Homology and Potential Cellular and Molecular Mechanisms for the Development of Unique Feather Morphologies in Early Birds

Abstract
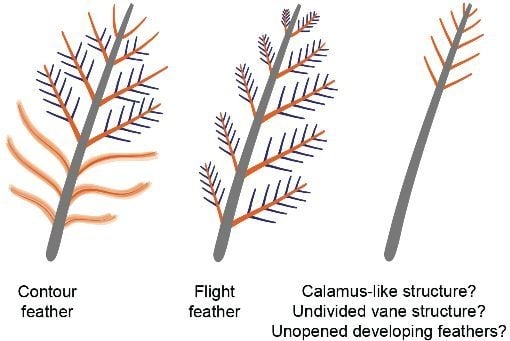
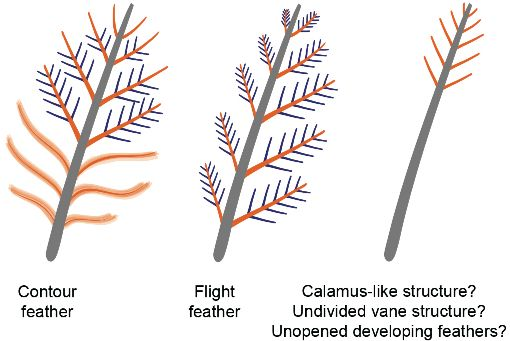
:
Institutional Abbreviations:
1. Introduction
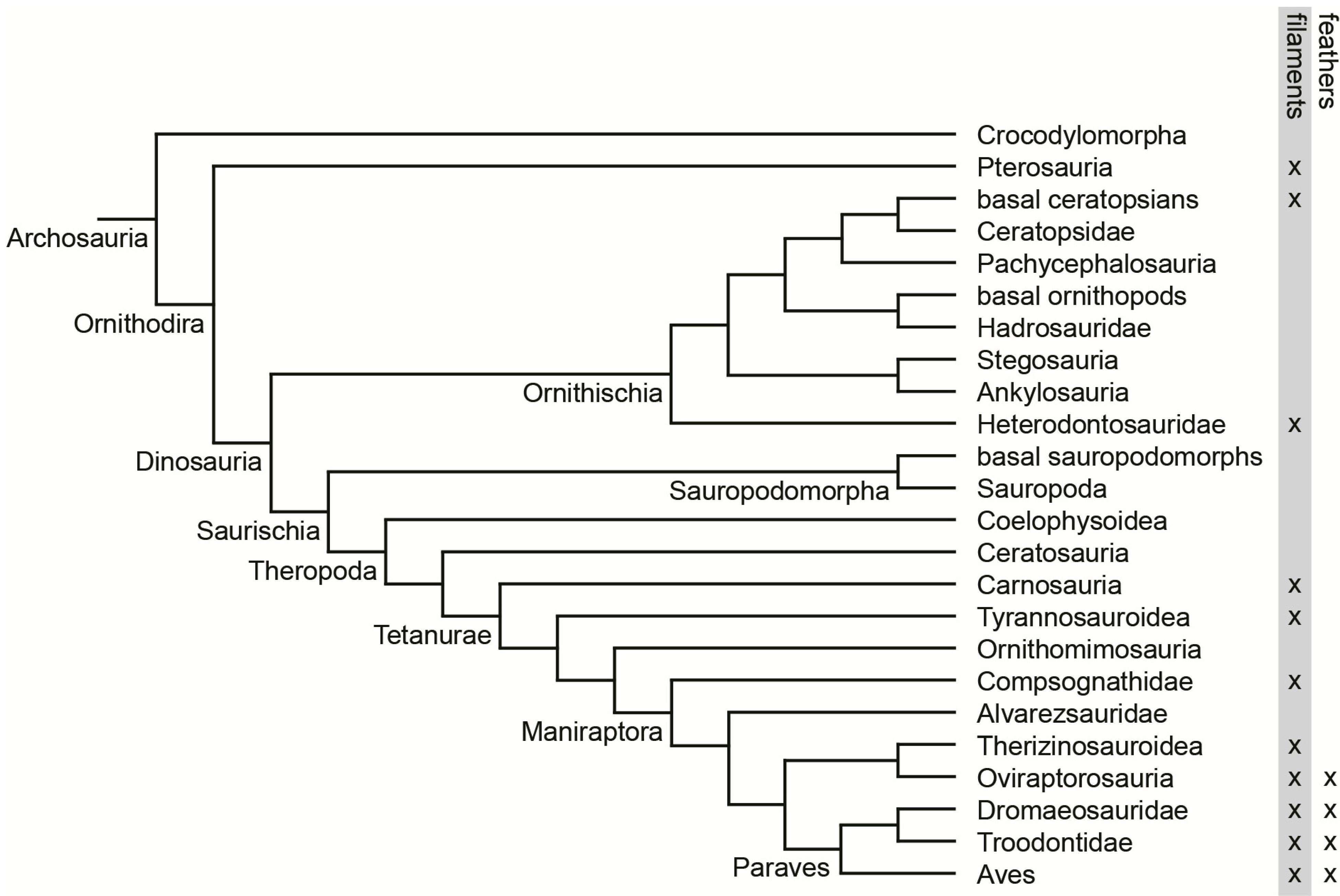
2. Morphological Description
2.1. Non-avian Maniraptoran Theropods
2.1.1. Similicaudipteryx yixianensis

2.1.2. Epidexipteryx hui

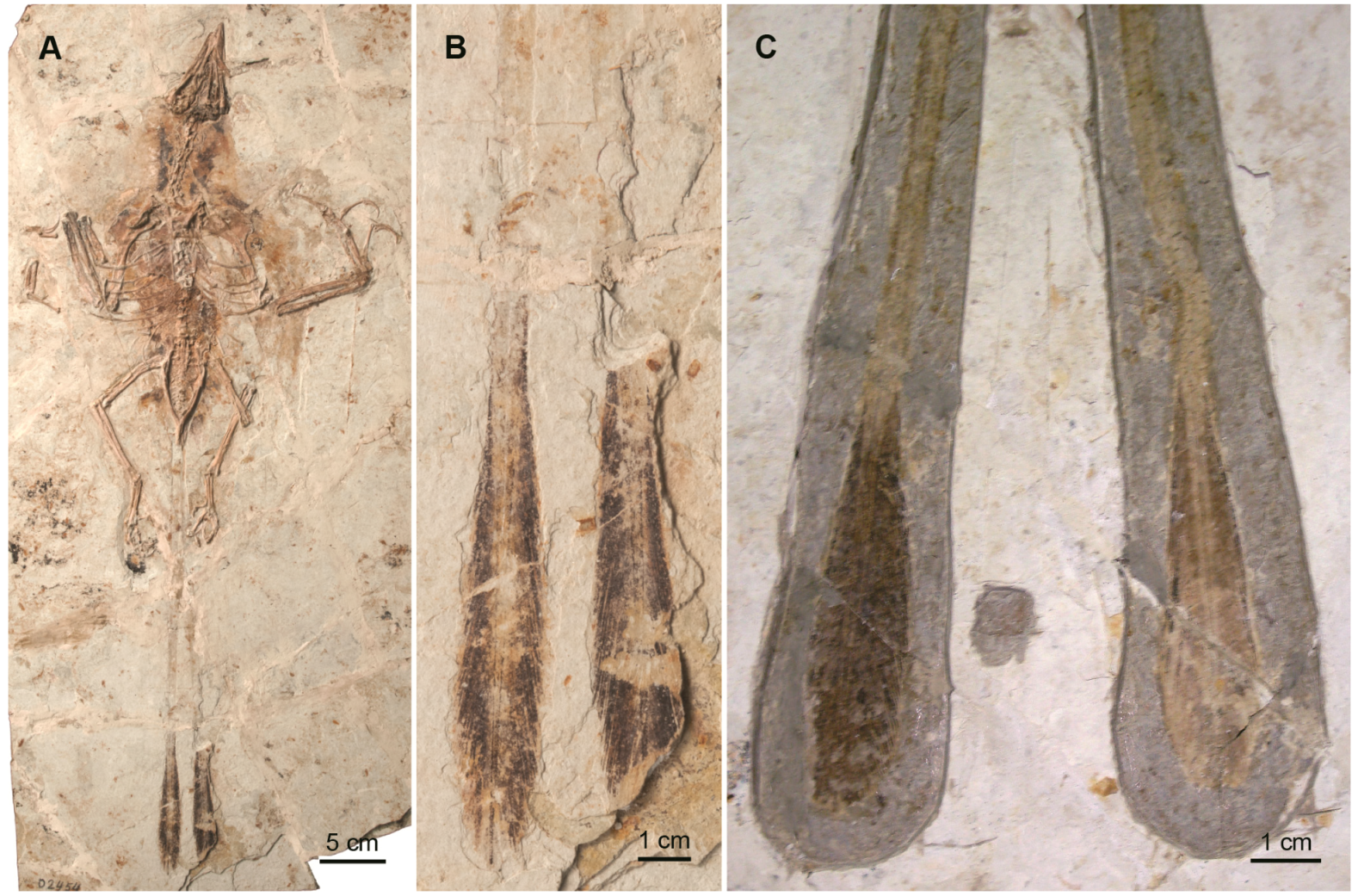
2.2. Basal Birds
2.2.1. Confuciusornithiformes
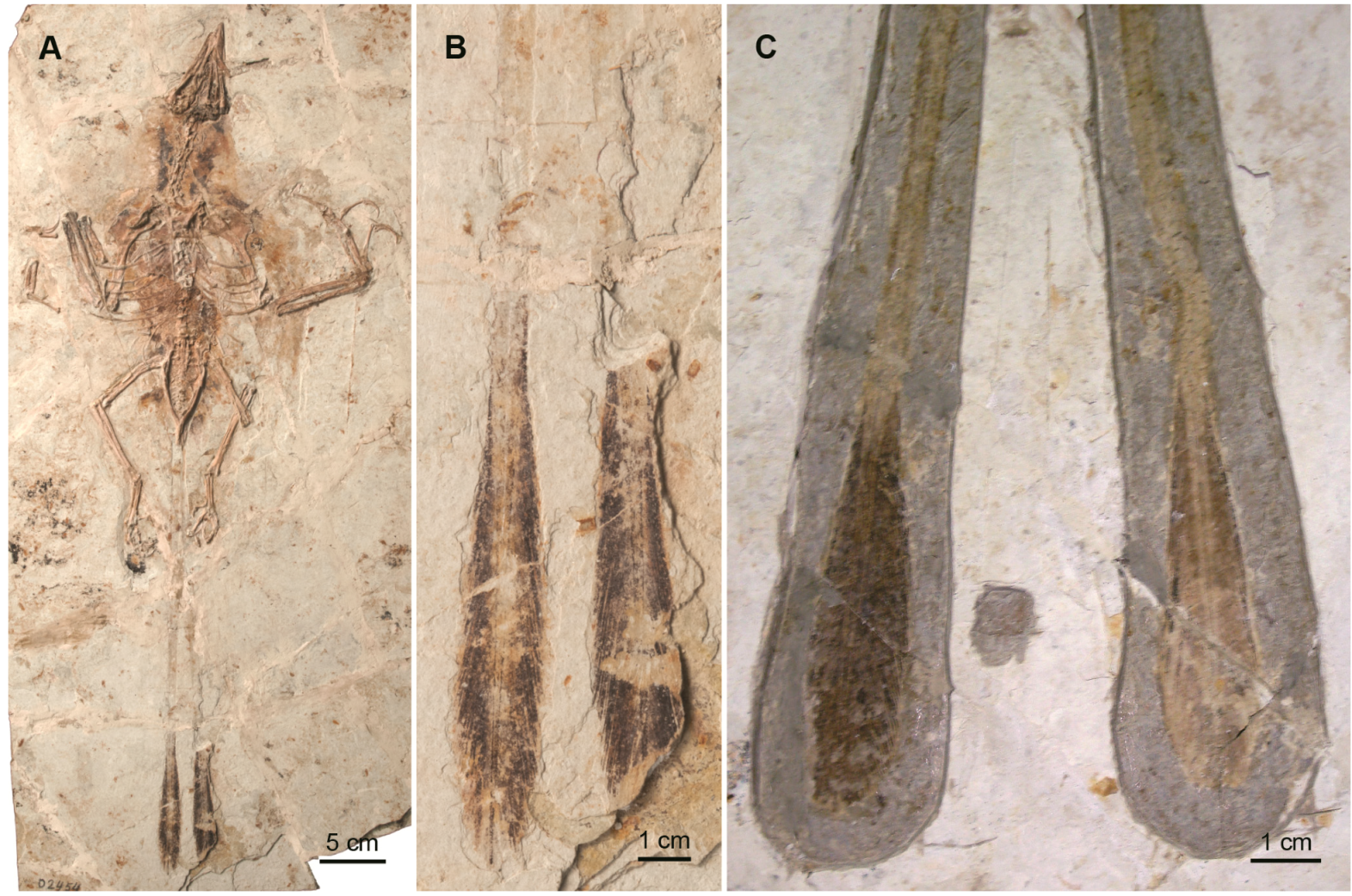
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
2.2.2. Enantiornithes



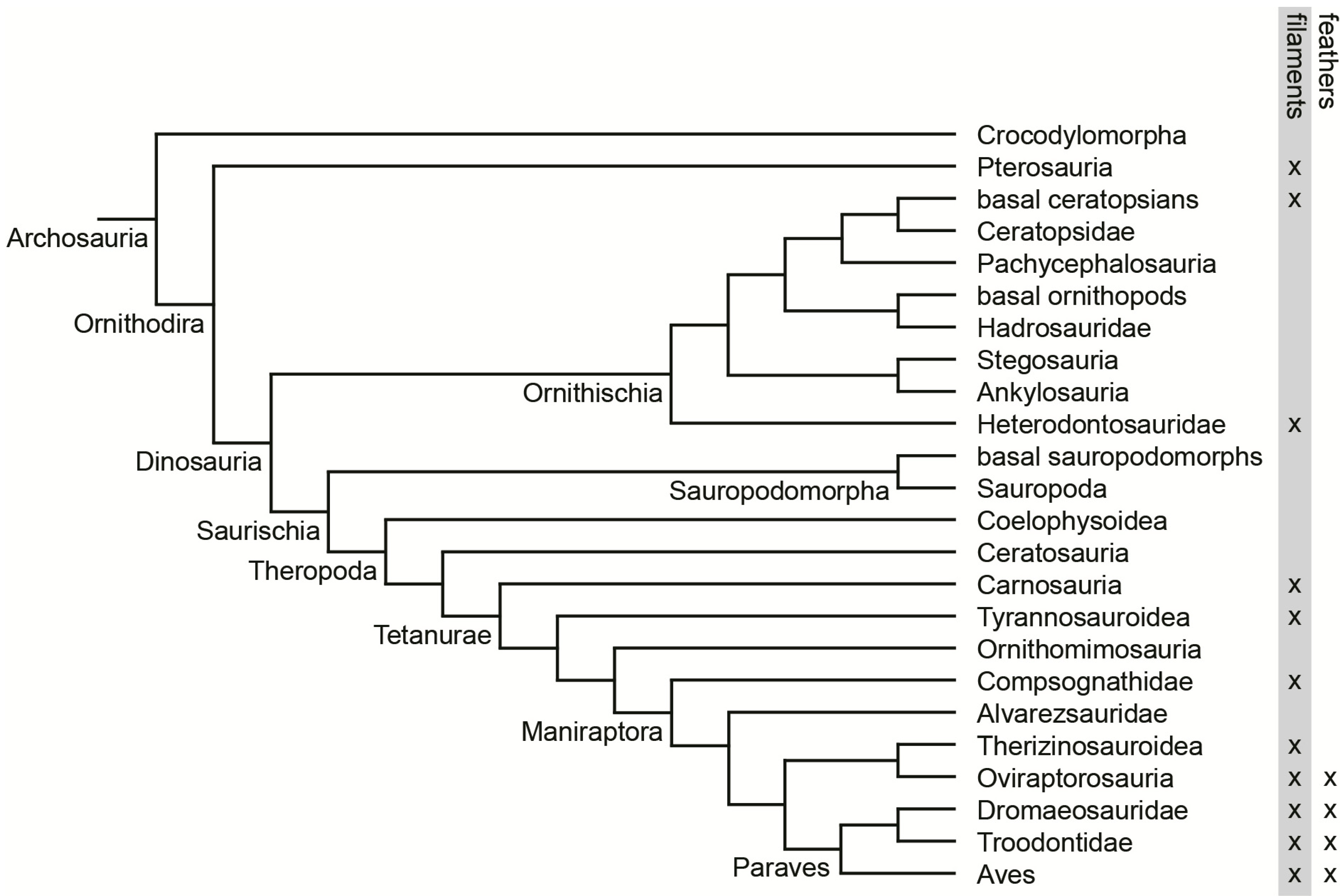
2.3. Comparison
3. Results and Discussion
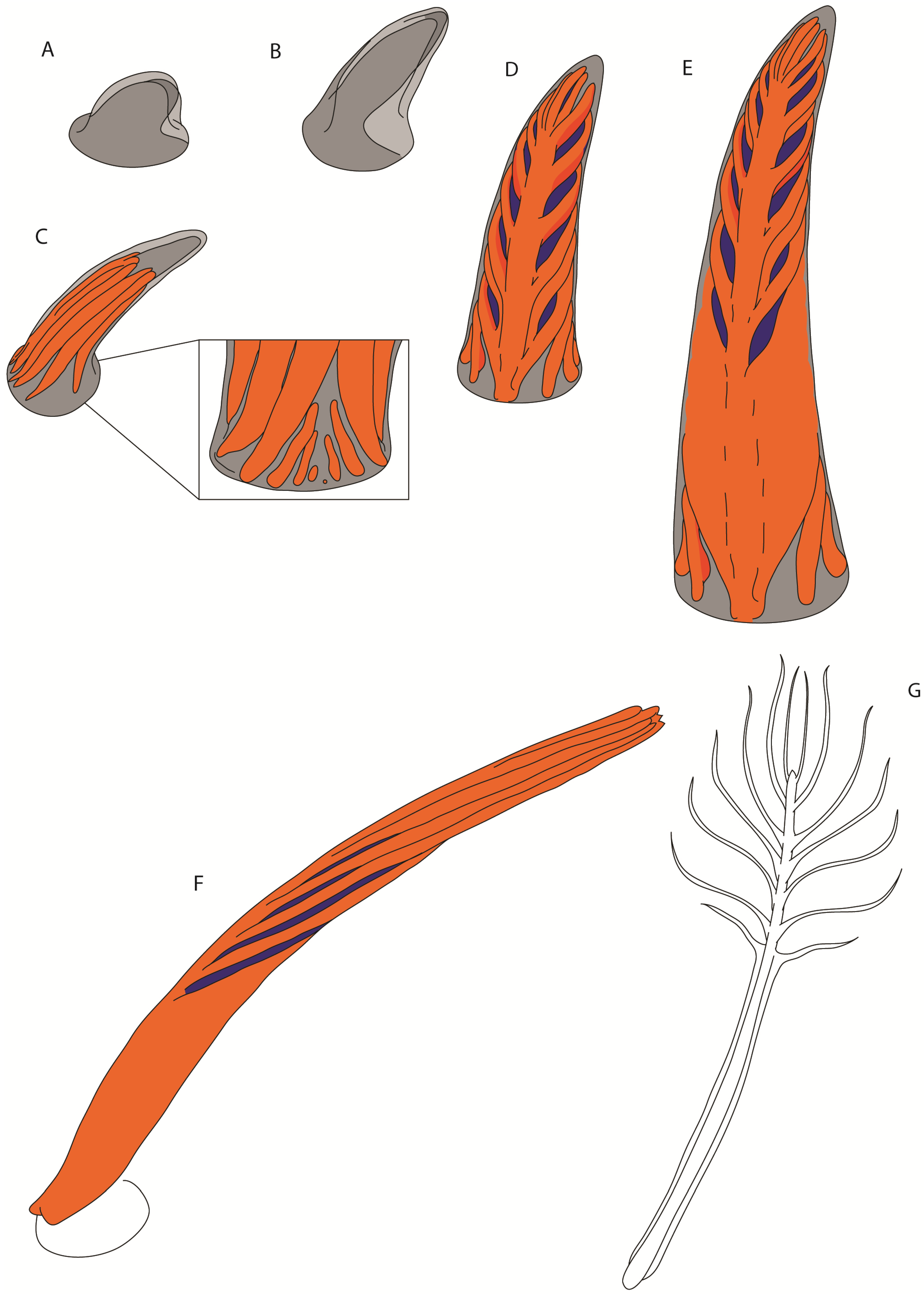
3.1. Rachis Dominated Feathers
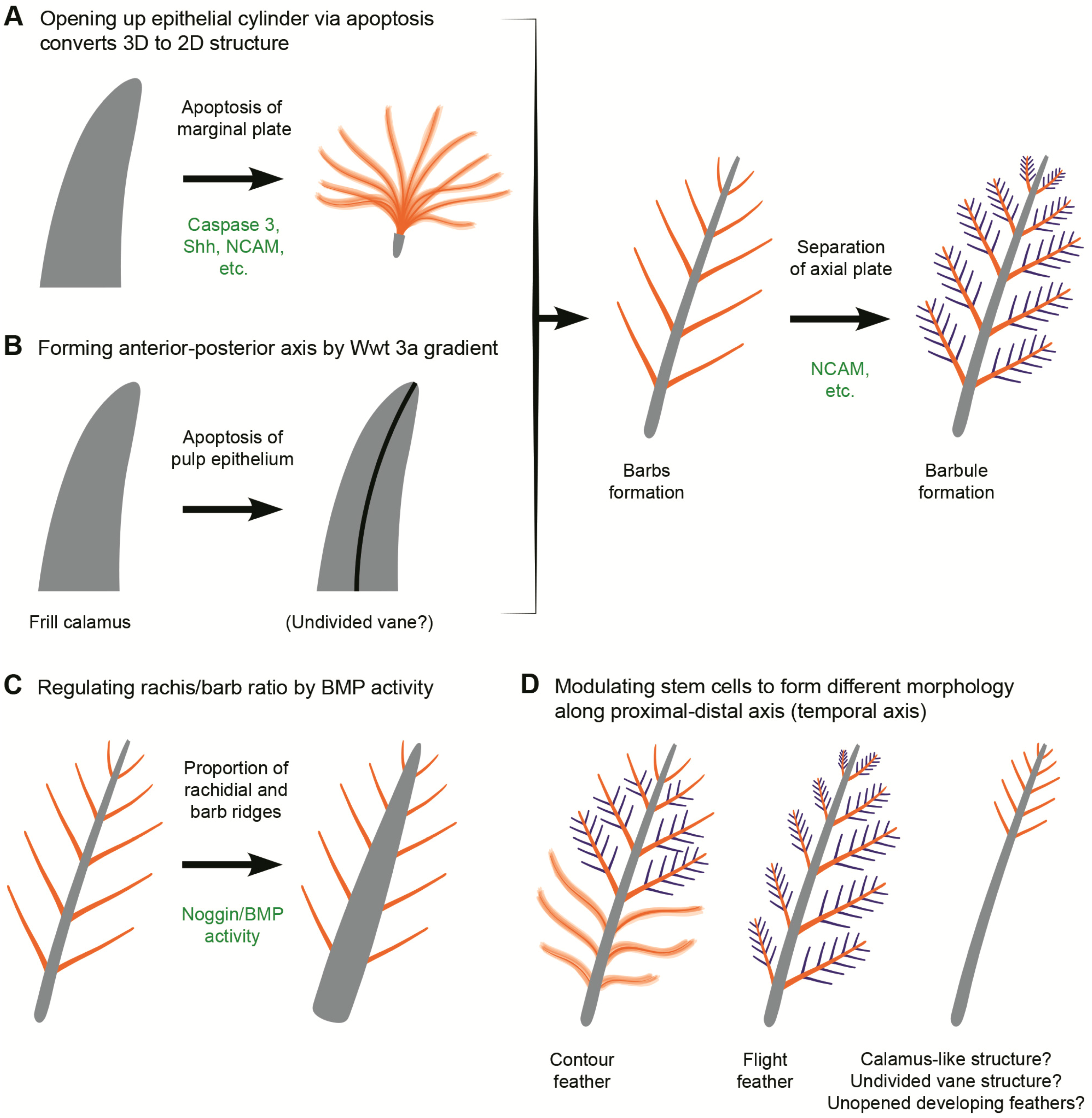
3.2. Potential Cellular and Molecular Basis for Evolution Novelty in Feather Morphogenesis
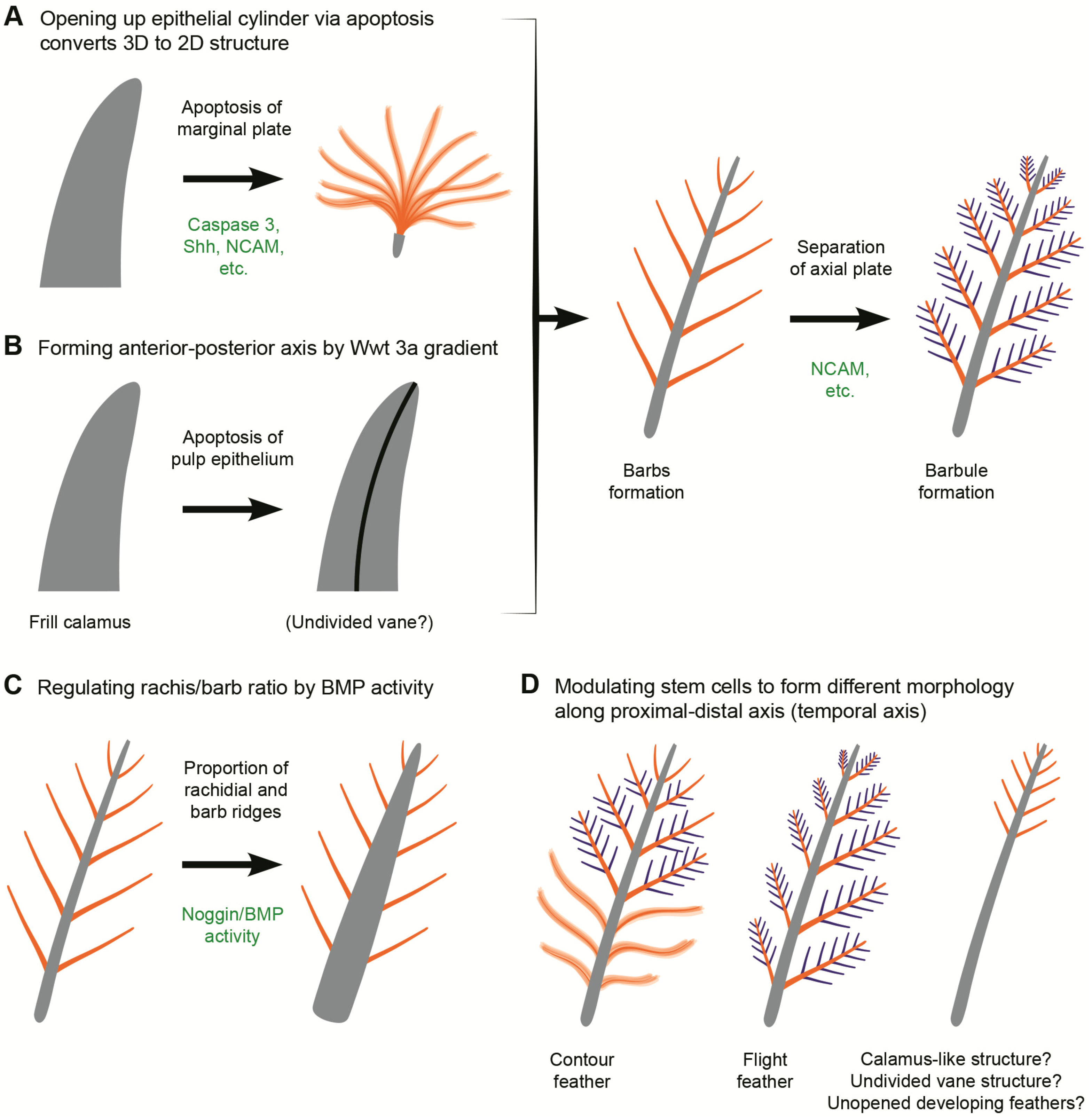
3.3. Function
3.4. Proximally Ribbon-like Pennaceous Feathers
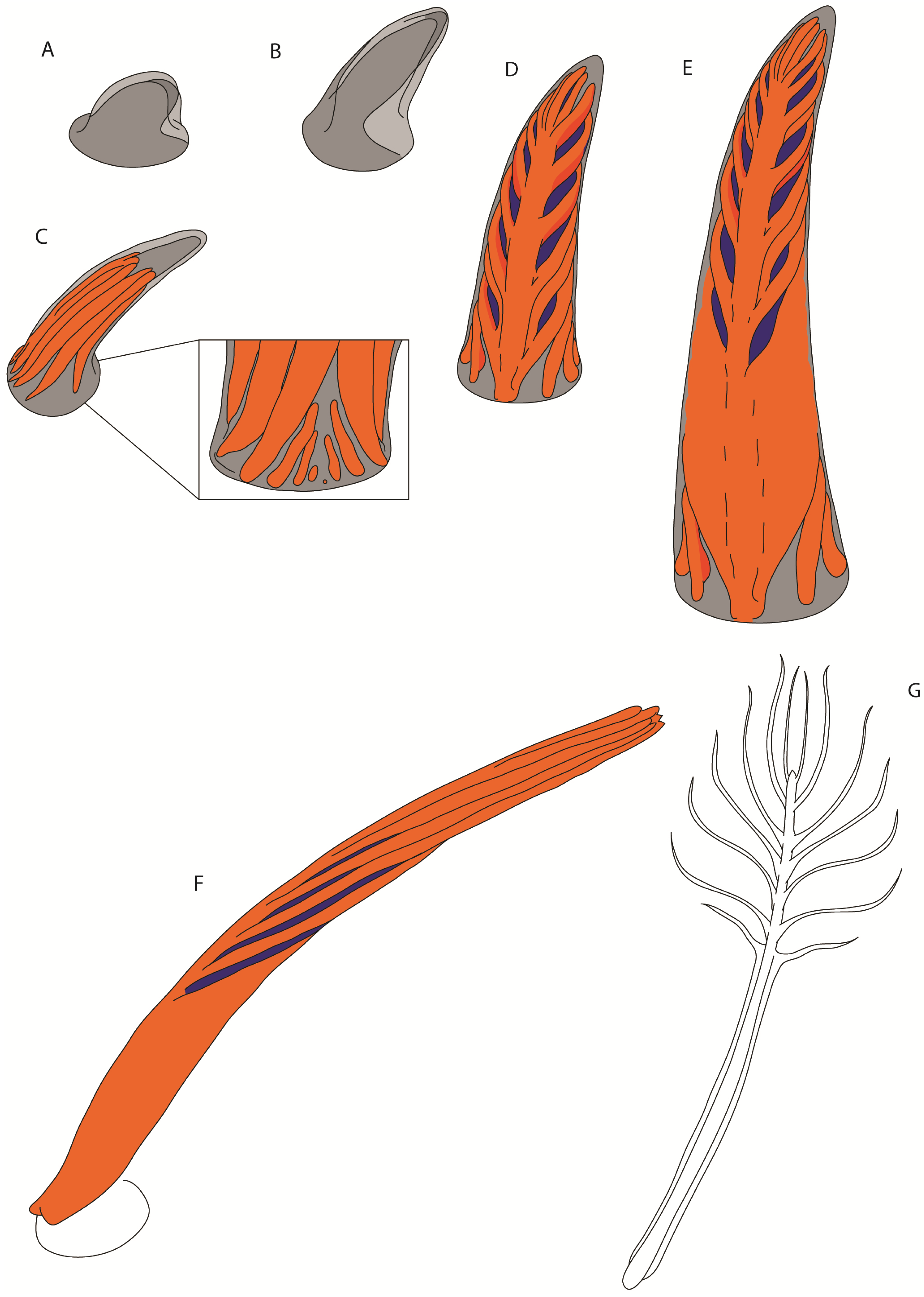
4. Conclusions
Acknowledgments
References
- Ji, Q.; Ji, S.A. On discovery of the earliest bird fossil in China and the origin of birds. Chin. Geol. 1996, 233, 30–33. [Google Scholar]
- Ji, Q.; Currie, P.J.; Norell, M.A.; Ji, S.A. Two feathered dinosaurs from northeastern China. Nature 1998, 393, 753–761. [Google Scholar] [CrossRef]
- Xu, X.; Zhao, Q.; Norell, M.A.; Sullivan, C.; Hone, D.W.; Erickson, G.M.; Wang, X.L.; Han, F.L.; Guo, Y. A new feathered maniraptoran dinosaur fossil that fills a morphological gap in avian origin. Chin. Sci. Bull. 2009, 54, 430–435. [Google Scholar] [CrossRef]
- Xu, X.; Zheng, X.T.; You, H.L. Exceptional dinosaur fossils show ontogenetic development of early feathers. Nature 2010, 464, 1339–1341. [Google Scholar]
- Zheng, X.T.; You, H.L.; Xu, X.; Dong, Z.M. An Early Cretaceous heterodontosaurid dinosaur with filamentous integumentary structures. Nature 2009, 458, 333–337. [Google Scholar]
- Zhang, F.C.; Zhou, Z.H.; Xu, X.; Wang, X.L.; Sullivan, C. A bizarre Jurassic maniraptoran from China with elongate ribbon-like feathers. Nature 2008, 455, 1105–1108. [Google Scholar] [CrossRef]
- Xu, X.; Wang, K.B.; Zhang, K.; Ma, Q.Y.; Xing, L.D.; Sullivan, C.; Hu, D.Y.; Cheng, S.Q.; Wang, S. A gigantic feathered dinosaur from the Lower Cretaceous of China. Nature 2012, 484, 92–95. [Google Scholar]
- Chuong, C.M.; Widelitz, R.B. Feather morphogenesis: A model of the formation of epithelial appendage. In Molecular Basis of Epithelial Appendage Morphogenesis; Chuong, C.M., Ed.; Landes Bioscience: Austin, TX, USA, 1998; pp. 57–74. [Google Scholar]
- Yu, M.; Wu, P.; Widelitz, R.B.; Chuong, C.M. The morphogenesis of feathers. Nature 2002, 420, 308–312. [Google Scholar] [CrossRef]
- Harris, M.P.; Fallon, J.F.; Prum, R.O. Shh-Bmp2 signaling module and the evolutionary origin and diversification of feathers. J. Exp. Zool. B Mol. Dev. Evol. 2002, 294, 160–176. [Google Scholar] [CrossRef]
- Chang, C.H.; Yu, M.K.; Wu, P.; Jiang, T.X.; Yu, H.S.; Widelitz, R.B.; Chuong, C.M. Sculpting skin appendages out of epidermal layers via temporally and spatially regulated apoptotic events. J. Invest. Dermatol. 2004, 122, 1348–1355. [Google Scholar] [CrossRef]
- Prum, R.O. Evolution of the morphological innovations of feathers. J. Exp. Zool. B Mol. Dev. Evol. 2005, 304, 570–579. [Google Scholar] [CrossRef]
- Lin, C.M.; Jiang, T.X.; Widelitz, R.B.; Chuong, C.M. Molecular signaling in feather morphogenesis. Curr. Opin. Cell Biol. 2006, 18, 730–741. [Google Scholar] [CrossRef]
- Alibardi, L. Cell structure of barb ridges in down feathers and juvenile wing feathers of the developing chick embryo: Barb ridge modification in relation to feather evolution. Ann. Anat. 2006, 188, 303–318. [Google Scholar] [CrossRef]
- Yue, Z.; Jiang, T.X.; Widelitz, R.B.; Chuong, C.M. Wnt3a gradient converts radial to bilateral feather symmetry via topological arrangement of epithelia. Proc. Natl. Acad. Sci. USA. 2006, 103, 951–955. [Google Scholar]
- Maderson, P.F.; Hillenius, W.J.; Hiller, U.; Dove, C.C. Towards a comprehensive model of feather regeneration. J. Morphol. 2009, 270, 1166–1208. [Google Scholar] [CrossRef]
- Mayr, G.; Peters, D.S.; Plowdowski, G.; Vogel, O. Bristle-like integumentary structures at the tail of the horned dinosaur Psittacosaurus. Naturwissenschaften 2002, 89, 361–365. [Google Scholar] [CrossRef]
- Xu, X.; Norell, M.A.; Kuang, X.; Wang, X.; Zhao, Q.; Jia, C. Basal tyrannosauroids from China and evidence for protofeathers in tyrannosauroids. Nature 2004, 431, 680–684. [Google Scholar] [CrossRef]
- Xu, X.; Guo, Y. The origin and early evolution of feathers: Insights from recent paleontological and neontological data. Vertebr. Palasiat. 2009, 47, 311–329. [Google Scholar]
- Hu, D.Y.; Hou, L.H.; Zhang, L.J.; You, H.L. A pre-Archaeopteryx troodontid theropod from China with long feathers on the metatarsus. Nature 2009, 461, 640–643. [Google Scholar]
- Xu, X.; Zhang, F. A new maniraptoran dinosaur from China with long feathers on the metatarsus. Naturwissenschaften 2005, 92, 173–177. [Google Scholar] [CrossRef]
- Xu, X.; Zhou, Z.; Wang, X.; Kuang, X.; Du, X. Four-winged dinosaurs from China. Nature 2003, 421, 335–340. [Google Scholar] [CrossRef]
- Xu, X.; You, H.L.; Kai, D.; Han, F.L. An Archaeopteryx-like theropod from China and the origin of Avialae. Nature 2011, 475, 465–475. [Google Scholar]
- Xu, X.; Wang, X.L.; Wu, X.C. A dromaeosaurid dinosaur with a filamentous integument from the Yixian Formation of China. Nature 1999, 401, 262–266. [Google Scholar]
- Ji, Q.; Norell, M.A.; Gao, K.Q.; Ji, S.A.; Ren, D. The distribution of integumentary structures in a feathered dinosaur. Nature 2001, 410, 1084–1088. [Google Scholar] [CrossRef]
- Xu, X.; Zhou, Z.; Prum, R.O. Branched integumental structures in Sinornithosaurus and the origin of feathers. Nature 2001, 410, 200–204. [Google Scholar] [CrossRef]
- Foth, C. On the identification of feather structures in stem-line representatives of birds: Evidence from fossils and actuopalaeontology. Palaeontol. Z. 2012, 86, 91–102. [Google Scholar]
- Butler, R.J.; Upchurch, P.; Norman, D.B. The phylogeny of the ornithischian dinosaurs. J. Syst. Palaeontol. 2008, 6, 1–40. [Google Scholar] [CrossRef]
- Brusatte, S.L.; Nesbitt, S.J.; Irmis, R.B.; Butler, R.J.; Benton, M.J.; Norell, M.A. The origin and early radiation of dinosaurs. Earth Sci. Rev. 2010, 101, 68–100. [Google Scholar] [CrossRef]
- Makovicky, P.J.; Zanno, L.E. Theropod diversity and the refinement of avian characteristics. In Living Dinosaurs: The Evolutionary History of Modern Birds; Dyke, G.D., Kaiser, G., Eds.; John Wiley & Sons: Oxford, UK, 2011; pp. 9–29. [Google Scholar]
- Rauhut, O.W. M.; Foth, C.; Tischlinger, H.; Norell, M.A. Exceptionally preserved juvenile megalosauroid theropod dinosaur with filamentous integument from the Late Jurassic of Germany. Proc. Natl. Acad. Sci. USA 2012, 109, 11746–11751. [Google Scholar]
- Chiappe, L.M.; Ji, S.; Ji, Q.; Norell, M.A. Anatomy and systematics of the Confuciusornithidae (Theropoda: Aves) from the Late Mesozoic of northeastern China. Bull. Am. Mus. Nat. Hist. 1999, 242, 1–89. [Google Scholar]
- Elzanowski, A. Biology of basal birds and the origin of avian flight. In Proceedings of the 5th Symposium of the Society of Avian Paleontology and Evolution, Beijing, China, 1–4 June 2000; Zhou, Z., Zhang, F., Eds.; Science Press: Beijing, China, 2002; pp. 211–226. [Google Scholar]
- Chuong, C.M.; Wu, P.; Zhang, F.C.; Xu, X.; Yu, M.; Widelitz, R.B.; Jiang, T.X.; Hou, L. Adaptation to the sky: Defining the feather with integument fossils from Mesozoic China and experimental evidence from molecular laboratories. J. Exp. Zool. B Mol. Dev. Evol. 2003, 298, 42–56. [Google Scholar]
- Zhang, F.; Zhou, Z.; Dyke, G.J. Feathers and “feather-like” integumentary structures in Liaoning birds and dinosaurs. Geol. J. 2006, 41, 395–404. [Google Scholar] [CrossRef]
- Wellnhofer, P. Archaeopteryx. Der Urvogel von Solnhofen; Friedrich Pfeil: München, Germany, 2008. [Google Scholar]
- Nudds, R.L.; Dyke, G.D. Narrow primary feather rachises in Confuciusornis and Archaeopteryx suggest poor flight ability. Science 2010, 328, 887–889. [Google Scholar] [CrossRef]
- Zheng, X.T.; Xu, X.; Zhou, Z.H.; Miao, D.; Zhang, F.C. Comment on “Narrow primary feather rachises in Confuciusornis and Archaeopteryx suggest poor flight ability. Science 2010, 330, 320. [Google Scholar]
- Hou, L.; Martin, L.D.; Zhou, Z.; Feduccia, A. Early adaptive radiation of birds: Evidence from fossils from northeastern China. Science 1996, 274, 1164–1167. [Google Scholar] [CrossRef]
- Zhang, F.; Zhou, Z. A primitive enantiornithine bird and the origin of feathers. Science 2000, 290, 1955–1960. [Google Scholar] [CrossRef]
- Li, L.; Duan, Y.; Hu, D.; Wang, L.; Cheng, S.; Hou, L. New eoentantiornithid bird from the Early Cretaceous Jiufotang Formation of western Liaoning, China. Acta Geol. Sin. Engl. Ed. 2006, 80, 38–41. [Google Scholar] [CrossRef]
- Prum, R.O.; Brush, A.H. The evolutionary origin and diversification of feathers. Q. Rev. Biol. 2002, 77, 261–295. [Google Scholar]
- He, T.; Wang, X.L.; Zhou, Z.H. A new genus and species of caudipterid dinosaur from the Lower Cretaceous Jiufotang Formation of western Liaoning, China. Vertebr. Palasiat. 2008, 46, 178–189. [Google Scholar]
- Prum, R.O. Moulting tail feathers in a juvenile oviraptorisaur. Nature 2010, 468, E1. [Google Scholar] [CrossRef]
- Chiappe, L.M.; Marugán-Lobón, J.; Ji, S.; Zhou, Z. Life history of a basal bird: Morphometrics of the Early Cretaceous Confuciusornis. Biol. Lett. 2008, 4, 719–723. [Google Scholar] [CrossRef]
- Ji, Q.; Chiappe, L.M.; Ji, S. A new late Mesozoic confuciusornithid bird from China. J. Vertebr. Paleontol. 1999, 19, 1–7. [Google Scholar]
- Zhang, F.; Zhou, Z.; Benton, M.J. A primitive confuciusornithid bird from China and its implications for early avian flight. Sci. China Ser. D Earth Sci. 2008, 51, 625–639. [Google Scholar]
- Stettenheim, P.R. The integumentary morphology of modern birds—An overview. Am. Zool. 2000, 40, 461–477. [Google Scholar] [CrossRef]
- Vinther, J.; Briggs, D.E. G.; Prum, R.O.; Saranathan, V. The colour of fossil feathers. Biol. Lett. 2008, 522–525. [Google Scholar]
- Zhang, F.C.; Kearns, S.L.; Orr, P.J.; Benton, M.J.; Zhou, Z.H.; Johnson, D.; Xu, X.; Wang, X.L. Fossilized melanosomes and the colour of Cretaceous dinosaurs and birds. Nature 2010, 463, 1075–1078. [Google Scholar] [CrossRef]
- Marugán-Lobón, J.; Chiappe, L.M.; Ji, S.A.; Zhou, Z.H.; Gao, C.H.; Hu, D.Y.; Meng, Q.J. Quantitative patterns of morphological variation in the appendicular skeleton of the Early Cretaceous bird Confuciusornis. J. Syst. Palaeontol. 2011, 9, 91–101. [Google Scholar] [CrossRef]
- Zheng, X.; Zhang, Z.; Hou, L. A new enantiornitine bird with four long rectrices from the Early Cretaceous of northern Hebei, China. Acta Geol. Sin. Engl. Ed. 2007, 81, 703–708. [Google Scholar] [CrossRef]
- Chen, P.J.; Dong, Z.; Zhen, S. An exceptionally well-preserved theropod dinosaur from the Yixian Formation of China. Nature 1998, 391, 147–152. [Google Scholar] [CrossRef]
- Xu, X.; Tang, Z.L.; Wang, X.L. A therizinosauroid dinosaur with integumentary structures from China. Nature 1999, 399, 350–354. [Google Scholar] [CrossRef]
- Xu, X.; Zhou, Z.; Wang, X. The smallest known non-avian theropod dinosaur. Nature 2000, 408, 705–708. [Google Scholar]
- Norell, M.; Ji, Q.; Gao, K.; Yuan, C.; Zhao, Y.; Wang, L. “Modern” feathers on a non-avian dinosaur. Nature 2002, 416, 36–37. [Google Scholar]
- Snow, D.W. Family Momotidae (motmots). In Handook of the Birds of the World; Hoyo, J., Elliott, A., Sargatal, J., Eds.; Lynx Edicions: Barcelona, Spain, 2001; Volume 6. [Google Scholar]
- Gill, F.B. Ornithology, 3rd ed; W.H. Freeman and Company: New York, NY, USA, 2007. [Google Scholar]
- Harris, M.P.; Williamson, S.; Fallon, J.F.; Meinhardt, H.; Prum, R.O. Molecular evidence for an activator-inhibitor mechanism in development of embryonic feather branching. Proc. Natl. Acad. Sci. USA 2005, 102, 11734–11739. [Google Scholar] [CrossRef]
- Feduccia, A. The Origin and Evolution of Birds; Yale University Press: New Haven, CT, USA, 1996. [Google Scholar]
- Peters, W.S.; Peters, D.S. Life history, sexual dimorphism and “ornamental” feathers in the Mesozoic bird Confuciusornis sanctus. Biol. Lett. 2009, 5, 817–820. [Google Scholar] [CrossRef]
- Peters, W.S.; Peters, D.S. Sexual size dimorphism is the most consistent explanation for the body size spectrum of Confuciusornis sanctus. Biol. Lett. 2010, 6, 531–532. [Google Scholar] [CrossRef]
- Padian, K.; Horner, J.R. The evolution of “bizarre structures” in dinosaurs: Biomechanics, sexual selection or species recognition? J. Zool. 2011, 238, 3–17. [Google Scholar] [CrossRef]
- Li, D.S.; Sullivan, C.; Zhou, Z.H.; Zhang, F.C. Basal birds from China: A brief review. Chin. Birds 2010, 1, 83–96. [Google Scholar] [CrossRef]
- O’Connor, J.K.; Sun, C.K.; Xu, X.; Wang, X.L.; Zhou, Z.H. A new species of Jeholornis with complete caudal integument. Hist. Biol. 2012, 24, 29–41. [Google Scholar]
- Xu, X.; Zheng, X.T.; You, H.L. Reply: Moulting tail feathers in a juvenile oviraptorosaur. Nature 2010, 468, E2. [Google Scholar]
- Brush, A.H. Evolving a protofeather and feather diversity. Am. Zool. 2000, 40, 631–639. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
O’Connor, J.K.; Chiappe, L.M.; Chuong, C.-m.; Bottjer, D.J.; You, H. Homology and Potential Cellular and Molecular Mechanisms for the Development of Unique Feather Morphologies in Early Birds. Geosciences 2012, 2, 157-177. https://doi.org/10.3390/geosciences2030157
O’Connor JK, Chiappe LM, Chuong C-m, Bottjer DJ, You H. Homology and Potential Cellular and Molecular Mechanisms for the Development of Unique Feather Morphologies in Early Birds. Geosciences. 2012; 2(3):157-177. https://doi.org/10.3390/geosciences2030157
Chicago/Turabian StyleO’Connor, Jingmai K., Luis M. Chiappe, Cheng-ming Chuong, David J. Bottjer, and Hailu You. 2012. "Homology and Potential Cellular and Molecular Mechanisms for the Development of Unique Feather Morphologies in Early Birds" Geosciences 2, no. 3: 157-177. https://doi.org/10.3390/geosciences2030157
APA StyleO’Connor, J. K., Chiappe, L. M., Chuong, C.-m., Bottjer, D. J., & You, H. (2012). Homology and Potential Cellular and Molecular Mechanisms for the Development of Unique Feather Morphologies in Early Birds. Geosciences, 2(3), 157-177. https://doi.org/10.3390/geosciences2030157