Hair Testosterone and Cortisol Concentrations in Pre- and Post-Rut Roe Deer Bucks: Correlations with Blood Levels and Testicular Morphometric Parameters

 ,
,  ,
,  and
and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Sampling
2.2. Testicular Analysis
2.3. Steroids Extraction from Plasma and Hair
2.4. Radioimmunoassay (RIA) for Testosterone and Cortisol
2.5. Statistical Analyses
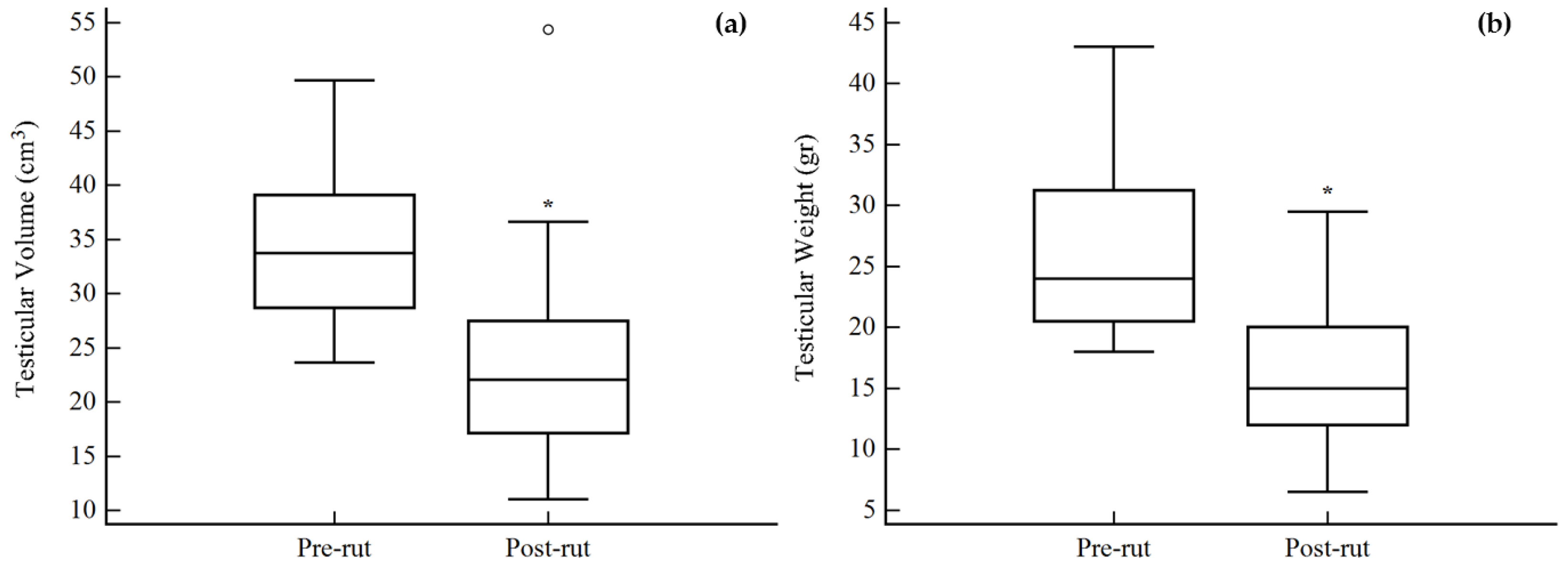
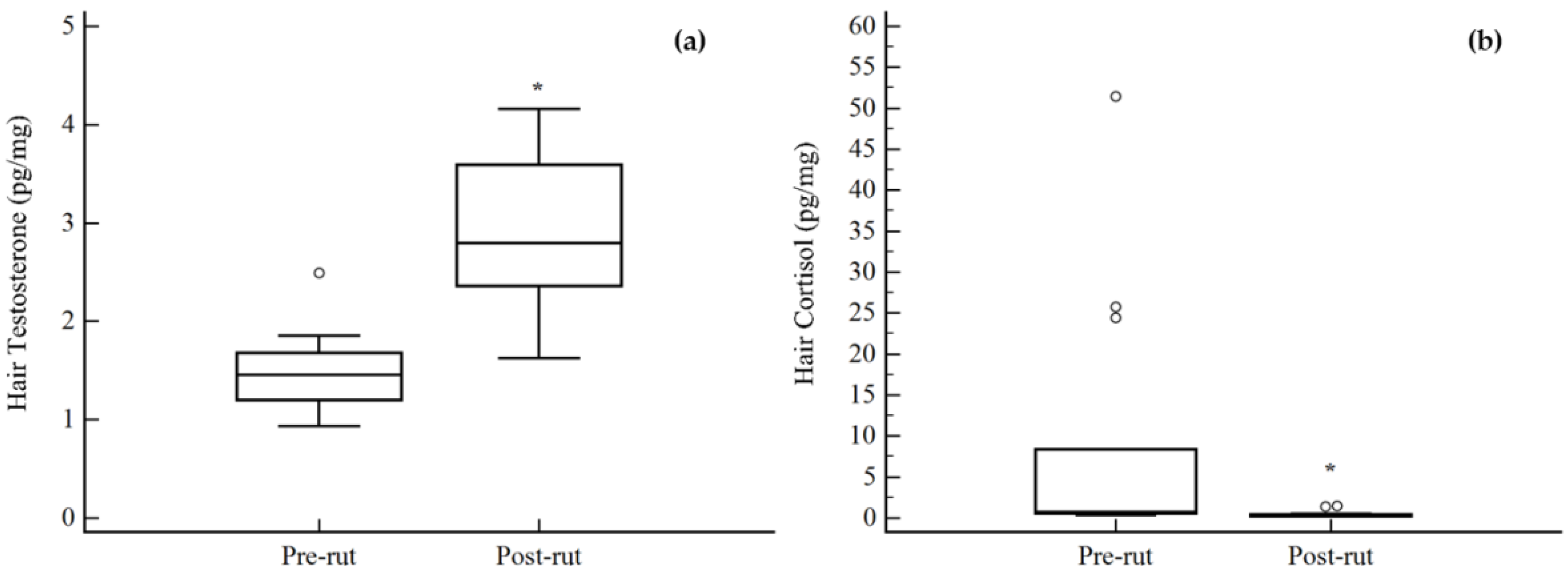
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Klonisch, T.; Schön, J.; Hombach-Klonisch, S.; Blottner, S. The roe deer as a model for studying seasonal regulation of testis function. Int. J. Androl. 2006, 29, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Goeritz, F.; Quest, M.; Wagener, A.; Fassbender, M.; Broich, A.; Hildebrandt, T.B.; Hofmann, R.R.; Blottner, S. Seasonal timing of sperm production in roe deer: Interrelationship among changes in ejaculate parameters, morphology and function of testis and accessory glands. Theriogenology 2003, 59, 1487–1502. [Google Scholar] [CrossRef]
- Hoem, S.A.; Melis, C.; Linnell, J.D.C.; Reidar, A. Fighting behaviour in territorial male roe deer Capreolus capreolus: The effects of antler size and residence. Eur. J. Wildl. Res. 2007, 53, 1–8. [Google Scholar] [CrossRef]
- Johnson, E.; Hornby, J. Seasonal changes of pelage in the roe deer Capreolus capreolus and its role in thermoregulation. J. Nat. Hist. 1975, 9, 619–628. [Google Scholar] [CrossRef]
- Roelants, H.; Schneider, F.; Göritz, F.; Streich, J.; Blottner, S. Seasonal changes of spermatogonial proliferation in roe deer, demonstrated by flow cytometric analysis of c-kit receptor, in relation to follicle-stimulating hormone, luteinizing hormone, and testosterone. Biol. Reprod. 2002, 66, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Blottner, S.; Hingst, O.; Meyer, H.H.D. Seasonal spermatogenesis and testosterone production in roe deer (Capreolus capreolus). J. Reprod. Fertil. 1996, 108, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Schön, J.; Göritz, F.; Streich, J.; Blottner, S. Histological organization of roe deer testis throughout the seasonal cycle: Variable and constant components of tubular and interstitial compartment. Anat. Embryol. (Berl.) 2004, 208, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Kozioł, K.; Koziorowski, M. Steroid hormones in peripheral blood plasma and androgen receptors in testis and epididymis of roe deer male (Capreolus capreolus) during the reproduction season. Small Rumin. Res. 2013, 115, 86–93. [Google Scholar] [CrossRef]
- Vera, F.; Zenuto, R.; Antenucci, C.D. Expanding the actions of cortisol and corticosterone in wild vertebrates: A necessary step to overcome the emerging challenges. Gen. Comp. Endocrinol. 2017, 246, 337–353. [Google Scholar] [CrossRef] [PubMed]
- Davenport, M.D.; Tiefenbacher, S.; Lutz, C.K.; Novak, M.A.; Meyer, J.S. Analysis of endogenous cortisol concentrations in the hair of rhesus macaques. Gen. Comp. Endocrinol. 2006, 147, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Sheriff, M.J.; Dantzer, B.; Delehanty, B.; Palme, R.; Boonstra, R. Measuring stress in wildlife: Techniques for quantifying glucocorticoids. Oecologia 2011, 166, 869–887. [Google Scholar] [CrossRef] [PubMed]
- Narayan, E.J. Evaluation of physiological stress in Australian wildlife: Embracing pioneering and current knowledge as a guide to future research directions. Gen. Comp. Endocrinol. 2017, 244, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Meyer, J.S.; Novak, M.A. Minireview: Hair Cortisol: A Novel Biomarker of Hypothalamic-Pituitary-Adrenocortical Activity. Endocrinology 2012, 153, 4120–4127. [Google Scholar] [CrossRef] [PubMed]
- Bacci, M.L.; Nannoni, E.; Govoni, N.; Scorrano, F.; Zannoni, A.; Forni, M.; Martelli, G.; Sardi, L. Hair cortisol determination in sows in two consecutive reproductive cycles. Reprod. Biol. 2014, 14, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Scorrano, F.; Carrasco, J.; Pastor-Ciurana, J.; Belda, X.; Rami-Bastante, A.; Bacci, M.L.; Armario, A. Validation of the long-term assessment of hypothalamic-pituitary-adrenal activity in rats using hair corticosterone as a biomarker. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2015, 29, 859–867. [Google Scholar] [CrossRef] [PubMed]
- Schell, C.J.; Young, J.K.; Lonsdorf, E.V.; Mateo, J.M.; Santymire, R.M. Investigation of techniques to measure cortisol and testosterone concentrations in coyote hair. Zoo Biol. 2017, 36, 220–225. [Google Scholar] [CrossRef] [PubMed]
- Sotohira, Y.; Suzuki, K.; Sano, T.; Arai, C.; Asakawa, M.; Hayashi, H. Stress assessment using hair cortisol of kangaroos affected by the lumpy jaw disease. J. Vet. Med. Sci. 2017, 79, 852–854. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.Y.; Kim, E.; Choi, M.H. Technical and clinical aspects of cortisol as a biochemical marker of chronic stress. BMB Rep. 2015, 48, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Stalder, T.; Kirschbaum, C. Analysis of cortisol in hair—State of the art and future directions. Brain Behav. Immun. 2012, 26, 1019–1029. [Google Scholar] [CrossRef] [PubMed]
- Dehnhard, M.; Clauss, M.; Lechner-Doll, M.; Meyer, H.H.D.; Palme, R. Noninvasive Monitoring of Adrenocortical Activity in Roe Deer (Capreolus capreolus) by Measurement of Fecal Cortisol Metabolites. Gen. Comp. Endocrinol. 2001, 123, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Wickings, E.J.; Nieschlag, E. Stability of testosterone and androstenedione in blood and plasma samples. Clin. Chim. Acta 1976, 71, 439–443. [Google Scholar] [CrossRef]
- Diver, M.J.; Hughes, J.G.; Hutton, J.L.; West, C.R.; Hipkin, L.J. The long-term stability in whole blood of 14 commonly-requested hormone analytes. Ann. Clin. Biochem. 1994, 31, 561–565. [Google Scholar] [CrossRef] [PubMed]
- Gaiani, R.; Chiesa, F.; Mattioli, M.; Nannetti, G.; Galeati, G. Androstenedione and testosterone concentrations in plasma and milk of the cow throughout pregnancy. J. Reprod. Fertil. 1984, 70, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Schams, D.; Barth, D. Annual profiles of reproductive hormones in peripheral plasma of the male roe deer (Capreolus capreolus). J. Reprod. Fertil. 1982, 66, 463–468. [Google Scholar] [CrossRef] [PubMed]
- Lightman, S.L.; Wiles, C.C.; Atkinson, H.C.; Henley, D.E.; Russell, G.M.; Leendertz, J.A.; McKenna, M.A.; Spiga, F.; Wood, S.A.; Conway-Campbell, B.L. The significance of glucocorticoid pulsatility. Eur. J. Pharmacol. 2008, 583, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Young, C.; Ganswindt, A.; McFarland, R.; de Villiers, C.; van Heerden, J.; Ganswindt, S.; Barrett, L.; Henzi, S.P. Faecal glucocorticoid metabolite monitoring as a measure of physiological stress in captive and wild vervet monkeys. Gen. Comp. Endocrinol. 2017, 253, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Hellhammer, J.; Fries, E.; Schweisthal, O.W.; Schlotz, W.; Stone, A.A.; Hagemann, D. Several daily measurements are necessary to reliably assess the cortisol rise after awakening: State and trait components. Psychoneuroendocrinology 2007, 32, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Bévalot, F.; Gaillard, Y.; Lhermitte, M.A.; Pépin, G. Analysis of corticosteroids in hair by liquid chromatography-electrospray ionization mass spectrometry. J. Chromatogr. B Biomed. Sci. Appl. 2000, 740, 227–236. [Google Scholar] [CrossRef]
- Lockwood, S.A.; Kattesh, H.G.; Rhinehart, J.D.; Strickland, L.G.; Krawczel, P.D.; Wilkerson, J.B.; Kirkpatrick, F.D.; Saxton, A.M. Relationships among temperament, acute and chronic cortisol and testosterone concentrations, and breeding soundness during performance testing of Angus bulls. Theriogenology 2017, 89, 140–145. [Google Scholar] [CrossRef] [PubMed]
- Boswell, T.; Woods, S.C.; Kenagy, G.J. Seasonal Changes in Body Mass, Insulin, and Glucocorticoids of Free-Living Golden-Mantled Ground Squirrels. Gen. Comp. Endocrinol. 1994, 96, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Zatra, Y.; Aknoun-Sail, N.; Kheddache, A.; Benmouloud, A.; Charallah, S.; Moudilou, E.N.; Exbrayat, J.-M.; Khammar, F.; Amirat, Z. Seasonal changes in plasma testosterone and cortisol suggest an androgen mediated regulation of the pituitary adrenal axis in the Tarabul’s gerbil Gerbillus tarabuli (Thomas, 1902). Gen. Comp. Endocrinol. 2017. [CrossRef] [PubMed]
- Handa, R.J.; Weiser, M.J. Gonadal steroid hormones and the hypothalamo–pituitary–adrenal axis. Front. Neuroendocrinol. 2014, 35, 197–220. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Parameter | Pre-Rut | Post-Rut | ||||
|---|---|---|---|---|---|---|
| Mean (SD) | Min–Max | N | Mean (SD) | Min–Max | n | |
| Testicular Weight g | 26.2 (7.5) | 18.0–43.0 | 12 | 16.6 (6.9) | 6.5–29.5 | 10 |
| Testicular Volume cm3 | 34.63 (7.14) | 23.62–49.65 | 12 | 24.72 (12.04) | 11.01–54.36 | 11 |
| Blood TEST ng/mL | 2.26 (1.42) | 0.88–4.59 | 8 | 0.15 (0.09) | 0.02–0.28 | 8 |
| Blood CORT ng/mL | 1.79 (2.57) | 0.41–8.10 | 8 | 3.32 (5.44) | 0.01–16.32 | 8 |
| Hair TEST pg/mg | 1.48 (0.39) | 0.93–2.49 | 14 | 2.89 (0.77) | 1.62–4.16 | 14 |
| Hair CORT pg/mg | 8.72 (15.07) | 0.36–51.47 | 14 | 0.49 (0.42) | 0.21–1.51 | 14 |
| Hair TEST | Hair CORT | Blood TEST | Blood CORT | Testicular Volume | Testicular Weight | |
|---|---|---|---|---|---|---|
| Hair TEST | −0.56 p = 0.0019 | −0.75 p = 0.0007 | −0.07 p = 0.8115 | −0.58 p = 0.0040 | −0.57 p = 0.0072 | |
| Hair CORT | 0.40 p = 0.1287 | 0.33 p = 0.2168 | 0.14 p = 0.5264 | 0.26 p = 0.2500 | ||
| Blood TEST | 0.07 p = 0.8075 | 0.72 p = 0.0037 | 0.76 p = 0.0044 | |||
| Blood CORT | −0.18 p = 0.5468 | 0.00 p = 0.9914 | ||||
| Testicular Volume | 0.96 p < 0.001 | |||||
| Testicular Weight |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ventrella, D.; Elmi, A.; Barone, F.; Carnevali, G.; Govoni, N.; Bacci, M.L. Hair Testosterone and Cortisol Concentrations in Pre- and Post-Rut Roe Deer Bucks: Correlations with Blood Levels and Testicular Morphometric Parameters. Animals 2018, 8, 113. https://doi.org/10.3390/ani8070113
Ventrella D, Elmi A, Barone F, Carnevali G, Govoni N, Bacci ML. Hair Testosterone and Cortisol Concentrations in Pre- and Post-Rut Roe Deer Bucks: Correlations with Blood Levels and Testicular Morphometric Parameters. Animals. 2018; 8(7):113. https://doi.org/10.3390/ani8070113
Chicago/Turabian StyleVentrella, Domenico, Alberto Elmi, Francesca Barone, Giacomo Carnevali, Nadia Govoni, and Maria Laura Bacci. 2018. "Hair Testosterone and Cortisol Concentrations in Pre- and Post-Rut Roe Deer Bucks: Correlations with Blood Levels and Testicular Morphometric Parameters" Animals 8, no. 7: 113. https://doi.org/10.3390/ani8070113
APA StyleVentrella, D., Elmi, A., Barone, F., Carnevali, G., Govoni, N., & Bacci, M. L. (2018). Hair Testosterone and Cortisol Concentrations in Pre- and Post-Rut Roe Deer Bucks: Correlations with Blood Levels and Testicular Morphometric Parameters. Animals, 8(7), 113. https://doi.org/10.3390/ani8070113