

The Supplementation of Berberine in High-Carbohydrate Diets Improves Glucose Metabolism of Tilapia (Oreochromis niloticus) via Transcriptome, Bile Acid Synthesis Gene Expression and Intestinal Flora

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Diets and Animals
2.2. Sample Collection and Chemical Analysis
2.3. Intestine Microbiological Analysis
2.4. Histological Analysis of Liver
2.5. Real-Time Quantitative RT-PCR Analysis of Gene Expression
2.6. Transcriptome Sequencing and Analysis
2.7. Challenge Test
2.8. Statistical Analysis
3. Results
3.1. Growth Performance
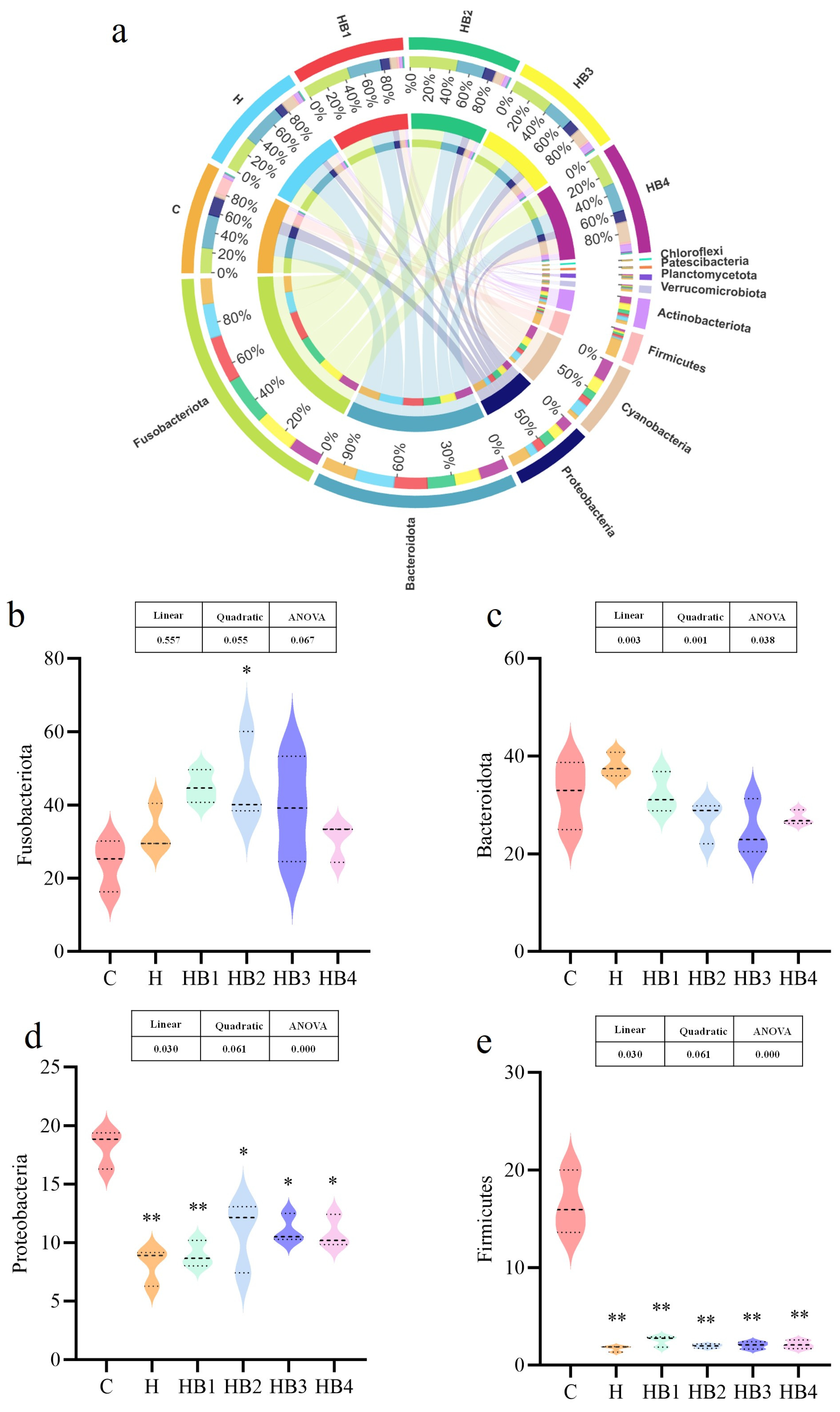
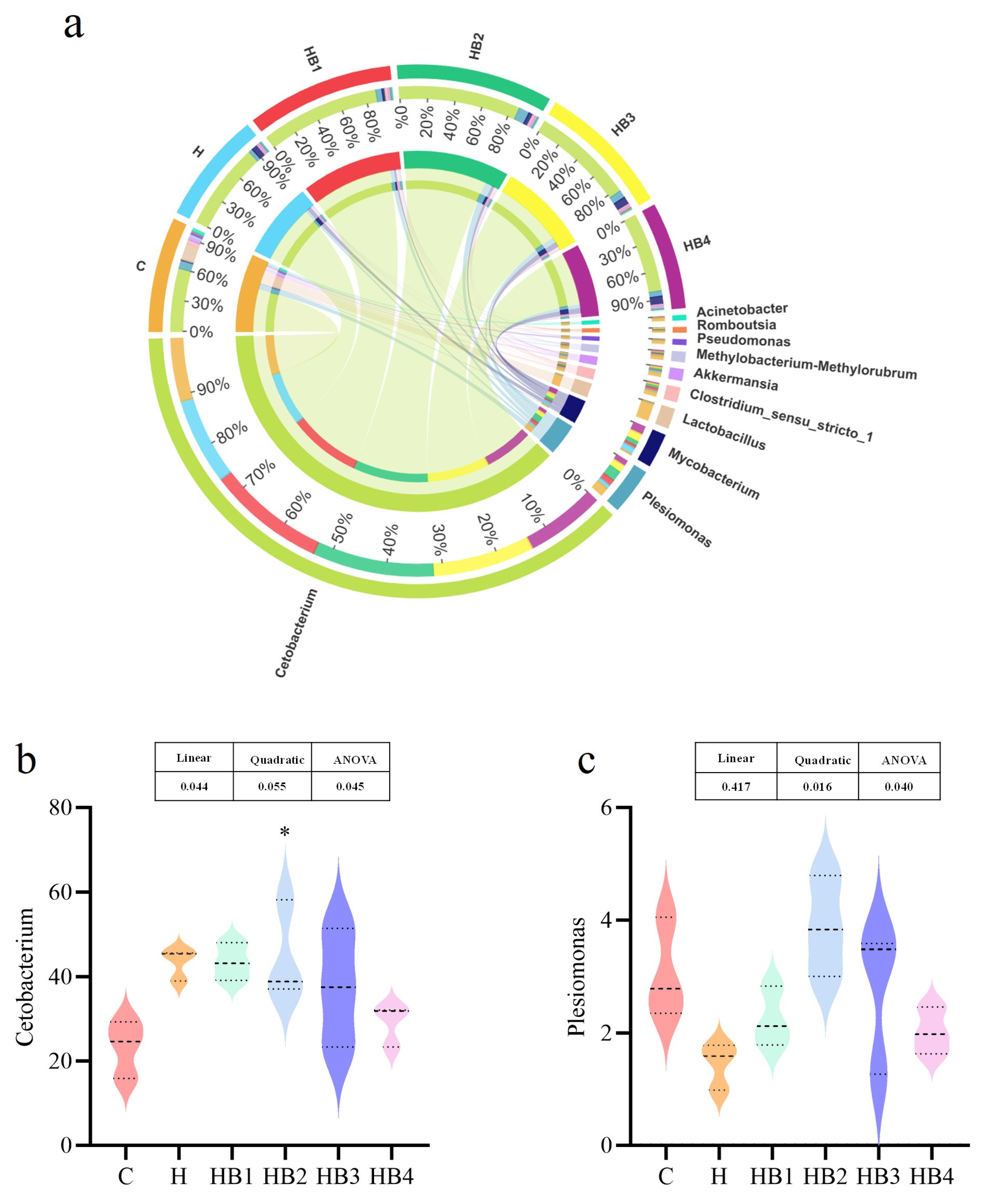
3.2. Intestinal Flora Analysis
3.2.1. Richness and Diversity Analysis
3.2.2. Comparison of the Intestinal Microbiota Composition
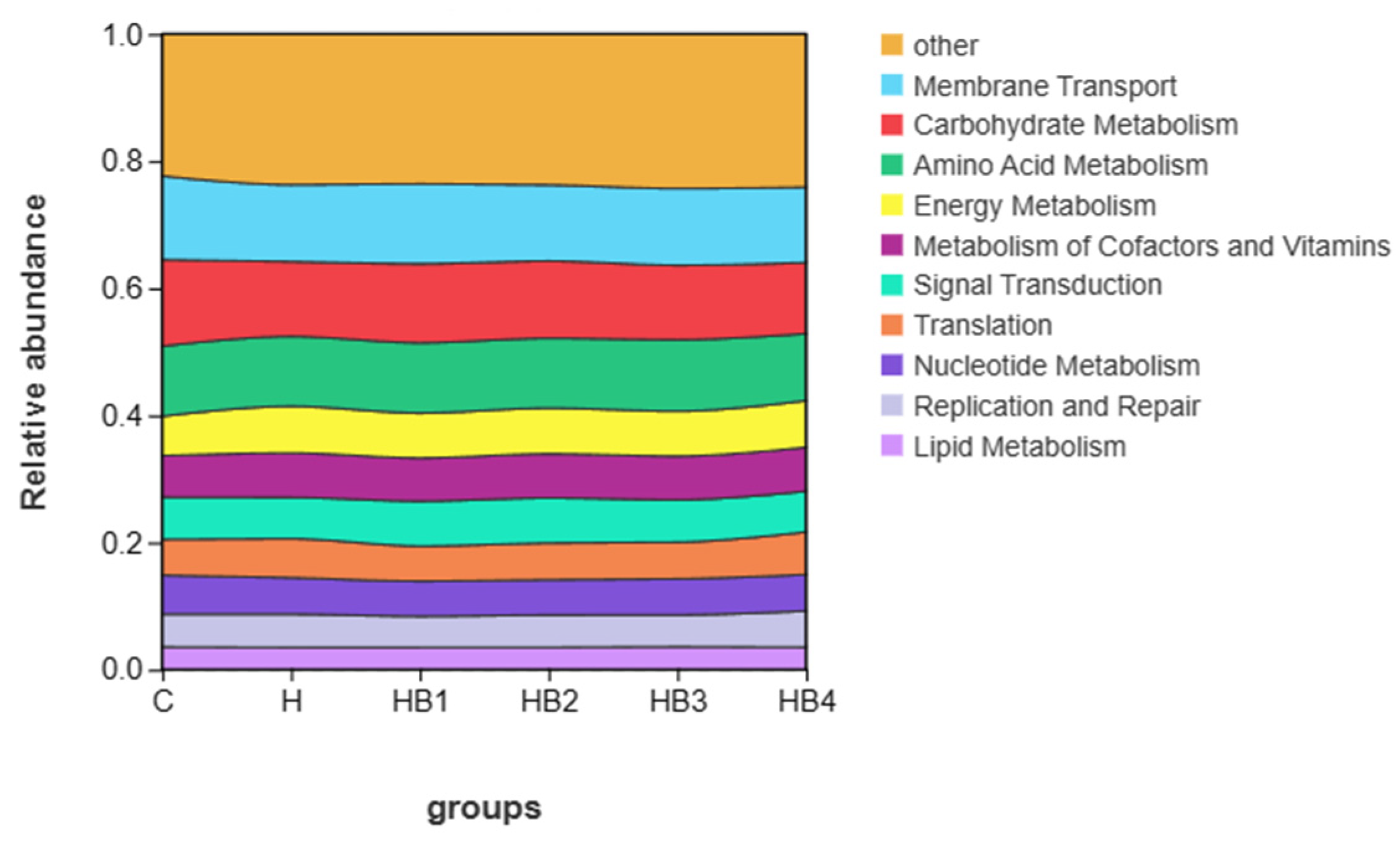
3.2.3. Functional Analysis of the Microbial Community in the Intestines
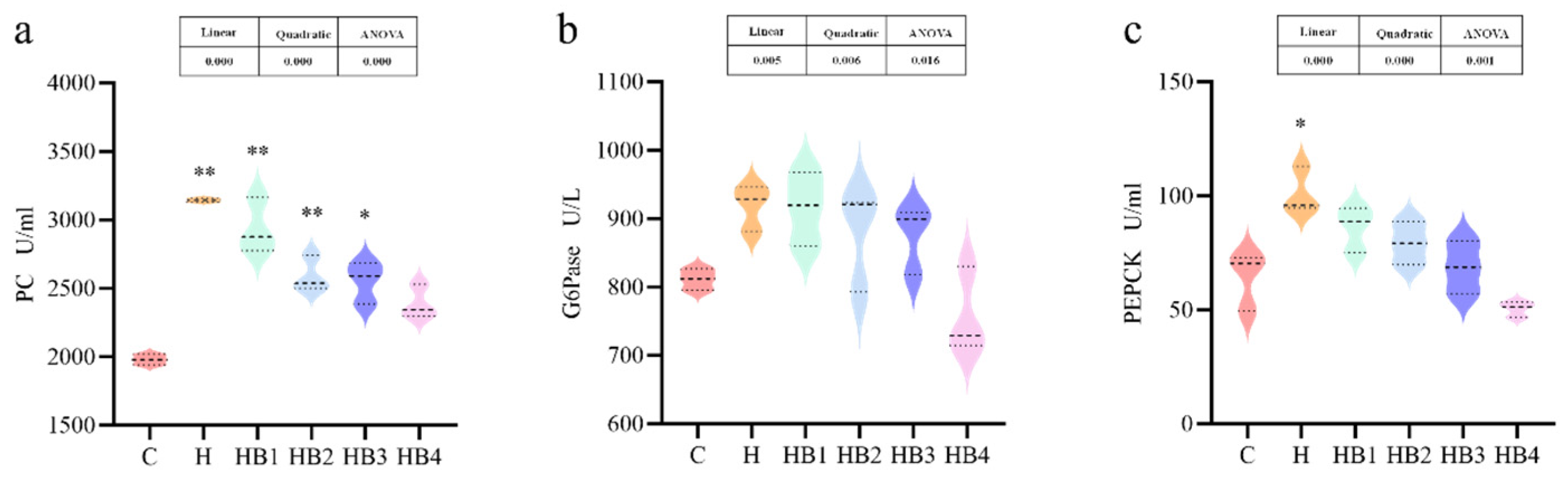
3.3. Enzyme Activity Related to Bile Acid Synthesis in the Liver
3.4. Bile Acids Content Assay
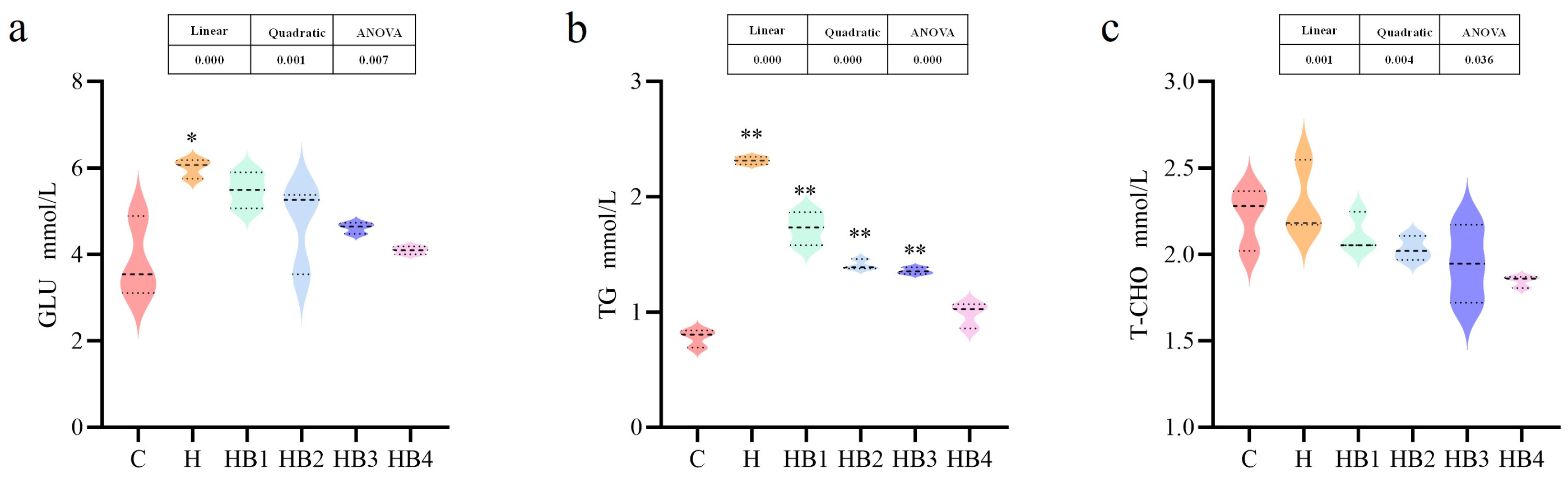
3.5. Serum Biochemical Composition Measurement
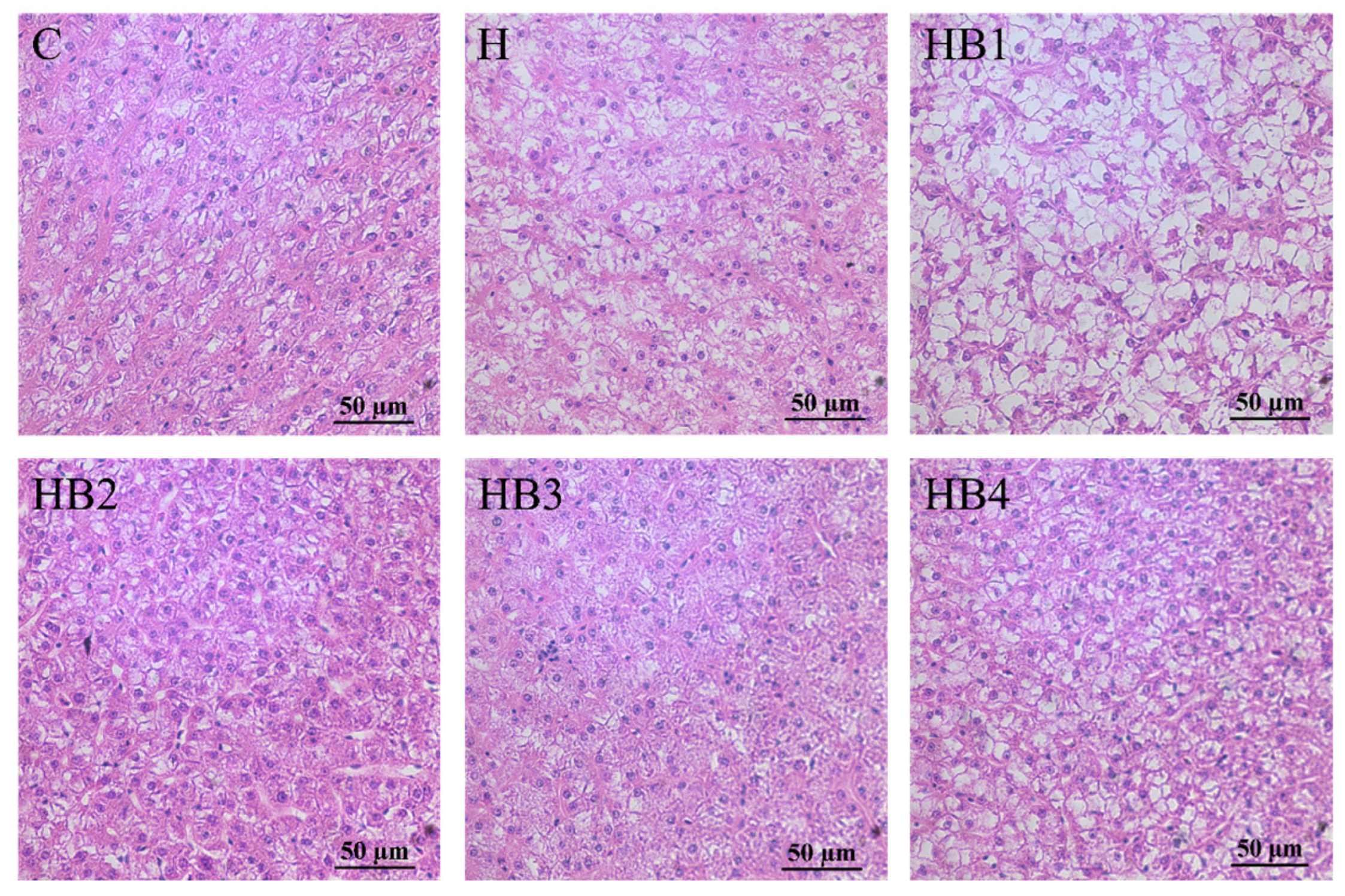
3.6. Liver Morphology
3.7. Glycogen Content Assay
3.8. Enzyme Activity Related to Glucose Metabolism in the Liver
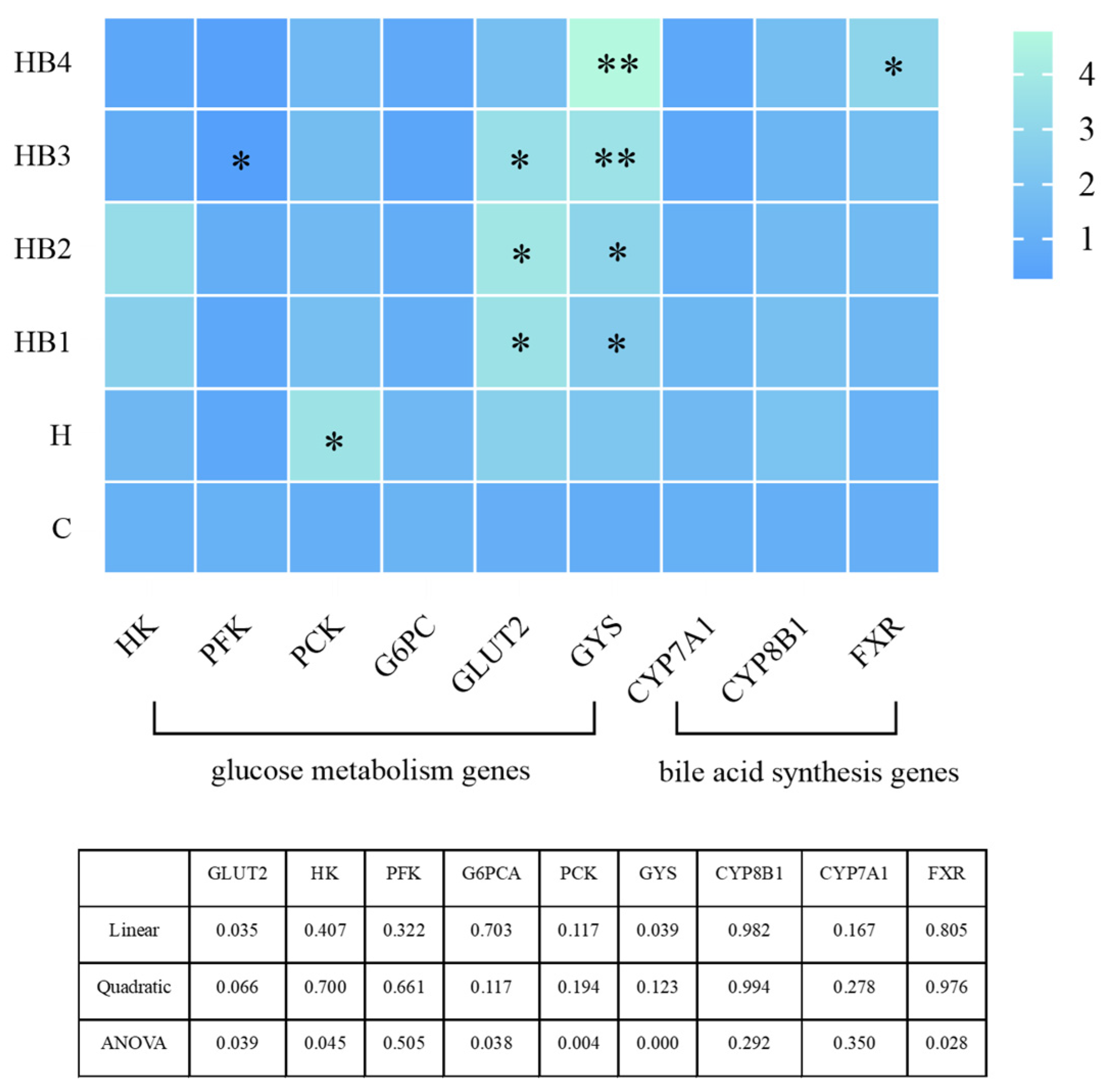
3.9. Gene Expression
3.10. Transcriptome Analysis
3.10.1. Sequence Alignment Analysis
3.10.2. Analysis of Differentially Expressed Genes (DEGs)
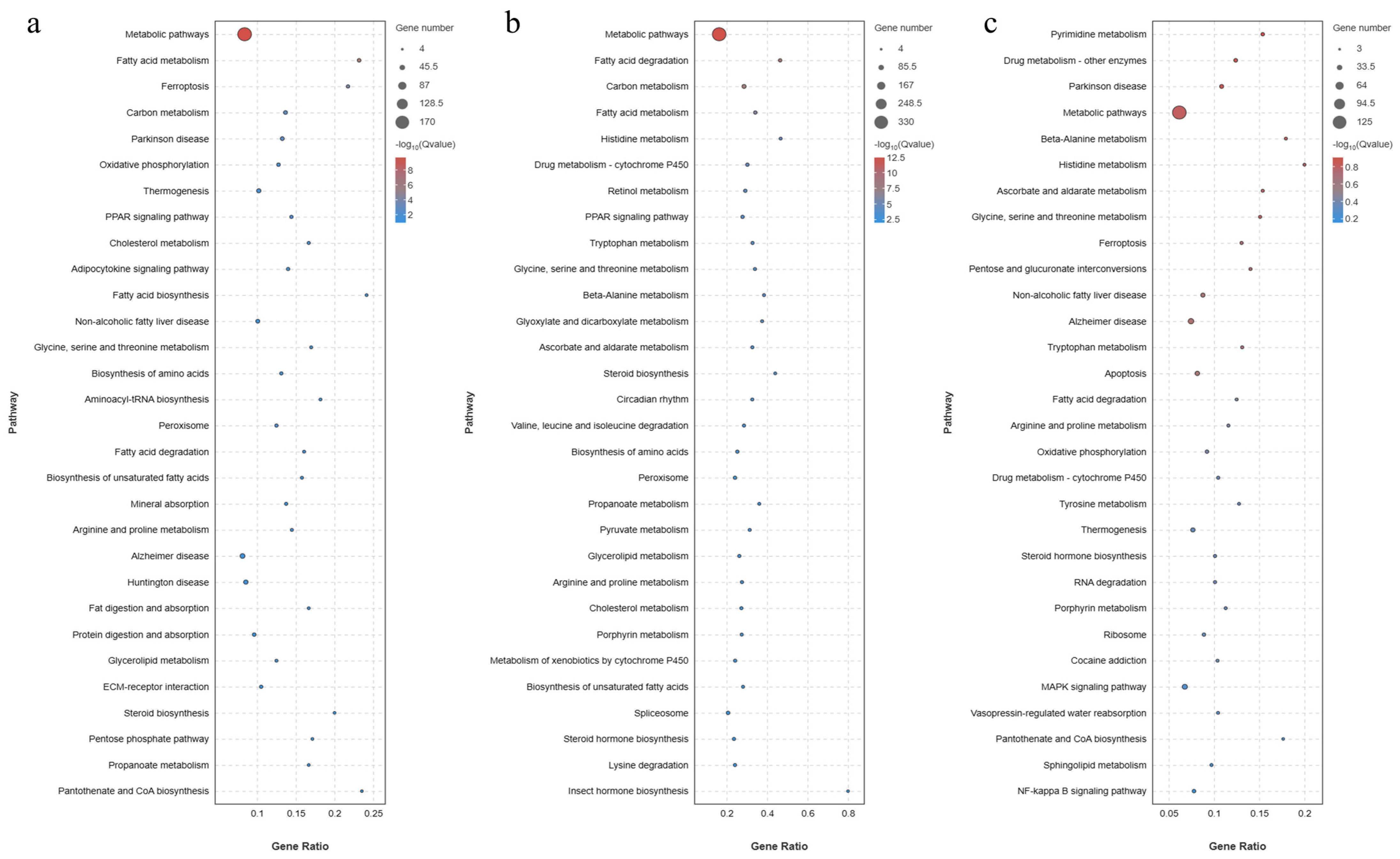
3.10.3. GO Enrichment and KEGG Enrichment Analysis of DEGs
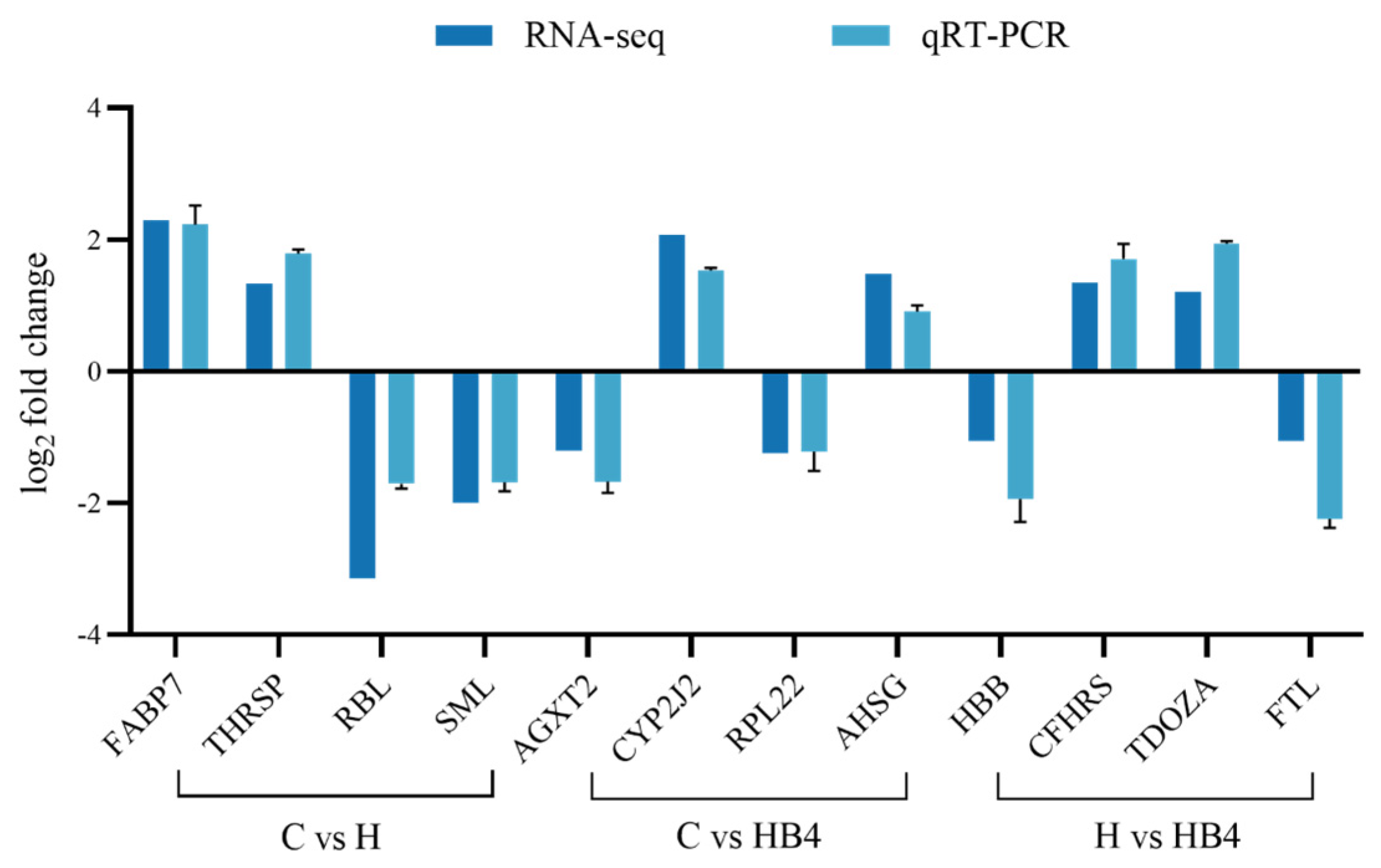
3.10.4. qPCR Verification
3.11. Challenge Test
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Elzaki, R.M. Does Fish Production Influence the GDP and Food Security in Gulf Cooperation Council Countries? Evidence from the Dynamic Panel Data Analysis. Aquaculture 2024, 578, 740058. [Google Scholar] [CrossRef]
- Zhao, L.; Liao, L.; Tang, X.; Liang, J.; Liu, Q.; Luo, W.; Adam, A.A.; Luo, J.; Li, Z.; Yang, S.; et al. High-Carbohydrate Diet Altered Conversion of Metabolites, and Deteriorated Health in Juvenile Largemouth Bass. Aquaculture 2022, 549, 737816. [Google Scholar] [CrossRef]
- He, Q.; Xiao, S.; Zhang, C.; Zhang, Y.; Shi, H.; Zhang, H.; Lin, F.; Liu, X.; Yang, H.; Wang, Q.; et al. Modulation of the Growth Performance, Biochemical Parameters, and Non-Specific Immune Responses of the Hybrid Grouper (Epinephelus fuscoguttatus ♀ × E. lanceolatus ♂) by Two Kinds of Chinese Herb. Aquac. Rep. 2021, 19, 100604. [Google Scholar] [CrossRef]
- Feng, M.; Luo, X.; Gu, C.; Li, Y.; Zhu, X.; Fei, J. Systematic Analysis of Berberine-Induced Signaling Pathway be tween miRNA Clusters and mRNAs and Identification of Mir-99a~125b Cluster Function by Seed-Targeting Inhibitors in Multiple Myeloma Cells. RNA Biol. 2015, 12, 82–91. [Google Scholar] [CrossRef]
- Yin, J.; Ye, J.; Jia, W. Effects and Mechanisms of Berberine in Diabetes Treatment. Acta Pharm. Sin. B 2012, 2, 327–334. [Google Scholar] [CrossRef]
- Zhou, M.; Deng, Y.; Liu, M.; Liao, L.; Dai, X.; Guo, C.; Zhao, X.; He, L.; Peng, C.; Li, Y. The Pharmacological Activity of Berberine, a Review for Liver Protection. Eur. J. Pharmacol. 2021, 890, 173655. [Google Scholar] [CrossRef]
- Pan, H.; Li, Z.; Xie, J.; Liu, D.; Wang, H.; Yu, D.; Zhang, Q.; Hu, Z.; Shi, C. Berberine Influences Blood Glucose via Modulating the Gut Microbiome in Grass Carp. Front. Microbiol. 2019, 10, 1066. [Google Scholar] [CrossRef]
- Meng, X.; Wu, S.; Hu, W.; Zhu, Z.; Yang, G.; Zhang, Y.; Qin, C.; Yang, L.; Nie, G. Clostridium butyricum Improves Immune Responses and Remodels the Intestinal Microbiota of Common Carp (Cyprinus carpio L.). Aquaculture 2021, 530, 735753. [Google Scholar] [CrossRef]
- Nowland, T.; Plush, K.; Barton, M.; Kirkwood, R. Development and Function of the Intestinal Microbiome and Potential Implications for Pig Production. Animals 2019, 9, 76. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Han, L.; Haidar Abbas Raza, S.; Yue, Q.; Sun, S.; Zhao, Y.; Lv, L.; Deng, Y.; Yuan, Z.; Alsharif, I.; et al. Polysaccharides in Berberis Dasystachya Improve Intestinal Flora Depending on the Molecular Weight and Ameliorate Type 2 Diabetes in Rats. J. Funct. Foods 2023, 100, 105381. [Google Scholar] [CrossRef]
- Jia, B.; Park, D.; Hahn, Y.; Jeon, C.O. Metagenomic Analysis of the Human Microbiome Reveals the Association between the Abundance of Gut Bile Salt Hydrolases and Host Health. Gut Microbes 2020, 11, 1300–1313. [Google Scholar] [CrossRef] [PubMed]
- Xiong, F.; Wu, S.-G.; Zhang, J.; Jakovlić, I.; Li, W.-X.; Zou, H.; Li, M.; Wang, G.-T. Dietary Bile Salt Types Influence the Composition of Biliary Bile Acids and Gut Microbiota in Grass Carp. Front. Microbiol. 2018, 9, 2209. [Google Scholar] [CrossRef]
- Chiang, J.Y.L. Bile Acids: Regulation of Synthesis. J. Lipid Res. 2009, 50, 1955–1966. [Google Scholar] [CrossRef]
- Wahlström, A.; Sayin, S.I.; Marschall, H.-U.; Bäckhed, F. Intestinal Crosstalk between Bile Acids and Microbiota and Its Impact on Host Metabolism. Cell Metab. 2016, 24, 41–50. [Google Scholar] [CrossRef]
- Collins, S.L.; Stine, J.G.; Bisanz, J.E.; Okafor, C.D.; Patterson, A.D. Bile Acids and the Gut Microbiota: Metabolic Interactions and Impacts on Disease. Nat. Rev. Microbiol. 2023, 21, 236–247. [Google Scholar] [CrossRef]
- Lin, S.; Wang, S.; Wang, P.; Tang, C.; Wang, Z.; Chen, L.; Luo, G.; Chen, H.; Liu, Y.; Feng, B.; et al. Bile Acids and Their Receptors in Regulation of Gut Health and Diseases. Prog. Lipid Res. 2023, 89, 101210. [Google Scholar] [CrossRef]
- Li, K.; Liu, L.; Clausen, J.H.; Lu, M.; Dalsgaard, A. Management Measures to Control Diseases Reported by Tilapia (Oreochromis spp.) and Whiteleg Shrimp (Litopenaeus vannamei) Farmers in Guangdong, China. Aquaculture 2016, 457, 91–99. [Google Scholar] [CrossRef]
- Chi, S.; Zhou, Q.; Yang, Q. Advances in nutrition research of tilapia. Feed Res. 2004, 9, 9–14. [Google Scholar] [CrossRef]
- Doan, H.V.; Hoseinifar, S.H.; Jaturasitha, S.; Dawood, M.A.O.; Harikrishnan, R. The Effects of Berberine Powder Supplementation on Growth Performance, Skin Mucus Immune Response, Serum Immunity, and Disease Resistance of Nile Tilapia (Oreochromis Niloticus) Fingerlings. Aquaculture 2020, 520, 734927. [Google Scholar] [CrossRef]
- Tian, J.; Jin, Y.; Yu, E.; Sun, J.; Xia, Y.; Zhang, K.; Li, Z.; Gong, W.; Wang, G.; Xie, J. Intestinal Farnesoid X Receptor Mediates the Effect of Dietary Berberine on Lipid Accumulation in Grass Carp (Ctenopharyngodon idella). Aquaculture 2022, 553, 738055. [Google Scholar] [CrossRef]
- Lee, M.H. Official and standardized methods of analysis. Trends Food Sci. Technol. 1996, 6, 382–383. [Google Scholar]
- Liu, Y.; Zhou, H.; Fan, J.; Huang, H.; Deng, J.; Tan, B. Low Methyl-Esterified Pectin Induces Abnormal Hepatic Lipid Deposition in Largemouth Bass, Micropterus Salmoides. Aquaculture 2023, 563, 738958. [Google Scholar] [CrossRef]
- Pan, L.; Li, W.; Xie, R.; Liu, H.; Tan, B.; Dong, X.; Yang, Q.; Chi, S.; Zhang, S. Effects of Clostridium butyricum on Growth Performance, Intestinal Health, and Disease Resistance of Hybrid Grouper (Epinephelus fuscoguttatus ♀ × E. lanceolatus ♂) Fed with Cottonseed Protein Concentrate (CPC) Replacement of Fishmeal. Aquac. Nutrition 2016, 457, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Li, W.; Liu, H.; Tan, B.; Dong, X.; Chi, S.; Yang, Q.; Zhang, S.; Fan, D.; Hu, Y. The Isolation, Identification, Whole-Genome Sequencing of Clostridium butyricum LV1 and Its Effects on Growth Performance, Immune Response, and Disease-Resistance of Litopenaeus vannamei. Microbiol. Res. 2023, 272, 127384. [Google Scholar] [CrossRef]
- Chen, Y.J.; Zhang, T.Y.; Chen, H.Y.; Lin, S.M.; Luo, L.; Wang, D.S. An evaluation of hepatic glucose metabolism at the transcription level for the omnivorous GIFT tilapia, Oreochromis niloticus during postprandial nutritional status transition from anabolism to catabolism. Aquaculture 2017, 473, 375–382. [Google Scholar] [CrossRef]
- Zhou, N.-N.; Wang, T.; Lin, Y.-X.; Xu, R.; Wu, H.-X.; Ding, F.-F.; Qiao, F.; Du, Z.-Y.; Zhang, M.-L. Uridine Alleviates High-Carbohydrate Diet-Induced Metabolic Syndromes by Activating Sirt1/AMPK Signaling Pathway and Promoting Glycogen Synthesis in Nile Tilapia (Oreochromis niloticus). Anim. Nutr. 2023, 14, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Yuan, K.; Shang, S.; Guo, Y. A Safer Hypoglycemic Agent for Type 2 Diabetes—Berberine Organic Acid Salt. J. Funct. Foods 2017, 38, 399–408. [Google Scholar] [CrossRef]
- Demirci, M.; Bahar Tokman, H.; Taner, Z.; Keskin, F.E.; Çağatay, P.; Ozturk Bakar, Y.; Özyazar, M.; Kiraz, N.; Kocazeybek, B.S. Bacteroidetes and Firmicutes Levels in Gut Microbiota and Effects of Hosts TLR2/TLR4 Gene Expression Levels in Adult Type 1 Diabetes Patients in Istanbul, Turkey. J. Diabetes Complicat. 2020, 34, 107449. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Zhu, X.; Zeng, M.; Qi, L.; Tang, X.; Wang, D.; Zhang, M.; Xie, Y.; Li, H.; Yang, X.; et al. A Diet High in Sugar and Fat Influences Neurotransmitter Metabolism and Then Affects Brain Function by Altering the Gut Microbiota. Transl. Psychiatry 2021, 11, 328. [Google Scholar] [CrossRef]
- Guo, Y.; Zhang, Y.; Huang, W.; Selwyn, F.P.; Klaassen, C.D. Dose-Response Effect of Berberine on Bile Acid Profile and Gut Microbiota in Mice. BMC Complement. Altern. Med. 2016, 16, 394. [Google Scholar] [CrossRef]
- Xia, X.; Yan, J.; Shen, Y.; Tang, K.; Yin, J.; Zhang, Y.; Yang, D.; Liang, H.; Ye, J.; Weng, J. Berberine Improves Glucose Metabolism in Diabetic Rats by Inhibition of Hepatic Gluconeogenesis. PLoS ONE 2011, 6, e16556. [Google Scholar] [CrossRef] [PubMed]
- Dempsey, J.L.; Wang, D.; Siginir, G.; Fei, Q.; Raftery, D.; Gu, H.; Yue Cui, J. Pharmacological Activation of PXR and CAR Downregulates Distinct Bile Acid-Metabolizing Intestinal Bacteria and Alters Bile Acid Homeostasis. Toxicol. Sci. 2019, 168, 40–60. [Google Scholar] [CrossRef] [PubMed]
- de Vos, W.M.; Tilg, H.; Van Hul, M.; Cani, P.D. Gut Microbiome and Health: Mechanistic Insights. Gut 2022, 71, 1020–1032. [Google Scholar] [CrossRef] [PubMed]
- Winston, J.A.; Theriot, C.M. Diversification of Host Bile Acids by Members of the Gut Microbiota. Gut Microbes 2020, 11, 158–171. [Google Scholar] [CrossRef]
- Gurung, M.; Li, Z.; You, H.; Rodrigues, R.; Jump, D.B.; Morgun, A.; Shulzhenko, N. Role of Gut Microbiota in Type 2 Diabetes Pathophysiology. EBioMedicine 2020, 51, 102590. [Google Scholar] [CrossRef]
- Sun, L.; Xie, C.; Wang, G.; Wu, Y.; Wu, Q.; Wang, X.; Liu, J.; Deng, Y.; Xia, J.; Chen, B.; et al. Gut Microbiota and Intestinal FXR Mediate the Clinical Benefits of Metformin. Nat. Med. 2018, 24, 1919–1929. [Google Scholar] [CrossRef]
- Sun, R.; Yang, N.; Kong, B.; Cao, B.; Feng, D.; Yu, X.; Ge, C.; Huang, J.; Shen, J.; Wang, P.; et al. Orally Administered Berberine Modulates Hepatic Lipid Metabolism by Altering Microbial Bile Acid Metabolism and the Intestinal FXR Signaling Pathway. Mol. Pharmacol. 2017, 91, 110–122. [Google Scholar] [CrossRef]
- Lefebvre, P.; Cariou, B.; Lien, F.; Kuipers, F.; Staels, B. Role of Bile Acids and Bile Acid Receptors in Metabolic Regulation. Physiol. Rev. 2009, 89, 147–191. [Google Scholar] [CrossRef]
- Xu, G.; Li, H.; Pan, L.; Shang, Q.; Honda, A.; Ananthanarayanan, M.; Erickson, S.K.; Shneider, B.L.; Shefer, S.; Bollineni, J.; et al. FXR-Mediated down-Regulation of CYP7A1 Dominates LXRα in Long-Term Cholesterol-Fed NZW Rabbits. J. Lipid Res. 2003, 44, 1956–1962. [Google Scholar] [CrossRef]
- Sb, B.; Cr, K. From Mice to Men: Insights into the Insulin Resistance Syndromes. Annu. Rev. Physiol. 2006, 68, 123–158. [Google Scholar] [CrossRef]
- Sun, X.; Zhang, Y.; Cheng, G.; Zhu, T.; Zhang, Z.; Xiong, L.; Hu, H.; Liu, H. Berberine Improves DSS-Induced Colitis in Mice by Modulating the Fecal-Bacteria-Related Bile Acid Metabolism. Biomed. Pharmacother. 2023, 167, 115430. [Google Scholar] [CrossRef]
- Wood, I.S.; Trayhurn, P. Glucose Transporters (GLUT and SGLT): Expanded Families of Sugar Transport Proteins. Br. J. Nutr. 2003, 89, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Xu, R.; Qiao, F.; Du, Z.-Y.; Zhang, M.-L. Effects of Mannan Oligosaccharides (MOS) on Glucose and Lipid Metabolism of Largemouth Bass (Micropterus Salmoides) Fed with High Carbohydrate Diet. Anim. Feed Sci. Technol. 2022, 292, 115449. [Google Scholar] [CrossRef]
- Dayal, R.; Srivastava, P.P.; Lakra, W.S.; Chowdhary, S.; Yadav, K.; Bhatnagar, A. Histopathological Alteration in the Livers of Striped Snakehead Murrel (Channa Striatus) Grow-out Fed with Different Dietary Fats. Eur. J. Exp. Biol. 2013, 3, 359–364. [Google Scholar]
- Gergely, P. Effect of Glucopyranosylidene-Spiro-Thiohydantoin on Glycogen Metabolism in Liver Tissues of Streptozotocin-Induced and Obese Diabetic Rats. Mol. Med. Rep. 2011, 4, 477–481. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, K.; Zhou, J.; Sullivan, M.A.; Liu, Y.; Gilbert, R.G.; Deng, B. Metformin and Berberine Suppress Glycogenolysis by Inhibiting Glycogen Phosphorylase and Stabilizing the Molecular Structure of Glycogen in Db/Db Mice. Carbohydr. Polym. 2020, 243, 116435. [Google Scholar] [CrossRef] [PubMed]
- Alberto González-Regueiro, J.; Moreno-Castañeda, L.; Uribe, M.; Carlos Chávez-Tapia, N. The Role of Bile Acids in Glucose Metabolism and Their Relation with Diabetes. Ann. Hepatol. 2017, 16, S15–S20. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Zhang, Z.; Ding, Q.; Yang, Y.; Bindelle, J.; Ran, C.; Zhou, Z. Intestinal Cetobacterium and Acetate Modify Glucose Homeostasis via Parasympathetic Activation in Zebrafish. Gut Microbes 2021, 13, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Jiang, X.; Liu, N.; Ren, M.; Wang, Z.; Li, M.; Chen, N.; Li, S. Effects of Dietary Berberine Hydrochloride Inclusion on Growth, Antioxidant Capacity, Glucose Metabolism and Intestinal Microbiome of Largemouth Bass (Micropterus salmoides). Aquaculture 2022, 552, 738023. [Google Scholar] [CrossRef]
- Tarnecki, A.M.; Burgos, F.A.; Ray, C.L.; Arias, C.R. Fish Intestinal Microbiome: Diversity and Symbiosis Unrav elled by Metagenomics. J. Appl. Microbiol. 2017, 123, 2–17. [Google Scholar] [CrossRef]
- Liao, X.; Wang, C.; Wang, B.; Qin, H.; Hu, S.; Wang, P.; Sun, C.; Zhang, S. Comparative Transcriptome Analysis of Litopenaeus vannamei Reveals That Triosephosphate Isomerase-Like Genes Play an Important Role During Decapod Iridescent Virus 1 Infection. Front. Immunol. 2020, 11, 1904. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Liu, K.; Tan, B.; Liu, H.; Dong, X.; Yang, Q.; Chi, S.; Zhang, S.; Wang, H. Transcriptome, Enzyme Activity and Histopathology Analysis Reveal the Effects of Dietary Carbohydrate on Glycometabolism in Juvenile Largemouth Bass, Micropterus Salmoides. Aquaculture 2019, 504, 39–51. [Google Scholar] [CrossRef]
- Liao, X.; Cheng, L.; Xu, P.; Lu, G.; Wachholtz, M.; Sun, X.; Chen, S. Transcriptome Analysis of Crucian Carp (Carassius auratus), an Important Aquaculture and Hypoxia-Tolerant Species. PLoS ONE 2013, 8, e62308. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Tong, Y.; Ren, Z.; Chen, S.; Mei, X.; Zhou, Q.; Dai, G. Hypoglycemic Mechanisms of Polygonatum sibiricum Polysaccharide in Db/Db Mice via Regulation of Glycolysis/Gluconeogenesis Pathway and Alteration of Gut Microbiota. Heliyon 2023, 9, e15484. [Google Scholar] [CrossRef] [PubMed]
- Vaz, F.M.; Ferdinandusse, S. Bile Acid Analysis in Human Disorders of Bile Acid Biosynthesis. Mol. Asp. Med. 2017, 56, 10–24. [Google Scholar] [CrossRef]
- Xu, X.; Gao, Z.; Yang, F.; Yang, Y.; Chen, L.; Han, L.; Zhao, N.; Xu, J.; Wang, X.; Ma, Y.; et al. Antidiabetic Effects of Gegen Qinlian Decoction via the Gut Microbiota Are Attributable to Its Key Ingredient Berberine. Genom. Proteom. Bioinform. 2020, 18, 721–736. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F. A Review on the Application of Herbal Medicines in the Disease Control of Aquatic Animals. Aquaculture 2020, 526, 735422. [Google Scholar] [CrossRef]
- Cao, J.; Liu, Z.; Zhang, D.; Guo, F.; Gao, F.; Wang, M.; Yi, M.; Lu, M. Distribution and Localization of Streptococcus agalactiae in Different Tissues of Artificially Infected Tilapia (Oreochromis niloticus). Aquaculture 2022, 546, 737370. [Google Scholar] [CrossRef]
- Sifa, L.; Chenhong, L.; Dey, M.; Gagalac, F.; Dunham, R. Cold Tolerance of Three Strains of Nile Tilapia, Oreo chromis Niloticus, in China. Aquaculture 2002, 213, 123–129. [Google Scholar] [CrossRef]
- Chen, Q.-Q.; Liu, W.-B.; Zhou, M.; Dai, Y.-J.; Xu, C.; Tian, H.-Y.; Xu, W.-N. Effects of Berberine on the Growth and Immune Performance in Response to Ammonia Stress and High-Fat Dietary in Blunt Snout Bream Megalobrama Amblycephala. Fish Shellfish Immunol. 2016, 55, 165–172. [Google Scholar] [CrossRef]
























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | C | H | H1 | H2 | H3 | H4 |
|---|---|---|---|---|---|---|
| white fishmeal | 25 | 25 | 25 | 25 | 25 | 25 |
| casein | 16 | 16 | 16 | 16 | 16 | 16 |
| gelatin | 4 | 4 | 4 | 4 | 4 | 4 |
| corn starch | 25 | 40 | 40 | 40 | 40 | 40 |
| paste starch | 4 | 4 | 4 | 4 | 4 | 4 |
| fish oil | 5 | 5 | 5 | 5 | 5 | 5 |
| soybean lecithin | 1 | 1 | 1 | 1 | 1 | 1 |
| vitamin C | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 |
| Premix a | 1 | 1 | 1 | 1 | 1 | 1 |
| Ca(H2PO4)2 | 2 | 2 | 2 | 2 | 2 | 2 |
| choline chloride | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 |
| ethoxyquin | 0.03 | 0.03 | 0.03 | 0.03 | 0.03 | 0.03 |
| attractant | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 |
| berberine | 0 | 0 | 0.0025 | 0.005 | 0.0075 | 0.01 |
| microcrystalline cellulose | 16.27 | 1.27 | 1.2675 | 1.265 | 1.2625 | 1.26 |
| total | 100 | 100 | 100 | 100 | 100 | 100 |
| crude protein | 40.91 | 41.67 | 40.89 | 40.61 | 40.63 | 41.53 |
| crude lipid | 6.94 | 6.14 | 6.25 | 6.61 | 5.89 | 5.66 |
| ash | 7.33 | 7.27 | 7.34 | 7.4 | 7.33 | 7.34 |
| carbohydrate | 27.38 | 41.05 | 40.96 | 41.39 | 40.31 | 40.91 |
| Gene | Sequence 5′-3′ | Reference/Accession No. |
|---|---|---|
| HK-F | CAGCACGGAACTCCATGATGACC | (Yong-Jun Chen, 2017) [25] |
| HK-R | GCACAAATGTGGGCAGCATCTTG | (Yong-Jun Chen, 2017) [25] |
| PFKl-F | GCCGCCTTCAACCTGGTTAAGA | (Yong-Jun Chen, 2017) [25] |
| PFKl-R | GTGCCGCAGAAGTCGTTGTCTA | (Yong-Jun Chen, 2017) [25] |
| G6PC-F | ATCGGAGACTGGCTCAACTTGGT | (Yong-Jun Chen, 2017) [25] |
| G6PC-R | TGGCATGACCTGAAGGACTTCCT | (Yong-Jun Chen, 2017) [25] |
| PCK-F | CGAGTGGAGAGCAAGACTGTGA | (Yong-Jun Chen, 2017) [25] |
| PCK-R | TTGGTGAGAGCTGAGCCTACG | (Yong-Jun Chen, 2017) [25] |
| GYS-F | TGACAAGGAGGCTGGTGAGAGG | (Yong-Jun Chen, 2017) [25] |
| GYS-R | ACTCGTGCATGGCTGAGAACTT | (Yong-Jun Chen, 2017) [25] |
| GLUT2-F | GGCACTCTAGCTCTGGCTGTGT | (Yong-Jun Chen, 2017) [25] |
| GLUT2-R | GGGTGGTGACCTGGGTCTTCTT | (Yong-Jun Chen, 2017) [25] |
| CYP7A1-F | GGGATAAGACACAGGCAACCA | XM_003456729.5 |
| CYP7A1-R | TGCGGAGGAATTGAAGTGGG | XM_003456729.5 |
| CYP8B1-F | GGGTAAAGAGGAGATGGGAATG | XM_019348226.2 |
| CYP8B1-R | GGAGCAGCCAGAATGAAGAA | XM_019348226.2 |
| FXR-F | AACCATCCTGACACCAGATCG | XM_025899550.1 |
| FXR-R | CAACAGAGGCTGGGAAAGGA | XM_025899550.1 |
| βactin | CAGTGCCCATCTACGAG | (Yong-Jun Chen, 2017) [25] |
| βactin | CCATCTCCTGCTCGAAGTC | (Yong-Jun Chen, 2017) [25] |
| Group | C | H | HB1 | HB2 | HB3 | HB4 | Linear | Quadratic | ANOVA |
|---|---|---|---|---|---|---|---|---|---|
| SR (%) | 98.90 ± 1.91 | 97.80 ± 1.91 | 97.80 ± 1.91 | 95.56 ± 1.96 | 94.46 ± 6.93 | 93.33 ± 6.65 | 0.118 | 0.307 | 0.566 |
| WGR (%) | 1486.08 ± 16.57 | 1433.87 ± 35.70 | 1435.85 ± 30.36 | 1448.69 ± 5.36 | 1495.85 ± 17.22 | 1501.53 ± 8.65 | 0.001 | 0.004 | 0.021 |
| SGR (%) | 5.03 ± 0.02 | 4.96 ± 0.06 | 4.97 ± 0.04 | 4.98 ± 0.01 | 5.04 ± 0.02 | 5.37 ± 0.34 | 0.075 | 0.095 | 0.332 |
| FCR | 0.97 ± 0.01 | 1.00 ± 0.03 | 1.00 ± 0.02 | 0.99 ± 0.00 | 0.96 ± 0.01 | 0.96 ± 0.00 | 0.002 | 0.008 | 0.031 |
| VSI (%) | 13.38 ± 0.67 | 11.55 ± 0.57 | 14.18 ± 1.50 | 14.35 ± 0.80 | 14.10 ± 0.48 | 13.87 ± 0.45 | 0.135 | 0.060 | 0.229 |
| HIS (%) | 1.56 ± 0.07 | 1.79 ± 0.20 | 2.21 ± 0.15 * | 2.12 ± 0.09 * | 1.74 ± 0.06 | 1.36 ± 0.10 | 0.042 | 0.001 | 0.003 |
| CF (g/cm3) | 3.67 ± 0.05 | 3.96 ± 0.05 | 3.57 ± 0.15 | 3.63 ± 0.08 | 3.83 ± 0.24 | 3.72 ± 0.05 | 0.645 | 0.399 | 0.305 |
| Group | C | H | HB1 | HB2 | HB3 | HB4 | Linear | Quadratic | ANOVA |
|---|---|---|---|---|---|---|---|---|---|
| sobs | 788.67 ± 107.29 | 544.67 ± 2.52 | 511.00 ± 70.15 * | 447.67 ± 28.59 ** | 483.00 ± 67.01 * | 490.33 ± 93.29 * | 0.238 | 0.198 | 0.001 |
| Chao 1 | 854.87 ± 79.71 | 611.64 ± 17.51 * | 619.00 ± 65.14 ** | 514.86 ± 15.39 ** | 568.23 ± 55.88 ** | 564.34 ± 74.53 ** | 0.275 | 0.186 | 0.000 |
| ace | 903.58 ± 57.91 | 592.33 ± 5.66 ** | 612.82 ± 46.76 ** | 515.50 ± 11.15 ** | 558.20 ± 66.89 ** | 571.58 ± 60.36 ** | 0.553 | 0.199 | 0.000 |
| Shannon | 4.97 ± 1.23 | 3.63 ± 0.63 | 3.21 ± 0.66 | 3.22 ± 0.85 | 3.61 ± 1.19 | 4.23 ± 0.44 | 0.183 | 0.238 | 0.195 |
| Simpson | 0.86 ± 0.08 | 0.80 ± 0.06 | 0.74 ± 0.07 | 0.71 ± 0.14 | 0.75 ± 0.18 | 0.86 ± 0.05 | 0.325 | 0.338 | 0.435 |
| Genes | Pathway | Gene ID | Log2 (fc) |
|---|---|---|---|
| C-vs-H | |||
| Pgm1 | Glycolysis/Gluconeogenesis | ncbi_100534546 | 0.29 |
| ENO3 | Glycolysis/Gluconeogenesis | ncbi_100699677 | 0.46 |
| ENO1 | Glycolysis/Gluconeogenesis | ncbi_100701182 | 2.34 |
| ADH5 | Glycolysis/Gluconeogenesis | ncbi_100702001 | 0.51 |
| GPI | Glycolysis/Gluconeogenesis | ncbi_100704840 | 4.32 |
| G6PC | Glycolysis/Gluconeogenesis | ncbi_100709522 | 0.46 |
| Cyp46a1 | Primary bile acid biosynthesis | ncbi_100703511 | 7.92 |
| C-vs-HB4 | |||
| tpi1b | Glycolysis/Gluconeogenesis | ncbi_100534540 | 0.52 |
| Pgm1 | Glycolysis/Gluconeogenesis | ncbi_100534546 | 0.43 |
| PDHA1 | Glycolysis/Gluconeogenesis | ncbi_100691217 | −1.71 |
| Acss2 | Glycolysis/Gluconeogenesis | ncbi_100692076 | 2.60 |
| Aldh3a2 | Glycolysis/Gluconeogenesis | ncbi_100693723 | 0.57 |
| ADH5 | Glycolysis/Gluconeogenesis | ncbi_100695823 | −0.86 |
| ACSS2 | Glycolysis/Gluconeogenesis | ncbi_100697261 | 0.74 |
| Aldh3a2 | Glycolysis/Gluconeogenesis | ncbi_100698670 | 0.59 |
| ENO3 | Glycolysis/Gluconeogenesis | ncbi_100699677 | 0.47 |
| Pgk1 | Glycolysis/Gluconeogenesis | ncbi_100701118 | 0.74 |
| ENO1 | Glycolysis/Gluconeogenesis | ncbi_100701182 | 2.79 |
| ADH5 | Glycolysis/Gluconeogenesis | ncbi_100702001 | 0.57 |
| ADH5 | Glycolysis/Gluconeogenesis | ncbi_100702534 | −2.03 |
| ADH5 | Glycolysis/Gluconeogenesis | ncbi_100702802 | 0.34 |
| PFKM | Glycolysis/Gluconeogenesis | ncbi_100702997 | 0.38 |
| ALDH16A1 | Glycolysis/Gluconeogenesis | ncbi_100705854 | 1.08 |
| ALDH2 | Glycolysis/Gluconeogenesis | ncbi_100707184 | 0.51 |
| G6PC | Glycolysis/Gluconeogenesis | ncbi_100709522 | 0.76 |
| aldh9A1 | Glycolysis/Gluconeogenesis | ncbi_109194280 | 0.34 |
| aldh9A1 | Glycolysis/Gluconeogenesis | ncbi_109196929 | 0.47 |
| HSD3B7 | Primary bile acid biosynthesis | ncbi_100534450 | 0.52 |
| Cyp46a1 | Primary bile acid biosynthesis | ncbi_100692681 | 0.55 |
| CYP8B1 | Primary bile acid biosynthesis | ncbi_109195682 | 0.78 |
| H-vs-HB4 | |||
| ALDH16A1 | Glycolysis/Gluconeogenesis | ncbi_100705854 | 0.75 |
| aldh9A1 | Glycolysis/Gluconeogenesis | ncbi_109196929 | 0.51 |
| HSD3B7 | Primary bile acid biosynthesis | ncbi_100534450 | 0.79 |
| CYP8B1 | Primary bile acid biosynthesis | ncbi_100693750 | 0.49 |
| CYP7B1 | Primary bile acid biosynthesis | ncbi_100694909 | 1.48 |
| Hsd17b4 | Primary bile acid biosynthesis | ncbi_100706762 | 0.36 |
| AKR1D1 | Primary bile acid biosynthesis | ncbi_109194203 | 0.18 |
| ch25h | Primary bile acid biosynthesis | ncbi_109194213 | −0.41 |
| CYP8B1 | Primary bile acid biosynthesis | ncbi_109195682 | 0.78 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, H.; Wei, M.; Tan, B.; Dong, X.; Xie, S. The Supplementation of Berberine in High-Carbohydrate Diets Improves Glucose Metabolism of Tilapia (Oreochromis niloticus) via Transcriptome, Bile Acid Synthesis Gene Expression and Intestinal Flora. Animals 2024, 14, 1239. https://doi.org/10.3390/ani14081239
Liu H, Wei M, Tan B, Dong X, Xie S. The Supplementation of Berberine in High-Carbohydrate Diets Improves Glucose Metabolism of Tilapia (Oreochromis niloticus) via Transcriptome, Bile Acid Synthesis Gene Expression and Intestinal Flora. Animals. 2024; 14(8):1239. https://doi.org/10.3390/ani14081239
Chicago/Turabian StyleLiu, Hongyu, Menglin Wei, Beiping Tan, Xiaohui Dong, and Shiwei Xie. 2024. "The Supplementation of Berberine in High-Carbohydrate Diets Improves Glucose Metabolism of Tilapia (Oreochromis niloticus) via Transcriptome, Bile Acid Synthesis Gene Expression and Intestinal Flora" Animals 14, no. 8: 1239. https://doi.org/10.3390/ani14081239