Intraspecific Length Variation and Shell Thickness of the Ostracod Cyprideis torosa (Jones, 1850) as a Potential Tool for Palaeosalinity Characterization

Abstract
:1. Introduction
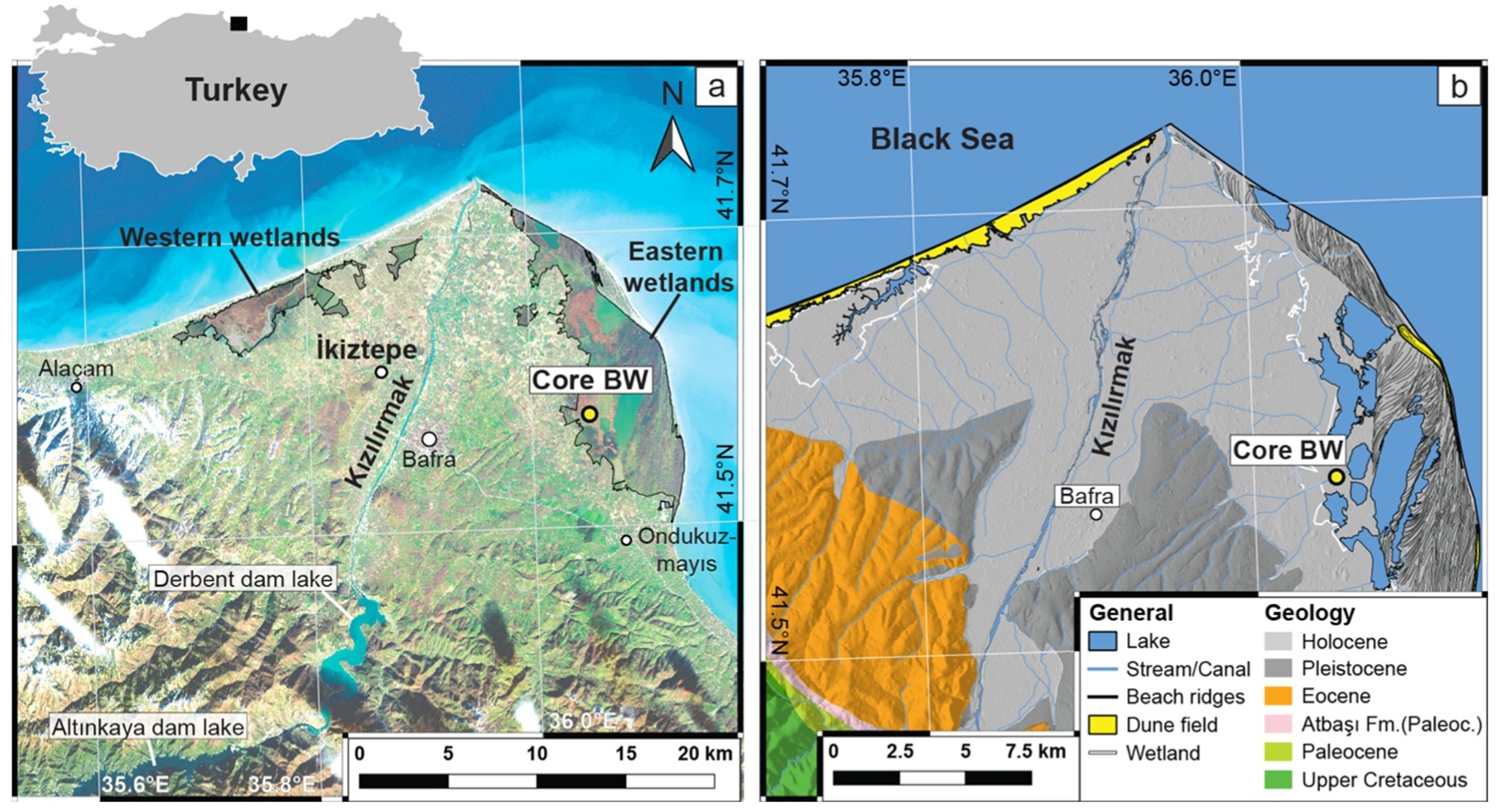
2. Materials and Methods
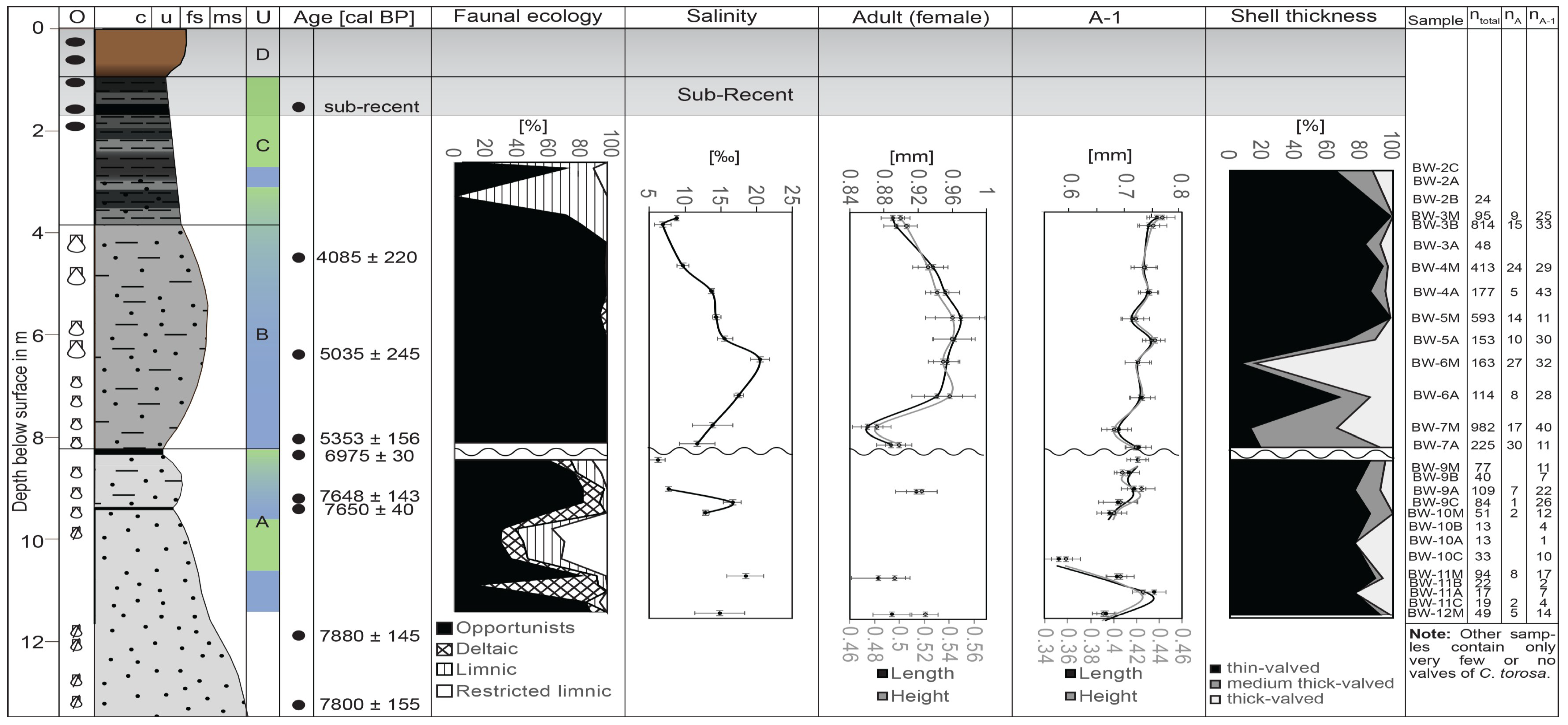
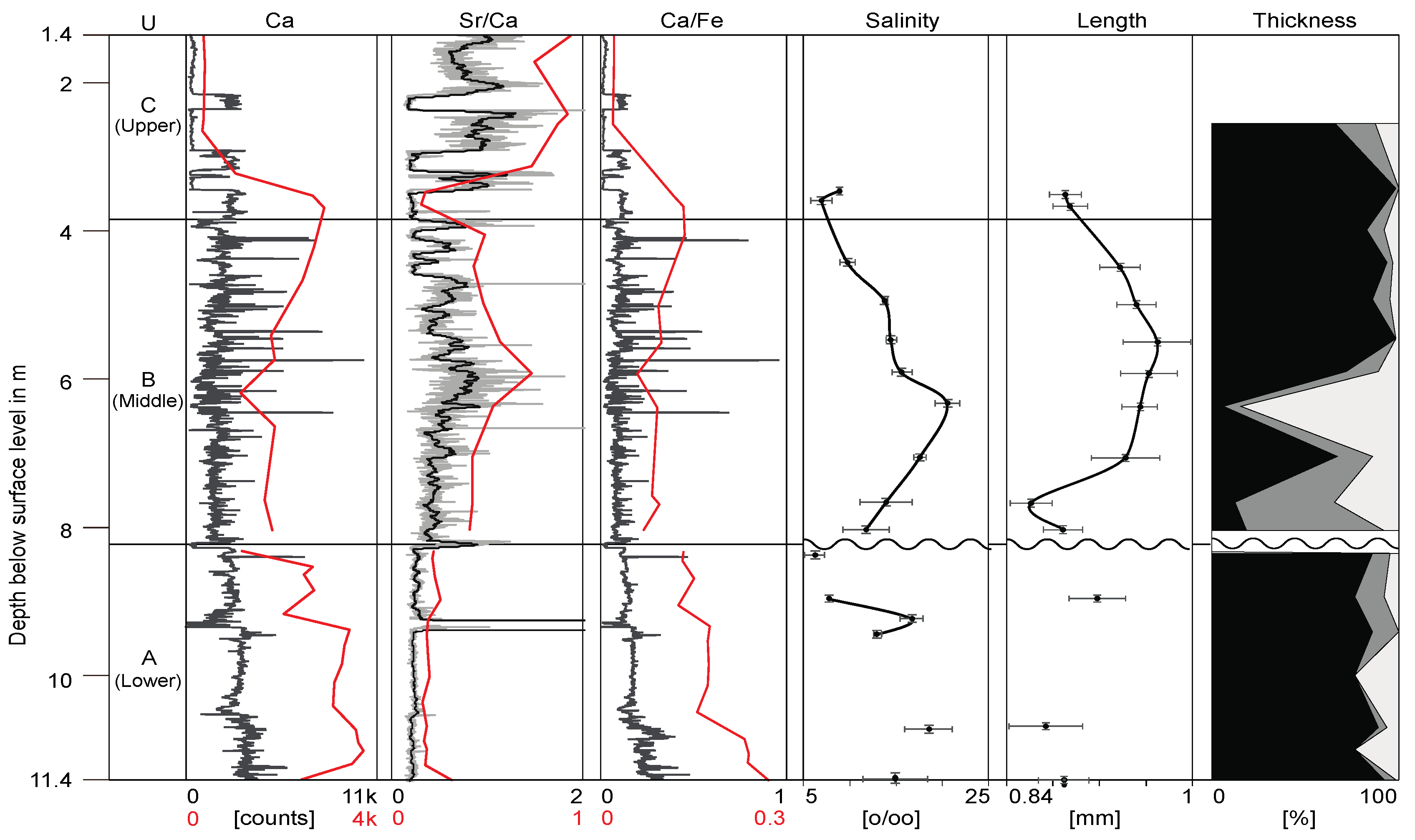
3. Results
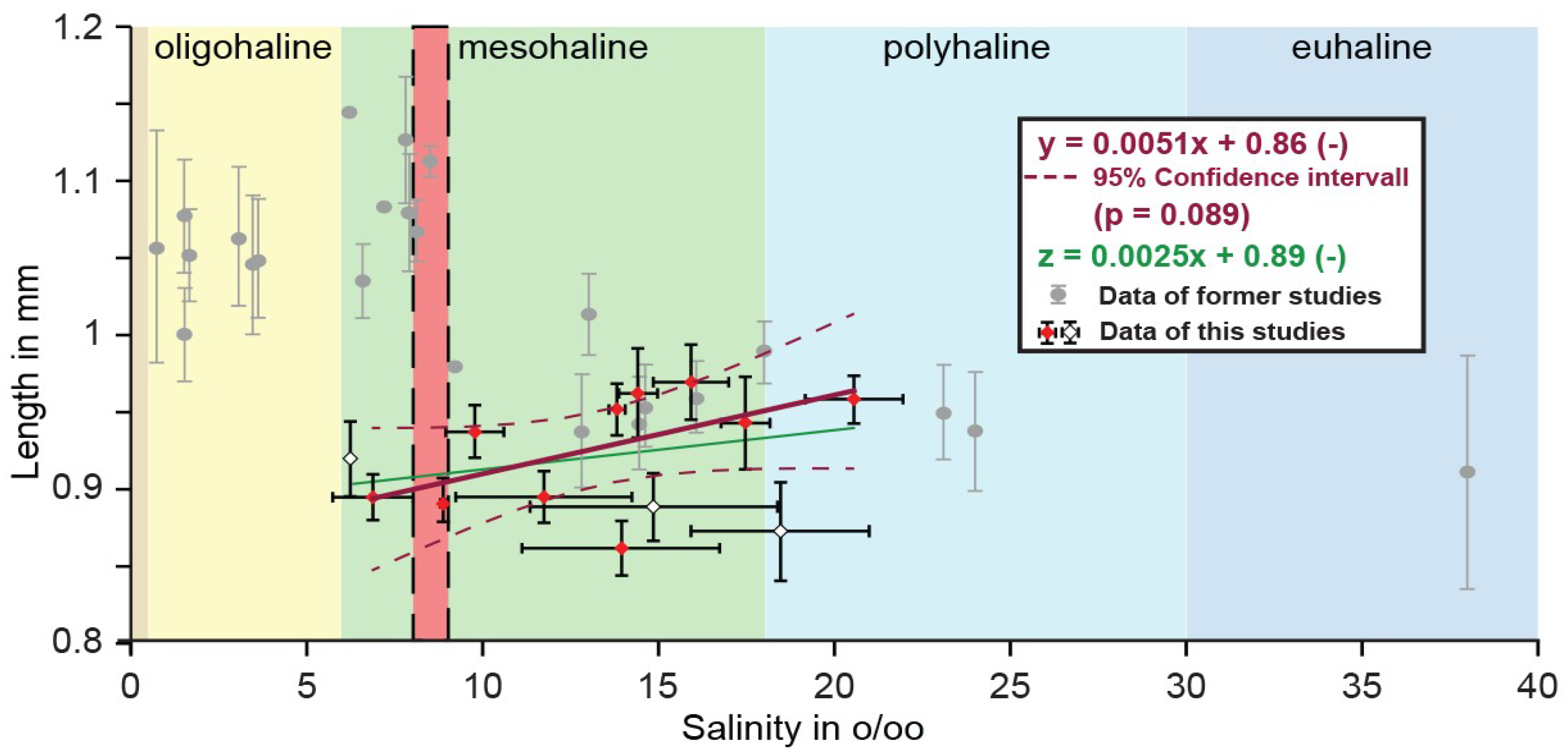
3.1. Length Variation
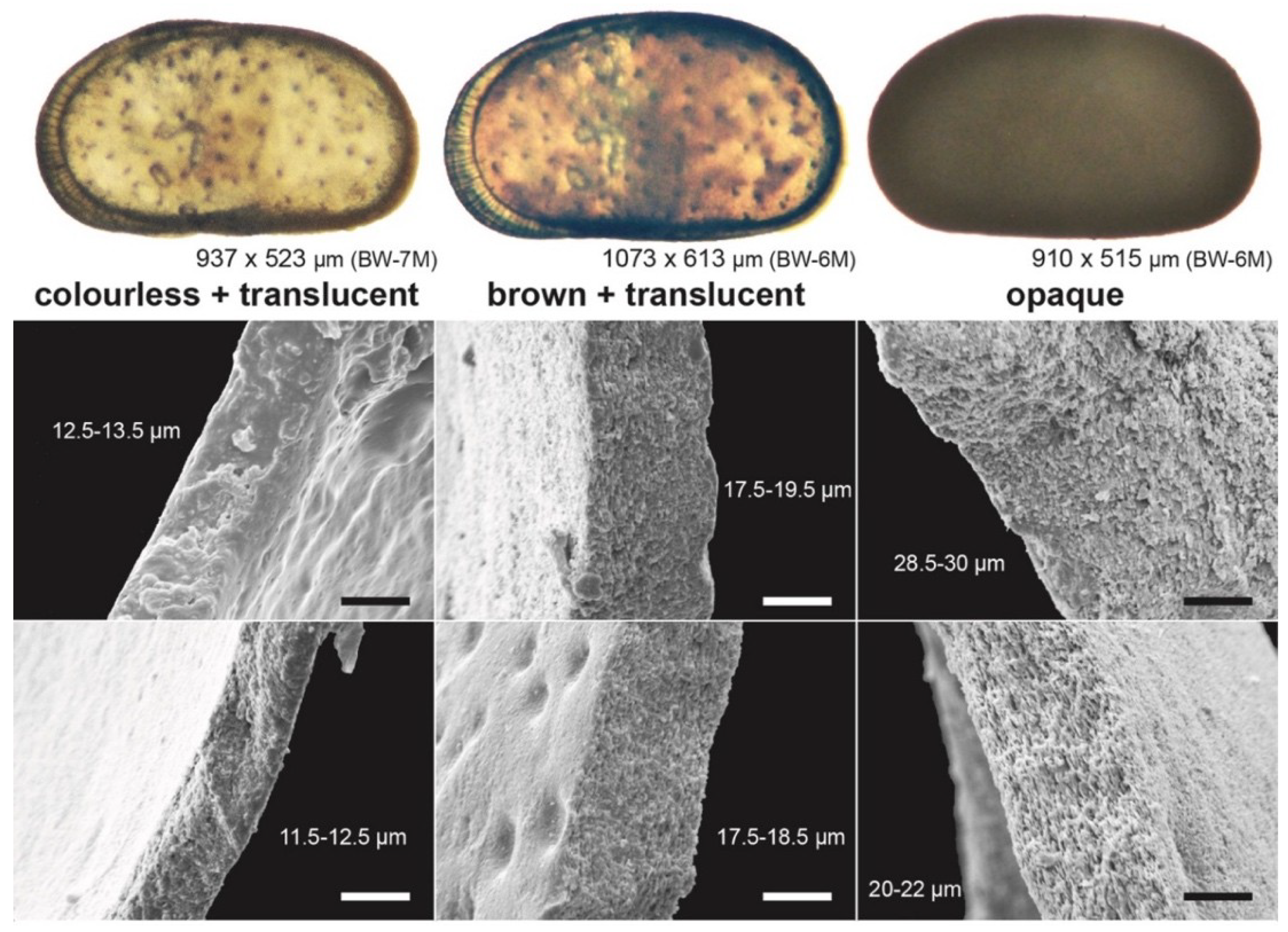
3.2. Valve Thickness
4. Discussion
4.1. Valve Size-Salinity Relationship
4.2. Valve Thickness
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Horne, D.J.; Cohen, A.; Martens, K. Taxonomy, morphology and biology of Quaternary and living Ostracoda. In The Ostracoda: Applications in Quaternary Research; Holmes, J.A., Chivas, A.R., Eds.; Geophysical Monograph Series; American Geophysical Union: Washington, DC, USA, 2002; Volume 131, pp. 5–36. [Google Scholar]
- Boomer, I.; Horne, D.J.; Slipper, I.J. The use of ostracods in palaeoenvironmental studies, or what can you do with an ostracod shell? Paleontol. Soc. Pap. 2003, 9, 153–180. [Google Scholar]
- Hartmann, G. Zur Morphologie und Ökologie rezenter Ostracoden und deren Bedeutung bei der Unterscheidung mariner und nichtmariner Sedimente. Fortschr. Geol. Rheinl. Westfal. 1963, 10, 67–80. [Google Scholar]
- Karanovic, I. Recent Freshwater Ostracods of the World: Crustacea, Ostracoda, Podocopida; Springer Science & Business Media: The Hague, The Netherlands, 2012. [Google Scholar]
- Heip, C. The life-cycle of Cyprideis torosa (Crustacea, Ostracoda). Oecologia 1976, 24, 229–245. [Google Scholar] [CrossRef] [PubMed]
- Turpen, J.B.; Angell, R.W. Aspects of molting and calcification in the ostracod Heterocypris. Boil. Bull. 1971, 140, 331–338. [Google Scholar] [CrossRef]
- De Deckker, P.; Forester, R.M. The use of ostracods to reconstruct continental palaeoenvironmental records. In Ostracoda in the Earth Sciences; Elsevier: Amsterdam, The Netherlands, 1988; pp. 175–199. [Google Scholar]
- Remane, A. Die Biologischen Grenzen Meer—Süßwasser und Meer—Land. Geol. Rundsch. 1958, 47, 11–24. [Google Scholar] [CrossRef]
- Keyser, D.; Walter, R. Calcification in ostracodes. Rev. Española Micropaleontol. 2004, 36, 1–11. [Google Scholar]
- Roca, J.R.; Wansard, G. Temperature influence on development and calcification of Herpetocypris brevicaudata Kaufmann, 1900 (Crustacea: Ostracoda) under experimental conditions. Hydrobiologia 1997, 347, 91–95. [Google Scholar] [CrossRef]
- Meisch, C. Crustacea: Ostracoda. In Süßwasserfauna von Mitteleuropa; Schwoerbel, J., Zwick, P., Eds.; Spektrum Akademischer Verlag: Heidelberg/Berlin, Germany, 2000; Volume 8, 522p. [Google Scholar]
- Van Harten, D. Size and environmental salinity in the modern euryhaline ostracod Cyprideis torosa (Jones, 1850), a biometrical study. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1975, 17, 35–48. [Google Scholar] [CrossRef]
- Van Harten, D. Cyprideis torosa (Ostracoda) revisited. Of salinity, nodes and shell size. In Proceedings of the Second European Ostracodologists’ Meeting; British Micropalaeontological Society: London, UK, 1996; Volume 191, p. 194. [Google Scholar]
- Van Harten, D. Variable noding in Cyprideis torosa (Ostracoda, Crustacea): An overview, experimental results and a model from Catastrophe Theory. Hydrobiologia 2000, 419, 131–139. [Google Scholar] [CrossRef]
- Mezquita, F.; Olmos, V.; Oltra, R. Population ecology of Cyprideis torosa (Jones, 1850) in a hypersaline environment of the Western Mediterranean (Santa Pola, Alacant) (Crustacea: Ostracoda). Ophelia 2000, 53, 119–130. [Google Scholar] [CrossRef]
- Dykan, N. Neogene–Quaternary Ostracodes of the Northern Part of the Black Sea; Publishing House “Chetverta Hvylja”: Kiev, Ukraine, 2016; 272p. [Google Scholar]
- Müller, G.W. Deutschlands Süßwasser-Ostracoden. In Originalabhandlungen aus dem Gesammtgebiete der Zoologie; Chun, C., Ed.; Zoologica 30, Schweizerbart Science Publishers: Leipzig, Germany, 1900. [Google Scholar]
- Van Morkhoven, F.P. Post-Palaeozoic Ostracoda: Their Morphology, Taxonomy, and Economic Use: 1. General; Elsevier: Amsterdam, The Netherlands, 1962; 204p. [Google Scholar]
- Frenzel, P.; Schulze, I.; Pint, A. Noding of Cyprideis torosa valves (Ostracoda)—A proxy for salinity? New data from field observations and a long-term microcosm experiment. Int. Rev. Hydrobiol. 2012, 97, 314–329. [Google Scholar] [CrossRef]
- Rosenfeld, A.; Vesper, B. The variability of the sieve-pores in Recent and fossil species of Cyprideis torosa (Jones, 1850) as an indicator for salinity and palaeosalinity. In Aspects of Ecology and Zoogeography of Recent and Fossil Ostracoda; Löffler, H., Danielopol, D., Eds.; Springer Science & Business Media: The Hague, The Netherlands, 1977; pp. 55–67. [Google Scholar]
- Boomer, I.; Frenzel, P.; Feike, M. Salinity-driven size variability in Cyprideis torosa (Ostracoda, Crustacea). J. Micropalaeontol. 2017, 36, 63–69. [Google Scholar] [CrossRef]
- Erginal, A.E.; Öztürk, M.Z. Kızılırmak Deltası plaj sırtlarının taramalı elektron mikroskobu analizleri ve tane boyu parametreleri ile incelenmesi (An assessment of beach ridges on the Kızılırmak delta based on scanning electron microscopy analyses and grain size parameters). Türk Coğrafya Derg. 2010, 54, 43–51. [Google Scholar]
- Berndt, C.; Yıldırım, C.; Çiner, A.; Strecker, M.; Ertunç, G.; Sarıkaya, M.A.; Özcan, O.; Ozturk, T.; Güneç Kiyak, N. Quaternary uplift of the northern margin of the Central Anatolian Plateau: New OSL dates of fluvial and delta-terrace deposits of the Kızılırmak River, Black Sea coast, Turkey. Quat. Sci. Rev. 2018, 201, 446–469. [Google Scholar] [CrossRef]
- Turoğlu, H. Kızılırmak Deltası ve Yakın Çevresinin Jeomorfolojik Özellikleri ve Insan Yaşamındaki Etkileri; Anadolu Araştırmaları; Istanbul University Publications: Istanbul, Turkey, 2010; Volume 4903, pp. 98–111. [Google Scholar]
- Ustaoğlu, M.R.; Mis, D.Ö.; Aygen, C. Observations on zooplankton in some lagoons in Turkey. J. Black Sea/Mediterr. Environ. 2012, 18, 208–222. [Google Scholar]
- Köse, B.; Ateș, S.; Çelik, H. Determination of late spring frost affections on some grape varieties grown in Samsun. Türkiye Tarımsal Araștırmalar Dergisi 2014, 1, 162–169. [Google Scholar]
- Berndt, C.; Frenzel, P.; Çiner, A.; Ertunç, G.; Yıldırım, C. Holocene marginal marine ostracod successions from the Kızılırmak River delta; Implications for depositional environments and sea level changes at the southern Black Sea coast. Sediment. Geol. 2019, in press. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Bradley, P.S. The shell structure of the Ostracoda and its Application to their Palaeontological investigation. Ann. Mag. Nat. Hist. 1941, 8, 1–33. [Google Scholar] [CrossRef]
- Rothwell, R.G.; Croudace, I.W. Twenty years of XRF core scanning marine sediments: What do geochemical proxies tell us? In Micro-XRF Studies of Sediment Cores; Croudace, I.W., Rothwell, R.G., Eds.; Springer: Dordrecht, The Netherlands, 2015; pp. 25–102. [Google Scholar]
- Martens, K. Effects of temperature and salinity on postembryonic growth in Mytilocypris henricae (Chapman) (Crustacea, Ostracoda). J. Crustac. Boil. 1985, 5, 258–272. [Google Scholar] [CrossRef]
- De Deckker, P. Ostracod palaeoecology. In The Ostracoda: Applications in Quaternary Research; Holmes, J.A., Chivas, A.R., Eds.; Geophysical Monograph Series; American Geophysical Union: Washington, DC, USA, 2002; Volume 131, pp. 121–134. [Google Scholar]
- Aladin, N.V. Osmoregulation in Cyprideis torosa from various seas of the USSR. Zool. Zhurnal 1989, 68, 40–50. [Google Scholar]
- Den Hartog, C. Typologie des Brackwassers. Helgoländer Wiss. Meeresunters. 1964, 10, 377. [Google Scholar] [CrossRef]
- Hamburger, K.; Møhlenberg, F.; Randløv, A.; Riisgård, H.U. Size, oxygen consumption and growth in the mussel Mytilus edulis. Mar. Boil. 1983, 75, 303–306. [Google Scholar] [CrossRef]
- McClain, C.; Rex, M. The relationship between dissolved oxygen concentration and maximum size in deep-sea turrid gastropods: An application of quantile regression. Mar. Boil. 2001, 139, 681–685. [Google Scholar]
- Delile, H.; Mazzini, I.; Blichert-Toft, J.; Goiran, J.P.; Arnaud-Godet, F.; Albarede, F.; Keay, S. Definition of a new approach in ancient harbor geoarchaeology: Geochemistry and ostracod analyses at Portus (Tiber delta, central Italy). In Proceedings of the 17th International Symposium on Ostracoda, Rome, Italy, 23–26 July 2013; Volume 37, pp. 103–105. [Google Scholar]
- Berndt, C. Combined Micropalaeontological and Heavy Mineral Investigation of Holocene Sediments from Elaia, the Ancient Harbour City of Pergamum (Turkey). Master’s Thesis, Institute of Earth Sciences, Friedrich Schiller University Jena, Jena, Germany, 2014. [Google Scholar]
- Remane, A. Die Brackwasserfauna. Verh. der Deutsch. Zool. Ges. 1934, 36, 34–74. [Google Scholar]
- Trahms, O.K. Beiträge zur Ökölogie küstennaher Brackwasser. 2. Die Bodenfauna und Bodenflora des Großen Jasmunder Boddens. Arch. Hydrobiol. 1939, 36, 1–35. [Google Scholar]
- Scott, H.W. Shell morphology of Ostracoda. Arthropoda 1961, 3, 21–37. [Google Scholar]
- Carbonel, P. Ostracods and the transition between fresh and saline waters. In Ostracoda in the Earth Sciences; Elsevier: Amsterdam, The Netherlands, 1988; pp. 157–173. [Google Scholar]
- Carbonel, P.; Pinson, J. Les Cyprideis Témoins de Lévolution des Sels en Milieu Laguno-Lacustre Sous Climat Semi-Aride. In Taxonomy, Biostratigraphy and Distribution of Ostracodes; Krstic, N., Ed.; Serbian Geological Society: Belgrade, SFR Yugoslavia, 1979; pp. 211–217. [Google Scholar]
- Carbonel, P. Les Ostracodes et Leur Intérêt Dans la Définition des Écosystèmes Estuariens et de Plateforme Continentale: Essais D’application à des Domaines Anciens; Mémoires de l’Institut de Géologie du Bassin d’Aquitaine, Université de Bordeaux I: Bordeaux, France, 1980; Volume 11. [Google Scholar]
- Frear, G.L.; Johnston, J. The solubility of calcium carbonate (calcite) in certain aqueous solutions at 25°. J. Am. Chem. Soc. 1929, 51, 2082–2093. [Google Scholar] [CrossRef]
- Kaesler, R.L.; Kontrovitz, M.; Taunton, S. Crushing strength of Puriana pacifica (Ostracoda), an experimental approach to taphonomy. J. Paleontol. 1993, 67, 1005–1010. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Berndt, C.; Frenzel, P.; Çiner, A. Intraspecific Length Variation and Shell Thickness of the Ostracod Cyprideis torosa (Jones, 1850) as a Potential Tool for Palaeosalinity Characterization. Geosciences 2019, 9, 83. https://doi.org/10.3390/geosciences9020083
Berndt C, Frenzel P, Çiner A. Intraspecific Length Variation and Shell Thickness of the Ostracod Cyprideis torosa (Jones, 1850) as a Potential Tool for Palaeosalinity Characterization. Geosciences. 2019; 9(2):83. https://doi.org/10.3390/geosciences9020083
Chicago/Turabian StyleBerndt, Christopher, Peter Frenzel, and Attila Çiner. 2019. "Intraspecific Length Variation and Shell Thickness of the Ostracod Cyprideis torosa (Jones, 1850) as a Potential Tool for Palaeosalinity Characterization" Geosciences 9, no. 2: 83. https://doi.org/10.3390/geosciences9020083