic agents in veterinary oncology. Vet. J. 2016, 207, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Rybicka, A.; Krol, M. Identification and characterization of cancer stem cells in canine mammary tumors. Acta Vet. Scand. 2016, 58, 86. [Google Scholar] [CrossRef] [PubMed]
- Im, K.; Jang, Y.; Shin, J.; Kim, N.; Lim, H.; Lee, S.; Kim, J.; Sur, J. CD44+/CD24– cancer stem cells are associated with higher grade of canine mammary carcinomas. Vet. Pathol. 2015, 52, 1041–1044. [Google Scholar] [CrossRef] [PubMed]
- Michishita, M.; Akiyoshi, R.; Yoshimura, H.; Katsumoto, T.; Ichikawa, H.; Ohkusu-Tsukada, K.; Nakagawa, T.; Sasaski, N.; Takahashi, K. Characterization of spheres derived from canine mammary gland adenocarcinoma cell lines. Res. Vet. Sci. 2011, 91, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Pang, L.; Argyle, D. The evolving cancer stem cell paradigm: Implications in veterinary oncology. Vet. J. 2015, 205, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Pang, L.; Cervantes-Arias, A.; Else, R.; Argyle, D. Canine mammary cancer stem cells are radio- and chemo-resistant and exhibit an epithelial-mesenchymal transition phenotype. Cancers 2011, 3, 1744. [Google Scholar] [CrossRef] [PubMed]
- Torres, C.G.; Olivares, A.; Stoore, C. Simvastatin exhibits antiproliferative effects on spheres derived from canine mammary carcinoma cells. Oncol. Rep. 2015, 33, 2235–2244. [Google Scholar] [CrossRef] [PubMed]
- Honscha, K.; Schirmer, A.; Reischauer, A.; Schoon, H.; Einspanier, A.; Gabel, G. Expression of ABC-transport proteins in canine mammary cancer: Consequences for chemotherapy. Reprod. Domest. Anim. 2009, 44, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Hasanabady, M.; Kalalinia, F. ABCG2 inhibition as a therapeutic approach for overcoming multidrug resistance in cancer. J. Biosci. 2016, 41, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Cruz, P.; Reyes, F.; Torres, C. Simvastatin modulates β- catenin/MDR1 expression on spheres derived from CF41.Mg canine mammary carcinoma cells. Pol. J. Vet. Sci. 2018, 21, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Dontu, G.; Wicha, M. Mammary stem cells, self-renewal pathways, and carcinogenesis. Breast Cancer Res. 2005, 7, 86. [Google Scholar] [CrossRef] [PubMed]
- Levi, M.; Peña, L.; Alonso-Díez, A.; Brunetti, B.; Muscatello, L.V.; Benazzi, C.; Pérez-Alenza, M.D.; Sarli, G. P-glycoprotein and breast cancer resistance protein in canine inflammatory and non-inflammatory grade III mammary carcinomas. Vet. Pathol. 2019, 56, 840–847. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Lopes, J.; Maschio, L.; Jardim-Perassi, B.; Moschetta, M.; Ferreira, L.; Martins, G.; Gelaleti, G.; De Campos Zuccari, D. Evaluation of melatonin treatment in primary culture of canine mammary tumors. Oncol. Rep. 2015, 33, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Hill, S.; Belancio, V.; Dauchy, R.; Xiang, S.; Brimer, S.; Mao, L.; Hauch, A.; Lundberg, P.; Summers, W.; Yuan, L.; et al. Melatonin: An inhibitor of breast cancer. Endocr. Relat. Cancer 2015, 22, 183–204. [Google Scholar] [CrossRef] [PubMed]
- Cucielo, M.; Freire, P.; Emilio-Silva, M.; Romagnoli, G.; Carvalho, R.; Kaneno, R.; Hiruma-Lima, C.; Delella, F.; Reiter, R.; Chuffa, L. Melatonin enhances cell death and suppresses the metastatic capacity of ovarian cancer cells by attenuating the signaling of multiple kinases. Pathol. Res. Prac. 2023, 248, 154637. [Google Scholar] [CrossRef] [PubMed]
- Di Bella, G.; Mascia, F.; Gualano, L.; Di Bella, L. Melatonin Anticancer Effects: Review. Int. J. Mol. Sci. 2013, 14, 2410–2430. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.; Gao, R.; Wang, Z.; Wang, X.; Fang, Y.; Gao, J.; Reiter, R.; Wang, J. Melatonin: A potential therapeutic option for breast cancer. Trends Endocrinol. Metab. 2020, 31, 859–871. [Google Scholar] [CrossRef] [PubMed]
- Maroufi, N.; Ashouri, N.; Mortezania, Z.; Ashoori, Z.; Vahedian, V.; Amirzadeh-Iranaq, M.; Fattahi, A.; Kazemzadeh, H.; Bizzarri, M.; Akbarzadeh, M.; et al. The potential therapeutic effects of melatonin on breast cancer: An invasion and metastasis inhibitor. Pathol. Res. Pract. 2020, 216, 153226. [Google Scholar] [CrossRef] [PubMed]
- Maroufi, N.; Vahedian, V.; Hemati, S.; Rashidi, M.; Akbarzadeh, M.; Zahedi, M.; Pouremamali, F.; Isazadeh, A.; Taefehshokr, S.; Hajazimian, S.; et al. Targeting cancer stem cells by melatonin: Effective therapy for cancer treatment. Pathol. Res. Pract. 2020, 216, 152919. [Google Scholar] [CrossRef] [PubMed]
- Moloudizargari, M.; Moradkhani, F.; Hekmatirad, S.; Fallah, M.; Asghari, M.; Reiter, R. Therapeutic targets of cancer drugs: Modulation by melatonin. Life Sci. 2021, 15, 118934. [Google Scholar] [CrossRef] [PubMed]
- Sadoughi, F.; Dana, P.; Asemi, Z.; Shafabakhash, R.; Mohammadi, S.; Heidar, Z.; Mirzamoradi, M.; Targhazeh, N.; Mirzaei, H. Molecular and cellular mechanisms of melatonin in breast cancer. Biochimie 2022, 202, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Goncalves ND, N.; Colombo, J.; Lopes, J.R.; Gelaleti, G.B.; Moschetta, M.G.; Sonehara, N.M.; Hellmén, E.; de Freitas Zanon, C.; Oliani, S.M.; Zuccari, D.A. Effect of melatonin in epithelial-mesenchymal transition markers and invasive properties of breast cancer stem cells of canine and human cell lines. PLoS ONE 2016, 11, e0150407. [Google Scholar] [CrossRef] [PubMed]
- Serrano, C.; Guzmán, S.; Arias, J.I.; Torres, C.G. Melatonin decreases in vitro viability and migration of spheres derived from CF41.Mg canine mammary carcinoma cells. BMC Vet. Res. 2019, 15, 390. [Google Scholar] [CrossRef] [PubMed]
- Azpeleta, C.; Delgado, M.J.; Metz, J.R.; Flik, G.; de Pedro, N. Melatonin as an anti-stress signal: Effects on an acute stress model and direct actions on interrenal tissue in goldfish. Front. Endocrinol. 2024, 14, 1291153. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Bok, E.; Lee, S.; Lee, H.-J.; Choe, Y.; Kim, N.H.; Lee, W.J.; Rho, G.-J.; Lee, S.-L. Metastasis prognostic factors and cancer stem cell-related transcription factors associated with metastasis induction in canine metastatic mammary gland tumors. J. Vet. Sci. 2021, 22, e62. [Google Scholar] [CrossRef] [PubMed]
- Jablonska, K.; Pula, B.; Zemla, A.; Owczarek, T.; Wojnar, A.; Rys, J.; Ambicka, A.; Podhorska-Okolow, M.; Ugorski, M.; Dziegiel, P. Expression of melatonin receptor MT1 in cells of human invasive ductal breast carcinoma. J. Pineal Res. 2013, 54, 334–345. [Google Scholar] [CrossRef] [PubMed]
- Mao, L.; Lin, Y.; Xiang, S.; Zeringue, S.B.; Dauchy, R.T.; Blask, D.E.; Hauch, A.; Hill, S.M. Molecular deficiency (ies) in MT1 melatonin signaling pathway underlies the melatonin-unresponsive phenotype in MDA-MB-231 human breast cancer cells. J. Pineal Res. 2014, 56, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Rögelsperger, O.; Ekmekcioglu, C.; Jäger, W.; Klimpfinger, M.; Königsberg, R.; Krenbek, D.; Sellner, F.; Thalhammer, T. Coexpression of the melatonin receptor 1 and nestin in human breast cancer specimens. J. Pineal Res. 2009, 46, 422–432. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; You, J.; Luo, W.; Chen, B.; Feng, Q.; Wu, B.; Jiang, L.; Luo, Q. Reduced tumorigenicity and drug resistance through the downregulation of octamer-binding protein 4 and Nanog transcriptional factor expression in human breast stem cells. Mol. Med. Rep. 2015, 11, 1647–1654. [Google Scholar] [CrossRef] [PubMed]
- Sen, U.; Shanavas, S.; Nagendra, A.H.; Nihad, M.; Chaudhury, D.; Rachamalla, H.K.; Banerjee, R.; Shenoy, P.S.; Bose, B. Significance of Oct-4 transcription factor as a pivotal therapeutic target for CD44+/24− mammary tumor initiating cells: Aiming at the root of the recurrence. Cell Biol. Int. 2023, 47, 742–753. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Han, Z.; Zhu, Y.; Chen, J.; Li, W. The role and specific mechanism of OCT4 in cancer stem cells: A review. Int. J. Stem. Cells 2020, 13, 312–325. [Google Scholar] [CrossRef] [PubMed]
- Lopes, J.; Arnosti, D.; Trosko, J.E.; Tai, M.H.; Zuccari, D. Melatonin decreases estrogen receptor binding to estrogen response elements sites on the oct4 gene in human breast cancer stem cells. Genes Cancer 2016, 7, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Maroufi, N.F.; Rashidi, M.; Vahedian, V.; Jahanbazi, R.; Mostafaei, S.; Akbarzadeh, M.; Kazemzadeh, H.; Nejabati, H.-R.; Isazadeh, A.; Rashidi, M.-R.; et al. Effect of Apatinib plus melatonin on vasculogenic mimicry formation by cancer stem cells from breast cancer cell line. Breast Cancer 2022, 29, 260–273. [Google Scholar] [CrossRef] [PubMed]
- Maroufi, N.F.; Amiri, M.; Dizaji, B.F.; Vahedian, V.; Akbarzadeh, M.; Roshanravan, N.; Haiaty, S.; Nouri, M.; Rashidi, M.-R. Inhibitory effect of melatonin on hypoxia-induced vasculogenic mimicry via suppressing epithelial-mesenchymal transition (EMT) in breast cancer stem cells. Eur. J. Pharmacol. 2020, 881, 173282. [Google Scholar] [CrossRef] [PubMed]
- Pongkan, W.; Piamsiri, C.; Dechvongya, S.; Punyapornwitthaya, V.; Boonyapakorn, C. Short-term melatonin supplementation decreases oxidative stress but does not affect left ventricular structure and function in myxomatous mitral valve degenerative dogs. BMC Vet. Res. 2022, 18, 24. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Sang, X.; Wang, M.; Liu, Y.; Liu, J.; Wang, X.; Liu, P.; Cheng, H. Melatonin potentiates the cytotoxic effect of Neratinib in HER2+ breast cancer through promoting endocytosis and lysosomal degradation of HER2. Oncogene 2021, 40, 6273–6283. [Google Scholar] [CrossRef] [PubMed]
- Sang, X.; Li, L.; Rui, C.; Liu, Y.; Liu, Z.; Tao, Z.; Cheng, H.; Liu, P. Induction of EnR stress by Melatonin enhances the cytotoxic effect of Lapatinib in HER2-positive breast cancer. Cancer Lett. 2021, 518, 82–93. [Google Scholar] [CrossRef] [PubMed]
- Leon-Blanco, M.M.; Guerrero, J.M.; Reiter, R.J.; Calvo, J.R.; Pozo, D. Melatonin inhibits telomerase activity in the MCF-7 tumor cell line both in vivo and in vitro. J. Pineal Res. 2003, 35, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Kukal, S.; Guin, D.; Rawat, C.; Bora, S.; Mishra, M.; Sharma, P.; Paul, P.; Kanojia, N.; Grewal, G.; Kukreti, S.; et al. Multidrug efflux transporter ABCG2: Expression and regulation. Cell Mol. Life Sci. 2021, 78, 6887–6939. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Zhang, C.; Chena, J.; Zhanga, S.; Pana, G.; Xina, Y.; Lin, L.; You, Z. Shenmai injection suppresses multidrug resistance in MCF-7/ADR cells through the MAPK/NF-kB signaling pathway. Pharm. Biol. 2020, 58, 276–285. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.; Frank, T. Teaching an old dog new tricks: Reactivated developmental signaling pathways regulate ABCB1 and chemoresistance in cancer. Cancer Drug Resist. 2021, 4, 424–452. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, H.; Chen, X. Drug-resistance and combating drug resistance in cancer. Cancer Drug Resist. 2019, 2, 141–160. [Google Scholar] [CrossRef] [PubMed]
- Martin, V.; Sanchez, A.; Herrera, F.; Gomez-Manzano, C.; Fueyo, J.; Alvarez-Vega, M.; Antolin, I.; Rodriguez, C. Melatonin-induced methylation of the ABCG2/BCRP promoter as a novel mechanism to overcome multidrug resistance in brain tumour stem cells. Br. J. Cancer 2013, 108, 2005–2012. [Google Scholar] [CrossRef] [PubMed]
- Fic, M.; Gomulkiewicz, A.; Grzegrzolka, J.; Podhorska-Okolow, M.; Zabel, M.; Dziegiel, P.; Jablonska, K. The impact of melatonin on colon cancer cells’ resistance to doxorubicin in an in vitro study. Int. J. Mol. Sci. 2017, 18, 1396. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, M.; Lin, C.; Su, S.; Reiter, R.; Chen, A.; Chen, M.; Yang, S. Effects of miR-34b/miR-892a upregulation and inhibition of ABCB1/ABCB4 on melatonin-induced apoptosis in VCR-resistant oral cancer cells. Mol. Ther. Nucleic Acids 2020, 19, 877–889. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Genbank Access Nº | Primer Pair Sequences |
|---|---|---|
| β actin | NC_000071.7 | FWD: 5′-CAAATGTGGATCAGCAAGCAG-3′ REV: 5′-GAAAGGGTGTAACGCAACTAAAG-3′ |
| MDR1 | NC_006596.2 | FWD: 5′-ACAGGAGATTGGCTGGTTTG-3′ REV: 5′-AAGTCCAAGAACAGGGCTGA-3′ |
| ABCG2 | NC_006614.2 | FWD: 5′-GACCTCCAACGACCTGAAGA-3′ REV: 5′-GAAGATTTGCCTCCACCTGT-3′ |
| MT1 | NR | FWD: 5′-TGTGCTTTCTAAACCTTTCTCCT-3′ REV: 5′- CACGAAGCCACTGATTTGGC-3′ |
| MT2 | NR | FWD: 5′-CTTGCTGACTTTTGCTCCCT-3′ REV: 5′-CGAATGACACTCAGCCCCAT-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cataldo, D.; Aravena, G.; Escobar, A.; Tapia, J.C.; Peralta, O.A.; Torres, C.G. Effect of Melatonin on Chemoresistance Exhibited by Spheres Derived from Canine Mammary Carcinoma Cells. Animals 2024, 14, 1229. https://doi.org/10.3390/ani14081229
Cataldo D, Aravena G, Escobar A, Tapia JC, Peralta OA, Torres CG. Effect of Melatonin on Chemoresistance Exhibited by Spheres Derived from Canine Mammary Carcinoma Cells. Animals. 2024; 14(8):1229. https://doi.org/10.3390/ani14081229
Chicago/Turabian StyleCataldo, Dania, Guillermo Aravena, Alejandro Escobar, Julio C. Tapia, Oscar A. Peralta, and Cristian G. Torres. 2024. "Effect of Melatonin on Chemoresistance Exhibited by Spheres Derived from Canine Mammary Carcinoma Cells" Animals 14, no. 8: 1229. https://doi.org/10.3390/ani14081229