Stress-Tolerance and Taxonomy of Culturable Bacterial Communities Isolated from a Central Mojave Desert Soil Sample

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil Sampling
2.2. Epifluorescent Microscopy
2.3. Bacteria Culturing and Isolation
2.4. Amplification and Sequencing of 16S rRNA Genes
2.5. Physiological Assays
2.6. Multisubstrate Testing
2.7. Data Analysis
3. Results
3.1. Cell Numbers
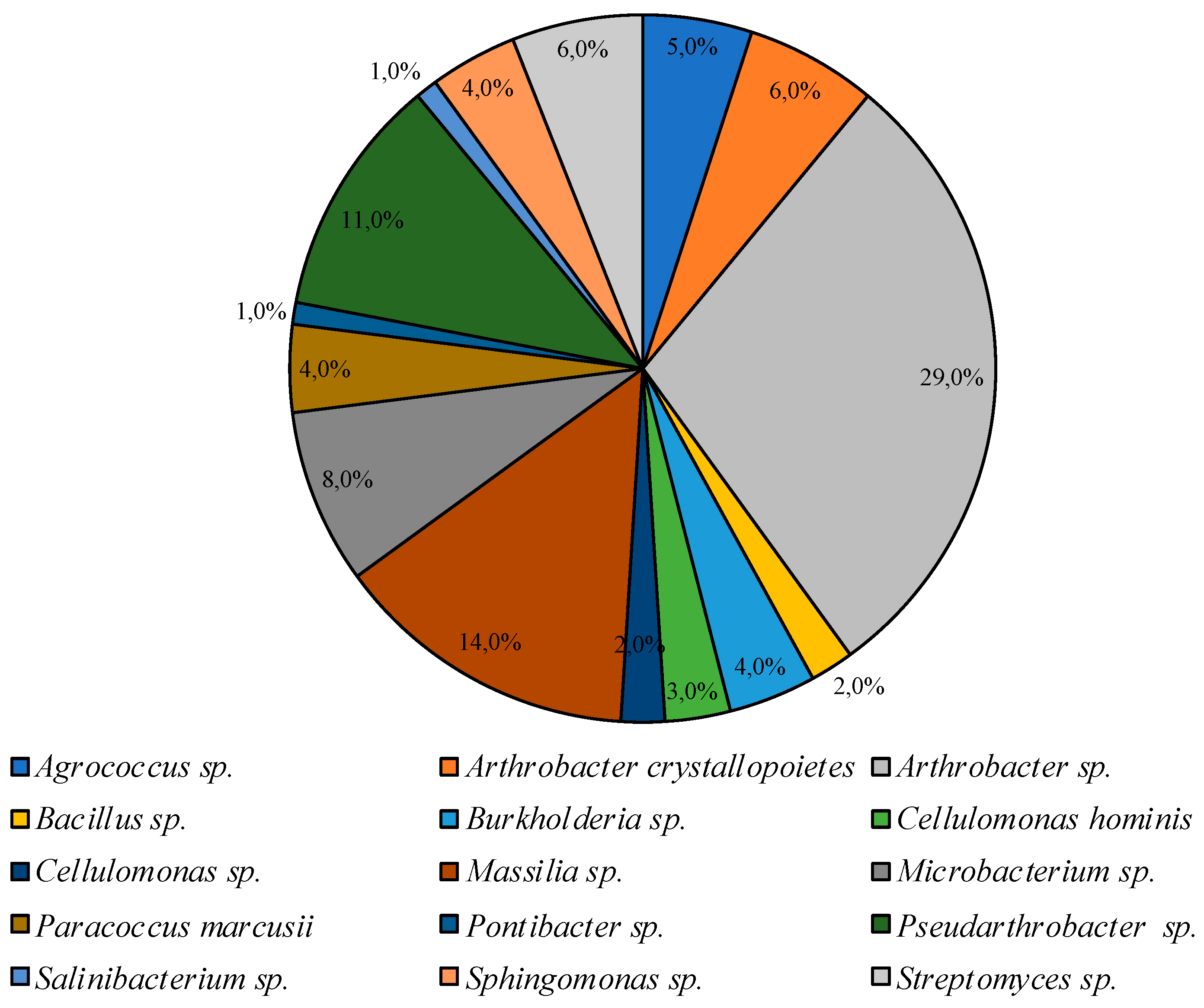
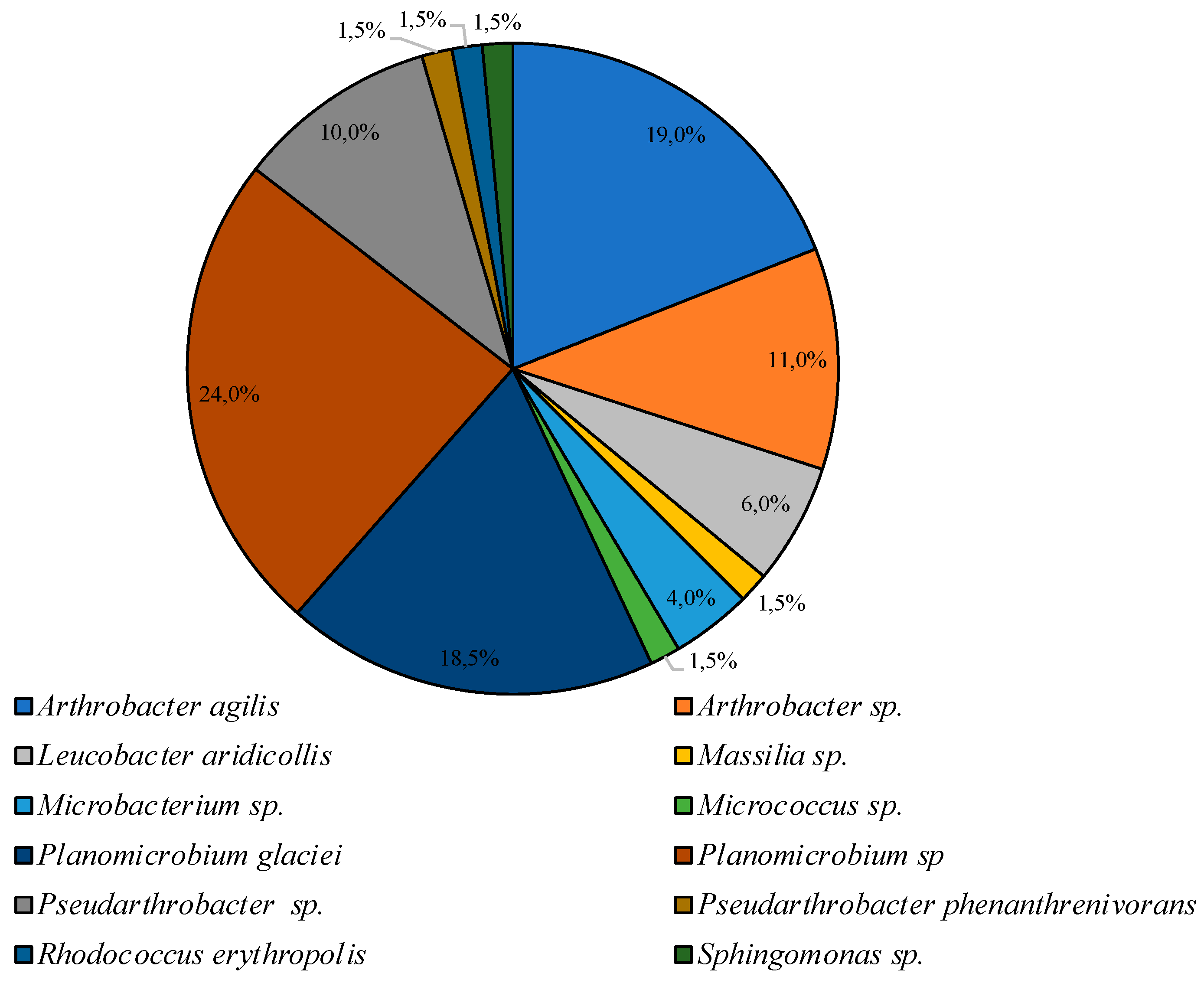
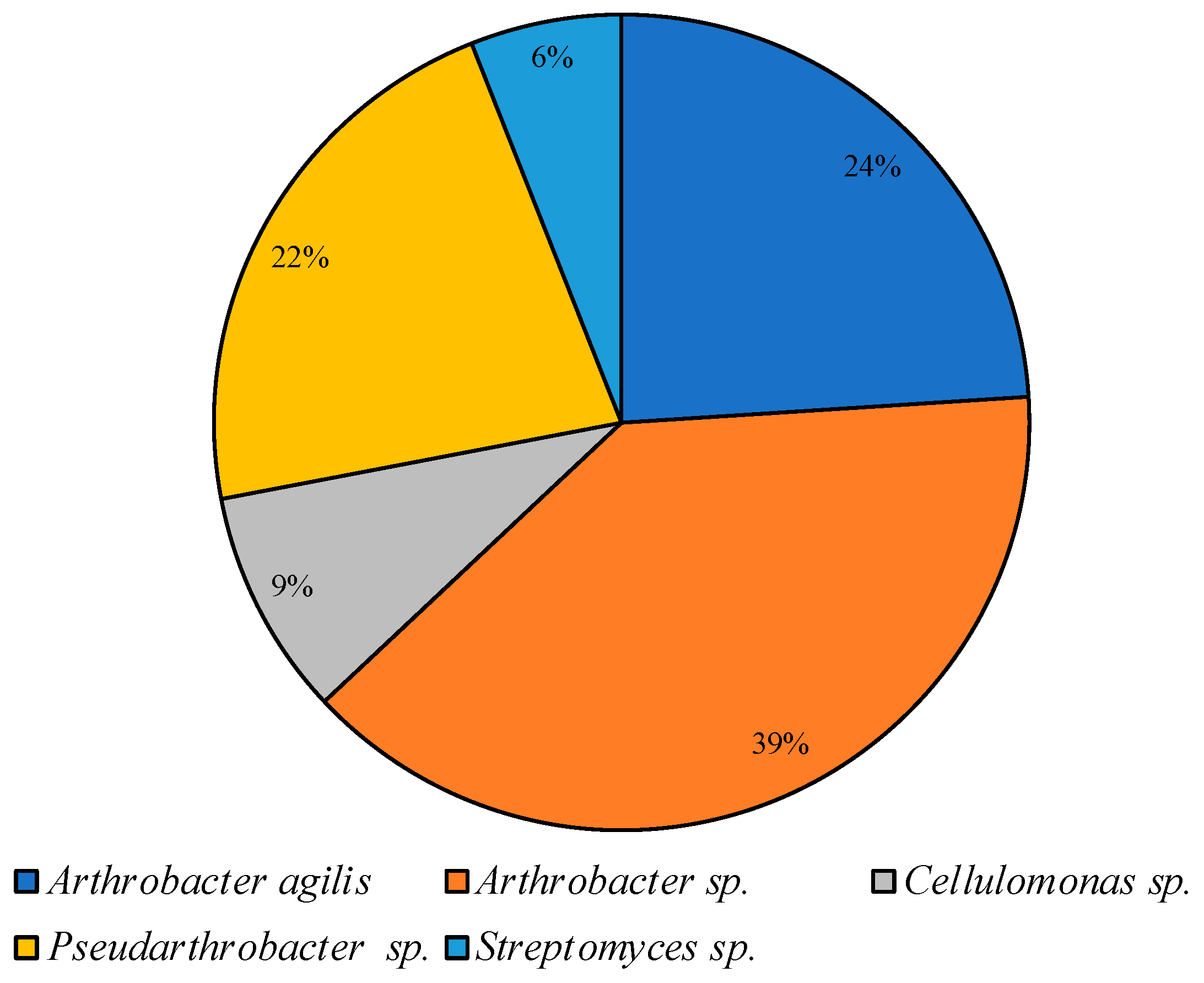
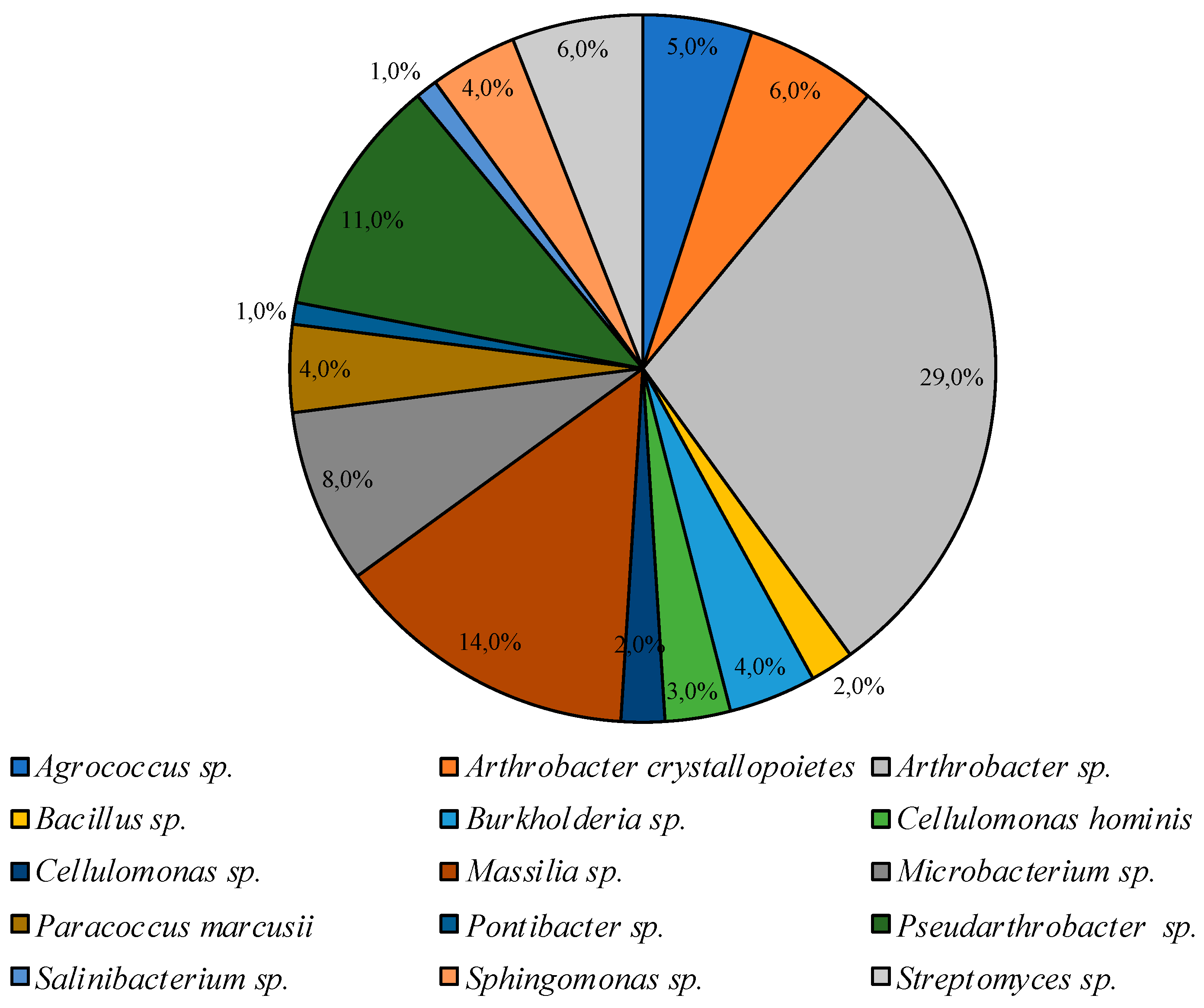
3.2. Culturable Communities’ Structure
3.3. Multisubstrate Testing
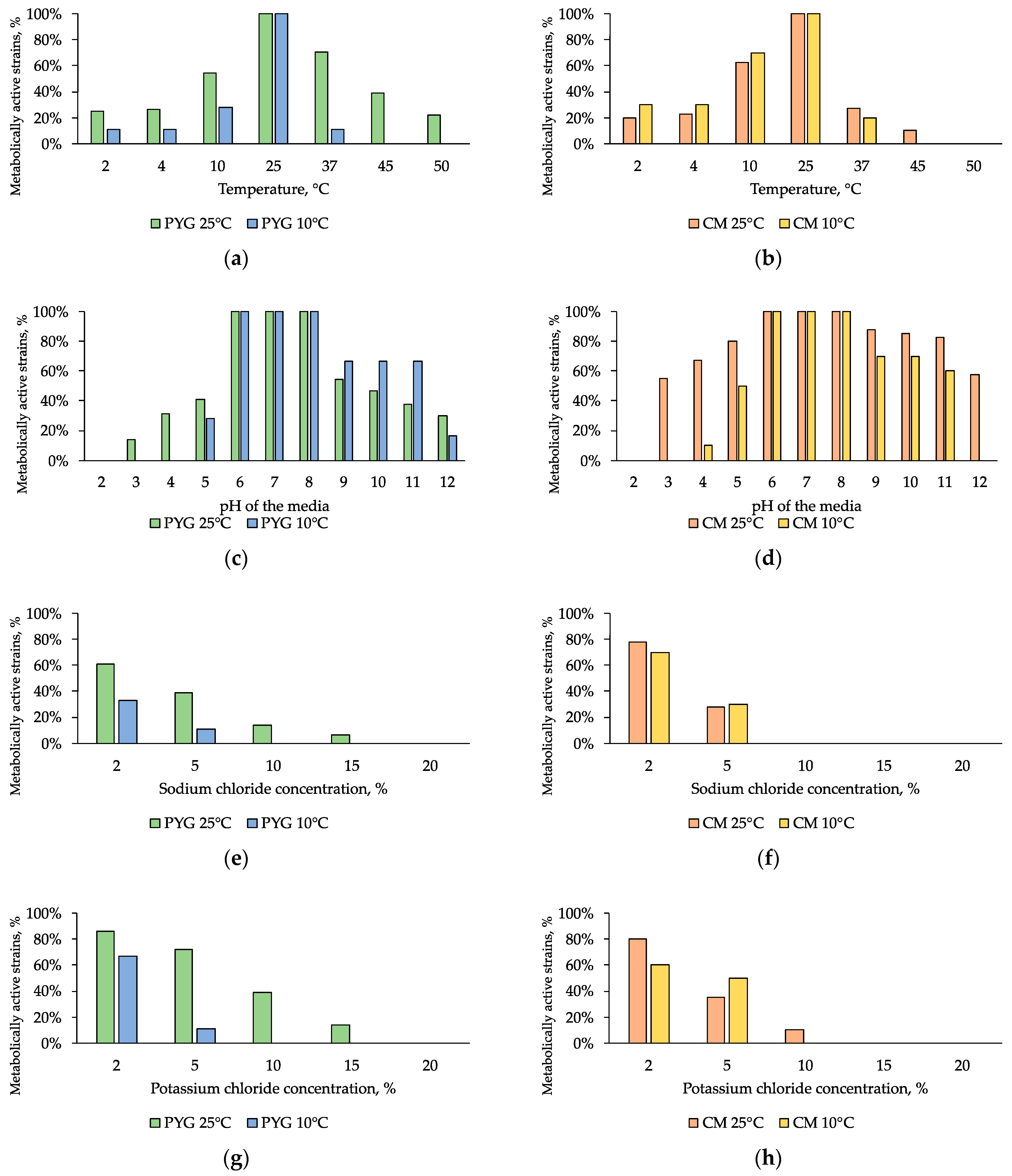
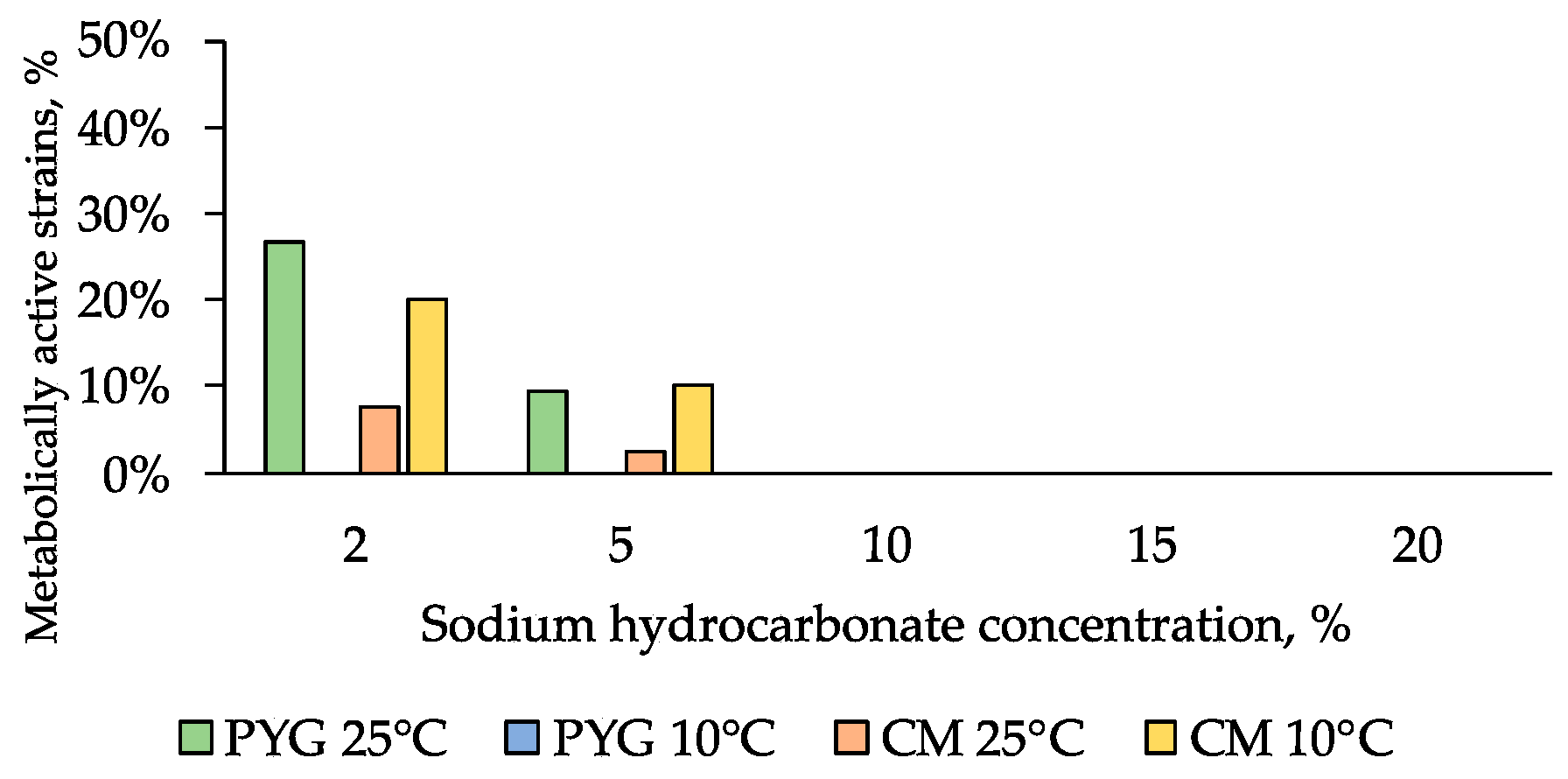
3.4. Physiological Assays
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain–Isolation Conditions–GenBank Accession Number | BLAST Search Results | Taxonomic Affiliation |
|---|---|---|
| KBP.AS.17–PYG 25 °C–MK403789 | Pseudarthrobacter siccitolerans [MF682005]—99.0% Pseudarthrobacter oxydans [MF681862]—99.0% Pseudarthrobacter oxydans [MF681851]—99.0% | Pseudarthrobacter oxydans |
| KBP.AS.62–PYG 25 °C–MK403790 | Arthrobacter sp. [KX928444]—99.5% Arthrobacter sp. [KT944733]—99.5% Arthrobacter sp. [KM507593]—99.5% | Arthrobacter sp. |
| KBP.AS.75–PYG 25 °C–MK403794 | Brachybacterium sp. [MH518254]—99.3% Brachybacterium paraconglomeratum [KY622837]—99.3% Brachybacterium conglomeratum [LN995481]—99.3% | Brachybacterium sp. |
| KBP.AS.105–PYG 25 °C–MK403787 | Janthinobacterium sp. [KT715033]—98.7% Janthinobacterium sp. [FJ605429]—98.7% Janthinobacterium sp. [FJ605404]—98.7% | Janthinobacterium sp. |
| KBP.AS.160–PYG 25 °C–MK403788 | Arthrobacter sp. [HM216918]—99.2% Uncultured Arthrobacter sp. [JN129287]—99.0% Arthrobacter agilis [JN009621]—99.0% | Arthrobacter sp. |
| KBP.AS.747–PYG 25 °C–MK403791 | Microbacterium aurantiacum [MH734538]—99.2% Microbacterium aurantiacum [MH715198]—99.2% Microbacterium sp. [MH702463]—99.2% | Microbacterium aurantiacum |
| KBP.AS.748–PYG 25 °C–MK403792 | Pseudarthrobacter siccitolerans [MF682005]—99.22% Pseudarthrobacter oxydans [MF681862]—99.2% Pseudarthrobacter oxydans [MF681851]—99.2% | Pseudarthrobacter sp. |
| KBP.AS.749–PYG 25 °C–MK403793 | Brevibacterium frigoritolerans [MK318239]—99.9% Brevibacterium frigoritolerans [MK318233]—99.9% Brevibacterium frigoritolerans [MK318218]—99.9% | Brevibacterium frigoritolerans |
| KBP.AS.750–PYG 25 °C–MK403795 | Planomicrobium okeanokoites [KF749386]—99.3% Planomicrobium okeanokoites [KF749394]—99.3% Planomicrobium okeanokoites [KF724965]—99.3% | Planomicrobium okeanokoites |
| KBP.AS.751–PYG 25 °C–MK403796 | Agrococcus lahaulensis [KF318382]—100% Agrococcus sp. [JX949729]—100% Agrococcus jenensis [EF672044]—100% | Agrococcus sp. |
| KBP.AS.752–PYG 25 °C–MK403797 | Agrococcus sp. [KC702716]—99.5% Agrococcus sp. [JX949729]—99.5% Agrococcus jenensis [EF672044]—99.4% | Agrococcus sp. |
| KBP.AS.753–PYG 25 °C–MK403798 | Agrococcus sp. [MG232346]—99.5% Agrococcus jenensis [AJ717350]—99.5% Agrococcus sp. [HF954438]—99.5% | Agrococcus sp. |
| KBP.AS.754–PYG 25 °C–MK403799 | Kocuria sp. [DQ192212]—99.3% Kocuria sp. [EF612289]—99.2% Kocuria sp. [DQ180950]—99.0% | Kocuria sp. |
| KBP.AS.755–PYG 25 °C–MK403800 | Planomicrobium glaciei [KF387709]—100% Planomicrobium sp. [JX949538]—100% Planomicrobium glaciei HQ232419]—100% | Planomicrobium glaciei |
| KBP.AS.756–PYG 25 °C–MK403801 | Leucobacter sp. [KY623368]—99.6% Leucobacter aridicollis [KR827428]—99.6% Leucobacter sp. [KP152582]—99.6% | Leucobacter aridicollis |
| KBP.AS.757–PYG 25 °C–MK403802 | Planomicrobium okeanokoites [KF724965]—99.5% Planomicrobium okeanokoites [KF749394]—99.5% Planomicrobium okeanokoites [KF749386]—99.5% | Planomicrobium okeanokoites |
| KBP.AS.758–PYG 25 °C–MK403803 | Microbacterium esteraromaticum [GU111572]—98.9% Microbacterium sp. [KU221074]—98.6% Microbacterium sp. [KU221073]—98.6% | Microbacterium sp. |
| KBP.AS.759–PYG 25 °C–MK403804 | Bacillus pumilus [MH910167]—99.9% Bacillus pumilus [MH910156]—99.9% Bacillus pumilus [MH908782]—99.9% | Bacillus pumilus |
| KBP.AS.760–PYG 25 °C–MK403805 | Microbacterium esteraromaticum [GU111572]—98.9% Microbacterium sp. MH231521]—98.2% Microbacterium sp. [MH231520]—98.2% | Microbacterium sp. |
| KBP.AS.761–PYG 25 °C–MK403806 | Rufibacter sp. [MH050957]—97.6% Rufibacter immobilis [MH050949]—97.6% Uncultured bacterium clone [JQ376150]—98.9% | Rufibacter sp. |
| KBP.AS.762–PYG 25 °C–MK403807 | Microbacterium esteraromaticum [GU111572]—99.2% Microbacterium sp. [MH231521]—98.5% Microbacterium sp. [MH231520]—98.5% | Microbacterium sp. |
| KBP.AS.763–PYG 25 °C–MK403808 | Massilia niabensis [NR_044571]—98.9% Oxalobacter sp. [DQ196473]—99.2% Massilia suwonensis [LN774622]—98.8% | Massilia sp. |
| KBP.AS.764–PYG 25 °C–MK403809 | Bacillus sp. [MF990764]—98.2% Bacillus pumilus [JX988407]—97.8% Bacillus pumilus [EU795022]—97.8% | Bacillus sp. |
| KBP.AS.765–PYG 25 °C–MK403810 | Massilia alkalitolerans [KY010279]—99.7% Uncultured bacterium clone [KU515079]—99.7% Massilia alkalitolerans [KP282807]—99.7% | Massilia alkalitolerans |
| KBP.AS.766–PYG 25 °C–MK403811 | Pseudarthrobacter siccitolerans [MF682005]—98.1% Pseudarthrobacter oxydans [MF681862]—98.1% Pseudarthrobacter oxydans [MF681851]—98.1% | Pseudarthrobacter sp. |
| KBP.AS.767–PYG 25 °C–MK403812 | Massilia varians [MF077216]—98.4% Massilia varians [MF077215]—98.4% Massilia varians [MF077214]—98.5% | Massilia varians |
| KBP.AS.768–PYG 25 °C–MK403813 | Planococcus donghaensis [LN774491]—100% Planomicrobium okeanokoites [KF749394]—100% Planomicrobium okeanokoites [KF749386]—100% | Planomicrobium sp. |
| KBP.AS.769–PYG 25 °C–MK403814 | Microbacterium pseudoresistens [LN890047]—98.9% Microbacterium pseudoresistens [KF687021]—98.9% Microbacterium sp. [MH671545]—98,2% | Microbacterium pseudoresistens |
| KBP.AS.770–PYG 25 °C–MK403815 | Arthrobacter agilis [KF924209]—99.3% Arthrobacter sp. [JX949695]—99.3% Arthrobacter sp. [JX949646]—99.3% | Arthrobacter sp. |
| KBP.AS.771–PYG 25 °C–MK403816 | Arthrobacter sp. [MH714643]—99.5% Arthrobacter sp. [MG757950]—99.5% Arthrobacter sp. [JX949321]—99.5% | Arthrobacter sp. |
| KBP.AS.772–PYG 25 °C–MK403817 | Planomicrobium glaciei [MG025800]—100% Planomicrobium glaciei [KF387709]—100% Planomicrobium sp. [JX949538]—100% | Planomicrobium glaciei |
| KBP.AS.773–PYG 25 °C–MK403818 | Uncultured bacterium [LC026838]—98.9% Cellulomonas sp. [KP126821]—98.4% Cellulomonas cellasea [KR922256]—98.4% | Cellulomonas sp. |
| KBP.AS.774–PYG 25 °C–MK403819 | Planomicrobium okeanokoites [KF749394]—99.8% Planomicrobium okeanokoites [KF749386]—99.8% Planomicrobium okeanokoites [KF724965]—99.8% | Planomicrobium okeanokoites |
| KBP.AS.775–PYG 25 °C–MK403820 | Planomicrobium okeanokoites [KF724965]—99.9% Planomicrobium okeanokoites [KF749394]—99.9% Planomicrobium okeanokoites [KF749386]—99.9% | Planomicrobium okeanokoites |
| KBP.AS.776–PYG 25 °C–MK403821 | Georgenia sp. [EF512742]—99.6% Georgenia sp. [AY880044]—99.5% Georgenia satyanarayanai [NR_117051]—99.5% | Georgenia sp. |
| KBP.AS.777–PYG 25 °C–MK403822 | Microbacterium barkeri [MG706019]—98.4% Microbacterium sp. [MG203882]—98.4% Microbacterium barkeri [KY928100]—98.4% | Microbacterium barkeri |
| KBP.AS.778–PYG 25 °C–MK403823 | Salinibacterium sp. [MH714650]—99.0% Salinibacterium sp. [MH790149]—99.0% Salinibacterium sp. [MH299814]—99.0% | Salinibacterium sp. |
| KBP.AS.779–PYG 25 °C–MK403824 | Brevibacterium frigoritolerans [MK318239]—100% Brevibacterium frigoritolerans [MK318233]—100% Brevibacterium frigoritolerans [MK318218]—100% | Brevibacterium frigoritolerans |
| KBP.AS.780–PYG 25 °C–MK403825 | Micrococcus sp. [JQ229694]—99.3% Micrococcus sp. [JQ229689]—99.3% Micrococcus sp. [KM613156]—99.2% | Micrococcus sp. |
| KBP.AS.781–PYG 25 °C–MK403826 | Kocuria sp. [KT900591]—98.9% Kocuria sediminis [MH178354]—98.85% Kocuria turfanensis [MF769333]—98.8% | Kocuria sp. |
| KBP.AS.782–PYG 25 °C–MK403827 | Labedella sp. [KM253033]—99.2% Labedella gwakjiensis [LK021191]—98.2% Labedella sp. [JX273669]—98.2% | Labedella sp. |
| KBP.AS.783–PYG 25 °C–MK403828 | Plantibacter flavus [CP019402]—100% Plantibacter sp. [HE662660]—100% Plantibacter sp. [HE662693]—100% | Plantibacter sp. |
| KBP.AS.784–PYG 25 °C–MK403829 | Arthrobacter agilis [KT804924]—99.9% Arthrobacter agilis [MF767263]—99.8% Arthrobacter agilis [KR811202]—99.8% | Arthrobacter agilis |
| KBP.AS.785–PYG 25 °C–MK403830 | Microbacterium esteraromaticum [GU111572]—99.1% Microbacterium aureliae [NR152652]—98.6% Microbacterium aureliae [KF793922]—98.6% | Microbacterium sp. |
| KBP.AS.786–PYG 25 °C–MK403831 | Pseudarthrobacter siccitolerans [MF682005]—99.0% Pseudarthrobacter oxydans [MF681862]—99.0% Pseudarthrobacter oxydans [MF681851]—99.0% | Pseudarthrobacter sp. |
| KBP.AS.787–CM 25 °C–MK403832 | Uncultured bacterium [KX507906]—98.2% Massilia niabensis [JF496256]—98.0% Massilia alkalitolerans [KY010279]—97.8% | Massilia sp. |
| KBP.AS.788–CM 25 °C–None | Was identified by morphology | Bacillus sp. |
| KBP.AS.789–CM 25 °C–MK403833 | Burkholderia sp. [KU060146] —95.7% Burkholderia sp. [KU060136]—95.7% Burkholderia sp. [KU060135]—95.7% | Burkholderia sp. |
| KBP.AS.790–CM 25 °C–MK403834 | Cellulomonas hominis [KF817811]—100% Cellulomonas hominis [MF928361]—100% Cellulomonas sp. [LC133615]—100% | Cellulomonas hominis |
| KBP.AS.791–CM 25 °C–MK403835 | Pseudarthrobacter siccitolerans [MF682005]—99.5% Pseudarthrobacter oxydans [MF681862]—99.5% Pseudarthrobacter oxydans [MF681851]—99.5% | Pseudarthrobacter sp. |
| KBP.AS.792–CM 25 °C–MK403836 | Massilia sp. [JQ511858]—100% Massilia sp. [KY635898]—99.6% Massilia sp. [KY635895]—99.6% | Massilia sp. |
| KBP.AS.793–CM 25 °C–MK403837 | Uncultured actinobacterium clone [KC994764]—98.8% Salinibacterium sp. [MH299814]—98.7% Salinibacterium sp. [CP026951]—98.7% | Salinibacterium sp. |
| KBP.AS.794–CM 25 °C–MK403838 | Arthrobacter sp. [LN871743]—99.7% Arthrobacter sp. [KU951449]—99.7% Arthrobacter sp. [KJ191079]—99.7% | Arthrobacter sp. |
| KBP.AS.795–CM 25 °C–MK403839 | Arthrobacter sp. [LN871743]—99.2% Arthrobacter sp. [KU951449]—99.2% Arthrobacter sp. [KJ191079]—99.2% | Arthrobacter sp. |
| KBP.AS.816–PYG 25 °C–MK403840 | Arthrobacter sp. [KJ191079]—99.8% Arthrobacter sp. [KJ191059]—99.8% Arthrobacter sp. [KJ191035]—99.8% | Arthrobacter sp. |
| KBP.AS.842–PYG 25 °C–MK403841 | Uncultured bacterium clone [KX507404]—99.1% Massilia suwonensis [NR116872]—99.1% Massilia sp. [KM187344]—98.8% | Massilia sp. |
| KBP.AS.843–PYG 25 °C–MK403842 | Mycetocola sp. [MH299815]—98.4% Mycetocola sp. [MH298813]—98.4% Mycetocola sp. [CP026949]—98.4% | Mycetocola sp. |
| KBP.AS.844–PYG 25 °C–MK403843 | Arthrobacter agilis [KT804924]—98.9% Arthrobacter agilis [MF767263]—98.8% Arthrobacter agilis [KR811202]—98.8% | Arthrobacter agilis |
| KBP.AS.845–PYG 25 °C–None | Was identified by morphology | Streptomyces sp. |
| KBP.AS.846–PYG 25 °C–MK403844 | Arthrobacter pascens [KY880876]—100% Pseudarthrobacter defluvii [KY880845]—100% Arthrobacter sp. [LM994042]—100% | Arthrobacter sp. |
| KBP.AS.847–CM 25 °C–MK403845 | Cellulomonas sp. [MH046844]—99.2% Cellulomonas sp. [HM587947]—99.2% Cellulomonas flavigena [KY951360]—99.0% | Cellulomonas sp. |
| KBP.AS.848–PYG 25 °C–MK403846 | Massilia alkalitolerans [KY010279]—99.6% Uncultured bacterium clone [KU515079]—99.6% Massilia alkalitolerans [KP282807]—99.6% | Massilia sp. |
| KBP.AS.849–PYG 25 °C–MK403847 | Microbacterium esteraromaticum [GU111572]—98.8% Microbacterium aureliae [NR152652]—98.2% Microbacterium aureliae [KF793922]—98.2% | Microbacterium sp. |
| KBP.AS.850–PYG 25 °C–MK403848 | Massilia sp. [MH734560]—99.6% Massilia sp. [KY635898]—99.6% Massilia sp. [KY635895]—99.6% | Massilia sp. |
| KBP.AS.854–CM 25 °C–MK403849 | Paracoccus marcusii [MH744726]—99.9% Paracoccus marcusii [MH725412]—99.9% Paracoccus marcusii [MH725411]—99.9% | Paracoccus marcusii |
| KBP.AS.855–PYG 25 °C–None | Were identified by morphology | Rufibacter sp. |
| KBP.AS.858–CM 25 °C–MK403850 | Arthrobacter crystallopoietes [CP018864]—98.7% Arthrobacter crystallopoietes [CP018863]—98.7% Arthrobacter sp. [KX645729]—98.7% | Arthrobacter crystallopoietes |
| KBP.AS.862–PYG 25 °C–MK403851 | Arthrobacter sulfonivorans [KR233773]—99.9% Uncultured soil bacterium clone [JX490026]—99.8% Uncultured bacterium clone [KC554813]—99.7% | Arthrobacter sp. |
| KBP.AS.866–CM 25 °C–MK403852 | Uncultured bacterium clone [EU647527]—98.8% Sphingomonas xinjiangensis [NR_108386]—98.6% Sphingomonas mucosissima [KM502884]—98.3% | Sphingomonas sp. |
| KBP.AS.867–PYG 25 °C–MK403853 | Microbacterium paraoxydans [MH819720]—99.9% Microbacterium paraoxydans [MH281749]—99.9% Microbacterium sp. [MH558380]—99.9% | Microbacterium paraoxydans |
| KBP.AS.868–PYG 25 °C–MK403854 | Pseudarthrobacter siccitolerans [MF682005]—99.2% Pseudarthrobacter oxydans [MF681862]—99.2% Pseudarthrobacter oxydans [MF681851]—99.2% | Pseudarthrobacter sp. |
| KBP.AS.869–PYG 25 °C–MK403855 | Kocuria rosea [KY194315]—98.8% Kocuria sp. [KY194287]—98.8% Kocuria turfanensis [KY194236]—98.8% | Kocuria sp. |
| KBP.AS.876–PYG 25 °C–MK403856 | Arthrobacter agilis [CP024915]—99.1% Arthrobacter agilis [MG279727]—99.1% Arthrobacter sp. [KJ191029]—99.1% | Arthrobacter agilis |
| KBP.AS.877–PYG 25 °C–MK403857 | Salinibacterium sp. [MH790149]—99.0% Salinibacterium sp. [MH299814]—99.0% Salinibacterium sp. [CP026951]—99.0% | Salinibacterium sp. |
| KBP.AS.899–CM 25 °C–MK403858 | Streptomyces sp. [MG930074]—100% Streptomyces sp. [MG930073]—100% Streptomyces atrovirens [MF662228]—100% | Streptomyces sp. |
| KBP.AS.916–PYG 10 °C–MK403859 | Planomicrobium glaciei [MG025800]—99.1% Planomicrobium glaciei [KY307898]—99.1% Uncultured bacterium clone [KU514918]—99.1% | Planomicrobium glaciei |
| KBP.AS.926–PYG 10 °C–MK403860 | Sphingomonas xinjiangensis [NR108386]—98.9% Uncultured bacterium clone [EU647527]—98.6% Sphingomonas mucosissima [KM502884]—98.6% | Sphingomonas sp. |
| KBP.AS.927–PYG 10 °C–MK403861 | Arthrobacter agilis [MG279728]—99.4% Arthrobacter agilis [MF767263]—99.4% Arthrobacter agilis [KU884338]—99.4% | Arthrobacter agilis |
| KBP.AS.928–PYG 10 °C–MK403862 | Uncultured bacterium [KX509715]—99.5% Uncultured bacterium [KX508022]—99.5% Massilia niabensis [NR044571]—99.5% | Massilia sp. |
| KBP.AS.929–CM 10 °C–MK403863 | Pseudarthrobacter siccitolerans [MF682005]—98.5% Pseudarthrobacter oxydans [MF681862]—98.5% Pseudarthrobacter oxydans [MF681851]—98.5% | Pseudarthrobacter sp. |
| KBP.AS.930–CM 10 °C–MK403864 | Pseudarthrobacter siccitolerans [MF682005]—98.7% Pseudarthrobacter oxydans [MF681862]—98.7% Pseudarthrobacter oxydans [MF681851]—98.7% | Pseudarthrobacter sp. |
| KBP.AS.931–CM 10 °C–MK403865 | Arthrobacter boritolerans [MH828349]—99.7% Paenarthrobacter sp. [MH734752]—99.7% Arthrobacter sp. [LC416398]—99.7% | Arthrobacter sp. |
| KBP.AS.932–CM 10 °C–MK403866 | Pseudarthrobacter sp. [MG860452]—99.0% Pseudarthrobacter sp. [MG860451]—99.0% Arthrobacter sp. [MG860340]—99.0% | Pseudarthrobacter sp. |
| KBP.AS.936–PYG 10 °C–MK403867 | Pseudarthrobacter phenanthrenivorans [MF077156]—99.8% Pseudarthrobacter phenanthrenivorans [MF077155]—99.8% Arthrobacter phenanthrenivorans [KC789777]—99.8% | Pseudarthrobacter phenanthrenivorans |
| KBP.AS.937–PYG 10 °C–MK403868 | Arthrobacter agilis [KF876857]—99.9% Arthrobacter agilis [KT763365]—99.8% Uncultured Arthrobacter sp. [JN129287]—99.7% | Arthrobacter agilis |
| KBP.AS.938–CM 25 °C–MK403869 | Streptomyces sp. [KU182931]—99.8% Streptomyces sp. [MG820052]—99.6% Streptomyces sp. [MG930074]—99.6% | Streptomyces sp. |
| KBP.AS.939–CM 25 °C–MK403870 | Uncultured bacterium clone [KU515164]—99.0% Uncultured bacterium clone [KX509026]—98.9% Massilia sp. [MG733567]—98.6% | Massilia sp. |
| KBP.AS.940–CM 25 °C–MK403871 | Microbacterium esteraromaticum [GU111572]—99.2% Microbacterium sp. [MH231521]—98.8% Microbacterium sp. [KU221074]—98.8% | Microbacterium sp. |
| KBP.AS.941–CM 25 °C–MK403872 | Pseudarthrobacter siccitolerans [MF682005]—99.4% Pseudarthrobacter oxydans [MF681862]—99.4% Pseudarthrobacter oxydans [MF681851]—99.4% | Pseudarthrobacter sp. |
| KBP.AS.943–CM 10 °C–MK403873 | Arthrobacter agilis [JQ684255]—99.8% Arthrobacter agilis [CP024915]—99.7% Arthrobacter agilis [MG279727]—99.7% | Arthrobacter agilis |
| KBP.AS.947–PYG 10 °C–MK403874 | Microbacterium esteraromaticumstrain [GU111572]—99.3% Microbacterium sp. [MH231521]—98.5% Microbacterium sp. [MH231520]—98.5% | Microbacterium sp. |
| KBP.AS.954–PYG 10 °C–MK403875 | Uncultured Microbacterium sp. Clone [MH894291]—99.9% Microbacterium sp. [MH813404]—99.9% Microbacterium sp. [MH714660]—99.9% | Microbacterium sp. |
| KBP.AS.955–PYG 10 °C–MK403876 | Pseudarthrobacter siccitolerans [MF682005]—99.5% Pseudarthrobacter oxydans [MF681862]—99.5% Pseudarthrobacter oxydans [MF681851]—99.5% | Pseudarthrobacter sp. |
| KBP.AS.956–PYG 10°C–MK403877 | Planomicrobium glaciei [KF387709]—99.9% Planomicrobium sp. [JX949538]—99.9% Planomicrobium sp. [JQ618344]—99.9% | Planomicrobium sp. |
| KBP.AS.957–CM 10 °C–MK403878 | Arthrobacter sp. [KX645729]—99.5% Arthrobacter sp. [KF889389]—99.5% Arthrobacter crystallopoietes [KM257018]—99.5% | Arthrobacter sp. |
| KBP.AS.958–CM 10 °C–MK403879 | Arthrobacter sp. [KY173031]—99.6% Arthrobacter sp. [LN614607]—99.6% Arthrobacter oryzae [KC934820]—99.6% | Arthrobacter sp. |
| KBP.AS.960–CM 25 °C–MK403880 | Microbacterium esteraromaticum [GU111572]—99.3% Microbacterium sp. [MH231521]—98.6% Microbacterium sp. [MH231520]—98.6% | Microbacterium sp. |
| KBP.AS.961–CM 10 °C–MK403881 | Cellulomonas hominis [KT992126]—99.1% Cellulomonas sp. [Y09658]—99.1% Cellulomonas sp. [MF361869]—98.9% | Cellulomonas sp. |
| KBP.AS.962–CM 10 °C–None | Was identified by morphology | Streptomyces sp. |
| KBP.AS.963–CM 10 °C–MK403882 | Arthrobacter agilis [CP024915]—99.7% Arthrobacter agilis [MG279727]—99.7% Arthrobacter sp. [KY476520]—99.7% | Arthrobacter agilis |
| KBP.AS.972–PYG 10 °C–MK403883 | Micrococcus sp. [MH202946]—99.5% Micrococcus sp. [AB188213]—99.5% Micrococcus sp. [EF540464] —99.5% | Micrococcus sp. |
| KBP.AS.973–PYG 10 °C–MK403884 | Rhodococcus sp. [MH285872]—100% Rhodococcus sp. [CP034152]—100% Rhodococcus erythropolis [MH251258]—100% | Rhodococcus erythropolis |
| KBP.AS.1003–PYG 10 °C–MK403768 | Pseudarthrobacter phenanthrenivorans [NR_074770]—100% Arthrobacter sp. [LN871743]—100% Arthrobacter sp. [KU951449]—100% | Arthrobacter sp. |
| KBP.AS.1005–PYG 10 °C–MK403769 | Leucobacter sp. [KY623368]—99.9% Leucobacter aridicollis [KR827428]—99.9% Leucobacter sp. [KP152582]—99.9% | Leucobacter aridicollis |
| KBP.AS.1007–PYG 10 °C–MK403770 | Arthrobacter sp. [KT944733]—100% Arthrobacter sp. [KM507593]—100% Arthrobacter pityocampae [MH472818]—100% | Arthrobacter sp. |
| KBP.AS.1008–PYG 10 °C–MK403771 | Uncultured bacterium clone [KU514918]—99.3% Planomicrobium sp. [JX949538]—99.0% Planomicrobium glaciei [JF411320]—99.0% | Planomicrobium glaciei |
| KBP.AS.1009–PYG 10 °C–MK403772 | Pseudarthrobacter siccitolerans [MF682005]—99.8% Pseudarthrobacter oxydans [MF681862]—99.8% Pseudarthrobacter oxydans [MF681851]—99.8% | Pseudarthrobacter sp. |
| KBP.AS.1010–PYG 10 °C–MK403773 | Planomicrobium glaciei [KF387709]—99.7% Planomicrobium sp. [JX949538]—99.7% Planomicrobium glaciei [HQ232419]—99.7% | Planomicrobium glaciei |
| KBP.AS.1011–PYG 10 °C–MK403774 | Arthrobacter phenanthrenivorans [JX840975]—99.3% Uncultured soil bacterium clone [DQ248289]—99.3% Arthrobacter sp. [AY238502]—99.3% | Arthrobacter sp. |
| KBP.AS.1012–PYG 10 °C–MK403775 | Planococcus donghaensis [LN774491]—100% Planomicrobium okeanokoites [KF749394]—100% Planomicrobium okeanokoites [KF724965]—100% | Planomicrobium sp. |
| KBP.AS.1021–PYG 10 °C–MK403776 | Arthrobacter sp. [KT424968]—100% Arthrobacter agilis [KM036066]—100% Arthrobacter agilis [JN377650]—99.9% | Arthrobacter agilis |
| KBP.AS.1024–PYG 10 °C–MK403777 | Pseudarthrobacter phenanthrenivorans [NR_074770]—100% Arthrobacter sp. [LN871743]—100% Arthrobacter phenanthrenivorans [KR085846]—100% | Arthrobacter sp. |
| KBP.AS.1026–CM 25 °C–MK403778 | Uncultured bacterium [AB696407]—98.5% Massilia sp. [JQ511857]—98.0% Massilia sp. [JQ511858]—97.8% | Massilia sp. |
| KBP.AS.1028–CM 25 °C–MK403779 | Agrococcus lahaulensis [KF318382]—99.8% Agrococcus jenensis [EF672044]—99.8% Agrococcus lahaulensis [MF351823]—99.6% | Agrococcus sp. |
| KBP.AS.1029–CM 25 °C–MK403780 | Uncultured bacterium clone [DQ125573]—99.5% Pseudarthrobacter oxydans [MH304399]—99.3% Arthrobacter sp. [MH714681]—99.3% | Arthrobacter sp. |
| KBP.AS.1030–CM 25 °C–MK403781 | Pseudarthrobacter siccitolerans [MF682005]—99.9% Pseudarthrobacter oxydans [MF681862]—99.9% Pseudarthrobacter oxydans [MF681851]—99.9% | Pseudarthrobacter sp. |
| KBP.AS.1031–CM 25 °C–MK403782 | Massilia sp. [MH707216]—99.9% Massilia alkalitolerans [KY010279]—99.9% Massilia varians [KX784920]—99.9% | Massilia sp. |
| KBP.AS.1032–CM 25 °C–MK403783 | Arthrobacter agilis [KF924209]—99.8% Arthrobacter sp. [JX949695]—99.8% Arthrobacter sp. [JX949646]—99.8% | Arthrobacter sp. |
| KBP.AS.1033–CM 25 °C–MK403784 | Microbacterium sp. [LT601250]—99.5% Uncultured bacterium clone [MG805031]—99.5% Microbacterium sp. [MF526604]—99.5% | Microbacterium sp. |
| KBP.AS.1037–CM 25 °C–MK403785 | Massilia varians [KC429599]—99.7% Massilia alkalitolerans [KY010279]—99.6% Massilia alkalitolerans [KP282807]—99.6% | Massilia sp. |
| KBP.AS.1044–CM 25 °C–MK403786 | Uncultured bacterium clone [KC442649]—87.5% Pontibacter sp. [HM579810]—87.5% Pontibacter diazotrophicus [NR_126288]—87.2% | Pontibacter sp. |
Appendix B
| GenBank Accession Number | Strain | Taxonomic Affiliation | Primers Used for Amplification | Primers Used for Sequencing |
|---|---|---|---|---|
| MK403789 | KBP.AS.17 | Pseudarthrobacter oxydans | 27 f + Un1492 r | 1100 r |
| MK403790 | KBP.AS.62 | Arthrobacter sp. | 341 f + 805 r | 805 r |
| MK403794 | KBP.AS.75 | Brachybacterium sp. | 27 f + Un1492 r | 1100 r |
| MK403787 | KBP.AS.105 | Janthinobacterium sp. | 27 f + Un1492 r | 1100 r |
| MK403788 | KBP.AS.160 | Arthrobacter sp. | 27 f + Un1492 r | 1100 r |
| MK403791 | KBP.AS.747 | Microbacterium aurantiacum | 27 f + 537 r | 537 r |
| MK403792 | KBP.AS.748 | Pseudarthrobacter sp. | 27 f + Un1492 r | 1100 r |
| MK403793 | KBP.AS.749 | Brevibacterium frigoritolerans | 27 f + Un1492 r | 1100 r |
| MK403795 | KBP.AS.750 | Planomicrobium okeanokoites | 341 f + 805 r | 805 r |
| MK403796 | KBP.AS.751 | Agrococcus sp. | 27 f + Un1492 r | 1100 r |
| MK403797 | KBP.AS.752 | Agrococcus sp. | 27 f + Un1492 r | 1100 r |
| MK403798 | KBP.AS.753 | Agrococcus sp. | 27 f + 537 r | 537 r |
| MK403799 | KBP.AS.754 | Kocuria sp. | 27 f + Un1492 r | 1100 r |
| MK403800 | KBP.AS.755 | Planomicrobium glaciei | 27 f + Un1492 r | 1100 r |
| MK403801 | KBP.AS.756 | Leucobacter aridicollis | 27 f + Un1492 r | 1100 r |
| MK403802 | KBP.AS.757 | Planomicrobium okeanokoites | 341 f + 805 r | 805 r |
| MK403803 | KBP.AS.758 | Microbacterium sp. | 27 f + Un1492 r | 1100 r |
| MK403804 | KBP.AS.759 | Bacillus pumilus | 27 f + Un1492 r | 1100 r |
| MK403805 | KBP.AS.760 | Microbacterium sp. | 27 f + Un1492 r | 1100 r |
| MK403806 | KBP.AS.761 | Rufibacter sp. | 27 f + 537 r | 537 r |
| MK403807 | KBP.AS.762 | Microbacterium sp. | 27 f + Un1492 r | 1100 r |
| MK403808 | KBP.AS.763 | Massilia sp. | 27 f + Un1492 r | 1100 r |
| MK403809 | KBP.AS.764 | Bacillus sp. | 341 f + 805 r | 805 r |
| MK403810 | KBP.AS.765 | Massilia alkalitolerans | 27 f + Un1492 r | 1100 r |
| MK403811 | KBP.AS.766 | Pseudarthrobacter sp. | 27 f + Un1492 r | 1100 r |
| MK403812 | KBP.AS.767 | Massilia varians | 27 f + Un1492r | 1100r |
| MK403813 | KBP.AS.768 | Planomicrobium sp. | 27 f + Un1492 r | 1100r |
| MK403814 | KBP.AS.769 | Microbacterium pseudoresistens | 27 f + 537 r | 537 r |
| MK403815 | KBP.AS.770 | Arthrobacter sp. | 27 f + Un1492 r | 1100 r |
| MK403816 | KBP.AS.771 | Arthrobacter sp. | 27 f+Un1492 r | 1100 r |
| MK403817 | KBP.AS.772 | Planomicrobium glaciei | 27 f + Un1492 r | 1100 r |
| MK403818 | KBP.AS.773 | Cellulomonas sp. | 27 f + Un1492 r | 1100 r |
| MK403819 | KBP.AS.774 | Planomicrobium okeanokoites | 27 f + Un1492 r | 1100 r |
| MK403820 | KBP.AS.775 | Planomicrobium okeanokoites | 27 f + Un1492 r | 1100 r |
| MK403821 | KBP.AS.776 | Georgenia sp. | 27 f + Un1492 r | 1100 r |
| MK403822 | KBP.AS.777 | Microbacterium barkeri | 27 f + Un1492 r | 1100 r |
| MK403823 | KBP.AS.778 | Salinibacterium sp. | 27 f + Un1492 r | 1100 r |
| MK403824 | KBP.AS.779 | Brevibacterium frigoritolerans | 27 f + Un1492 r | 1100 r |
| MK403825 | KBP.AS.780 | Micrococcus sp. | 27 f + Un1492 r | 1100 r |
| MK403826 | KBP.AS.781 | Kocuria sp. | 27 f + Un1492 r | 1100 r |
| MK403827 | KBP.AS.782 | Labedella sp. | 27 f + Un1492 r | 1100 r |
| MK403828 | KBP.AS.783 | Plantibacter sp. | 27 f+ 537 r | 537 r |
| MK403829 | KBP.AS.784 | Arthrobacter agilis | 27 f + Un1492 r | 1100 r |
| MK403830 | KBP.AS.785 | Microbacterium sp. | 27 f + Un1492 r | 1100 r |
| MK403831 | KBP.AS.786 | Pseudarthrobacter sp. | 27 f + Un1492 r | 1100 r |
| MK403832 | KBP.AS.787 | Massilia sp. | 27 f + Un1492 r | 1100 r |
| None | KBP.AS.788 | Bacillus sp. | Was identified by morphology | |
| MK403833 | KBP.AS.789 | Burkholderia sp. | 341 f + 805 r | 805 r |
| MK403834 | KBP.AS.790 | Cellulomonas hominis | 27 f + Un1492 r | 1100 r |
| MK403835 | KBP.AS.791 | Pseudarthrobacter sp. | 27 f + Un1492 r | 1100 r |
| MK403836 | KBP.AS.792 | Massilia sp. | 27 f + Un1492 r | 1100 r |
| MK403837 | KBP.AS.793 | Salinibacterium sp. | 27 f + Un1492 r | 1100 r |
| MK403838 | KBP.AS.794 | Arthrobacter sp. | 27 f + Un1492 r | 1100 r |
| MK403839 | KBP.AS.795 | Arthrobacter sp. | 27 f + Un1492 r | 1100 r |
| MK403840 | KBP.AS.816 | Arthrobacter sp. | 27 f + Un1492 r | 1100 r |
| MK403841 | KBP.AS.842 | Massilia sp. | 27 f + Un1492 r | 1100 r |
| MK403842 | KBP.AS.843 | Mycetocola sp. | 27 f + Un1492 r | 1100 r |
| MK403843 | KBP.AS.844 | Arthrobacter agilis | 27 f + Un1492 r | 1100 r |
| None | KBP.AS.845 | Streptomyces sp. | Was identified by morphology | |
| MK403844 | KBP.AS.846 | Arthrobacter sp. | 27 f+ 537 r | 537 r |
| MK403845 | KBP.AS.847 | Cellulomonas sp. | 27 f + Un1492 r | 1100 r |
| MK403846 | KBP.AS.848 | Massilia sp. | 27 f + Un1492 r | 1100 r |
| MK403847 | KBP.AS.849 | Microbacterium sp. | 27 f + Un1492 r | 1100 r |
| MK403848 | KBP.AS.850 | Massilia sp. | 27 f + Un1492 r | 1100 r |
| MK403849 | KBP.AS.854 | Paracoccus marcusii | 27 f + Un1492 r | 1100r |
| None | KBP.AS.855 | Rufibacter sp. | Was identified by morphology | |
| MK403850 | KBP.AS.858 | Arthrobacter crystallopoietes | 27 f + Un1492 r | 1100 r |
| MK403851 | KBP.AS.862 | Arthrobacter sp. | 27 f + Un1492 r | 1100 r |
| MK403852 | KBP.AS.866 | Sphingomonas sp. | 27 f + Un1492 r | 1100 r |
| MK403853 | KBP.AS.867 | Microbacterium paraoxydans | 27 f + Un1492 r | 1100 r |
| MK403854 | KBP.AS.868 | Pseudarthrobacter sp. | 27 f + Un1492 r | 1100 r |
| MK403855 | KBP.AS.869 | Kocuria sp. | 341 f + 805 r | 805 r |
| MK403856 | KBP.AS.876 | Arthrobacter agilis | 27 f + Un1492 r | 1100 r |
| MK403857 | KBP.AS.877 | Salinibacterium sp. | 27 f + Un1492 r | 1100 r |
| MK403858 | KBP.AS.899 | Streptomyces sp. | 27 f + Un1492 r | 1100 r |
| MK403859 | KBP.AS.916 | Planomicrobium glaciei | 27 f + Un1492 r | 1100 r |
| MK403860 | KBP.AS.926 | Sphingomonas sp. | 27 f + Un1492 r | 1100 r |
| MK403861 | KBP.AS.927 | Arthrobacter agilis | 27 f + Un1492 r | 1100 r |
| MK403862 | KBP.AS.928 | Massilia sp. | 27 f + Un1492 r | 1100 r |
| MK403863 | KBP.AS.929 | Pseudarthrobacter sp. | 27 f + Un1492 r | 1100 r |
| MK403864 | KBP.AS.930 | Pseudarthrobacter sp. | 27 f + Un1492 r | 1100 r |
| MK403865 | KBP.AS.931 | Arthrobacter sp. | 27 f + Un1492 r | 1100 r |
| MK403866 | KBP.AS.932 | Pseudarthrobacter sp. | 27 f + Un1492 r | 1100 r |
| MK403867 | KBP.AS.936 | Pseudarthrobacter phenanthrenivorans | 27 f + Un1492 r | 1100 r |
| MK403868 | KBP.AS.937 | Arthrobacter agilis | 27 f + Un1492 r | 1100 r |
| MK403869 | KBP.AS.938 | Streptomyces sp. | 27 f + Un1492 r | 1100 r |
| MK403870 | KBP.AS.939 | Massilia sp. | 27 f + Un1492 r | 1100 r |
| MK403871 | KBP.AS.940 | Microbacterium sp. | 27 f + Un1492 r | 1100 r |
| MK403872 | KBP.AS.941 | Pseudarthrobacter sp. | 27 f + Un1492 r | 1100 r |
| MK403873 | KBP.AS.943 | Arthrobacter agilis | 27 f + Un1492 r | 1100 r |
| MK403874 | KBP.AS.947 | Microbacterium sp. | 27 f + Un1492 r | 1100 r |
| MK403875 | KBP.AS.954 | Microbacterium sp. | 27 f + Un1492 r | 1100 r |
| MK403876 | KBP.AS.955 | Pseudarthrobacter sp. | 27 f + Un1492 r | 1100 r |
| MK403877 | KBP.AS.956 | Planomicrobium sp | 27 f + Un1492 r | 1100 r |
| MK403878 | KBP.AS.957 | Arthrobacter sp. | 27 f + Un1492 r | 1100 r |
| MK403879 | KBP.AS.958 | Arthrobacter sp. | 27 f + Un1492 r | 1100 r |
| MK403880 | KBP.AS.960 | Microbacterium sp. | 27 f + Un1492 r | 1100 r |
| MK403881 | KBP.AS.961 | Cellulomonas sp. | 27 f + Un1492 r | 1100 r |
| None | KBP.AS.962 | Streptomyces sp. | Was identified by morphology | |
| MK403882 | KBP.AS.963 | Arthrobacter agilis | 27 f + Un1492 r | 1100 r |
| MK403883 | KBP.AS.972 | Micrococcus sp. | 27 f + Un1492 r | 1100 r |
| MK403884 | KBP.AS.973 | Rhodococcus erythropolis | 27 f + Un1492 r | 1100 r |
| MK403768 | KBP.AS.1003 | Arthrobacter sp. | 27 f + Un1492 r | 1100 r |
| MK403769 | KBP.AS.1005 | Leucobacter aridicollis | 27 f + Un1492 r | 1100 r |
| MK403770 | KBP.AS.1007 | Arthrobacter sp. | 27 f + Un1492 r | 1100 r |
| MK403771 | KBP.AS.1008 | Planomicrobium glaciei | 27 f + Un1492 r | 1100 r |
| MK403772 | KBP.AS.1009 | Pseudarthrobacter sp. | 27 f + Un1492 r | 1100 r |
| MK403773 | KBP.AS.1010 | Planomicrobium glaciei | 27 f + Un1492 r | 1100 r |
| MK403774 | KBP.AS.1011 | Arthrobacter sp. | 27 f + Un1492 r | 1100 r |
| MK403775 | KBP.AS.1012 | Planomicrobium sp | 341 f + 805 r | 805 r |
| MK403776 | KBP.AS.1021 | Arthrobacter agilis | 27 f + Un1492 r | 1100 r |
| MK403777 | KBP.AS.1024 | Arthrobacter sp. | 27 f + Un1492 r | 1100 r |
| MK403778 | KBP.AS.1026 | Massilia sp. | 27 f + Un1492 r | 1100 r |
| MK403779 | KBP.AS.1028 | Agrococcus sp. | 27 f + Un1492 r | 1100 r |
| MK403780 | KBP.AS.1029 | Arthrobacter sp. | 27 f + Un1492 r | 1100 r |
| MK403781 | KBP.AS.1030 | Pseudarthrobacter sp. | 27 f + Un1492 r | 1100 r |
| MK403782 | KBP.AS.1031 | Massilia sp. | 27 f + Un1492 r | 1100 r |
| MK403783 | KBP.AS.1032 | Arthrobacter sp. | 27 f + Un1492 r | 1100 r |
| MK403784 | KBP.AS.1033 | Microbacterium sp. | 341 f + 805 r | 805 r |
| MK403785 | KBP.AS.1037 | Massilia sp. | 27 f + Un1492 r | 1100 r |
| MK403786 | KBP.AS.1044 | Pontibacter sp. | 341 f + 805 r | 805 r |
Appendix C
| PYG 25 °C | PYG 10 °C | CM 25 °C | CM 10 °C |
|---|---|---|---|
| Agrococcus | Agrococcus | ||
| Arthrobacter | Arthrobacter | Arthrobacter | Arthrobacter |
| Bacillus | Bacillus | ||
| Brachybacterium | |||
| Burkholderia | |||
| Cellulomonas | Cellulomonas | Cellulomonas | |
| Georgenia | |||
| Janthinobacterium | |||
| Kocuria | |||
| Labedella | |||
| Leucobacter | Leucobacter | ||
| Massilia | Massilia | Massilia | |
| Microbacterium | Microbacterium | Microbacterium | |
| Micrococcus | |||
| Mycetocola | |||
| Paracoccus | Paracoccus | ||
| Planomicrobium | Planomicrobium | ||
| Plantibacter | |||
| Pontibacter | |||
| Pseudarthrobacter | Pseudarthrobacter | Pseudarthrobacter | |
| Rhodococcus | |||
| Rufibacter | |||
| Salinibacterium | Salinibacterium | ||
| Sphingomonas | Sphingomonas | ||
| Streptomyces | Streptomyces | Streptomyces |
References
- Chyba, C.F.; Hand, K.P. Astrobiology: The study of the living universe. Annu. Rev. Astron. Astrophys. 2005, 43, 31–74. [Google Scholar] [CrossRef]
- Cockell, C.S.; Schwendner, P.; Perras, A.; Rettberg, P.; Beblo-Vranesevic, K.; Bohmeier, M.; Rabbow, E.; Moissl-Eichinger, C.; Wink, L.; Marteinsson, V.; et al. Anaerobic microorganisms in astrobiological analogue environments: From field site to culture collection. Int. J. Astrobiol. 2017, 1–15. [Google Scholar] [CrossRef]
- Cloutis, E.A.; Craig, M.A.; Kruzelecky, R.V.; Jamroz, W.R.; Scott, A.; Hawthorne, F.C.; Mertzman, S.A. Spectral reflectance properties of minerals exposed to simulated Mars surface conditions. Icarus 2008, 195, 140–168. [Google Scholar] [CrossRef]
- Horneck, G.; Klaus, D.M.; Mancinelli, R.L. Space microbiology. Microbiol. Mol. Biol. Rev. 2010, 74, 121–156. [Google Scholar] [CrossRef]
- Rummel, J.D.; Beaty, D.W.; Jones, M.A.; Bakermans, C.; Barlow, N.G.; Boston, P.J.; Chevrier, V.F.; Clark, B.C.; de Vera, J.-P.P.; Gough, R.V.; et al. A new analysis of Mars “special regions”: Findings of the second MEPAG Special Regions Science Analysis Group (SR-SAG2). Astrobiology 2014, 14, 887–968. [Google Scholar] [CrossRef]
- Carr, M.H. The Surface of Mars; Cambridge University Press: Cambridge, UK, 2007; Volume 6. [Google Scholar]
- Gilichinsky, D.; Rivkina, E.; Shcherbakova, V.; Laurinavichuis, K.; Tiedje, J. Supercooled water brines within permafrost—an unknown ecological niche for microorganisms: A model for astrobiology. Astrobiology 2003, 3, 331–341. [Google Scholar] [CrossRef]
- Smith, H.D.; Baqué, M.; Duncan, A.G.; Lloyd, C.R.; McKay, C.P.; Billi, D. Comparative analysis of cyanobacteria inhabiting rocks with different light transmittance in the Mojave Desert: A Mars terrestrial analogue. Int. J. Astrobiol. 2014, 13, 271–277. [Google Scholar] [CrossRef]
- Dartnell, L.R.; Desorgher, L.; Ward, J.M.; Coates, A.J. Modelling the surface and subsurface martian radiation environment: Implications for astrobiology. Geophys. Res. Lett. 2007, 34, L02207. [Google Scholar] [CrossRef]
- Bay, S.; Ferrari, B.; Greening, C. Life without water: How do bacteria generate biomass in desert ecosystems? Microbiol. Aust. 2018, 39, 28–32. [Google Scholar] [CrossRef]
- Paenibacilius, B. Microbial diversity of extreme regions: An unseen heritage and wealth. Indian. J. Plant Genet. Resour. 2016, 29, 246–248. [Google Scholar] [CrossRef]
- Wierzchos, J.; Ascaso, C.; McKay, C.P. Endolithic cyanobacteria in halite rocks from the hyperarid core of the Atacama Desert. Astrobiology 2006, 6, 415–422. [Google Scholar] [CrossRef]
- McKay, C.P.; Friedmann, E.I.; Gómez-Silva, B.; Cáceres-Villanueva, L.; Andersen, D.T.; Landheim, R. Temperature and moisture conditions for life in the extreme arid region of the Atacama Desert: Four years of observations including the El Nino of 1997–1998. Astrobiology 2003, 3, 393–406. [Google Scholar] [CrossRef]
- Vítek, P.; Jehlička, J.; Edwards, H.G.; Hutchinson, I.; Ascaso, C.; Wierzchos, J. The miniaturized Raman system and detection of traces of life in halite from the Atacama Desert: Some considerations for the search for life signatures on Mars. Astrobiology 2012, 12, 1095–1099. [Google Scholar] [CrossRef]
- Benardini, J.N.; Sawyer, J.; Venkateswaran, K.; Nicholson, W.L. Spore UV and acceleration resistance of endolithic Bacillus pumilus and Bacillus subtilis isolates obtained from Sonoran desert basalt: Implications for lithopanspermia. Astrobiology 2003, 3, 709–717. [Google Scholar] [CrossRef]
- Doran, P.T.; Lyons, W.B.; McKnight, D.M. (Eds.) Life in Antarctic Deserts and Other Cold Dry Environments: Astrobiological Analogs; Cambridge University Press: Cambridge, UK, 2010; Volume 5, ISBN 978-0-521-88919-3. [Google Scholar]
- Rampelotto, P.H. Resistance of microorganisms to extreme environmental conditions and its contribution to astrobiology. Sustainability 2010, 2, 1602–1623. [Google Scholar] [CrossRef]
- Vítek, P.; Jehlička, J.; Edwards, H.G.; Hutchinson, I.; Ascaso, C.; Wierzchos, J. Miniaturized Raman instrumentation detects carotenoids in Mars-analogue rocks from the Mojave and Atacama deserts. Phil. Trans. R. Soc. A 2014, 372, 20140196. [Google Scholar] [CrossRef]
- Salas, E.; Abbey, W.; Bhartia, R.; Beegle, L.W. The Mojave Desert: A Martian Analog Site for Future Astrobiology Themed Missions; Jet Propulsion Laboratory, NASA: Pasadena, CA, USA, 2011. Available online: http://hdl.handle.net/2014/43479 (accessed on 9 April 2019).
- Bishop, J.L.; Schelble, R.T.; McKay, C.P.; Brown, A.J.; Perry, K.A. Carbonate rocks in the Mojave Desert as an analogue for Martian carbonates. Int. J. Astrobiol. 2011, 10, 349–358. [Google Scholar] [CrossRef]
- Purohit, H.J.; Kaila, V.C.; Vaidya, V.C.; Khardenavis, A.A. Optimization and Applicability of Bioprocesses; Springer Nature: Singapore, 2017; ISBN 978-981-10-6863-8. [Google Scholar]
- Makhalanyane, T.P.; Valverde, A.; Gunnigle, E.; Frossard, A.; Ramond, J.B.; Cowan, D.A. Microbial ecology of hot desert edaphic systems. FEMS Microbiol. Rev. 2015, 39, 203–221. [Google Scholar] [CrossRef] [Green Version]
- Abbey, W.; Salas, E.; Bhartia, R.; Beegle, L.W. The Mojave vadose zone: A subsurface biosphere analogue for Mars. Astrobiology 2013, 13, 637–646. [Google Scholar] [CrossRef]
- McHugh, T.A.; Compson, Z.; van Gestel, N.; Hayer, M.; Ballard, L.; Haverty, M.; Hines, J.; Irvine, N.; Krassner, D.; Lyons, T.; et al. Climate controls prokaryotic community composition in desert soils of the southwestern United States. FEMS Microbiol. Ecol. 2017, 93. [Google Scholar] [CrossRef]
- Preston, L.J.; Dartnell, L.R. Planetary habitability: Lessons learned from terrestrial analogues. Int. J. Astrobiol. 2014, 13, 81–98. [Google Scholar] [CrossRef]
- Navarro-González, R.; Navarro, K.F.; de la Rosa, J.; Iñiguez, E.; Molina, P.; Miranda, L.D.; Morales, P.; Cienfuegos, E.; Coll, P.; Raulin, F.; et al. The limitations on organic detection in Mars-like soils by thermal volatilization–gas chromatography–MS and their implications for the Viking results. Proc. Natl. Acad. Sci. USA 2006, 103, 16089–16094. [Google Scholar] [CrossRef] [Green Version]
- Greeley, R.; Bridges, N.T.; Kuzmin, R.O.; Laity, J.E. Terrestrial analogs to wind-related features at the Viking and Pathfinder landing sites on Mars. J. Geophys. Res. Planets 2002, 107, 5-1–5-22. [Google Scholar] [CrossRef]
- Davila, A.F.; Schulze-Makuch, D. The last possible outposts for life on Mars. Astrobiology 2016, 16, 159–168. [Google Scholar] [CrossRef]
- Peters, G.H.; Abbey, W.; Bearman, G.H.; Mungas, G.S.; Smith, J.A.; Anderson, R.C.; Douglas, S.; Beegle, L.W. Mojave Mars simulant—Characterization of a new geologic Mars analog. Icarus 2008, 197, 470–479. [Google Scholar] [CrossRef]
- Schlesinger, W.H.; Pippen, J.S.; Wallenstein, M.D.; Hofmockel, K.S.; Klepeis, D.M.; Mahall, B.E. Community composition and photosynthesis by photoautotrophs under quartz pebbles, southern Mojave Desert. Ecology 2003, 84, 3222–3231. [Google Scholar] [CrossRef]
- Navarro-González, R.; Rainey, F.A.; Molina, P.; Bagaley, D.R.; Hollen, B.J.; de la Rosa, J.; Small, A.M.; Quinn, R.C.; Grunthaner, F.J.; Cáceres, L.; et al. Mars-like soils in the Atacama Desert, Chile, and the dry limit of microbial life. Science 2003, 302, 1018–1021. [Google Scholar] [CrossRef]
- Mogul, R.; Vaishampayan, P.; Bashir, M.; McKay, C.P.; Schubert, K.; Bornaccorsi, R.; Gomez, E.; Payton, G.; Capra, J.; Bacon, L.; et al. Microbial community and biochemical dynamics of biological soil crusts across a gradient of surface coverage in the central Mojave Desert. Front. Microbiol. 2017, 8, 1974. [Google Scholar] [CrossRef]
- Fierer, N.; Leff, J.W.; Adams, B.J.; Nielsen, U.N.; Bates, S.T.; Lauber, C.L.; Owens, S.; Gilbert, J.A.; Wall, D.H.; Caporaso, J.G. Cross-biome metagenomic analyses of soil microbial communities and their functional attributes. Proc. Natl. Acad. Sci. USA 2012, 109, 21390–21395. [Google Scholar] [CrossRef] [Green Version]
- Schelble, R.T.; McDonald, G.D.; Hall, J.A.; Nealson, K.H. Community structure comparison using FAME analysis of desert varnish and soil, Mojave Desert, California. Geomicrobiol. J. 2005, 22, 353–360. [Google Scholar] [CrossRef]
- Ewing, S.A.; Southard, R.J.; Macalady, J.L.; Hartshorn, A.S.; Johnson, M.J. Soil microbial fingerprints, carbon, and nitrogen in a Mojave Desert creosote-bush ecosystem. Soil Sci. Soc. Am. J. 2007, 71, 469–475. [Google Scholar] [CrossRef]
- Smith, H.D.; Duncan, A.G.; Neary, P.L.; Lloyd, C.R.; Anderson, A.J.; Sims, R.C.; McKay, C.P. In situ microbial detection in Mojave Desert soil using native fluorescence. Astrobiology 2012, 12, 247–257. [Google Scholar] [CrossRef]
- Bryant, E.; Rech, S. The effect of moisture on soil microbial communities in the Mojave Desert. Astrobiology 2008, 8, 427. [Google Scholar] [CrossRef]
- Garrity, G.M.; Heimbuch, B.K.; Gagliardi, M. Isolation of zoosporogenous actinomycetes from desert soils. J. Ind. Microbiol. 1996, 17, 260–267. [Google Scholar] [CrossRef]
- Garcia, D.E.; Lopez, B.R.; de-Bashan, L.E.; Hirsch, A.M.; Maymon, M.; Bashan, Y. Functional metabolic diversity of the bacterial community in undisturbed resource island soils in the southern Sonoran Desert. Land. Degrad. Dev. 2018, 29, 1467–1477. [Google Scholar] [CrossRef]
- Pointing, S.B.; Belnap, J. Microbial colonization and controls in dryland systems. Nat. Rev. Microbiol. 2012, 10, 551. [Google Scholar] [CrossRef]
- Loughney, K.M.; Badgley, C. Facies, Environments, and Fossil Preservation in the Barstow Formation, Mojave Desert, California. Palaios 2017, 32, 396–412. [Google Scholar] [CrossRef]
- Reynolds, R.L.; Reheis, M.; Yount, J.; Lamothe, P. Composition of aeolian dust in natural traps on isolated surfaces of the central Mojave Desert—Insights to mixing, sources, and nutrient inputs. J. Arid Environ. 2006, 66, 42–61. [Google Scholar] [CrossRef]
- Cheptsov, V.S.; Vorobyova, E.A.; Osipov, G.A.; Manucharova, N.A.; Polyanskaya, L.M.; Gorlenko, M.V.; Pavlov, A.K.; Rosanova, M.S.; Lomasov, V.N. Microbial activity in Martian analog soils after ionizing radiation: Implications for the preservation of subsurface life on Mars. AIMS Microbiol. 2018, 4, 541–562. [Google Scholar] [CrossRef]
- Belov, A.A.; Cheptsov, V.S.; Vorobyova, E.A. Soil bacterial communities of Sahara and Gibson deserts: Physiological and taxonomical characteristics. AIMS Microbiol. 2018, 4, 685–710. [Google Scholar] [CrossRef]
- Schinner, F.; Öhlinger, R.; Kandeler, E.; Margesin, R. (Eds.) Methods in Soil Biology; Springer Science & Business Media: Berlin, Germany, 2012; ISBN 978-3-642-60966-4. [Google Scholar]
- Depository of Live Systems. Available online: https://depo.msu.ru (accessed on 8 April 2019).
- Marchesi, J.R.; Sato, T.; Weightman, A.J.; Martin, T.A.; Fry, J.C.; Hiom, S.J.; Wade, W.G. Design and evaluation of useful bacterium-specific PCR primers that amplify genes coding for bacterial 16S rRNA. Appl. Environ. Microbiol. 1998, 64, 795–799. [Google Scholar]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef]
- DeLong, E.F. Archaea in coastal marine environments. Proc. Natl. Acad. Sci. USA 1992, 89, 5685–5689. [Google Scholar] [CrossRef]
- Lane, D.J. 16S/23S rRNA sequencing, In Nucleic Acid Techniques in Bacterial Systematic; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 1991; pp. 115–175. [Google Scholar]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Chromas and ChromasPro DNA Sequencing Software—Technelysium Pty Ltd. Available online: http://www.technelysium.com.au (accessed on 8 April 2019).
- Clustal Omega < Multiple Sequence Alignment < EMBL-EBI. Available online: http://www.ebi.ac.uk/Tools/msa/clustalo/ (accessed on 8 April 2019).
- BLAST: Basic Local Alignment Search Tool. Available online: http: //blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 8 April 2019).
- Mega Home. Available online: https://www.megasoftware.net/ (accessed on 8 April 2019).
- Gorlenko, M.V.; Majorova, T.N.; Kozhevin, P.A. Disturbances and their influence on substrate utilization patterns in soil microbial communities. In Microbial Communities; Springer: Berlin, Germany, 1997; pp. 84–93. [Google Scholar]
- Gorlenko, M.V.; Kozhevin, P.A. Multisubstrate Testing of Natural Microbial Communities; MAKS Press: Moscow, Russia, 2005. (In Russian) [Google Scholar]
- Chernov, I.Y.; Lysak, L.V. Methodical Materials for Seminars on the Course-General Ecology; MAKS Press: Moscow, Russia, 2005. (In Russian) [Google Scholar]
- Titus, J.H.; Nowak, R.S.; Smith, S.D. Soil resource heterogeneity in the Mojave Desert. J. Arid Environ. 2002, 52, 269–292. [Google Scholar] [CrossRef] [Green Version]
- Hereford, R.; Webb, R.H.; Longpre, C.I. Precipitation history and ecosystem response to multidecadal precipitation variability in the Mojave Desert region, 1893–2001. J. Arid Environ. 2006, 67, 13–34. [Google Scholar] [CrossRef]
- Manucharova, N.A.; Vlasenko, A.N.; Men’ko, E.V.; Zvyagintsev, D.G. Specificity of the chitinolytic microbial complex of soils incubated at different temperatures. Microbiology 2011, 80, 205–215. [Google Scholar] [CrossRef]
- Barer, M.R. Viable but non-culturable and dormant bacteria: Time to resolve an oxymoron and a misnomer? J. Med. Microbiol. 1997, 46, 629–631. [Google Scholar] [CrossRef]
- Christner, B.C.; Mosley-Thompson, E.; Thompson, L.G.; Zagorodnov, V.; Sandman, K.; Reeve, J.N. Recovery and identification of viable bacteria immured in glacial ice. Icarus 2000, 144, 479–485. [Google Scholar] [CrossRef]
- Rainey, F.A.; Ray, K.; Ferreira, M.; Gatz, B.Z.; Nobre, M.F.; Bagaley, D.; Small, A.M. Extensive diversity of ionizing-radiation-resistant bacteria recovered from Sonoran Desert soil and description of nine new species of the genus Deinococcus obtained from a single soil sample. Appl. Environ. Microbiol. 2005, 71, 5225–5235. [Google Scholar] [CrossRef]
- Luo, X.; Wang, J.; Zeng, X.C.; Wang, Y.; Zhou, L.; Nie, Y.; Dai, J.; Fang, C. Mycetocola manganoxydans sp. nov., an actinobacterium isolated from the Taklamakan desert. Int. J. Syst. Evol. Microbiol. 2012, 62, 2967–2970. [Google Scholar] [CrossRef] [Green Version]
- Mayilraj, S.; Suresh, K.; Schumann, P.; Kroppenstedt, R.M.; Saini, H.S. Agrococcus lahaulensis sp. nov., isolated from a cold desert of the Indian Himalayas. Int. J. Syst. Evol. Microbiol. 2006, 56, 1807–1810. [Google Scholar] [CrossRef] [Green Version]
- Gontia, I.; Kavita, K.; Schmid, M.; Hartmann, A.; Jha, B. Brachybacterium saurashtrense sp. nov., a halotolerant root-associated bacterium with plant growth-promoting potential. Int. J. Syst. Evol. Microbiol. 2011, 61, 27992804. [Google Scholar] [CrossRef]
- Lin, Y.C.; Yokota, A. Plantibacter auratus sp. nov., in the family Microbacteriaceae. Int. J. Syst. Evol. Microbiol. 2006, 56, 2337–2339. [Google Scholar] [CrossRef] [Green Version]
- Hatayama, K.; Esaki, K.; Ide, T. Cellulomonas soli sp. nov. and Cellulomonas oligotrophica sp. nov., isolated from soil. Int. J. Syst. Evol. Microbiol. 2013, 63, 60–65. [Google Scholar] [CrossRef]
- Ahmed, I.; Kudo, T.; Abbas, S.; Ehsan, M.; Iino, T.; Fujiwara, T.; Ohkuma, M. Cellulomonas pakistanensis sp. nov., a moderately halotolerant Actinobacteria. Int. J. Syst. Evol. Microbiol. 2014, 64, 2305–2311. [Google Scholar] [CrossRef] [Green Version]
- Cheptsov, V.; Vorobyova, E.; Belov, A.; Pavlov, A.; Tsurkov, D.; Lomasov, V.; Bulat, S. Survivability of soil and permafrost microbial communities after irradiation with accelerated electrons under simulated Martian and open space conditions. Geosciences 2018, 8, 298. [Google Scholar] [CrossRef]
- Mahato, N.K.; Tripathi, C.; Nayyar, N.; Singh, A.K.; Lal, R. Pontibacter ummariensis sp. nov., isolated from a hexachlorocyclohexane-contaminated soil. Int. J. Syst. Evol. Microbiol. 2016, 66, 1080–1087. [Google Scholar] [CrossRef] [Green Version]
- Rassner, S.M.E.; Anesio, A.M.; Girdwood, S.E.; Hell, K.; Gokul, J.K.; Whitworth, D.E.; Edwards, A. Can the bacterial community of a high Arctic glacier surface escape viral control? Front. Microbiol. 2016, 7, 956. [Google Scholar] [CrossRef]
- Zhang, D.C.; Liu, H.C.; Xin, Y.H.; Yu, Y.; Zhou, P.J.; Zhou, Y.G. Salinibacterium xinjiangense sp. nov., a psychrophilic bacterium isolated from the China No. 1 glacier. Int. J. Syst. Evol. Microbiol. 2008, 58, 2739–2742. [Google Scholar] [CrossRef] [Green Version]
- Han, S.K.; Nedashkovskaya, O.I.; Mikhailov, V.V.; Kim, S.B.; Bae, K.S. Salinibacterium amurskyense gen. nov., sp. nov., a novel genus of the family Microbacteriaceae from the marine environment. Int. J. Syst. Evol. Microbiol. 2003, 53, 2061–2066. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.C.; Liu, H.C.; Xin, Y.H.; Yu, Y.; Zhou, P.J.; Zhou, Y.G. Planomicrobium glaciei sp. nov., a psychrotolerant bacterium isolated from a glacier. Int. J. Syst. Evol. Microbiol. 2009, 59, 1387–1390. [Google Scholar] [CrossRef] [Green Version]
- Fagliarone, C.; Mosca, C.; Ubaldi, I.; Verseux, C.; Baqué, M.; Wilmotte, A.; Billi, D. Avoidance of protein oxidation correlates with the desiccation and radiation resistance of hot and cold desert strains of the cyanobacterium Chroococcidiopsis. Extremophiles 2017, 21, 981–991. [Google Scholar] [CrossRef]
- Shukla, M.; Chaturvedi, R.; Tamhane, D.; Vyas, P.; Archana, G.; Apte, S.; Bandekar, J.; Desai, A. Multiple-stress tolerance of ionizing radiation-resistant bacterial isolates obtained from various habitats: Correlation between stresses. Curr. Microbiol. 2007, 54, 142–148. [Google Scholar] [CrossRef]
- Clark, B.C.; van Hart, D.C. The salts of Mars. Icarus 1981, 45, 370–378. [Google Scholar] [CrossRef]
- Lushchak, V.I. Adaptive response to oxidative stress: Bacteria, fungi, plants and animals. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2011, 153, 175–190. [Google Scholar] [CrossRef]
- Cheptsov, V.S.; Vorobyova, E.A.; Manucharova, N.A.; Gorlenko, M.V.; Pavlov, A.K.; Vdovina, M.A.; Lomasov, V.N.; Bulat, S.A. 100 kGy gamma-affected microbial communities within the ancient Arctic permafrost under simulated Martian conditions. Extremophiles 2017, 21, 1057–1067. [Google Scholar] [CrossRef]
- Musilova, M.; Wright, G.; Ward, J.M.; Dartnell, L.R. Isolation of radiation-resistant bacteria from Mars analog Antarctic Dry Valleys by preselection, and the correlation between radiation and desiccation resistance. Astrobiology 2015, 15, 1076–1090. [Google Scholar] [CrossRef]
- Gholami, M.; Etemadifar, Z.; Bouzari, M. Isolation a new strain of Kocuria rosea capable of tolerating extreme conditions. J. Environ. Radioact. 2015, 144, 113–119. [Google Scholar] [CrossRef]
- Yu, L.Z.H.; Luo, X.S.; Liu, M.; Huang, Q. Diversity of ionizing radiation-resistant bacteria obtained from the Taklimakan Desert. J. Basic Microbiol. 2015, 55, 135–140. [Google Scholar] [CrossRef]
- Mao, J.; Tang, Q.; Zhang, Z.; Wang, W.; Wei, D.; Huang, Y.; Liu, Z.; Shi1, Y.; Goodfellow, M. Streptomyces radiopugnans sp. nov., a radiation-resistant actinomycete isolated from radiation-polluted soil in China. Int. J. Syst. Evol. Microbiol. 2007, 57, 2578–2582. [Google Scholar] [CrossRef] [Green Version]
- Marizcurrena, J.J.; Morel, M.A.; Braña, V.; Morales, D.; Martinez-López, W.; Castro-Sowinski, S. Searching for novel photolyases in UVC-resistant Antarctic bacteria. Extremophiles 2017, 21, 409–418. [Google Scholar] [CrossRef]
- Farías, M.E.; Fernández-Zenoff, V.; Flores, R.; Ordóñez, O.; Estévez, C. Impact of solar radiation on bacterioplankton in Laguna Vilama, a hypersaline Andean lake (4650 m). J. Geophys. Res. Biogeosci. 2009, 114. [Google Scholar] [CrossRef] [Green Version]
- Mojib, N.; Farhoomand, A.; Andersen, D.T.; Bej, A.K. UV and cold tolerance of a pigment-producing Antarctic Janthinobacterium sp. Ant5-2. Extremophiles 2013, 17, 367–378. [Google Scholar] [CrossRef]
- Vallalar, B. Investigation of the Growth and Survival of Bacteria from Mars Analog Environments When Exposed to Mars-like Conditions. Master of Science (MS), Bharathi Vallalar, Louisiana State University and Agricultural and Mechanical College. Available online: https://digitalcommons.lsu.edu/gradschool_theses/3601 (accessed on 9 April 2019).
- Horvath, R.S. Microbial co-metabolism and the degradation of organic compounds in nature. Bacteriol. Rev. 1972, 36, 146–155. [Google Scholar]
- Romero, D.; Traxler, M.F.; López, D.; Kolter, R. Antibiotics as signal molecules. Chem. Rev. 2011, 111, 5492–5505. [Google Scholar] [CrossRef]
- Miller, M.B.; Bassler, B.L. Quorum sensing in bacteria. Annu. Rev. Microbiol. 2001, 55, 165–199. [Google Scholar] [CrossRef]






| Index | PYG 25 °C | PYG 10 °C | CM 25 °C | CM 10 °C |
|---|---|---|---|---|
| Shannon index | 3.87 | 2.93 | 3.56 | 1.44 |
| Evenness index | 0.87 | 0.88 | 0.91 | 0.72 |
| Index | PYG 25 °C–PYG 10 °C | CM 25 °C–CM 10 °C | PYG 25 °C–CM 25 °C | PYG 10 °C–CM 10 °C |
|---|---|---|---|---|
| Common genera number | 8 | 4 | 12 | 3 |
| Jaccard index | 0.33 | 0.27 | 0.48 | 0.27 |
| Chekanovskiy–Sørensen index | 0.50 | 0.42 | 0.65 | 0.43 |
| Sørensen modified index | 0.55 | 0.42 | 0.57 | 0.34 |
| Most assimilated substrates | Amino acids | Alanine | Weakly assimilated substrates | Amino acids | Arginine |
| Glutamine | Asparagine | ||||
| Proline | Serine | ||||
| Lipid-like substance | Tween 80 | Threonine | |||
| Carbohydrates | Pullulan | Phenylalanine | |||
| Soluble starch | Amino acid polymer | Peptone | |||
| Sucrose | Salt of carbon acid | Propionate | |||
| Arabinose | Carbohydrates | Dextran 500 | |||
| Glucose | Xylose | ||||
| Maltose | |||||
| Alcohol | Sorbitol | ||||
| Salt of carbon acid | Lactate | ||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Belov, A.A.; Cheptsov, V.S.; Vorobyova, E.A.; Manucharova, N.A.; Ezhelev, Z.S. Stress-Tolerance and Taxonomy of Culturable Bacterial Communities Isolated from a Central Mojave Desert Soil Sample. Geosciences 2019, 9, 166. https://doi.org/10.3390/geosciences9040166
Belov AA, Cheptsov VS, Vorobyova EA, Manucharova NA, Ezhelev ZS. Stress-Tolerance and Taxonomy of Culturable Bacterial Communities Isolated from a Central Mojave Desert Soil Sample. Geosciences. 2019; 9(4):166. https://doi.org/10.3390/geosciences9040166
Chicago/Turabian StyleBelov, Andrey A., Vladimir S. Cheptsov, Elena A. Vorobyova, Natalia A. Manucharova, and Zakhar S. Ezhelev. 2019. "Stress-Tolerance and Taxonomy of Culturable Bacterial Communities Isolated from a Central Mojave Desert Soil Sample" Geosciences 9, no. 4: 166. https://doi.org/10.3390/geosciences9040166