Litter Survival Differences between Divergently Selected Lines for Environmental Sensitivity in Rabbits


Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Animals
2.2. Traits
2.3. Statistical Analysis
3. Results
3.1. Correlated Response to Selection in Litter Survival and Pre-Weaning Weight
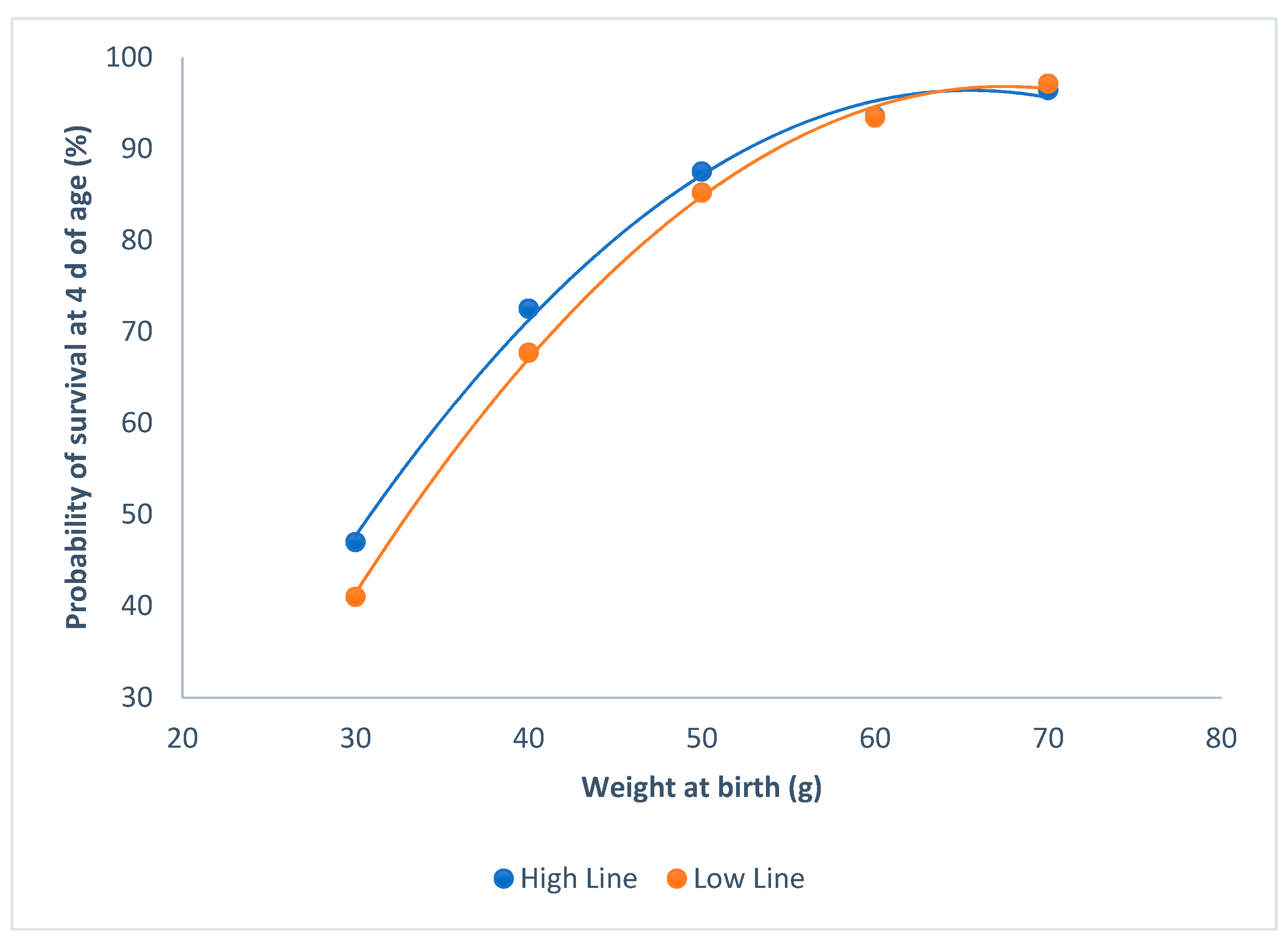
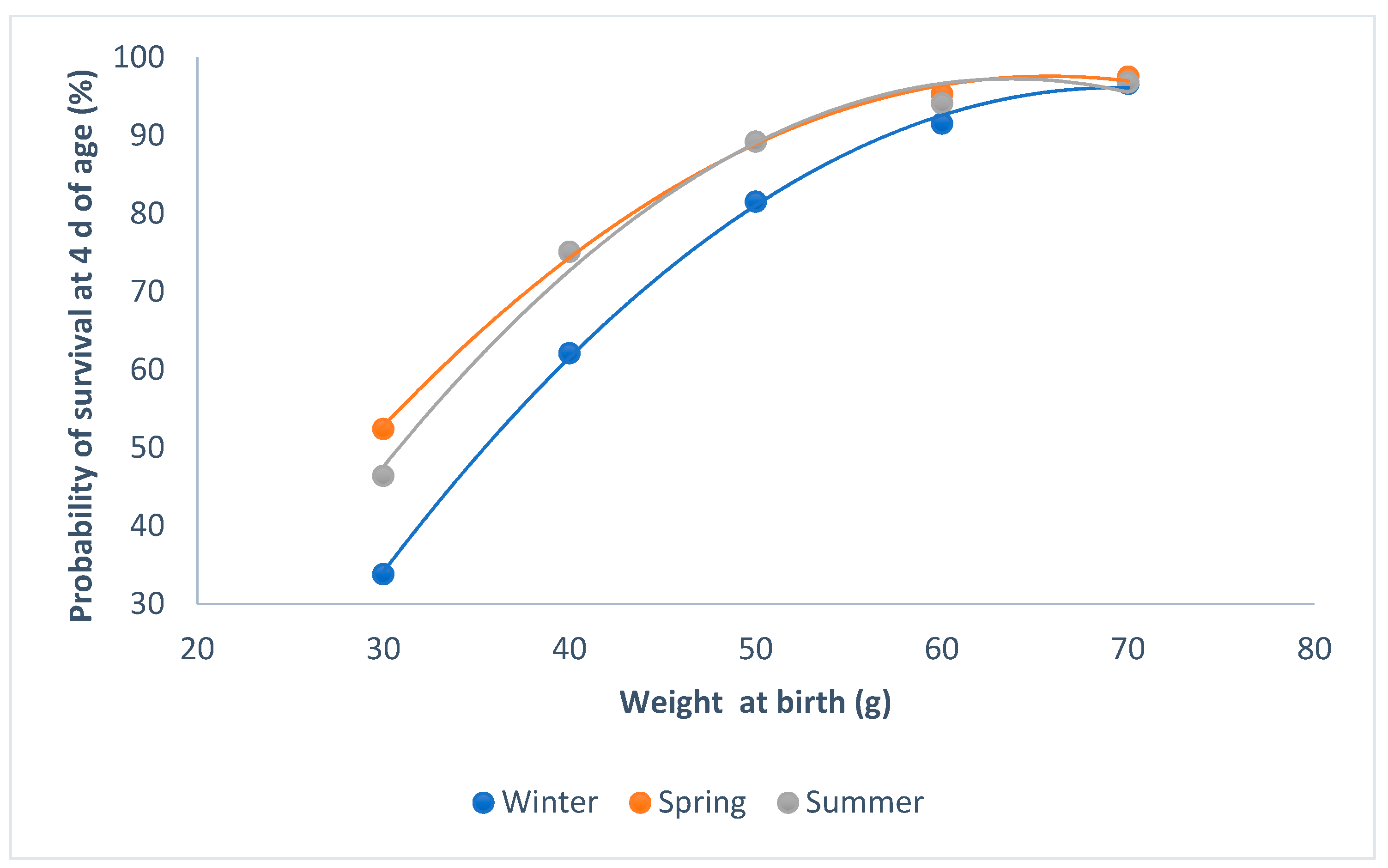
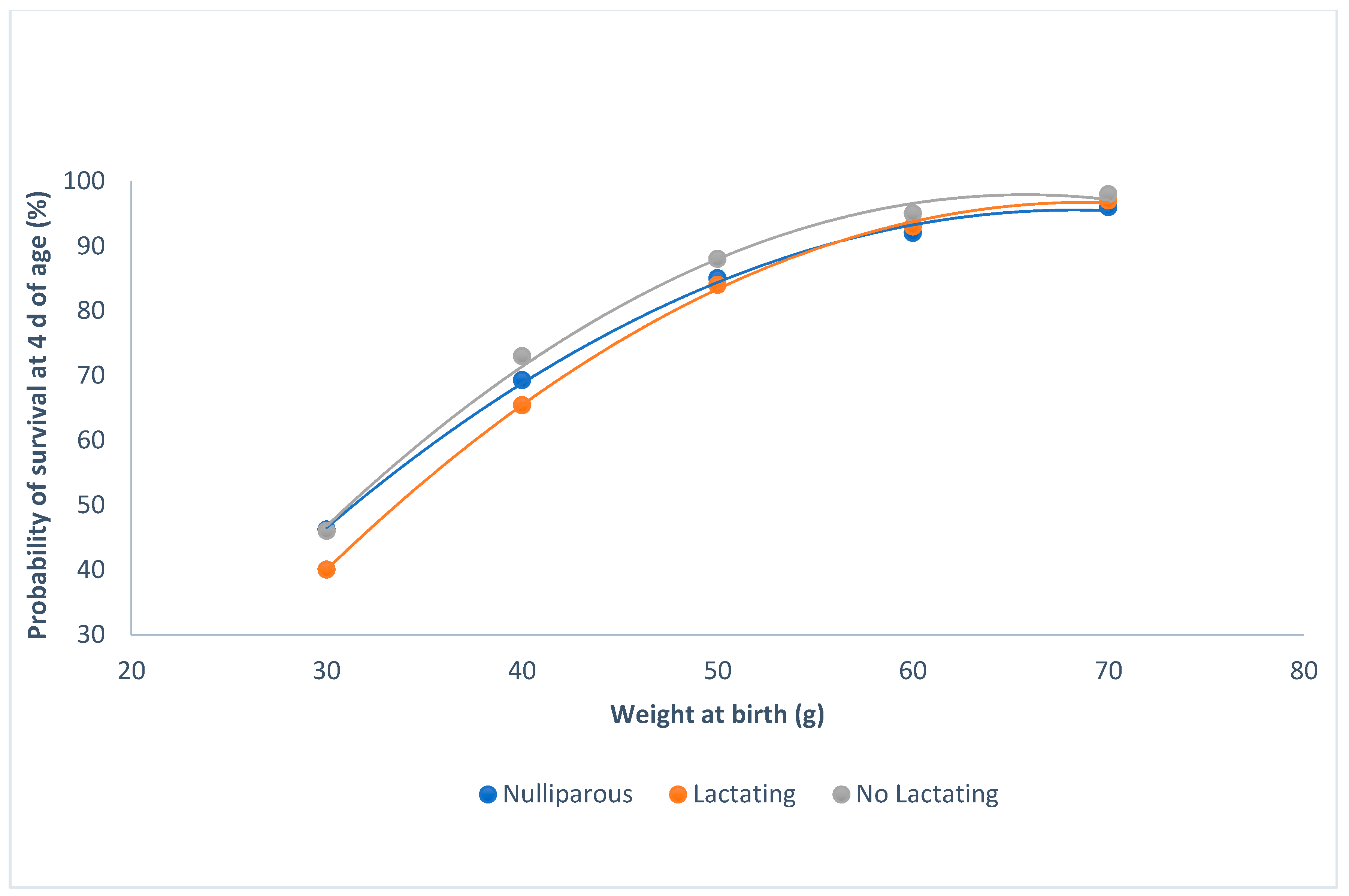
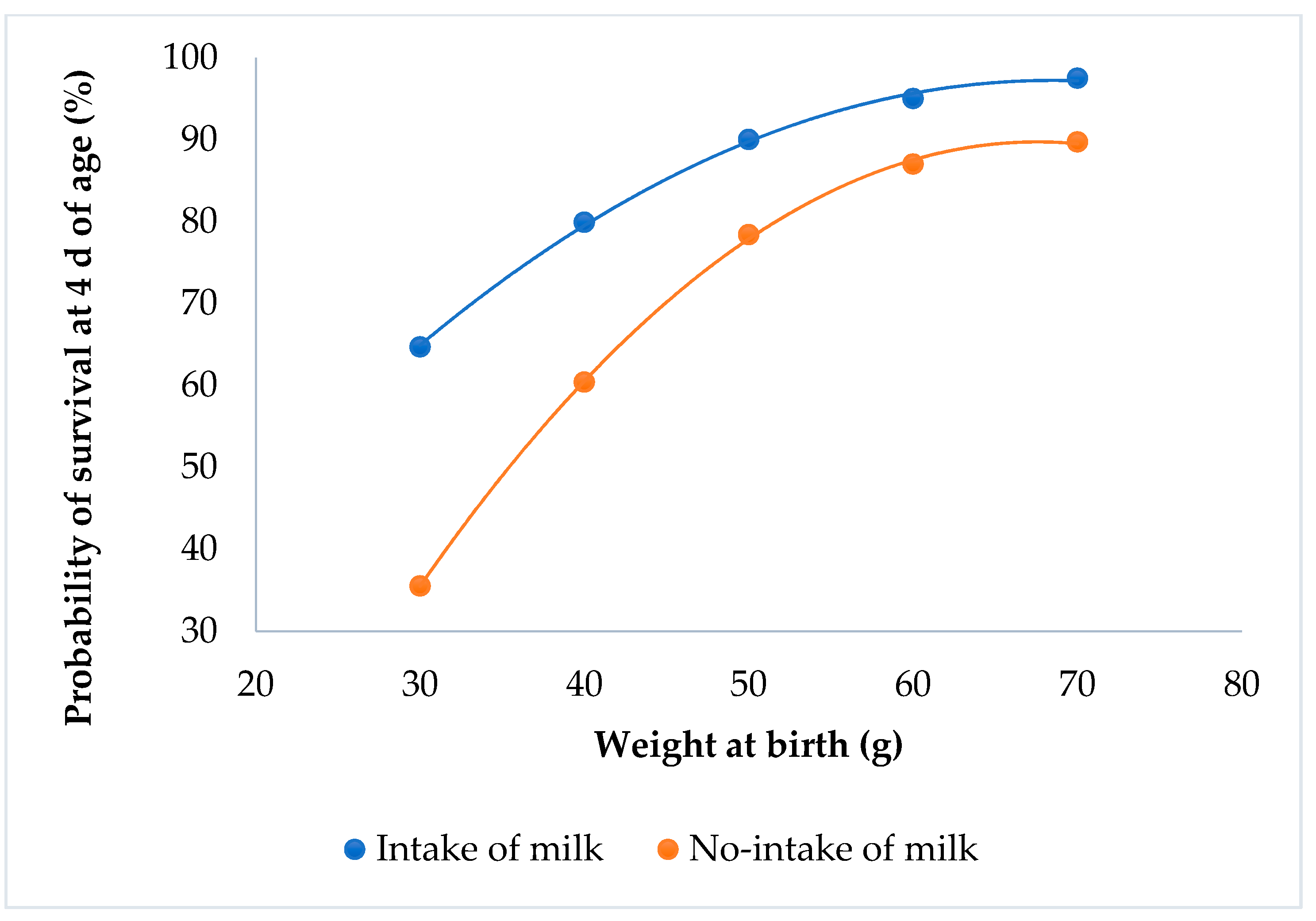
3.2. Survival at 4 d of Age and Individual Weight at Birth
4. Discussion
4.1. Correlated Response to Selection in Litter Survival and Pre-Weaning Weight
4.2. Survival at 4 d of Age and Individual Weight at Birth
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- García, M.; Baselga, M. Estimation of genetic response to selection in litter size of rabbits using a cryopreserved control population. Livest. Prod. Sci. 2002, 74, 45–53. [Google Scholar] [CrossRef]
- Sánchez, J.P.; Theilgaard, P.; Mínguez, C.; Baselga, M. Constitution and evaluation of a long-lived productive rabbit line. J. Anim. Sci. 2008, 86, 515–525. [Google Scholar] [CrossRef] [PubMed]
- Larzul, C.; Ducrocq, V.; Tudela, F.; Juin, H.; Garreau, H. The length of productive life can be modified through selection: An experimental demonstration in the rabbit. J. Anim. Sci. 2014, 92, 2395–2401. [Google Scholar] [CrossRef] [PubMed]
- Rauw, W.M.; Kanis, E.; Noordhuizen-Stassen, E.N.; Grommers, F.J. Undesirable side effects of selection for high production efficiency in farm animals: A review. Livest. Prod. Sci. 1998, 56, 15–33. [Google Scholar] [CrossRef]
- Rosell, J.; De La Fuente, L. Culling and mortality in breeding rabbits. Prev. Vet. Med. 2009, 88, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Blasco, A.; Martínez-Álvaro, M.; García, M.L.; Ibáñez-Escriche, N.; Argente, M.J. Selection for genetic environmental sensitivity of litter size in rabbits. Genet. Sel. Evol. 2017, 49, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Argente, M.J.; García, M.L.; Zbyňovská, K.; Petruška, P.; Capcarová, M.; Blasco, A. Correlated response to selection for litter size environmental variability in rabbits’ resilience. Animal 2019. [Google Scholar] [CrossRef]
- Bolet, G.; Esparbié, J.; Falieres, J. Relations entre le nombre de foetus par corne utérine, la taille de portée à la naissance et la croissance pondérale des lapereaux. Ann. Zootech. 1996, 45, 185–200. [Google Scholar] [CrossRef]
- Poignier, J.; Szendrö, Z.S.; Levai, A.; Radnai, I.; Biro-Nemeth, E. Effect of birth weight and litter size on growth and mortality in rabbit. World Rabbit Sci. 2000, 8, 103–109. [Google Scholar] [CrossRef]
- Blasco, A.; Blasco, P.D.A. Bayesian Data Analysis for Animal Scientists; Springer International Publishing: New York, NY, USA, 2017. [Google Scholar]
- Legarra, A.; Varona, L.; López de Maturana, E. TM Threshold Model. Available online: http://snp.toulouse.inra.fr/~alegarra/manualtm.pdf (accessed on 5 July 2019).
- Sorensen, D.; Gianola, D. Likelihood, Bayesian, and MCMC Methods. Quantitative Genetics, 1st ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Geyer, C.M. Practical markow chain Monte Carlo (with discussion). Stat. Sci. 1992, 7, 467–511. [Google Scholar]
- SAS. SAS/STAT User’s Guide 9.4; SAS Institute: Cary, NC, USA, 2017. [Google Scholar]
- Peiró, R.; Badawy, A.Y.; Blasco, A.; Santacreu, M.A. Correlated responses on growth traits after two-stage selection for ovulation rate and litter size in rabbits. Animal 2019. [Google Scholar] [CrossRef] [PubMed]
- Pascual, J.J.; Savietto, D.; Cervera, C.; Baselga, M. Resources allocation in reproductive rabbit does: A review of feeding and genetic strategies for suitable performance. World Rabbit. Sci. 2013, 21, 123–144. [Google Scholar] [CrossRef]
- Bolet, G.; Garreau, H.; Joly, T.; Theau-Clément, M.; Falières, J.; Hurtaud, J.; Bodin, L. Genetic homogenisation of birth weight in rabbits: Indirect selection response for uterine horn characteristics. Livest. Sci. 2007, 111, 28–32. [Google Scholar] [CrossRef] [Green Version]
- Garreau, H.; Bolet, G.; Larzul, C.; Robert-Granié, C.; Saleil, G.; SanCristobal, M.; Bodin, L. Results of four generations of a canalising selection for rabbit birth weight. Livest. Sci. 2008, 119, 55–62. [Google Scholar] [CrossRef]
- Hamilton, H.H.; Lukefahr, S.D.; McNitt, J.I. Maternal nest quality and its influence on litter survival and weaning performance in commercial rabbits. J. Anim. Sci. 1997, 75, 926. [Google Scholar] [CrossRef]
- García-Ximénez, F.; Vicente, J.; Viudes-De-Castro, M. Neonatal performances in 3 lines of rabbit (litter sizes, litter and individual weights). Anim. Res. 1995, 44, 255–261. [Google Scholar]
- Hull, D.; Segall, M.M. The contribution of brown adipose tissue to heat production in the new-born rabbit. J. Physiol. 1965, 181, 449–457. [Google Scholar] [CrossRef]
- Spencer, S.A.; Hull, D. The effect of over-feeding newborn rabbits on somatic and visceral growth, body composition and long-term growth potential. Br. J. Nutr. 1984, 51, 389–402. [Google Scholar] [CrossRef]
- Venge, O. The influence of nursing behaviour and milk production n early growth in rabbits. Anim. Behav. 1963, 11, 500–506. [Google Scholar] [CrossRef]
- Schaal, B.; Coudert, P.; Rideaud, P.; Fortun-Lamothe, L.; Hudson, R.; Orgeur, P. Immediate postnatal sucking in the rabbit: Its influence on pup survival and growth. Reprod. Nutr. Dev. 2000, 40, 19–32. [Google Scholar] [Green Version]
- Argente, M.; Santacreu, M.; Climent, A.; Blasco, A. Phenotypic and genetic parameters of birth weight and weaning weight of rabbits born from unilaterally ovariectomized and intact does. Livest. Prod. Sci. 1999, 57, 159–167. [Google Scholar] [CrossRef]
- Xiccato, G.; Trocino, A.; Sartori, A.; Queaque, P.I. Effect of parity order and litter weaning age on the performance and body energy balance of rabbit dos. Livest. Prod. Sci. 2004, 16, 239–251. [Google Scholar] [CrossRef]
- Rebollar, P.; Pérez-Cabal, M.; Pereda, N.; Lorenzo, P.L.; Arias-Álvarez, M.; García-Rebollar, P. Effects of parity order and reproductive management on the efficiency of rabbit productive systems. Livest. Sci. 2009, 121, 227–233. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Season | Temperature (°C) | Relative Humidity (%) | ||||
|---|---|---|---|---|---|---|
| Average | Minimum | Maximum | Average | Minimum | Maximum | |
| Winter | 14.4 | 7.5 | 22.2 | 63.2 | 26.6 | 100 |
| Spring | 20.3 | 15.2 | 28.6 | 78.7 | 43.3 | 100 |
| Summer | 26.9 | 19.2 | 33.8 | 76.9 | 32.1 | 100 |
| 20–34 | 35–44 | 45–54 | 55–64 | 65–80 | |
|---|---|---|---|---|---|
| Line H | 73 (29) | 316 (214) | 644 (578) | 494 (468) | 234 (222) |
| Line L | 128 (54) | 339 (226) | 756 (652) | 661 (609) | 338 (321) |
| Line H | Line L | |||||
|---|---|---|---|---|---|---|
| Mean | SD | CV | Mean | SD | CV | |
| Litter size | 7.69 | 2.98 | 0.38 | 8.35 | 2.43 | 0.29 |
| Survival | ||||||
| At birth | 0.89 | 0.25 | 0.28 | 0.86 | 0.25 | 0.29 |
| At 4 days of age | 0.88 | 0.25 | 0.28 | 0.87 | 0.25 | 0.29 |
| At weaning | 0.61 | 0.34 | 0.56 | 0.67 | 0.34 | 0.51 |
| Litter weight | ||||||
| Total at birth (g) | 431 | 118 | 0.27 | 450 | 119 | 0.26 |
| Live at birth (g) | 412 | 127 | 0.31 | 410 | 128 | 0.31 |
| At weaning (g) | 2518 | 1183 | 0.47 | 2460 | 1185 | 0.48 |
| Individual weight | ||||||
| Live at birth (g) | 53.5 | 11.2 | 0.21 | 54.0 | 11.2 | 0.21 |
| Dead at birth (g) | 46.3 | 11.2 | 0.24 | 46.0 | 11.1 | 0.24 |
| At weaning (g) | 493 | 151 | 0.31 | 477 | 150 | 0.31 |
| Weight distance | ||||||
| Live at birth (g) | 4.9 | 4.6 | 0.94 | 5.4 | 4.6 | 0.85 |
| Dead at birth (g) | 6.9 | 4.6 | 0.67 | 6.8 | 4.6 | 0.67 |
| Weaned (g) | 53.3 | 45.7 | 0.86 | 47.8 | 46.4 | 0.97 |
| H | L | H-L | HPD95% | P | ||
|---|---|---|---|---|---|---|
| Litter size | ||||||
| At birth | 7.7 | 8.3 | −0.6 | −1.1; | −0.2 | 1.00 |
| Survival | ||||||
| At birth | 0.89 | 0.87 | 0.02 | −0.03; | 0.06 | 0.79 |
| At 4 days of age | 0.88 | 0.87 | 0.01 | −0.04; | 0.05 | 0.67 |
| At weaning | 0.62 | 0.67 | −0.05 | −0.12; | 0.01 | 0.93 |
| Litter weight | ||||||
| Total at birth (g) | 440 | 443 | −3 | −15; | 8 | 0.73 |
| Live at birth (g) | 409 | 411 | −2 | −14; | 9 | 0.65 |
| At weaning (g) | 2461 | 2404 | 57 | −84; | 205 | 0.78 |
| Individual weight | ||||||
| Live at birth (g) | 53.5 | 54.1 | −0.4 | −1.7; | 0.8 | 0.75 |
| Dead at birth (g) | 46.3 | 46.1 | −0.2 | −2.4; | 1.9 | 0.60 |
| At weaning (g) | 495 | 480 | 15 | −17; | 47 | 0.82 |
| Weight distance | ||||||
| Live at birth (g) | 4.9 | 5.4 | −0.5 | −0.9; | 0.0 | 0.97 |
| Dead at birth (g) | 7.0 | 6.8 | 0.2 | −0.9; | 1.3 | 0.68 |
| Weaned (g) | 54.1 | 47.8 | 6.3 | 0.2; | 12.3 | 0.98 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Agea, I.; García, M.-L.; Blasco, A.; Argente, M.-J. Litter Survival Differences between Divergently Selected Lines for Environmental Sensitivity in Rabbits. Animals 2019, 9, 603. https://doi.org/10.3390/ani9090603
Agea I, García M-L, Blasco A, Argente M-J. Litter Survival Differences between Divergently Selected Lines for Environmental Sensitivity in Rabbits. Animals. 2019; 9(9):603. https://doi.org/10.3390/ani9090603
Chicago/Turabian StyleAgea, Ivan, María-Luz García, Agustín Blasco, and María-José Argente. 2019. "Litter Survival Differences between Divergently Selected Lines for Environmental Sensitivity in Rabbits" Animals 9, no. 9: 603. https://doi.org/10.3390/ani9090603