4.1. Growth Performance, Energetics, and Carcass Traits
According to [
32], the primary impact of ZH on ruminant production is linked to the enhancement of both LW and the dressing carcass. This is attributed to an increase in muscle protein synthesis and retention, which occurs concurrently with the suppression of lipogenesis and an increase in lipolysis. ZH activates a biological mechanism that improves growth performance indicators and energy retention due to higher muscle tissue concentration compared to adipose tissue. This is confirmed by calculations that estimate the energy intake of the diet [
27,
29], indicating an apparent increase in the NEm, NEg, and DMI observed/expected ratios. Multiple studies have demonstrated the effectiveness of the use of ZH in lamb production. For instance, research conducted with ZH in lambs with LW, doses, and results similar to those of the present study, showed improvements in growth indicators [
33,
34,
35,
36]. These studies have reported an increase in ADG, feed efficiency, and energy retention provided by the diet (12.5–25.0%, 13.0–36.0%, and 9.5–35.0%, respectively). Similarly, the results on carcass quality traits from some authors [
34,
37,
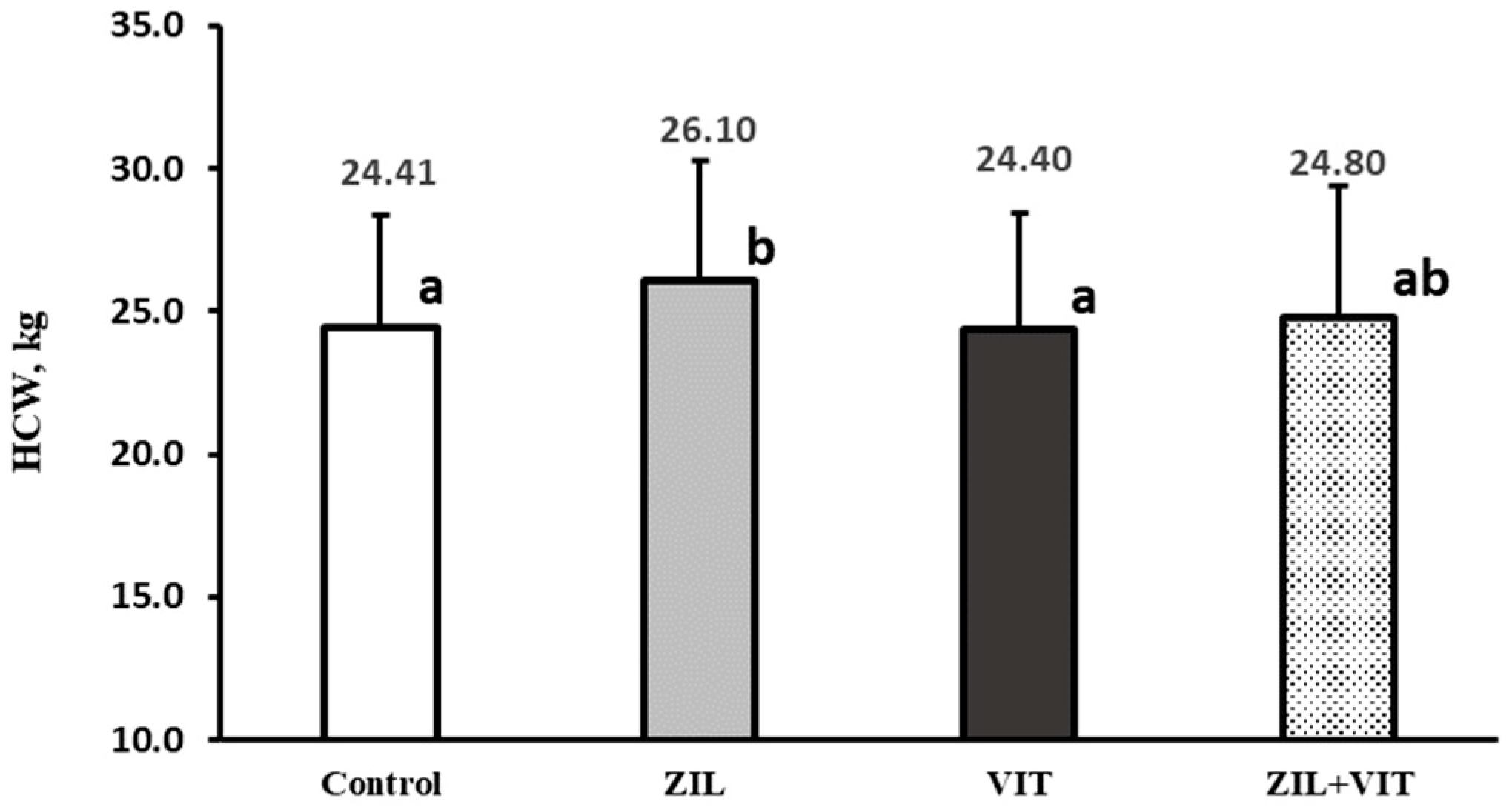
38] coincide with those presented in our study. These studies indicate that the effects of ZH supplementation are observed in relation to carcass quality, such as an increase in the HCW (7.0–11.2%), dressing carcass (2.0–8.8%), LTM area (11.2–15.4%), and a reduction in perirenal–pelvic fat (21.7–26.1%).
Vitamin D
3 acts as a precursor of 1,25-dihydroxycholecalciferol, a hormone involved in calcium and phosphorus homeostasis [
39]. Previous studies have shown that feeding supra-nutritional doses of D3 in ruminants can cause hypercalcemia and reduced DMI and body weight [
22]. In a study by [
22], steers were supplemented with 5 × 10
6 IU of D3 for 10 d during the ZH supplementation stage (21 d), resulting in a significant decrease in ADG, DMI, and feed efficiency by 47.6%, 11.5%, and 52.0%, respectively. Similarly, [
16] reductions in ADG and feed efficiency of up to 25.1% and 23.0% were observed, respectively, when testing the supplementation of 5 × 10
6 IU of D3 in steers for 24 d. Our findings align with these results, where administering D3 led to an 18.1% reduction in ADG and a 25.9% reduction in feed efficiency over a period of 29 d, although the overall DMI was not affected. However, DMI during the last 7 d was reduced by 15.7%.
Studies with supra-nutritional doses of D3 supplementation were carried out in short periods before slaughter. This approach should ensure that the carcass quality is not compromised by DMI reduction due to D3 supplementation. However, some studies suggest that there may be a slight impact on the quality of the carcass. Studies have shown that supplementing D3 in steers and lambs before slaughter can result in a significant reduction in weight and fat thickness. Authors [
22] found that supplementing 5 × 10
6 IU of D3 for 10 d led to a reduction in HCW and back fat thickness by 3.8% and 16.3%, respectively. Studies [
40] reported that supplementing 6 million IU of D3 for 4–6 d prior to slaughter resulted in a reduction in FLW by 4.7%, HCW by 5.3%, and back fat thickness by 19.1%. Similarly, [
41] observed a 19.0% reduction in adjusted fat thickness in lambs when supplementing 7.5 × 10
6 IU of D3 for 4 d before slaughter, without any changes in dressing carcass and LTM area.
On the contrary, a study conducted by [
16] observed a 20.2% increase in back fat thickness in steer carcasses when feeding 5 × 10
6 IU of D3, without affecting HCW and dressing carcass. However, providing high doses of D3 may lead to a reduction in DMI, which can reduce the supply of nutrients for body maintenance and result in catabolism of fat tissue. This can ultimately lead to a reduction in the USDA yield grade. Nonetheless, other studies [
16] did not show this pattern of results when administering D3. Although the negative effect of D3 on DMI, ADG, and feed efficiency is an expected response, ZH supplementation could compensate for this effect. The negative interaction is the result of the inability of ZH to compensate for the reduction in DMI. It seems that ZH cannot have a favorable response in growth performance when the energy available in the diet is reduced due to low DMI.
4.2. Meat Quality
Meat tenderness is a crucial quality trait that affects consumer acceptability, satisfaction, and repeat purchase [
42]. Muscle hypertrophy brought about by ZH supplementation consequently increases the toughness of the meat. Various factors can contribute to an increase in WBSF values due to ZH supplementation. One reason for this is the alteration to the activity of the myofibrillar enzyme system, which may occur due to decreased calpain concentrations and increased calpastatin activity [
10,
18,
43]. Research has also shown that an increase in muscle fiber diameter and hypertrophy [
12] and a decrease in intramuscular fat levels [
13] can raise WBSF values. The calpain–calpastatin proteolytic system has been identified as the key factor that determines meat tenderness.
However, some authors suggest that aging can help mitigate the negative impact of meat toughness [
44]. The effectiveness of the aging strategy in improving the WBSF of muscles treated with ZH is not consistent. ZH has been shown to increase WBSF in cattle, which has been widely documented [
8]. Supplementing with ZH for 20 d resulted in tougher meat (7.0–21.4%) in the LTM across different periods of aging. Other studies [
45,
46] have reported similar increases in WBSF (8.8–29.8%) in the LTM with ZH supplementation ranging from 20 to 40 d. The impact of ZH on muscle WBSF in lambs is variable. According to studies by [
34,
35], supplementing with ZH for 30 d can increase the toughness of the LTM by 20.0 to 56.4%, which is consistent with our research showing a 21.5% increase in WBSF. However, other reports by [
34,
47] have not found any differences in WBSF between lambs treated with ZH and the control group.
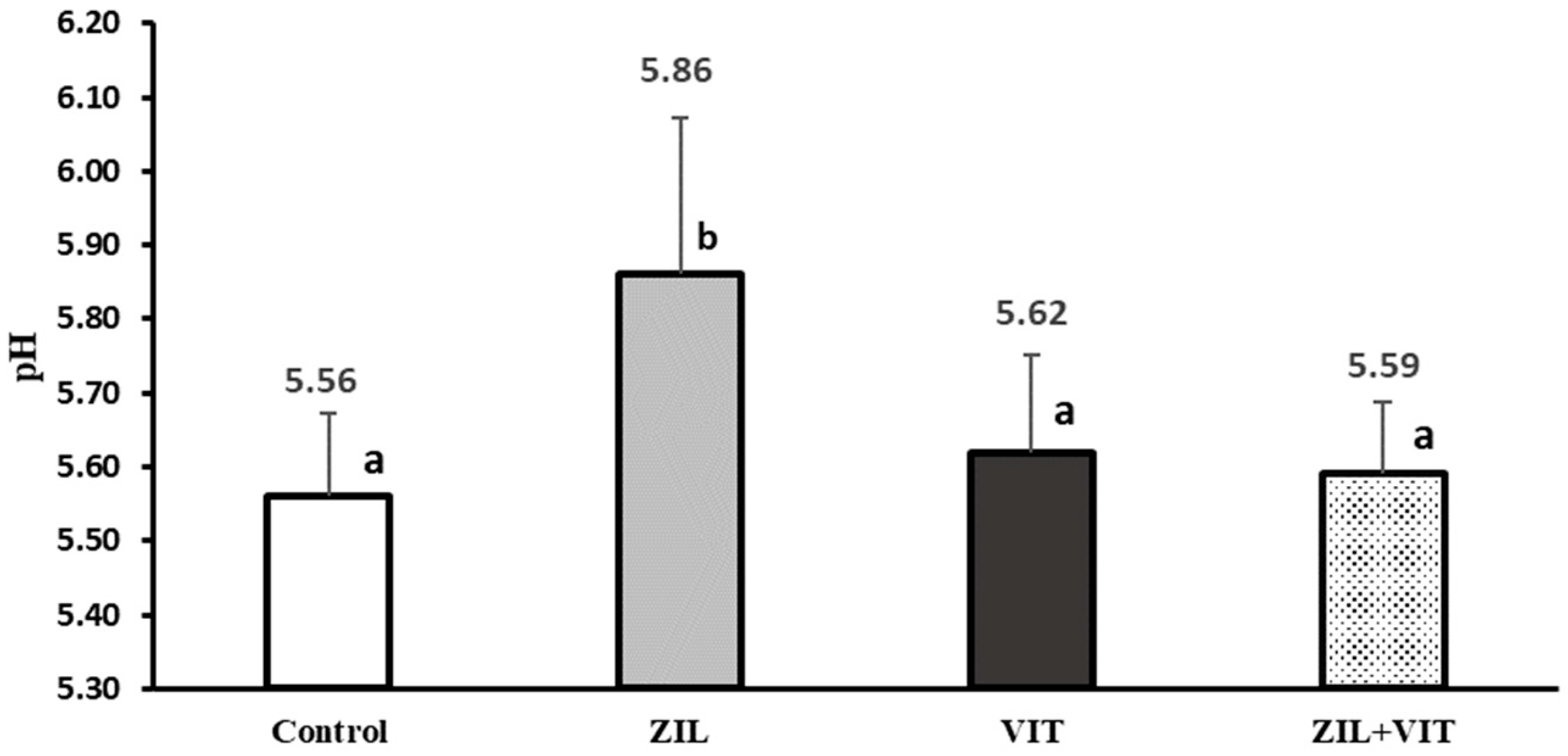
The final pH of the meat is a crucial factor in determining its quality, and lambs supplemented with ZH have been observed to produce meat with an elevated pH of 3.2 to 8.9% [
31]. Our experiment’s results align with this last result. The findings on the color of lamb meat, when supplemented with ZH, are inconsistent. According to research [
34,
48], there were no discernible differences in the luminosity (L*), redness (a*), and yellowness (b*) color characteristics of the LTM after administering ZH for 28–30 d. However, other studies have reported a decrease in luminosity (L*) and redness (a*) by up to 13.8% and 24.2%, respectively. In the current experiment, ZH only resulted in a 7.0% decrease in luminosity without any impact on the redness of the meat.
A higher concentration of calcium in muscles is decisive for the function of this system, which is key to determining the postmortem quality characteristics of meat tenderness [
20]. The proteolytic system of the calpain–calpastatin muscle system is a calcium-dependent enzymatic system. Therefore, it increases the activity of calpain enzymes that degrade myofibrillar proteins [
20] and promotes meat tenderness [
17,
18,
19]. Vitamin D
3 supplemented before slaughter increases the concentration of intramuscular calcium [
16,
17,
18]; therefore, it could increase the activity of the calpain enzyme system and promote meat tenderness [
15]. Studies have shown that D3 supplementation can indeed increase meat tenderness [
16,
17,
18]. In this regard, studies on cattle, with D3 doses ranging from 1 to 7 million IU, have been shown to reduce the WBSF of the LTM [
18,
40]. They used supra-nutritional doses of D3 in the diet for a period ranging from 3 to 7 d before slaughter. Other authors [
8,
22] have reported that supplementing with 0.5–5 × 10
6 IU/d of D3 between 10 and 20 d before slaughter did not affect the WBSF values of meat, even with 14 to 35 d of aging. However, [
22] found that a treatment of 0.25 × 10
6 IU/d with D3 for 165 d before slaughter tended to produce tougher meat at 21 d of aging. The findings align with those reported by [
41] in their study on lambs supplemented with 750,000 IU/d of D3 for 4 d, which resulted in meat with 11.4% toughness. However, in the current experiment, no significant difference was observed in the WBSF of muscle samples collected from animals supplemented with D3 for 7 d before slaughter.
The water content of meat products is one of the essential quality characteristics; it also has a substantial influence on product quality, as a higher loss of water gives an expectation of a less optimal quality, which, if excessive, can have an adverse effect on product appearance [
49]. Studies with lambs supplemented with ZH showed increased water purge loss of meat between 20 and 44.0% [
34,
50]. Our experiment showed similar results; the WHC of lamb meat was reduced with ZH supplementation. Likewise, in the current study, meat samples from lambs treated with ZH (
Table 6) showed higher pH values and, conversely, luminosity (L*) and red intensity (a*) were lower (
Figure 5). Higher pH values in meat are associated with the supply of β-agonists, which may be due to lower deposition of muscle glycogen, affecting pH drops [
47].
If the final pH of the meat is high, the physical state of the proteins will be above their isoelectric point. Proteins will associate with more water in the muscle and, therefore, the fibers will be tight (less space between cells). This meat will appear to have lower luminosity (L*) due to the fact that its surface does not scatter light to the same extent as the more open surface of the meat with a lower final pH [
51]. The oxygen penetration depth and formation of oxymyoglobin (red myoglobin form) is dependent upon the partial pressure of oxygen and its ability to diffuse into the muscle structure. It is possible that changes in the structure and spacing of the muscle could alter this depth and, hence, the oxygen consumption rate. In high-pH dark muscle, the lack of spaces between cells and muscle bundles prevents oxygen diffusion into the tissue, and there is a greater demand for oxygen by myoglobin in the interior; hence, at the surface, there is less oxymyoglobin, which is observed as a reduction in redness (a*) values [
52].
Different studies have implemented D3 to mitigate the negative impact of meat toughness caused by ZH supplementation. Authors [
18,
22] have reported slight improvements in beef tenderness in cattle that were supplemented with ZH and given D3 at doses of 1 to 7 million IU per d for 3 to 6 d. Other studies [
46] in steers supplemented with 1–9 × 10
6 IU between 3 and 9 d before slaughter did not reduce meat toughness produced by ZH supplementation. ZH supplementation resulted in tougher meat in 7.0–21.4% of the LTM on different aging periods, and the supplementation of 0.5 × 10
6 IU D3 did not reduce the negative effect of zilpaterol. [
8] In our experiment, supplementing with 1.5 × 10
6 IU of D3 did not result in reduced WBSF values of the LTM of feedlot lambs with ZH supplementation, which is similar to the outcome mentioned earlier.
4.3. Fatty Acids
β-agonists have been studied for their effect on the fatty acid profile of ruminants. In a study by [
3,
47], lambs treated with ZH showed an increase in polyunsaturated fatty acids in intramuscular fat. In their study on lambs, [
53] found that administering Salbutamol, a β-agonist, led to a decrease in saturated fatty acids and an increase in total unsaturated fatty acids. Nonetheless, the research conducted by [
31] revealed that the addition of ZH to the diet of lambs did not result in any alteration to the fatty acid composition of the intramuscular fat. However, the findings of our current study demonstrated that supplementing lambs with ZH did not impact the overall levels of saturated or unsaturated fatty acids present in intramuscular fat but tended to reduce monounsaturated fatty acid (C16:1). The variation in the results obtained through the use of β-agonists in the fatty acid profile of meat can be attributed to various factors. These factors include genetic influence by breed, the type of β-adrenergic receptor that is stimulated (β
1 or β
2), and the concentration of β-adrenergic receptors that increase with age due to tissue development [
54]. Additionally, the composition of fatty acids in meat can be affected by factors, including the degree of fattening and the composition of the diet consumed by ruminants [
55].
Research with D3 in ruminants has focused on calcium metabolism and its effect on meat tenderness. Thus, research on the use of D3 in muscle fatty acid metabolism is scarce. In this sense, the fatty acid profile of meat is better related to vitamin E supplementation because of the oxidative stability of fats [
15]. However, studies in mice have shown that D3 deficiency plays an important role in the metabolism of fatty acids in the blood and liver [
56]. In addition, it has been observed that supplementation of D3 also stimulated the expression of genes promoting fatty acid oxidation, such as carnitine palmitoyl-transferase I α or β, medium- and long-chain acyl-CoA dehydrogenases, as well as the expression of the pyruvate dehydrogenase kinase 4 mRNA, which is known to encode an enzyme that favors the utilization of fatty acids over glucose [
57].
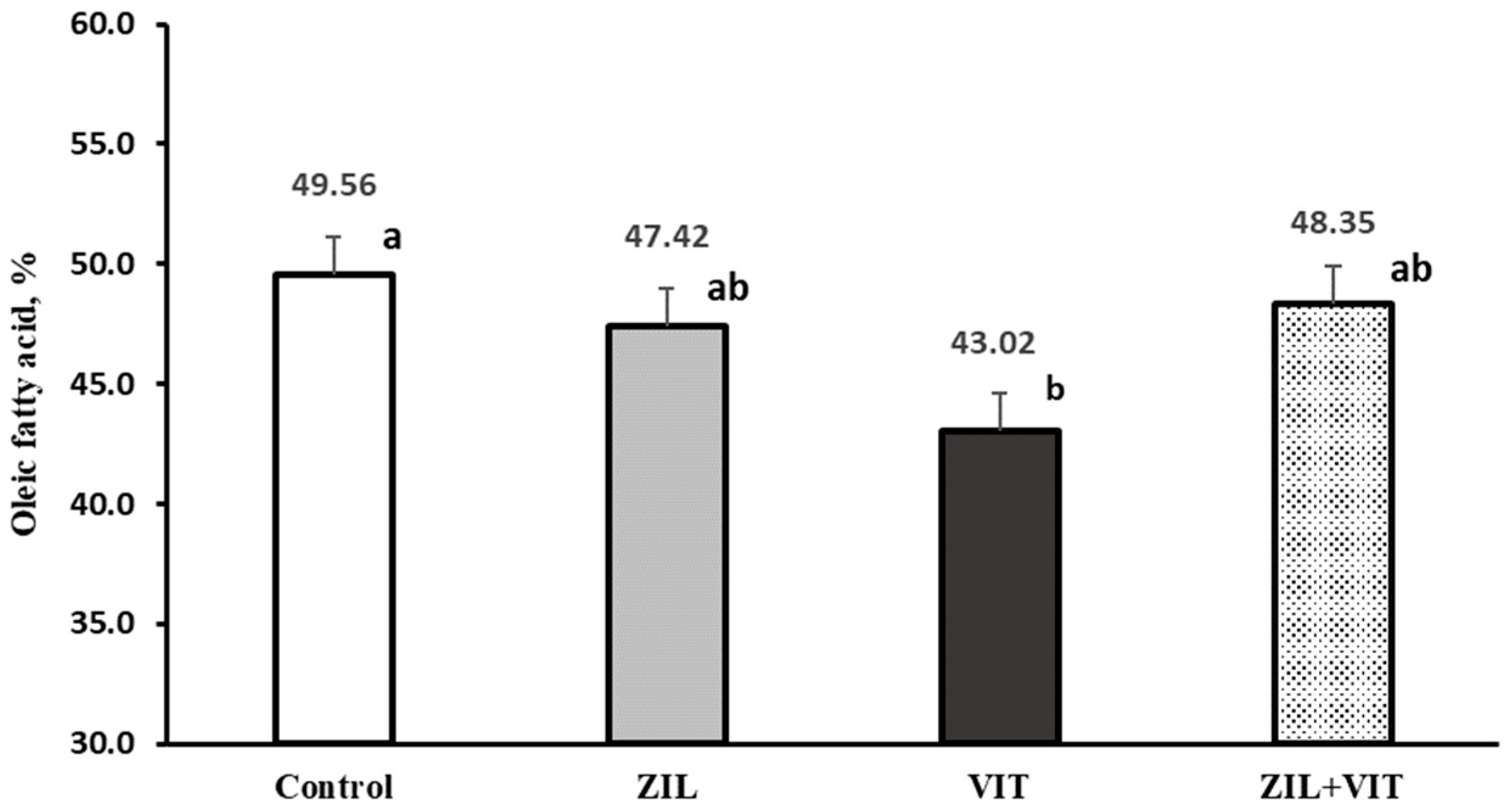
However, in the present experiment, it is observed (
Figure 8) that D3 supplementation results in a reduction in oleic fatty acid. Likewise, an interaction was observed, and a change in the metabolism of PUFAs and UFAs in intramuscular fat was noted when the lambs were supplemented with ZIL+VIT. The reason for this interaction is not clear. Probably, the negative effect of DMI by D3 produces this response, which may be altered by the lipid metabolism of ZH in muscle.

 ,
, 










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}