
Polyamine and Ethanolamine Metabolism in Bacteria as an Important Component of Nitrogen Assimilation for Survival and Pathogenicity


1
Interfaculty Institute of Microbiology and Infection Medicine Tübingen (IMIT), Department of Microbiology and Biotechnology, University of Tübingen, Auf der Morgenstelle 28, 72076 Tübingen, Germany
2
Cluster of Excellence ‘Controlling Microbes to Fight Infections’, University of Tübingen, 72076 Tübingen, Germany
*
Author to whom correspondence should be addressed.
Med. Sci. 2022, 10(3), 40; https://doi.org/10.3390/medsci10030040
Submission received: 30 June 2022
/
Revised: 23 July 2022
/
Accepted: 25 July 2022
/
Published: 29 July 2022
(This article belongs to the Special Issue Polyamines: Essential Molecules Regulating Development, Health, and Disease)
Abstract
:Nitrogen is an essential element required for bacterial growth. It serves as a building block for the biosynthesis of macromolecules and provides precursors for secondary metabolites. Bacteria have developed the ability to use various nitrogen sources and possess two enzyme systems for nitrogen assimilation involving glutamine synthetase/glutamate synthase and glutamate dehydrogenase. Microorganisms living in habitats with changeable availability of nutrients have developed strategies to survive under nitrogen limitation. One adaptation is the ability to acquire nitrogen from alternative sources including the polyamines putrescine, cadaverine, spermidine and spermine, as well as the monoamine ethanolamine. Bacterial polyamine and monoamine metabolism is not only important under low nitrogen availability, but it is also required to survive under high concentrations of these compounds. Such conditions can occur in diverse habitats such as soil, plant tissues and human cells. Strategies of pathogenic and non-pathogenic bacteria to survive in the presence of poly- and monoamines offer the possibility to combat pathogens by using their capability to metabolize polyamines as an antibiotic drug target. This work aims to summarize the knowledge on poly- and monoamine metabolism in bacteria and its role in nitrogen metabolism.
1. Nitrogen Metabolism
1.1. Fundamentals of Nitrogen Assimilation in Prokaryotes
Nitrogen belongs to the group of vital elements. It is a key macronutrient required for the growth of living organisms. It is an essential component of amino acids, amino sugars, coenzymes, purines and pirimidines in nucleic acids [1,2], polyamines, and monoamines [3]. Microorganisms can use nitrogen sources with different redox states, including molecular nitrogen (redox state 0), ammonium (redox state −3), nitrate (redox state +5), and nitrite (redox state +3). Energetically, ammonium is the most preferred nitrogen source as a compound in the most reduced form.
Pathways for nitrogen assimilation comprise utilization pathways from the extracellular environment and biosynthetic pathways for intracellular production. Depending on the organism, these pathways are coordinated in order to control the intracellular amounts of nitrogen-containing compounds. Depending on the intracellular pool of nitrogen, expression of enzymes of nitrogen metabolism must be well coordinated. Nitrogen control has previously been precisely investigated in Gram-negative bacteria Escherichia coli, Salmonella typhimuium, Pseudomonas aeruginosa, as well as Gram-positive actinobacterial species Corynebacterium glutamicum, Streptomyces coelicolor, and Mycobacterium tuberculosis [4,5,6,7,8].
Some nitrogen-containing compounds such as ammonium can enter the cell by diffusion. Other compounds, e.g., amino acids, nitrate, nitrite, and urea, can be taken up by transporters [9]. A favored inorganic nitrogen source is ammonium which can diffuse across cell membranes or be transported into the cells via specialized Amt transporters. Subsequently, it can be directly used in the synthesis of glutamine and glutamate, which are key building blocks for biomolecules and subsequent biomass generation. Other inorganic nitrogen sources including nitrate, nitrite, urea, and diatomic nitrogen need to be reduced to ammonium before they can be assimilated. This process requires extra energy—in the form of ATP and electrons [5]. Organic nitrogen sources such as amino acids and amino sugars can be incorporated into the metabolism directly or in the deaminated and deamidated forms [8,10,11,12,13]. Alternative complex organic and non-organic nitrogen sources including monoamines and polyamines first have to be neutralized and detoxified with subsequent reduction [14,15].
Bacteria possess two enzyme systems for ammonium assimilation based on the glutamine synthetase/glutamate synthase (GS/GOGAT) and glutamate dehydrogenase (GDH) activity, which are ubiquitous in bacteria. Under different nitrogen concentrations, these two distinctive pathways form glutamine and glutamate, respectively. The intracellular ammonium is integrated into cellular metabolism by the glutamine synthetase. It is active under low concentrations of ammonium as a component of the glutamine synthetase/glutamate synthase (GS/GOGAT) pathway. Glutamine synthetase (GS) catalyzes the ATP-dependent synthesis of glutamate and ammonia to glutamine. Glutamate is generated from glutamine and 2-oxyglutarate by glutamine-2-oxoglutarate-aminotransferase (GOGAT) [5,6,15]. Glutamate synthase (GOGAT) transfers the amide group from glutamine to 2-oxoglutarate (2-OG), producing two molecules of glutamate. Under high concentrations of ammonium, the glutamate dehydrogenase (GDH) is active. It catalyzes the synthesis of glutamate using substrates ammonium and 2-oxyglutarate as well as NADPH. However, the GDH remains inactive under limiting conditions due to the low substrate affinity and high Km value [6] (Figure 1). Glutamine and glutamate can be subsequently incorporated into diverse biosynthetic reactions in the cell. Glutamine provides nitrogen for the synthesis of aromatic compounds (purines, pyrimidines), amino acids (arginine, histidine, tryptophane, asparagine), amino sugars (glucosamine), and others (e.g., aminobenzoate). Glutamate is the donor of nitrogen in transamination reactions [16,17] (Figure 1).
1.2. Nitrogen Assimilation in Gram-Negative Bacteria
For survival in a competitive and stressful environment under nutrient limitation, bacteria have developed a complex metabolism and regulatory machinery that controls the amount of nitrogen in the cell at transcriptional and post-transcriptional levels. Nitrogen control has been extensively investigated predominantly in enteric bacteria from the Enterobacteriaceae family: Escherichia coli, Salmonella typhimurium, Klebsiella pneumoniae, and Klebsiella aerogenes. A key role has the central response regulator NtrC, which is coupled with a specific sensor kinase NtrB sensing the limitation of nitrogen. In addition, the status of nitrogen and the regulation of enzyme activity at a global level are controlled via the following proteins: encoded by glnD uridylyltransferase/uridylyl-removing enzyme UTase; an adenylylate transferase/deadenylylase GlnE; small signal transducing proteins PII GlnK and GlnB [6].
1.2.1. Nitrogen Assimilation and Control in Escherichia coli
In E. coli, nitrogen starvation conditions induce the differential expression of genes that are regulated by the two-component system NtrB/NtrC. The PII signal protein GlnB is essential in this regulatory cascade sensing the cellular level of ATP, 2-oxoglutarate or glutamate [6]. The role of GlnK has also been demonstrated in E. coli. During nitrogen starvation, it regulates the expression of Ntr genes [18]. The PII protein can occur in two different forms in E. coli: PII and urydylylated PII-UMP. The bifunctional uridylyltransferase GlnD is responsible for two different states of the PII protein, whereas both activities are influenced by the amounts in the cell of the 2-oxoglutarate/glutamine and ATP/ADP. High carbon and ATP amounts signal the nitrogen limitation stimulating the activity of the uridylyltransferase. High concentrations of glutamine stimulate the uridylyl-removing activity of UTase. Under nitrogen-limiting conditions, urydylylated PII protein senses the phosphorylation of the sensor kinase NtrB [6]. Phosphorylated NtrB can activate the response regulator NtrC via the transfer of the phospho-group [19]. Phosphorylation of NtrC changes its conformation forming a dimer of two NtrC molecules. The dimeric form of NtrC enhances the transcription of nitrogen metabolism genes, including the gene glnA encoding glutamine synthetase through binding to the σ54 sigma factor-dependent promoters. Furthermore, in free-living anaerobic nitrogen-fixing bacteria such as Rhizobia sp. and Klebsiella sp., strict regulation of the nif genes for nitrogen fixation is present and depends on nitrogen availability. For example, in Klebsiella pneumoniae, the NtrB/NtrC system has been demonstrated to be required for control of the nifL gene for nitrogen fixation [20].
Nitrogen assimilation was also described to be controlled on the post-translational level. It occurs via covalent adenylylation and deadenylylation of each subunit of the glutamine synthetase enzyme [21,22]. This requires the bifunctional protein GlnE, which is regulated by the PII signal protein. In E. coli, modification of GSI by the covalent addition of AMP leads to a reduction of enzyme activity [23]. GlnE has two distinct domains, the N-terminal deadenylylation domain and the C-terminal adenylylation domain, that catalyze two different reactions. These domains are connected by an interregion linker. Besides the GS binding region, GlnE also has independent glutamine, PII, and PII-UMP binding sites. Under glutamine-rich conditions, glutamine and PII bind to GlnE. This causes the linker region to bring the adenylylation and deadenylylation domains together allowing adenylylation [23]. Under glutamine-limiting conditions, PII-UMP binds to GlnE. This causes a conformation change and allows deadenylylation [24].
1.2.2. Nitrogen Assimilation and Control in Cyanobacteria
In cyanobacteria, nitrogen is catabolized via the coupled reaction in the GS/GOGAT pathway [25]. In contrast to other bacteria, in cyanobacteria, the GS activity is mainly regulated by small inhibitory proteins. Such proteins have been characterized in the Synechocystis PCC 6803 strain: inactivating factor 7 (IF7) encoded by gifA and inactivating factor 17 (IF17) encoded by gifB [26]. The global nitrogen control factor NtcA controls the expression of these genes responding to the 2-OG levels of the cell. There is no sensor protein interacting with NtcA. For the regulation of NtcA activity, PII is dispensable. Most cyanobacterial genomes harbor only one glnB gene, which encodes a PII protein. The others possess both glnB and glnK [27]. NtcA has been demonstrated to respond to 2-OG [28]. A global repressor of carbon-regulated genes, NdhR has been observed to use 2-OG as corepressor [29]. 2-OG appears as a main indicator of the status of central metabolism, carbon, and nitrogen catabolism. Furthermore, the regulation can be mediated by a non-coding RNA, a glutamine riboswitch in the 5′UTR of gifB, and by nsiR4. This glutamine riboswitch links the glutamine status of the cells to regulation of the central nitrogen assimilation reaction [27].
Several signaling pathways exist at different levels in cyanobacteria. Nitrogen-regulated genes nirB and ntcB have been characterized for example in Synechococcus, where they constitute an operon (nirB-ntcB) transcribed different from nirA. Under nitrogen limitation, transcription of nirB-ntcB is increased and is NtcA-dependent, whereas ntcB codes a member of the LysR family of transcriptional activators [6]. Cyanobacteria are able to maintain nitrogen fixation that can occur in cells heterocysts, which differentiate from the vegetative cell filaments. Nitrogen fixation as well as heterocyst development are controlled by ammonium repression. Ammonium represses expression of the nitrate reductase, nitrate/nitrite transport system, nitrite reductase, GSI, and the GSIII structural gene glnN [6,25,30].
1.3. Nitrogen Assimilation in Gram-Positive Bacteria
Gram positive bacteria are classified in two phyla: Firmicutes with low GC DNA content (subdivided into the classes Bacilli and Clostridia) and Actinobacteria with high GC DNA content (subdivided into multiple orders). The order Actinomycetales is the largest one, containing a variety of species and unclassified isolates. The nitrogen assimilatory enzymes of the GOGAT/GS pathway as well as GlnK and GlnE are conserved in Gram-positive bacteria such as Clostridium acetobutylicum, Bacillus subtilis, and Streptomyces coelicolor. However, while the enteric bacteria possess the Ntr control system, there is no reported presence of this system in Gram-positive bacteria. Instead, they possess alternative transcriptional regulatory mechanisms.
1.3.1. Nitrogen Assimilation and Its Control in Bacillus subtilis
The model organism for Gram-positive bacteria with low GC content is Bacillus subtilis. Nitrogen metabolism has been extensively investigated in this organism. A preferred nitrogen source for this bacterium is ammonium [31]. No GdhA activity has been reported in B. subtilis—ammonium can be assimilated only via the GS/GOGAT pathway [32]. Furthermore, no post-translational modification of the GS enzyme has been observed, but feedback inhibition of GS by glutamine has been described. Under nitrogen depletion, three global regulators, GlnR, TnrA, and CodY, are present in cells for best possible growth [2]. Another regulator, GltC, controls the transcription of the glutamate synthetase (GOGAT) gene in dependence to the intracellular concentration of 2-oxoglutarate [33]. The transcriptional regulator TnrA that belongs to the MreR family of DNA-binding proteins has been shown to be functional only under nitrogen-limiting conditions in B. subtilis. The TnrA regulator positively controls the transcription of genes involved in ammonium uptake (amtB, ureABC, nasBC, and nasDEF) and nitrogen signaling (glnK), as well as its own transcription. In addition, it exerts a negative effect on the glnRA operon encoding GlnR and the GS, on the gltAB operon encoding GOGAT, and on gltC encoding GltC [34]. A homologue of the TnrA regulator, GlnR, can target almost the same binding sequences as TnrA. GlnR is active under nitrogen excess and acts mainly as a repressor. It represses the transcription of the glnA gene encoded in the glnRA operon as well as the transcription of tnrA, gltAB, ureABC, and glnR [35,36]. In B. subtilis, CodY-dependent regulation occurs under carbon and nitrogen depletion conditions when bacteria have to rely on amino acids for growth. The repressor protein CodY controls the transcription of urease encoding operon ureABC, dipeptide degradative operon dpp, histidine degradative operon hut, and isoleucine/valine degradative operon bkd [34].
1.3.2. Nitrogen Assimilation and Its Control in Corynebacterium glutamicum
The model organism for Gram-positive bacteria with high GC content is Corynebacterium glutamicum, which has been extensively studied because of its application in industry for the production of amino acids such as L-glutamine. C. glutamicum also serves for studies of important human pathogens including Corynebacterium diphteriae, Corynebacterium jeikeium, Mycobacterium leprae, and Mycobacterium tuberculosis. C. glutamicum possesses the genes amtA and amtB for ammonium uptake [37], which are transcribed under nitrogen starvation conditions [38]. AmtA specifically transports methylammonium; AmtB transports ammonium with high affinity and methylammonium with low affinity [39,40]. In C. glutamicum, amtB forms a cluster with glnK coding for the signaling protein PII. Besides the nitrogen-sensing function, GlnK has reportedly been involved in the transcriptional regulation of nitrogen metabolism genes in combination with the global transcriptional regulator AmtR [41]. In C. glutamicum, ammonium is assimilated via the GS/GOGAT pathway under nitrogen limitation or via the Gdh pathway under nitrogen excess. Two GS enzymes have been described in C. glutamicum: GlnA (also annotated as GlnA1, GSI) and GlnA2 (GSI). GlnA is the essential functional glutamine synthetase which is subjected to post-translational modification by the adenylyl transferase enzyme GlnE [42,43]. GlnA2 has been described as a non-essential enzyme which is not subjected to post-translational modifications. In contrast to E. coli, GlnE activity in C. glutamicum is not regulated by GlnK [44]. A central transcriptional regulator of nitrogen metabolism genes in C. glutamicum is AmtR, which belongs to the TetR family of regulatory proteins typically acting as transcriptional repressors. The GlnK protein can interact with AmtR under nitrogen-limiting conditions. As a result of the protein–protein interaction, AmtR is removed from the upstream regions of its target genes and leaves them for transcription. AmtR has been reported to control its own transcription as well as the transcription of 34 other target genes including the amtB–glnK–glnD operon, the gltBD, urtABCDE, gluABCD operons, genes dapD, gdh, codA, the ureABCEFGD operon, genes amtA, glnA1, and crnT [40,45].
1.3.3. Nitrogen Assimilation and Its Control in Streptomyces coelicolor
Intensive investigations of primary and secondary metabolism in S. coelicolor A3 (2) lead to establishment of this Gram-positive bacterium with high GC content as a model organism for Streptomyces and Actinobacteria [46]. In its natural soil habitat, S. coelicolor is an obligatory aerobe bacterium with filamentous growth and GC content of 63–78%. It belongs to the phylum Actinobacteria and genus Streptomyces, and is closely related to human pathogens from genus Mycobacterium and Rhodococcus [47,48,49]. Streptomycetes including S. coelicolor feature an impressive adaptability to environmental stress, a complex metabolism and life cycle, and high metabolic potential, as well as the ability to synthesize a large variety of useful natural products. Streptomyces spp. fulfill a key ecological role in the soil—they are able to naturally recycle the remains of other organisms by their utilization [50].
In the natural soil environment, S. coelicolor lives under varying nutrient conditions. Therefore, it is capable of assimilation of different sources of carbon and nitrogen such as ammonium (NH4+), nitrite (NO2−), nitrate (NO3−), amino acids histidine and arginine, amino sugars, peptides, urea as well as chitin, cellulose and xylose, releasing chitinases, cellulases, and xylanases for extracellular utilization of such organic material [51,52,53]. The genome of the S. coelicolor M145 strain was the first to be fully sequenced and is remarkably large: a linear chromosome contains 8,667,507 base pairs and 7825 predicted genes [49]. Diverse multiple gene clusters are present in the genome of S. coelicolor for the synthesis of natural products, such as actinorhodin, undecylprodigiosin, methylenomycin, perimycin, and calcium-dependent antibiotic [53]. Regulatory networks that control the metabolism and morphological differentiation of S. coelicolor in respond to nitrogen availability signals allow S. coelicolor to survive under variable nutrient conditions [54].
S. coelicolor is constantly exposed to environmental stress. This occurs in the form of limited availability of nutrient sources and local excess of toxic compounds as a result of the decomposition of organic material. The cellular response to nitrogen limitation in S. coelicolor can be controlled on a transcriptional level—it involves the global nitrogen response regulator GlnR [55,56]. It belongs to the OmpR family of regulators and influences by activation the expression of the operon amtB-glnK-glnD (the ammonium transporter AmtB, the PII signal protein GlnK, and the adenylyl transferase GlnD) that are conserved genes in Actinomycetales [57,58,59,60,61,62]. Other GlnR target genes include glutamine synthetase encoding genes glnA and glnII, a glutamate dehydrogenase encoding gene gdhA, nitrate/nitrite reduction genes nirB and nasA, urea cleavage gene ureA, and a HemD-like transcriptional regulator encoding gene nnaR, as well as seven further genes with an unknown function [57,58,61,63,64].
Two transcriptional regulators, GlnR and NnaR, control nitrate assimilatory genes (nirBD, narK, nasA) in S. coelicolor. Under nitrogen-limited conditions, GlnR can activate the expression of these genes that is enhanced by the synergistic binding of GlnR and NnaR in the presence of nitrate [65]. Another regulator, GlnRII, can bind to the upstream regions of amtB-glnK-glnD, glnA, and glnII as well as sco1863 showing the same binding capacity as GlnR [60]. It was hypothesized that GlnRII has a particular role in the glnII regulation and is not a functional homologue of GlnR. It was found in Streptomyces spp., but has not been found in Corynebacterium and Mycobacterium [57,66].
Complex nitrogen metabolism of S. coelicolor requires additional control by further transcriptional regulators: Crp [67], PhoP [68,69], ArgR [70], AfsR [71], DasR [72,73], and AfsQ1 [74]. Crp regulates the interplay of primary and secondary metabolism, and the genes amtB-glnK-glnD, glnA, and glnII [67]. PhoP negatively regulates the transcription of the amtB-glnK-glnD operon as well as genes glnA, glnII, and glnR under conditions of phosphate limitation [26]. AfsR controls expression of glnR in response to unknown nutrient stress stimulus [71]. AfsQ1 is required for the carbon, nitrogen, and phosphate metabolism regulation under glutamate presence [74,75].
In dependence from the level of ammonium, the uptake in S. coelicolor can be controlled at post-translational level—the ammonium transporter AmtB interacts with the nitrogen sensor protein PII (GlnK) [57,76,77]. When concentrations of ammonium are high, GlnK can be inactivated by adenylylation mediated by the adenylyltransferase GlnD or by proteolysis [57]. In order to avoid the depletion of the intracellular glutamate pool, the activity levels of glutamine synthetases can be controlled at post-translational level [45]. When nitrogen conditions are variable, GlnA activity can be regulated through the reversible adenylylation/deadenylylation by an adenylyltransferase GlnE [60]. However, no post-translational modifications of the GOGAT enzyme in Actinobacteria have so far been reported. The regulation of GSI by GlnE has also been demonstrated in E. coli, where GlnE can be controlled by GlnK, GlnB (nitrogen regulatory protein P-II), and GlnD. However, in S. coelicolor, the PII proteins GlnK and GlnD seem to be not essential for these processes [45,77,78].
1.3.4. Nitrogen Assimilation and Its Control in Mycobacterium tuberculosis
Nitrogen metabolism has been extensively studied in pathogenic Actinobacteria from the genus Mycobacterium, which includes soil bacteria such as Mycobacterium smegmatis as well as mammal and human pathogens including Mycobacterium tuberculosis (causative agent of tuberculosis) and Mycobacterium leprae (causative agent of leprosy). In M. tuberculosis, the glutamine synthetase and nitrogen assimilatory protein GlnA (also referred as GlnA1) have been associated with pathogenicity and virulence [79,80]. Understanding of nitrogen assimilation in this bacterium is important for the comprehension of infection mechanisms. Furthermore, this knowledge allows the development of novel therapeutic strategies to control M. tuberculosis and multidrug resistance (MDR) strains. M. tuberculosis possesses an ammonium transporter protein, AmtB, and it does not have an active glutamate dehydrogenase (GDH) enzyme. Thus, GS/GOGAT is the only way to assimilate nitrogen. M. tuberculosis genome contains one glutamine synthetase encoding gene glnA (also referred as glnA1) and three GS-like enzymes encoded by glnA2, glnA3, and glnA4 [81,82]. All of these enzymes belong to the GSI (prokaryotic) type of glutamine synthetases. Although all GS-like enzymes have been reported to be active in cells, only GSI encoded by glnA1 has been found to be essential for M. tuberculosis growth. The activity of the GlnA1 enzyme can be down-regulated under nitrogen excess by the bifunctional adenylyl transferase GlnE. Under nitrogen starvation, GlnE deadenylylates GlnA1 and restores its activity. In contrast to E. coli and similar to S. coelicolor, in M. tuberculosis, GlnE activity is not regulated by GlnK and GlnD [23]. At transcriptional level, GlnR, which is a functional homologue of the global transcriptional regulator GlnR from S. coelicolor, controls the nitrogen assimilation. As occurs in S. coelicolor, during nitrogen limitation in M. tuberculosis, GlnR regulates the transcription of glnA as well as the transcription of operons amtB-glnK-glnD, gltBD, and nirBD, and at least 33 other genes in M. tuberculosis and more than 100 genes in M. smegmatis [83,84]. A putative TetR-like transcriptional regulator, AmtR, has also been found in M. tuberculosis (Rv3160c) demonstrating only 27.9% amino acid sequence identity to the AmtR protein from C. glutamicum [45].
1.4. The Central Role of Glutamine Synthetases in the Bacterial Nitrogen Metabolism
Glutamine synthetases (GS; EC 6.1.1.3) are enzymes found in all forms of life, with the central role in nitrogen assimilation [85]. GS enzymes catalyze the formation of L-glutamine through the condensation of ammonia with L-glutamate in an ATP-dependent manner. Glutamine, together with glutamate, serves as an essential component for protein biosynthesis. It is a major nitrogen source for biosynthetic reactions in the cell and one of the major nontoxic ammonia carriers [86,87]. It has been assumed that glutamine synthetases might be among the most ancient existing enzymes in nature [88]. Functional GSs have been described to occur in three forms. GS type-1 (GSI) has been found in most prokaryotes [6,21,22] as well as in mammals and plants [89,90]. GSI enzymes are subdivided into two GS isoenzymes: GSI-α and GSI-β. GSI-α enzymes are generally found in thermophilic bacteria, low G + C Gram-positive bacteria, and euryarchaeota, whereas GS I-β enzymes are found in other bacteria [34]. GS type-2 (GSII) are generally found in eukaryotes, in some Gram-positive high GC-content bacteria [6,45,91], and in symbiotic Gram-negative soil bacteria such as Rhizobium sp. [88,92]. GS type-3 (GSIII) are generally found in cyanobacteria [93], in the Gram-negative anaerobe bacterium Bacteriodes fragilis [85], and some protozoans [94]. Another octamer form of GS, GSIV, has so far only been found in the plant-associated bacteria Rhizobium leguminosarum [95], Rhizobium meliloti [96], and Agrobacterium tumefaciens [97]. The biosynthetic activity of GSIV is significantly lower than that of GSI and GSII, which is the reason for discussions about whether the enzyme primarily fulfills a function other than the synthesis of glutamine [96]. However, this could not yet be demonstrated.
Structurally, all GSs are composed of two closed-ring structures. Active sites are formed between protomers [98]. GSI is a dodecameric protein with about 360 amino acids length and a subunit between 44–60 kDa that has been found in bacteria and archea [99,100,101,102]. GSII is a dodecamer with about 450 amino acids length, composed of subunits between 35–50 kDa [88,100,101,102]. GSIII is a hexameric enzyme with about 730 amino acids length, composed of subunits of approximately 75 kDa each [93,103].
The crystal structure of the glutamine synthases has been previously elucidated and characterized in Salmonella typhimurium [104,105], Helicobacter pylori [106], Bacillus subtilis [107], and Mycobacterium tuberculosis [108]. The GSI GlnA has been structurally elucidated in S. typhimurium (PDB: 2GLS). It consists of 12 subunits and demonstrates a high sequence similarity to prokaryotic GS enzymes [98,109]. The crystal structure of GSII (GlnII) with a resolution of 2.55 Å (PDB: 4BAX) has been determined in S. coelicolor and described by X-ray diffraction. GlnII is a dodecamer comprising 10 subunits organized in 2 rings that demonstrates high sequence similarity to eukaryotic GS enzymes [7,66,110]. The glnII gene has been detected in Streptomyces sp. but is not present in other Actinobacteria such as Corynebacterium and Mycobacterium [91] and is not transcribed at a constant level at all growth phases, though it is preferentially transcribed during mycelial differentiation [60]. The crystal structure of the GSIII (PDB: 3O6X) has been determined in B. fragilis—it is a dodecamer with two hexameric rings [103].
Furthermore, a computational analysis of glnA-genes across actinobacterial genomes revealed the existence of a supposed common glnA ancestor, from which the glnA, glnII, and glnA-like genes in different Actinobacteria are derived [88,111]. For example, in S. coelicolor and in M. tuberculosis, three genes were identified as glnA-like: glnA2, glnA3, and glnA4 [57,112]. GlnA and GlnII were demonstrated to be functional glutamine synthetases, and glnA2, glnA3, and glnA4 were shown to encode GS-like enzymes that do not exhibit a glutamine synthetase activity in both S. coelicolor [57,112,113] and M. tuberculosis [80].
In S. coelicolor, analysis of GlnA2, GlnA3, and GlnA4 revealed that they share similar structural features [7]. A phenotypic analysis of glnA2, glnA3, and glnA4 knock-out mutants with different nitrogen sources demonstrated their involvement in polyamine and ethanolamine metabolism [113,114,115]. Biochemical studies on GlnA2 and GlnA3 demonstrated that they are active as γ-glutamylpolyamine synthetases [113,115]. Furthermore, it has been shown that GlnA4 is a γ-glutamylethanolamide synthetase [114]. The presence of different GS-like proteins highlights the occurrence of these specialized proteins required for the survival, colonization, and propagation in specific habitats.
2. Polyamine and monoamine Metabolism
2.1. Polyamine Metabolism in Bacteria
2.1.1. Distribution and Role of Polyamines
Polyamines are aliphatic polycations that are composed of a polycarbone chain and amino groups. Widely distributed natural polyamines are putrescine (1,4-diaminobutane), spermidine (N-(3 aminopropyl)-1,4-butadiamine), spermine (N, N′-bis (3-aminopropyl-1,4-butanediamine)), and cadaverine (pentane-1,5-diamine) [116,117,118]. Further polyamines that are not frequently occurring in nature have been reported, including thermine, thermospermine, caldopentamine, and others. These molecules have been found in the extreme thermophile Thermus thermophiles [119]. A variety of other linear polyamines have been found in (hyper)thermophilic archea and bacteria, including norspermidine, norspermine, caldopentamine, homocaldopentamine, thermopentamine, and caldohexamine [120], as well as branched-chain polyamines such as N4-bis(aminopropyl)spermidine [121].
Polyamines are present in diverse bacterial habitats. In soil, the polyamine concentration per gram of humus ranges are: putrescine: between 0.28 and 0.56 nmol/g, spermidine: 0.23–0.62 nmol/g, and spermine: 0.16–0.43 nmol/g [122]. In mammal cells, a natural environment for pathogenic bacteria, elevated polyamine levels have been reported exceeding physiological concentrations in human skin, in regenerating tissues (more than 1 mM), and in body fluids and blood (more than 0.01 mM) [123,124,125]. In addition, polyamine levels increase in human lungs during an inflammation process and shortly after apoptosis [126]. However, increased polyamine concentrations can lead to bacterial cell death. For example, it has been reported that 0.15 mM of exogenous putrescine is toxic for the cyanobacterium Anacystis nidulans [127]. Other reports described inhibition of E. coli growth in the presence of 4 mM spermidine [128].
Natural intracellular polyamine concentrations range in different species. E. coli can synthesize high amounts of putrescine (up to 32 mM total concentration) and spermidine (up to 6.88 mM). However, in most bacteria, the content of intracellular putrescine ranges between 0.1–0.2 mM [3,129]. Spermidine has been shown to be essential for planktonic growth of some Gram-negative bacteria, such as ε-proteobacterium Campylobacter jejuni [130] and γ-proteobacterium Pseudomonas aeruginosa PAO1 [131]. The intracellular spermidine content can vary between 1–3 mM. In contrast to other polyamines, the presence of cadaverine and spermine in bacterial cells remains not well investigated. In E. coli, spermine is not biosynthesized de novo, but it can be taken up from the environment [132]. In other bacteria, spermine was found in cells when present in the medium [133]. Cadaverine has been characterized in bacteria and plants. It is the least prevalent of polyamines that naturally occur in cells and is of low importance for bacteria [134]. It is normally absent in E. coli [133].
Polyamines, especially spermidine, have been described as essential in Archaea and eukaryotes. However, their role in bacteria is less understood [13,118]. It has been reported that polyamines are not required for normal growth in Gram-positive bacteria such as Bacillus subtilis [135] and S. coelicolor [136]. In γ-proteobacteria, polyamines have been shown to be required for growth—their limitation reduced the growth rate in Yersinia pestis [137], Vibrio cholera [138], Salmonella typhimurium [139], and Escherichia coli [140]. The intracellular polyamine amount is tightly coordinated with the cellular metabolism. Polyamines have been demonstrated to be accumulated intracellularly under stress conditions [117]. In bacteria, polyamines are important for homeostasis: their functions include an influence on transcription [140,141,142,143] and translation [144,145,146,147], the biosynthesis of siderophores [136,148,149,150], cell growth stimulation [151,152], and biofilm formation [138,153,154]. Furthermore, polyamines play a role in stress response: they confer the response to oxidative stress [128,140,155,156], SOS system activation [157], acid resistance [155,158,159], and antibiotic resistance [160,161,162,163].
2.1.2. Importance of Polyamines for Intracellular Pathogens
In mouse macrophages, the concentration of polyamines putrescine, spermidine, and spermine reportedly vary from 250–1750 pmol/5 × 106 macrophages, depending on the metabolic state of the cell [164]. Polyamines synthesized by the host can favor intracellular survival of human pathogens causing deadly diseases. Such bacteria are able to colonize and manipulate immune cells, escaping the response. This makes the treatment of infections extremely difficult. Macrophages represent the first line of human immune defense. Most intracellular pathogens residing in macrophages cause a time-dependent up-regulation of the metabolic regulator (PPARγ) in infected macrophages, resulting in increased expression of M2 markers and down-modulation of the M1 response [165]. PPARγ induces the arginine metabolism leading to the synthesis of the polyamine spermine from putrescine via spermidine [166]. Bacteria that are able to colonize and survive in macrophages, providing access to intracellular polyamines, include Klebsiella pneumonia causing pneumonia infection [167], Salmonella typhimurium causing typhoid fever [168], Brucella abortus causing brucellosis [169], Acinetobacter baumannii causing meningitis or lung infections [170], and Mycobacterium tuberculosis causing tuberculosis [171].
2.1.3. Occurrence of Polyamines in Bacterial Cells
Polyamine Biosynthesis
In bacteria and in all kingdoms of life, polyamines can be synthesized from the amino acids methionine, ornithine, lysine, and arginine [117,172]. Bacterial polyamine biosynthetic pathways are configured in multiple pathways that have been studied in E. coli, P. aeruginosa, Salmonella enterica, Campylobacter jejuni, and S. coelicolor [118]. Generally, bacteria produce a diamine (putrescine or cadaverine) and triamine (spermidine). Some bacteria can produce longer-chain polyamines (including spermine), some produce only diamines, and others, such as pathogens, often do not produce any polyamines [173].
Putrescine is synthesized by ornithine decarboxylation involving an ornithine decarboxylase (ODC). Furthermore, putrescine can be generated from arginine by an arginine decarboxylase (ADC) and agmatinase. Both ODC and ADC pathways have been extensively investigated in E. coli [117,174]. In P. aeruginosa, agmatine conversion to putrescine in a two-step reaction has been demonstrated. It involves hydrolytic deimination of agmatine to N-carbamoylputrescine catalyzed by agmatine deiminase AguA. Subsequently, N-carbamoylputrescine amidohydrolase AguB catalyzes hydrolysis of the carbamoyl group yielding ammonia, carbon dioxide, and putrescine [131,175]. The second reaction has also been shown in Enterococcus faecalis, where putrescine transcarbamylase releases carbamoylphosphate from N-carbamoylputrescine and inorganic phosphate yielding putrescine [176]. Spermidine and spermine are derived from putrescine via addition of aminopropyl groups, which are supplied by the methionine derivative—decarboxylated S-adenosylmethionine (dcAdoMet, dSAM) produced by a S-adenosylmethionine decarboxylase (AdoMetDC, SpeD). The addition of aminopropyl groups occurs through spermidine and spermine synthetases (SpdS/SpeE and SpmS) [117,174,177]. Cadaverine has been studied in E. coli, Lactobacillus spp. and Vibrio sp., where it is synthesized by lysine decarboxylases (LDCs)-dependent lysine decarboxylation [178] (Figure 2). In S. coelicolor, low amounts of putrescine, spermidine, spermine, and cadaverine (ca. 0.05–0.1 μmol/g) have been detected when grown in a complex medium [120]. It has been reported that putrescine, spermidine, and diaminopropan can occur in S. coelicolor in the late-stationary phase in the minimal liquid medium (NMMP), while cadaverine can be produced under iron limitation [136], hinting towards a possibility to synthesize these polyamines de novo in S. coelicolor. Knowledge of polyamine biosynthetic pathways have allowed advances in engineering of the industrial bacterium C. glutamicum for efficient production of the most promising polyamines putrescine and cadaverine [179].
Polyamine Uptake
On the one hand, some bacteria do not possess a polyamine biosynthetic pathway and must import polyamines from the extracellular environment. On the other hand, the uptake of polyamines is generally economical for bacteria and allows energy to be saved, because the synthesis of S-adenosylmethionine (AdoMet, SAM), for which biosynthesis requires ATP, is consumed in the AdoMet decarboxylase–spermidine synthetase pathway. The uptake of external polyamines is of central importance in order to optimize growth, metabolism, cell-to-cell communication, and adaptation to the environment [14].
At physiological pH, polyamines are positively charged hydrophilic molecules. They cannot pass through cellular membranes by diffusion. Thus, an active transport system is needed for polyamine uptake from the extracellular environment. Polyamine transport has been extensively investigated in predominantly pathogenic bacteria, including Vibrio cholerae, Proteus mirabilis, Aggregatibacter actinomycetemcomitans, Streptococcus pneumonia, and Escherichia coli.
Several polyamine transporters have been described in E. coli: PuuP for putrescine [180,181], the putrescine–ornithine antiporter PotE [182], PotFGHI for putrescine [183], the proton-dependent importer PlaP for putrescine [184], the cadaverine–lysine antiporter CadB [185], the importer PotABCD for spermidine [186,187], and the spermidine transporter MdtJI [188] (Figure 3; Table 1). In V. cholerae, three homologues of PotD from E. coli have been described: NspS, PotD1, and PotD2 [189].
Polyamine uptake has been shown to influence the pathogenicity. The periplasmic substrate-binding protein PotD1 has been shown to be responsible for spermidine uptake that hinders biofilm formation. Instead, a polyamine norspermidine enhances biofilm formation via the NspS/MbaA signaling system in V. cholerae [190]. In A. actinomycetemcomitans, a homolog of PotD from E. coli was also reported to correlate with biofilm formation causing periodontitis [191]. In S. pneumonia, the uptake of spermidine by the spermidine importer PotABCD was reported to be important for pathogenicity in mice [133,192]. In P. mirabilis, putrescine uptake was demonstrated to be carried out by the proton-dependent putrescine importer PlaP [193].
2.1.4. Polyamine Assimilation in Bacteria
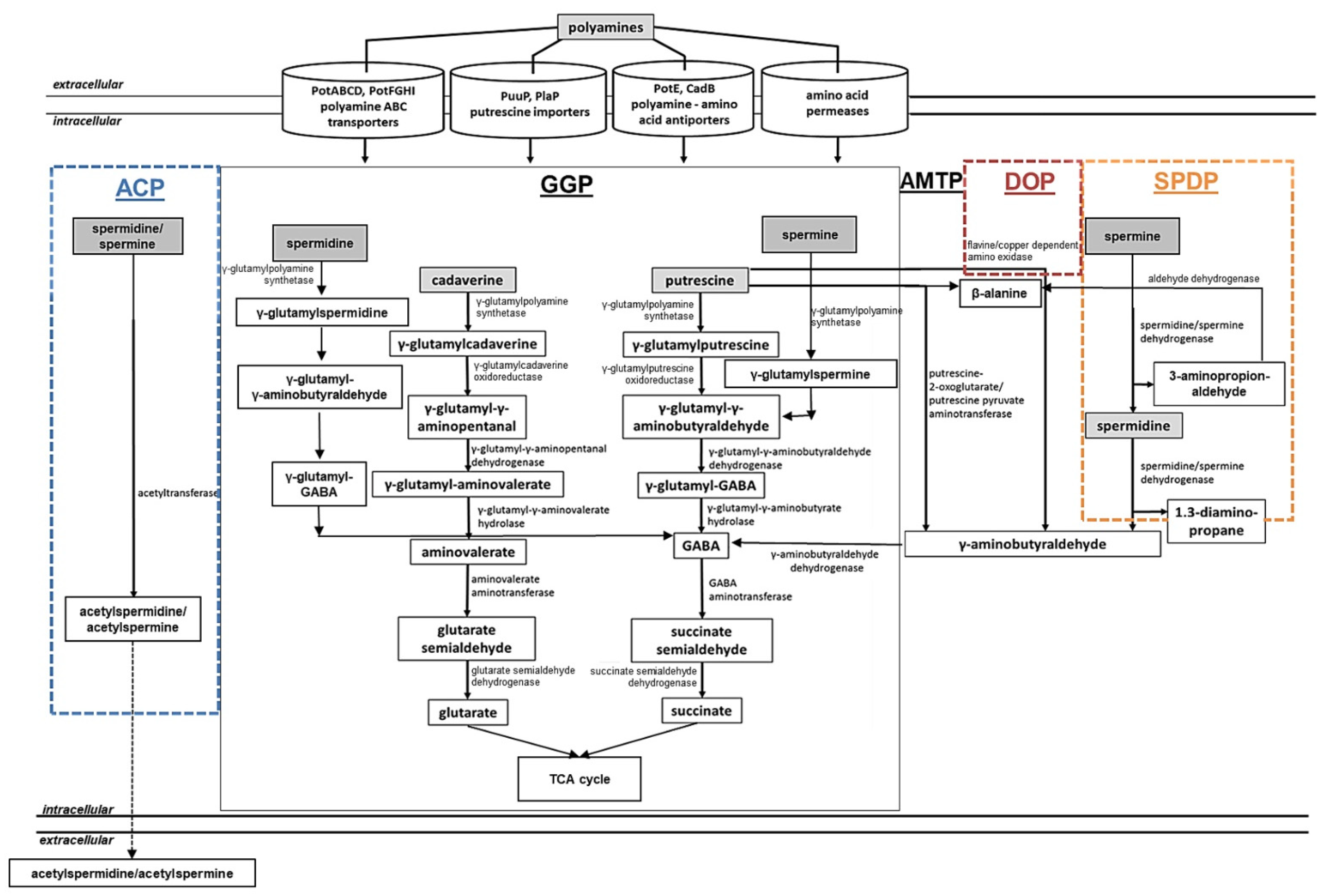
Rather low amounts of polyamines can be synthesized intracellularly for maintenance of cellular functions. However, in the extracellular environment, polyamines can be present in excess, resulting in locally elevated and toxic amounts of these compounds. Under these conditions, detoxification of increasing concentrations of intracellular polyamines is required to avoid cell death. On the other hand, some bacteria do not have a complete pathway for polyamine biosynthesis (Enterococcus faecalis) or lack it completely (Staphylococcus aureus), thus relying on polyamine uptake and utilization [194]. It has been reported that bacteria can utilize polyamines as a source of nitrogen and carbon to control the intracellular polyamine pool. This process has been investigated in the Gram-negative bacteria E. coli and P. aeruginosa as well as in the Gram-positive bacteria Staphylococcus aureus, Bacillus subtilis, C. glutamicum, and S. coelicolor [7,14,15,180,195,196,197,198]. The following pathways have been characterized in E. coli, P. aeruginosa, B. subtilis, and S. coelicolor: the gamma-glutamylation pathways (Figure 3, GGP), the aminotransferase pathway (Figure 3, AMTP), the direct oxidation pathway (Figure 3, DOP), the spermine/spermidine dehydrogenase pathway (Figure 3, SPDP), and the acetylation pathway (Figure 3, ACP).
Polyamine Utilization in E. coli
The gamma-glutamylation pathway (also referred as the putrescine utilization pathway) has been characterized in E. coli and P. aeruginosa [180,199,200,201] (Figure 3, GGP). In E. coli, extracellular putrescine can be transported into the cell by the PuuP transporter and afterwards glutamylated by the γ-glutamylputrescine synthetase (PuuA) in an ATP-dependent manner [180,200] (Table 1). In further steps, the pathway involves the γ-glutamylpolyamine oxidoreductase PuuB, the γ-aminobutyraldehyde dehydrogenase PuuC, the γ-glutamyl-GAΒA hydrolase PuuD, the GABA aminotransferase PuuE, and the succinate semialdehyde dehydrogenase leading to the production of succinate [199,202] (Figure 3, GGP; Table 1). A cadaverine-specific utilization pathway has not been reported in E. coli, but the activity of PuuA with cadaverine was comparable of that towards putrescine in vitro [14,203]. Presumably both the aminotransferase and the glutamylation pathway of putrescine are involved in cadaverine metabolism in E. coli.
The aminotransferase pathway has been investigated in E. coli [201,204,205] (Figure 3, AMTP). In this pathway, putrescine is metabolized to γ-aminobutyric acid (GABA) via the intermediate γ-aminobutyraldehyde. This metabolic route involves the enzymes putrescine aminotransferase PatA, the γ-aminobutyraldehyde dehydrogenase PatD, the GABA aminotransferase GabT, and the succinic semialdehyde dehydrogenase GabD ending with the formation of succinate that can be further used in the TCA cycle [14,159] (Figure 3, AMTP; Table 1). The activity of the PatA enzyme on cadaverine is reportedly comparable to that towards putrescine.
The acetylation pathway has been described for the utilization of spermidine in E. coli and also in B. subtilis [206,207,208] (Figure 3, ACP). It occurs via the spermidine acetyltransferase that acetylates spermidine to acetylspermidine using acetyl-CoA [209]. However, the further fate of acetylspermidine in E. coli is not yet known (Figure 3, ACP).
Interestingly, the intracellular spermidine concentrations may be also reduced by the glutathionylspermidine synthetase/amidase (GSP synthetase) in E. coli and Haemophilus influenzae. In the reaction catalyzed by this enzyme, spermidine is ligated with glutathione forming glutathionylspermidine in an ATP-dependent manner [210,211].
Polyamine Utilization in P. aeruginosa
In P. aeruginosa PAO1, the gamma-glutamylation pathway (Figure 3, GGP) includes almost identical metabolic steps as that in E. coli [196,212], but is represented by seven pauA genes, four pauB genes, one pauC gene, and two pauD genes that are thought to be responsible for polyamine catabolism [195,196] (Table 1). Interestingly, each PauA1-PauA7 enzyme seems to have different substrate specificity towards different mono- and polyamines being involved in the first step of the pathway [195,196].
The aminotransferase pathway (Figure 3, AMTP) of P. aeruginosa PAO1 includes a putrescine-pyruvate aminotransferase that generates γ-aminobutyraldehyde and L-alanine [213,214]. The KauB protein that corresponds to PatD from E. coli oxidizes in the following step, with γ-aminobutyraldehyde forming GABA, which is further catabolized to succinate by GabT and GabD [14,212] (Figure 3, AMTP; Table 1).
The direct oxidation pathway (Figure 3, DOP) has been investigated in P. aeruginosa and Staphylococcus aureus. It requires an amine oxidase [195,215,216,217,218,219] (Figure 3, DOP). The spermine/spermidine dehydrogenase pathway has been described in P. aeruginosa [195,212], for which the structure of the essential enzyme spermidine dehydrogenase has been reported [216] (Figure 3, SPDP). In P. aeruginosa PAO1, the spermidine dehydrogenase (SpdH) can cleave spermidine into 1,3-diaminopropane and γ-aminobutyraldehyde and spermine into spermidine and 3-aminopropanaldehyde. KauB oxidizes 3-aminopropanaldehyde to β-alanine, which is further catabolized to acetyl-CoA [195,212] (Table 1).
Polyamine Utilization in S. coelicolor
In contrast to Gram-negative bacteria E. coli and P. aeruginosa, the utilization of polyamines has barely been investigated in Gram-positive bacteria. While some studies report the acetylation of polyamines by S. aureus [197], B. subtilis [206], and C. glutamicum [198], extensive investigations of polyamine catabolism have been reported in S. coelicolor. A gamma-glutamylation pathway for polyamine utilization in S. coelicolor has been described in a combined in silico and transcriptional analysis [7] (Figure 3, GGP). The first step is catalyzed by GlnA2 and GlnA3 enzymes that are able to glutamylate the polyamines putrescine, spermidine, spermine, and cadaverine [113,115]. Based on in silico and transcriptional studies, the following steps of the pathway were postulated. In the second step, γ-glutamylpolyamines are further reduced by the γ-glutamylpolyamine oxidoreductase (SCO5671). This enzyme is an ortholog of the γ-glutamylpolyamine oxidoreductases PauB1-B4 in P. aeruginosa and PuuB in E. coli (Table 1). The subsequent step is catalyzed by the dehydrogenases (SCO5657 and SCO5666) that are orthologs of (γ-glutamyl-) γ-aminobutyraldehyde dehydrogenases PuuC and PatD from E. coli. The fourth pathway step needs hydrolases (SCO5657, SCO5666, and SCO6961), resulting in production of γ-aminobutyric acid (GABA) or aminovalerate. Subsequently, an ortholog of GabT from E. coli the GABA aminotransferase (SCO5676) catalyzes the production of succinate semialdehyde or glutarate semialdehyde (Table 1). Remarkably, it has been shown that the expression of sco5676 is induced by arginine [70], which is a precursor of putrescine biosynthesis, as well as in the presence of polyamines [113]. Finally, a homolog of the succinic semialdehyde dehydrogenase GabD from E. coli, the SCO5679 protein, catalyzes the last step producing glutarate or succinate that feed the tricarboxylic acid (TCA) cycle. RNAseq analysis showed that the expression of sco5679 was induced by polyamines, supporting the hypothesis that SCO5679 is involved in the last step of the polyamine gamma-glutamylation pathway in S. coelicolor [7,115] (Figure 3, GGP; Table 1).
The aminotransferase pathway of S. coelicolor includes an aminotransferase. The enzyme SCO5655 is a homolog of the putrescine aminotransferase (PatA) from E. coli (Table 1). The RT-PCR analysis demonstrated that the expression of sco6960, sco6961, and sco5655 was enhanced in the presence of polyamines [113], indicating their involvement in polyamine assimilation (Figure 3, AMTP).
![Medsci 10 00040 g003]()
Figure 3.
Combined model of bacterial polyamine utilization pathways (adapted from [7,115]). Central reactions and metabolic routes of polyamine catabolic pathways, as well as up-to-date polyamine uptake systems described in bacteria, are shown. ACP, acetylation pathway; GGP, gamma-glutamylation pathways (black box); AMTP, aminotransferase pathway; DOP, direct oxidation pathway; SPDP, spermine/spermidine dehydrogenase pathway. Dashed arrows represent predicted and straight arrows confirmed metabolic pathways. Pathways described for the following bacteria—in black: E. coli, P. aeruginosa, and S. coelicolor; dashed orange box: P. aeruginosa; dashed brown box: S. aureus and P. aeruginosa; dashed blue box: E. coli, B. subtilis, and C. glutamicum [7,14,195,197,198,201,206,207,208,212,215,217,218,219].
Figure 3.
Combined model of bacterial polyamine utilization pathways (adapted from [7,115]). Central reactions and metabolic routes of polyamine catabolic pathways, as well as up-to-date polyamine uptake systems described in bacteria, are shown. ACP, acetylation pathway; GGP, gamma-glutamylation pathways (black box); AMTP, aminotransferase pathway; DOP, direct oxidation pathway; SPDP, spermine/spermidine dehydrogenase pathway. Dashed arrows represent predicted and straight arrows confirmed metabolic pathways. Pathways described for the following bacteria—in black: E. coli, P. aeruginosa, and S. coelicolor; dashed orange box: P. aeruginosa; dashed brown box: S. aureus and P. aeruginosa; dashed blue box: E. coli, B. subtilis, and C. glutamicum [7,14,195,197,198,201,206,207,208,212,215,217,218,219].
2.1.5. Regulation of Polyamine Assimilation Genes in Bacteria
Polyamines are involved in the regulation of transcription and translation processes required for bacterial proliferation. Therefore, a strict control of the intercellular polyamine content is of importance. Regulation of the genes from the gamma-glutamylation and aminotransferase pathways has been extensively investigated in E. coli. It allows the detection of elevated polyamine concentrations aiming at their subsequent intracellular utilization. It has been demonstrated in E. coli that the gene control involves the nitrogen regulatory protein C (NtrC), the nitrogen assimilation control protein (Nac), an alternative sigma factor σS, and the alternative sigma factor for nitrogen-controlled genes σ54 [6,14,20]. For instance, the putrescine aminotransferase encoding gene patA is regulated by NtrC, σS, and σ54 [159,203,220]. Its expression can also be subjected to catabolite repression [214].
The gamma-glutamylation pathway is encoded in the puuPADRCBE gene cluster in E. coli [180]. Similar to patA, this gene cluster contains the σS dependent promoters [86,221] and the NtrC-σ54 dependent promoter [86,222]. The genes of the gamma-glutamylation pathway are regulated by the puuR encoded repressor PuuR, which represses the expression of puu genes in the puuAP and puuDRCBE operons [202,223]. It has been demonstrated in E. coli that the expression of the puu genes can be induced by putrescine as well as through the shift from anaerobic to aerobic conditions. FNR and ArcA recognition sites were described in the puuA-puuD intergenic region [224].
In S. coelicolor, the regulation of polyamine utilization genes involves two regulators. It has been demonstrated that the regulation of the γ-glutamylpolyamine synthetase encoding gene glnA3 of the gamma-glutamylation pathway is controlled by a specific regulator SCO5656 (EpuRII) [115]. In RT-PCR and EMSA analysis, EpuRII revealed control of several polyamine-associated genes, including glnA3, sco5676, and sco5977 (Table 1). Another regulator of polyamine utilization genes is the global regulator of the nitrogen metabolism GlnR. This has been demonstrated to control the transcription of the second γ-glutamylpolyamine synthetase encoding gene glnA2. According to EMSA analysis, especially strong binding of glnA2 to the promoter area was observed for the acetylated version of GlnR [115].
2.2. Monoamine Metabolism in Bacteria
2.2.1. Distribution and Role of the Monoamine Ethanolamine
Ethanolamine is a naturally occurring common monoamine. It is a primary alcohol and primary amine that belongs to the class of aliphatic amino alcohols. Ethanolamine is a building block of biomembranes and usually occurs in the form of the second-most-abundant head group for phospholipids—phosphatidylethanolamine [225,226,227]. Phosphatidylethanolamine is a substrate and precursor in several biological pathways and comprises 25–45% of all phospholipids in a cell [228]. Ethanolamine forms choline after methylation and is a nitrogenous base in phospholipids and an essential vitamin [227]. Ethanolamine is a main precursor of phosphoglycerides that are important elements in the structure of choline in cellular membranes [229]. Ethanolamine is an abundant compound in processed food and the intestinal tract content [230,231]. It is prevalent in the gastrointestinal tract environment. Ethanolamine and glycerol occur as a breakdown product after the cleavage of phosphatidylethanolamine by phosphodiesterases [232,233].
The incorporation into cell membranes, the importance for cellular homeostasis, and the biological role of ethanolamine have been reported in a number of bacterial genera, such as Mycobacterium spp. [234], Corynebacterium spp., Enterococcus spp., Listeria spp. and Clostridium spp. [235,236], Chromohalobacter salexigens [237], and Streptomyces spp. [114]. Ethanolamine has been observed to be a donor of nitrogen and carbon for gut-associated Gram-negative bacteria such as Escherichia spp., and Salmonella spp. [235,236]. Since ethanolamine as an alternative nitrogen source favors a competitive advantage for intestinal pathogens, its utilization has been described as a positive virulence factor [236,238].
2.2.2. Ethanolamine Biosynthesis and Uptake in Bacteria
The direct biosynthetic pathway for ethanolamine is the decarboxylation from serine (Figure 4). This metabolic route has been investigated in plants and mammals, but it is barely studied in bacteria. Yeast and bacteria possess a phosphatidylserine decarboxylase and can synthesize phosphatidylserine in a reaction catalyzed by a phosphatidylserine synthase. This enzyme condenses the phosphatidyl moiety of cytidine diphosphate (CDP)-diacylglycerol with serine, resulting in phosphatidylserine [229,239] (Figure 4).
Bacteria can also take up extracellular ethanolamine in order to optimize the metabolism and environment adaptation. The uptake of ethanolamine is possible through the passage into the cell by carrier-mediated transport or diffusion [241]. In E. coli and S. typhimurium, it can be transported into the cell by the specialized transport protein EutH, which is related to permeases [242]. Most Actinobacteria and Proteobacteria can obtain ethanolamine from the extracellular environment using a transporter encoded by the eat gene, which is a functional, non-homologous equivalent to eutH from S. typhimurium [243].
2.2.3. Ethanolamine Assimilation in Bacteria
Ethanolamine utilization as a source of carbon and nitrogen has been reported and investigated in Salmonella, Enterococcus, Arthrobacter, Erwinia, Flavobacterium, Klebsiella, Mycobacterium, Pseudomonas, Achromobacter, Corynebacterium, Clostridium, Vibrio and Escherichia [235,243,244,245,246,247,248], Chromohalobacter [237], and Streptomyces [114]. Members of the Enterobacteriaceae family such as E. coli and S. typhimurium, as well as the members of the phylus Firmicutes, possess long eut operons encoding genes for ethanolamine utilization in a eut-pathway (Figure 5, EUTP). These operons have considerable differences in gene content, organization, and regulation [235,243]. Actinobacteria and most Proteobacteria feature short eut operons, which contain the transporter encoding gene eut together with eutBC and sometimes eutR [235,243]. There are species that contain both long and short eut operons: Klebsiella pneumonia and Pseudomonas fluorescens [243].
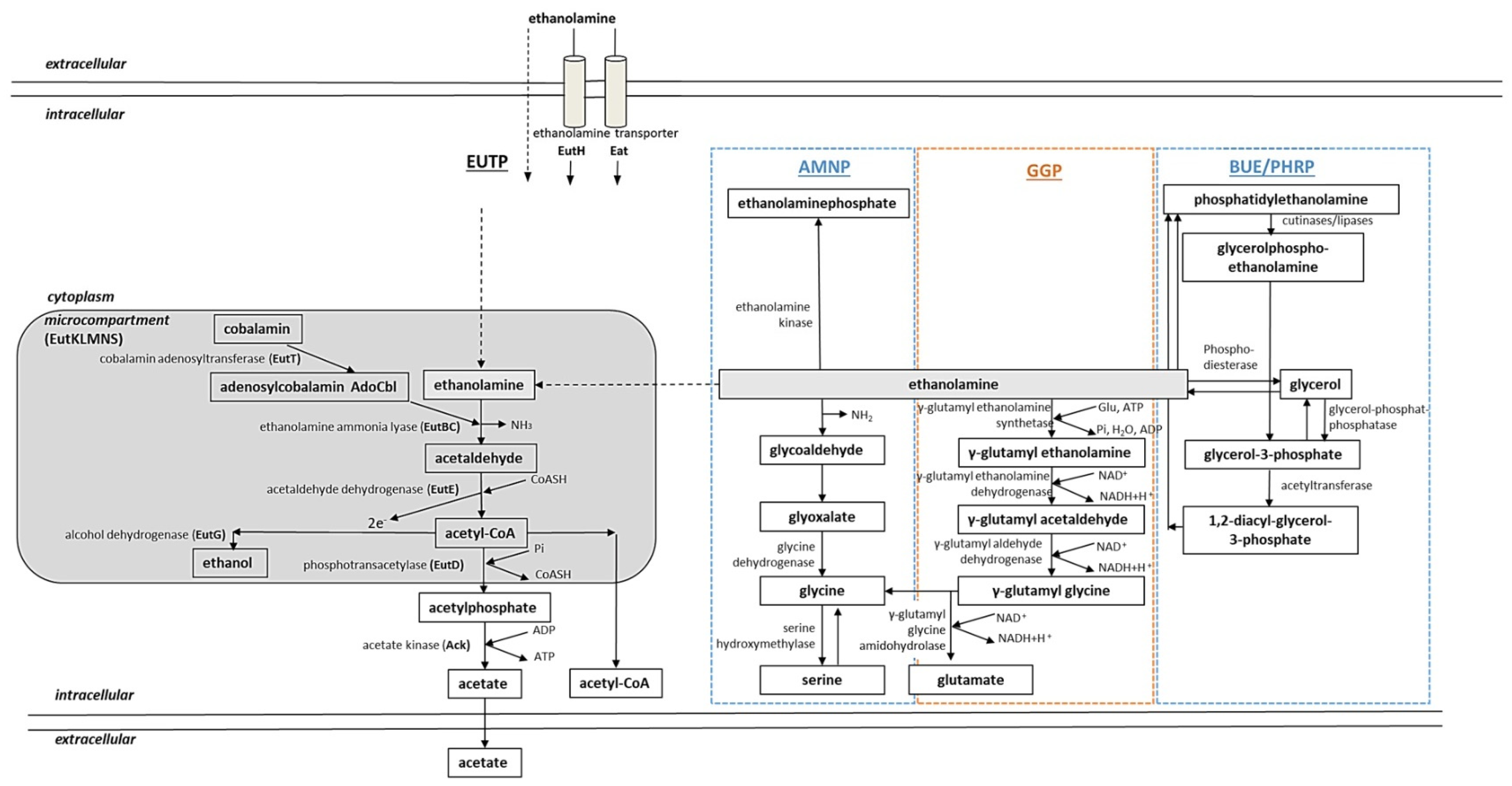
It has been observed that organisms ranging from the Proteobacteria to Actinobacteria possess the capability for ethanolamine metabolism that does not require eut genes. In addition to canonical ethanolamine utilization pathways that involve eut genes and the metabolosome, an alternative conversion of ethanolamine has been reported [114,234,237,249,250,251]. The following pathways have been characterized in E. coli, S. typhimurium, Mycobacterium sp. 607, M. tuberculosis, C. salexigens, and S. coelicolor: the eut pathway (Figure 5, EUTP), the amination pathway (Figure 5, AMNP), polar head recycling (Figure 5, PHRP), biosynthetic utilization of ethanolamine (Figure 5, BUE), and the gamma-glutamylation pathway (Figure 5, GGP).
Ethanolamine Assimilation in E. coli and S. typhimurium
E. coli and S. typhimurium possess similar ethanolamine utilization mechanisms involving proteins encoded by eut genes [231,246] (Figure 5, EUTP). Ethanolamine utilization involves 17 Eut proteins encoded by genes from the ethanolamine utilization operon eut [242,252,253]. Ethanolamine utilization in E. coli and S. typhimurium takes place in a bacterial microcompartment (BMC) [254], also known as the metabolosome [231,255], which is required to retain acetaldehyde. Microcompartments protect the cell from toxic effects by acetaldehyde and prevent the loss of this volatile C-source [256,257].
Ethanolamine can enter the cell by diffusion or via the ethanolamine transporter EutH. After it reaches the microcompartment, the ethanolamine ammonia lyase EutBC breaks down ethanolamine into ammonia and acetaldehyde [252,258,259]. This process requires a cofactor AdoCbl (adenosylcobalamin), which is generated from cobalamin by a corrinoid cobalamin adenosyltransferase EutT [253]. The ammonia serves as a supply of reduced nitrogen. The acetaldehyde is transformed into acetyl-CoA by an acetaldehyde dehydrogenase EutE [252,260]. Acetyl-CoA is used in the TCA cycle, the glyoxylate cycle, and lipid biosynthesis [243]. Acetyl-CoA is converted into acetylphosphate by a phosphotransacetylase EutD or into ethanol by an alcohol dehydrogenase EutG [242]. Acetylphosphate is converted into acetate by an acetate kinase Ack generating ATP [261,262] (Figure 5, EUTP).
The ethanolamine-specific microcompartment of S. typhimurium also contains other structural proteins for microcompartment formation encoded by eutK, eutM, eutS, eutL, and eutN [231,254]. Other proteins encoded by the eut cluster can indirectly influence ethanolamine utilization. Such proteins include a reactivating factor for the ethanolamine ammonia lyase EutA and EutJ (chaperone of EutG and EutE), as well as EutP and EutQ [243].
Ethanolamine Assimilation in S. coelicolor and M. tuberculosis
In S. coelicolor, ethanolamine utilization has been shown to occur via gamma-glutamylation [114] (Figure 5, GGP). In the first step of the pathway, ethanolamine is glutamylated by the γ-glutamylethanolamide synthetase GlnA4 (SCO1613). Based on bioinformatical and transcriptional analysis, it was possible to describe the subsequent steps of the pathway: it involves a γ-glutamylethanolamine dehydrogenase SCO1611, a γ-glutamylaldehyde dehydrogenase SCO1612, and a γ-glutamylglycine amidohydrolase SCO1615. The end products of the pathway are glycine and glutamate [114]. These pathway steps have also been proposed for C. salexigens [237,251] (Figure 5, GGP; Table 2).
Studies in vivo revealed that Mycobacterium sp. are not only able to use host fatty acids from the lipid droplets, but also acquire carbon and nitrogen from phospholipids of the host [217]. The human pathogenic Actinobacteria had to evolve a direct metabolic pathway to strictly control the intracellular ethanolamine due to its potential toxic effect when in excess. Accumulation of ethanolamine might lead to its uncontrolled binding to negatively charged molecules such as DNA or RNA, alkalization of the cellular milieu, and cellular death. It has been demonstrated that Mycobacterium sp. including M. tuberculosis are able to degrade and recycle their own as well as host phospholipids and use them as nutrients [263]. During the recycling process, phosphatidylethanolamine is cleaved by phospholipases. Subsequently, the lipid polar head (glycerolphosphoethanolamine) is converted to glycerol-3-phosphate and ethanolamine by glycerophosphoryl diester phosphodiesterases (Figure 5, PHRP). The phosphodiesterase is essential in vivo during survival of M. tuberculosis in macrophages [264]. Glycerol-3-phosphate can then be channeled for glycolysis, gluconeogenesis, and acylation or broken down to glycerol and phosphate. In Mycobacterium sp., ethanolamine can also be transformed into phosphatidylethanolamine (Figure 5, BUE), which occurs in a pathway involving phosphatidylserine synthase with the intermediate phosphatidylserine (as described in the Section 2.2.2). Furthermore, in Mycobacterium sp., ethanolamine can be transformed to glycoaldehyde and further to glyoxate, which is converted to glycine by glycine dehydrogenase [234,249,250,265] (Figure 5, AMNP).
![Medsci 10 00040 g005]()
Figure 5.
Combined model of bacterial ethanolamine utilization pathways (adapted from [7,114]). Central routes of ethanolamine catabolism with reaction products and involved enzymes, as well as up-to-date polyamine uptake systems known in bacteria, are shown. EUTP: eut-pathway; AMNP: amination pathway; GGP, gamma-glutamylation pathway; BUE: biosynthetic utilization of ethanolamine; PHRP: polar head recycling pathway. Dashed arrows represent diffusion. Gray, rounded rectangle represents the bacterial microcompartment. Pathways in black: E. coli and S. typhimurium; dashed blue box: Mycobacterium sp. and M. tuberculosis; dashed orange box: C. salexigens and S. coelicolor [7,234,236,237,243,249,250,251,254,264].
Figure 5.
Combined model of bacterial ethanolamine utilization pathways (adapted from [7,114]). Central routes of ethanolamine catabolism with reaction products and involved enzymes, as well as up-to-date polyamine uptake systems known in bacteria, are shown. EUTP: eut-pathway; AMNP: amination pathway; GGP, gamma-glutamylation pathway; BUE: biosynthetic utilization of ethanolamine; PHRP: polar head recycling pathway. Dashed arrows represent diffusion. Gray, rounded rectangle represents the bacterial microcompartment. Pathways in black: E. coli and S. typhimurium; dashed blue box: Mycobacterium sp. and M. tuberculosis; dashed orange box: C. salexigens and S. coelicolor [7,234,236,237,243,249,250,251,254,264].
2.2.4. Regulation of Ethanolamine Assimilation Genes in Bacteria
Intracellular ethanolamine content can drastically increase as a result of uptake or cell lysis. In order to reduce elevated ethanolamine concentrations and escape the cytotoxic effects, strict control of the ethanolamine utilization genes is essential. The regulation mechanism of ethanolamine-associated genes in bacteria has been investigated in E. coli, S. typhimurium (the EutR system), and Enterococcus faecalis (the EutV-EutW system). In S. typhimurium, a EutR regulator positively regulates the transcription of the eut operon in the presence of ethanolamine [252,266]. EutR belongs to the AraC family of transcriptional regulators. It binds the promoter of eut in the presence of ethanolamine and AdoCbl as well as the promoter of the eutR gene, providing a low level of constitutive expression [267,268]. Autoregulation of eutR allows the maintenance of induced expression despite competition between EutR and EutBC for AdoCbl, which is required for synthesis of EutBC and for the regulation of the eut operon by EutR [243,267].
E. faecalis does not have the eutR gene. In this organism, ethanolamine utilization is regulated by a two-component system composed of the response regulator EutV and the sensor histidine kinase EutW [245]. It has been demonstrated that eut operon regulation can involve other regulators. For example, the expression of the eut operon in S. typhimurium can also be modulated by the global regulator of invasion genes CsrA, which can lead to increased pathogenicity [266,269]. Furthermore, the expression of the eut operon in E. faecalis can be influenced by the global transcriptional regulator of gelatinase and serine protease encoding genes Fsr [243,269].
In contrast to S. typhimurium and E. faecalis, regulation of ethanolamine utilization in Actinobacteria remains less investigated. It has been shown that in S. coelicolor, the gene sco1614 (epuRI) encodes a regulator of the γ-glutamylethanolamide synthetase encoding gene glnA4. The role of EpuRI as a negative transcriptional regulator of the genes associated with ethanolamine utilization has been proven by transcriptional analysis [114].
3. Recent Advances in Drug Development Targeting Bacterial Nitrogen, Mono- and Polyamine Metabolism
The investigation of the crucial role of nitrogen metabolism and specifically polyamine and monoamine metabolism for survival of pathogenic bacteria and human parasites led to the discovery of a number of potential drugs that might inhibit key enzymes. A large group of such compounds include GS inhibitors that have been extensively investigated in connection to the M. tuberculosis infection and can be described in two categories. The first group are small, highly polar amino acid analogues that target the conserved amino acid binding site, e.g., two of the most widely used GS inhibitors are methionine sulfoximine (MSO) and phosphinothricin (PPT). The second group are the larger, more hydrophobic heterocycles that compete with ATP targeting the nucleotide-binding site, e.g., purine analogs [98,270]. Furthermore, azaserine, an inhibitor of GOGAT in M. tuberculosis, has been identified [271]. GDH has also been described as an imported drug target in Mycobacterium spp.
Another group of inhibitors include compounds that target the polyamine biosynthesis (see pathway, Figure 2). Such inhibitors include the D,L-α-difluoromethylornithine (DFMO). It is a fluorinated ornithine analog that targets polyamine biosynthesis by inhibiting the ODC. DFMO is an approved drug, not only for cancer treatment [272] but also to treat trypanosomiasis [273] and Streptococcus pneumoniae infections [274]. Other validated ODC inhibitors include the putrescine analogs 3-aminooxy-1-aminopropane (APA) and 1,4-diamino-2-butanone (DAB), the agmatine analog 1-guanidinooxy-3-aminopropane (GAPA), and the spermine analog MDL 27695 (N,N′-bis(3-((phenylmethyl)amino)propyl)-1,7-diaminoheptane) that have been shown to be effective against Leishmania spp. [273]. Further inhibitors of polyamine metabolism studied in Leishmania spp. are AdoMetDC inhibitors, e.g., 5-(((Z)-4-amino-2-butenyl)methylamino)-5-deoxyadenosine (MDL 73811) and CGP 40215A (a diamidine and bicyclic analog of MGBG), as well as SpdSyn inhibitors, e.g., hypericin [172,273]. Other polyamine analogs include transport inhibitors such as Ant4 analogs identified in a study with T. cruzi [275] and AMXT 1501, which has been described as a potent inhibitor of polyamine and capsule biosynthesis in S. pneumoniae infections [274]. Polyamine biosynthesis and transport have been identified as drug targets in some bacteria as S. pneumoniae and Salmonella enterica serovar Typhimurium [276,277].
Remarkably, the development of drugs targeting ethanolamine metabolism is far less investigated. It has been reported that ethanolamine analogs with substitutions in the amino group of one of the methylene hydrogens of ethanolamine effectively inhibit ethanolamine transport in T. cruzi [278]. Although polyamine and ethanolamine utilization have been reported to be excellent drug targets in pathogenic bacteria, the development of inhibitor candidates still requires further study.
4. Conclusions
Mono- and polyamine metabolism has been investigated in a number of human pathogenic parasites and bacteria as a drug target. The knowledge of these metabolic networks is of medical importance, allowing the development of new drugs based on the validation of key enzymes involved in biosynthesis and utilization of these compounds. Multiple studies in pathogenic parasites including Leishmania sp., Trypanosoma sp., Toxoplasma sp., Trichomonas sp., Cryptosporidium sp., Crithidia sp., and Leptomonas sp. [279,280], and bacteria S. typhimurium, B. abortus, M. tuberculosis, Chlamydia pneumoniae, Legionella pneumophila, Listeria monocytogenes, and others [281] prove the crucial role of polyamines for their proliferation [282,283] leading to the validation of specific enzymes as drug targets. The interconnection between the mono-/polyamine biosynthesis, uptake, and assimilation remains crucial to find new drug targets. Since most human pathogens rely not only on polyamine biosynthesis, but also on polyamine detoxification with possible subsequent utilization or efflux in order to proliferate and maintain infection, targeting mono-/polyamine metabolism can extend the options for combating bacterial infections. Interestingly, targeting polyamine metabolism is currently also receiving much attention as a potential anti-cancer treatment [284,285], leading to the occurrence of validated inhibitors.
Investigations of mono- and polyamine metabolism in model bacteria including E. coli, S. typhimurium, P. aeruginosa, and S. coelicolor suggest new applications. For example, gamma-glutamylation pathways for polyamine utilization are rather of advantage for bacteria and involve specialized glutamylating enzymes for polyamine detoxification. Such enzymes have been discovered in pathogens P. aeruginosa [195] and S. coelicolor [113]. These gamma-glutamyl-polyamine/monoamine synthetases modify substrates by adding a glutamyl group in order to ensure the incorporation of these molecules into carbon and/or nitrogen metabolism [7]. Such glutamylation reaction is widespread in nature and might dramatically change chemical features of compounds through the change of stability in solution and by reduction of toxicity, for instance, of gamma-glutamypolyamines/monoamines [286]. Homologs of the glutamylation enzymes GlnA2, GlnA3, and GlnA4 from S. coelicolor have been found in other actinobacteria, including human pathogens belonging to Mycobacterium spp. and Rhodococcus spp. [7]. These enzymes are promising drug targets due to their key relevance in pathogenicity. Almost all actinobacteria, including M. tuberculosis causing tuberculosis infection, possess GlnA3 homologues as well as other homologues of polyamine uptake systems and enzymes from the predicted polyamine utilization pathway. Interestingly, the presence of GlnA3 (Rv1878) and some homologues involved in polyamine uptake (Rv1877) and utilization steps GabT (GABA transaminase, Rv2555) were reported in a guinea pig model of tuberculosis, where the bacterial proteome during early and chronic stages of this disease in vivo was investigated [287]. This report provided evidence that homologues involved in the polyamine utilization in S. coelicolor are necessary for M. tuberculosis surviving during tuberculosis infection.
Since there is an urgent need to find new anti-bacterial drugs with new modes of action that would be efficient on bacterial infections and shorten the treatment duration while avoiding relapses and the emergence of resistances as well as improving compliance, new strategies for the treatment of multidrug-resistant (MDR) infections are of particular concern. Whereas several antibiotics are effective in treating bacterial infections, these drugs target a small number of essential functions in the cell. Therefore, investigation of the pathways for mono-/polyamine metabolism that are required for bacterial growth, survival, and pathogenicity would provide new targets for the rational design of more effective agents that could be active against multidrug resistant strains.
Author Contributions
Conceptualization, S.K.; methodology, S.K.; formal analysis, S.K.; investigation, S.K.; resources, W.W.; data curation, S.K.; writing—original draft preparation, S.K.; writing—review and editing, S.K., W.W.; visualization, S.K.; supervision, S.K., W.W.; project administration, W.W.; funding acquisition, W.W. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the German Research Foundation (Deutsche Forschungsgemeinschaft, DFG)-funded Research Training Group GRK1708 (I and II) and the BMBF (Fördermaßnahme “Targetvalidierung für die pharmazeutische Wirkstoffentwicklung”) project GPS-TBT (FKZ: 16 GW0183K), as well as project GSS-TUBTAR (FKZ: 16 GW0253K). SK was a member of the DFG Research Training Group GRK1708 (I and II).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are involved in this study.
Acknowledgments
We thank all former coworkers and students in our department who contributed to the ongoing work on polyamines, in particular, Agnieszka Mitulski (formerly Bera), who initiated the analyses of polyamines, and Andreas Kulik for his permanent help in analytics.
Conflicts of Interest
Both authors declare no conflict of interest.
References
- Aharonowitz, Y. Nitrogen metabolite regulation of antibiotic biosynthesis. Annu. Rev. Microbiol. 1980, 34, 209–233. [Google Scholar] [PubMed]
- Arcondéguy, T.; Jack, R.; Merrick, M. PII signal transduction proteins, pivotal players in microbial nitrogen control. Microbiol. Mol. Biol. Rev. 2001, 65, 80–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, S. A Guide to the Polyamines; Oxford University Press: Oxford, UK, 1998; p. 610. [Google Scholar]
- Tyler, B. Regulation of the assimilation of nitrogen compounds. Annu. Rev. Biochem. 1978, 47, 1127–1162. [Google Scholar] [CrossRef] [PubMed]
- Magasanik, B. Genetic control of nitrogen assimilation in bacteria. Annu. Rev. Genet. 1982, 16, 135–168. [Google Scholar] [CrossRef]
- Merrick, M.J.; Edwards, R.A. Nitrogen control in bacteria. Microbiol. Rev. 1995, 59, 604–622. [Google Scholar] [CrossRef] [PubMed]
- Krysenko, S.; Matthews, A.; Busche, T.; Bera, A.; Wohlleben, W. Poly- and Monoamine Metabolism in Streptomyces coelicolor: The New Role of Glutamine Synthetase-Like Enzymes in the Survival under Environmental Stress. Microb. Physiol. 2021, 31, 233–247. [Google Scholar] [CrossRef]
- Sperber, A.M.; Herman, J.K. Metabolism Shapes the Cell. J. Bacteriol. 2017, 199, e00039-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, G.; Mullins, J.G.L.; Merrick, M. Membrane topology of the Mep/Amt family of ammonium transporters. Mol. Microbiol. 2000, 37, 331–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shapiro, S.; Vining, L.C. Suppression of nitrate utilization by ammonium and its relationship to chloramphenicol production in Streptomyces venezuelae. Can. J. Microbiol. 1984, 30, 798–804. [Google Scholar] [CrossRef] [PubMed]
- Oshima, T.; Soda, K. Biochemistry and biotechnology of amino acid dehydrogenases. Adv. Biochem. Eng. Biotechnol. 1990, 42, 187. [Google Scholar]
- Moir, J.W.B.; Wood, N.J. Nitrate and nitrite transport in bacteria. Cell. Mol. Life Sci. 2001, 58, 215–224. [Google Scholar] [CrossRef]
- Fischer, M.; Alderson, J.; van Keulen, G.; White, J.; Sawers, R.G. The obligate aerobe Streptomyces coelicolor A3(2) synthesizes three active respiratory nitrate reductases. Microbiology 2010, 156, 3166–3179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, H.; Kurihara, S. Polyamine Catabolism in Prokaryotes. In Polyamines: A Universal Molecular Nexus for Growth, Survival, and Specialized Metabolism; Kusano, T., Suzuki, H., Eds.; Springer: Tokyo, Japan, 2015; pp. 47–59. [Google Scholar]
- Yuan, J.; Doucette, C.D.; Fowler, W.U.; Feng, X.J.; Piazza, M.; Rabitz, H.A.; Wingreen, N.S.; Rabinowitz, J.D. Metabolomics-driven quantitative analysis of ammonia assimilation in E. coli. Mol. Syst. Biol. 2009, 5, 302. [Google Scholar] [CrossRef] [PubMed]
- Krysenko, S.; Wohlleben, W. Überlebenswichtig: Glutaminsynthetasehomologe Proteine in Streptomyceten. Biospektrum 2022, 28, 23–26. [Google Scholar] [CrossRef]
- Reitzer, L.J.; Magasanik, B. Ammonia assimilation and the biosynthesis of glutamine, glutamate, aspartate, asparagine, L-alanine, and D-alanine. In Escherichia coli and Salmonella typhimurium: Cellular and Molecular Biology, Volume 1; Neidhardt, F.C., Ingraham, J.L., Low, K.B., Magasanik, B., Schaechter, M., Umbarger, H.E., Eds.; American Society for Microbiology: Washington, DC, USA, 1987; pp. 302–320. [Google Scholar]
- Blauwkamp, T.A.; Ninfa, A.J. Physiological role of the GlnK signal transduction protein of Escherichia coli: Survival of nitrogen starvation. Mol. Microbiol. 2002, 46, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Ninfa, A.J.; Jiang, P.; Atkinson, M.R.; Peliska, J.A. Integration of antagonistic signals in the regulation of nitrogen assimilation in Escherichia coli. Curr. Top. Cell. Regul. 2001, 36, 31–75. [Google Scholar]
- He, L.; Soupene, E.; Kustu, S. NtrC is required for control of Klebsiella pneumoniae NifL activity. J. Bacteriol. 1997, 179, 7446–7455. [Google Scholar] [CrossRef] [Green Version]
- Shapiro, B.M.; Stadtman, E.R. Glutamine synthetase (Escherichia coli). Methods Enzymol. 1970, 17, 910–922. [Google Scholar]
- Stadtman, E.R.; Ginsburg, A. The glutamine synthetase of Escherichia coli: Structure and control. In The Enzymes; Boyer, P.D., Ed.; Academic Press: New York, NY, USA, 1974; Volume 10, pp. 755–807. [Google Scholar]
- Carroll, P.; Pashley, C.A.; Parish, T. Functional analysis of GlnE, an essential adenylyl transferase in Mycobacterium tuberculosis. J. Bacteriol. 2008, 190, 4894–4902. [Google Scholar] [CrossRef] [Green Version]
- Jiang, P.; Pioszak, A.A.; Ninfa, A.J. Structure/function analysis of glutamine synthetase adenylyltransferase (ATase, E.C. 2.7.7.49) of Escherichia coli. Biochemistry 2007, 46, 4117–4132. [Google Scholar] [CrossRef]
- Bolay, P.; Muro-Pastor, M.I.; Florencio, F.J.; Klähn, S. The Distinctive Regulation of Cyanobacterial Glutamine Synthetase. Life 2018, 8, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Dominguez, M.; Reyes, J.C.; Florencio, F.J. Glutamine synthetase inactivation by protein-protein interaction. Proc. Natl. Acad. Sci. USA 1999, 96, 7161–7166. [Google Scholar] [CrossRef] [Green Version]
- Forchhammer, K.; Selim, K.A. Carbon/nitrogen homeostasis control in cyanobacteria. FEMS Microbiol. Rev. 2020, 44, 33–53. [Google Scholar] [PubMed]
- Vazquez-Bermudez, M.F.; Herrero, A.; Flores, E. 2-Oxoglutarate increases the binding affinity of the NtcA (nitrogen control) transcription factor for the Synechococcus glnA promoter. FEBS Lett. 2002, 512, 71–74. [Google Scholar] [CrossRef] [Green Version]
- Jiang, P.; Peliska, J.A.; Ninfa, A.J. Enzymological characterization of the signal-transducing uridylyltransferase/uridylyl-removing enzyme (EC 2.7.7.59) of Escherichia coli and its interaction with the PII protein. Biochemistry 1998, 37, 12782–12794. [Google Scholar] [CrossRef] [PubMed]
- Muro-Pastor, M.I.; Florencio, F.J. Regulation of ammonium assimilation in cyanobacteria. Plant. Physiol. Biochem. 2003, 41, 595–603. [Google Scholar] [CrossRef]
- Atkinson, M.R.; Fisher, S.H. Identification of genes and gene products whose expression is activated during nitrogen-limited growth in Bacillus subtilis. J. Bacteriol. 1991, 173, 23–27. [Google Scholar] [CrossRef] [Green Version]
- Schreier, H.J. Biosynthesis of glutamine and glutamate and the assimilation of ammonia in Bacillus subtilis and other Gram-positive bacteria. In Bacillus subtilis and Other Gram-Positive Bacteria: Biochemistry, Physiology, and Molecular Genetics; Sonenshein, A.L., Hoch, J.A., Losick, R., Eds.; American Society for Microbiology: Washington, DC, USA, 1993; pp. 281–298. [Google Scholar]
- Picossi, S.; Belitsky, B.R.; Sonenshein, A.L. Molecular mechanism of the regulation of Bacillus subtilis gltAB expression by GltC. J. Mol. Biol. 2007, 365, 1298–1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, S.H. Regulation of nitrogen metabolism in Bacillus subtilis: Vive la difference! Mol. Microbiol. 1999, 32, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Brandenburg, J.L.; Wray, L.V., Jr.; Beier, L.; Jarmer, H.; Saxild, H.H.; Fisher, S.H. Roles of PucR, GlnR, and TnrA in regulating expression of the Bacillus subtilis ure P3 promoter. J. Bacteriol. 2002, 184, 6060–6064. [Google Scholar] [CrossRef] [Green Version]
- Kormelink, T.; Koenders, E.; Hagemeijer, Y.; Overmars, L.; Siezen, R.J.; de Vos, W.M.; Francke, C. Comparative genome analysis of central nitrogen metabolism and its control by GlnR in the class bacilli. BMC Genom. 2012, 13, 191. [Google Scholar] [CrossRef] [Green Version]
- Meier-Wagner, J.; Nolden, L.; Jakoby, M.; Siewe, R.; Krämer, R.; Burkovski, A. Multiplicity of ammonium uptake systems in Corynebacterium glutamicum: Role of Amt and AmtB. Microbiology 2001, 147, 135–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siewe, R.M.; Weil, B.; Burkovski, A.; Eikmanns, B.J.; Eikmanns, M.; Krämer, R. Functional and genetic characterization of the (methyl) ammonium uptake carrier of Corynebacterium glutamicum. J. Biol. Chem. 1996, 271, 5398–5403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walter, B.; Kuspert, M.; Ansorge, D.; Kramer, R.; Burkovski, A. Dissection of ammonium uptake systems in Corynebacterium glutamicum: Mechanism of action and energetics of AmtA and AmtB. J. Bacteriol. 2008, 190, 2611–2614. [Google Scholar] [CrossRef] [Green Version]
- Amon, J.; Titgemeyer, F.; Burkovski, A. Common patterns—Unique features: Nitrogen metabolism and regulation in Gram-positive bacteria. FEMS Microbiol. Rev. 2010, 34, 588–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beckers, G.; Strosser, J.; Hildebrandt, U.; Kalinowski, J.; Farwick, M.; Kramer, R.; Burkovski, A. Regulation of AmtR-controlled gene expression in Corynebacterium glutamicum: Mechanism and characterization of the AmtR regulon. Mol. Microbiol. 2005, 58, 580–595. [Google Scholar] [CrossRef]
- Jakoby, M.; Krämer, R.; Burkovski, A. Nitrogen regulation in Corynebacterium glutamicum: Isolation of genes involved and biochemical characterization of corresponding proteins. FEMS Microbiol. Lett. 1999, 173, 303–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nolden, L.; Beckers, G.; Mockel, B.; Pfefferle, W.; Nampoothiri, K.M.; Kramer, R.; Burkovski, A. Urease of Corynebacterium glutamicum: Organization of corresponding genes and investigation of activity. FEMS Microbiol. Lett. 2000, 189, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Burkovski, A. Ammonium assimilation and nitrogen control in Corynebacterium glutamicum and its relatives: An example for new regulatory mechanisms in actinomycetes. FEMS Microbiol. Rev. 2003, 27, 617–628. [Google Scholar] [CrossRef] [Green Version]
- Harper, C.; Hayward, D.; Wiid, I.; van Helden, P. Regulation of nitrogen metabolism in Mycobacterium tuberculosis: A comparison with mechanisms in Corynebacterium glutamicum and Streptomyces coelicolor. IUBMB Life 2008, 60, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Hopwood, D.A. Forty years of genetics with Streptomyces: From in vivo through in vitro to in silico. Microbiology 1999, 145, 2183–2202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchanan, R.E. Studies in the nomenclature and classification of the bacteria. II. The Primary Sub-divisions of the Schizomycetes. J. Bacteriol. 1917, 2, 155–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, F.; Bibb, M.J. Codon usage in the G+C-rich Streptomyces genome. Gene 1992, 113, 55–65. [Google Scholar] [CrossRef]
- Bentley, S.D.; Chater, K.F.; Cerdeño-Tárraga, A.M.; Challis, G.L.; Thomson, N.R.; James, K.D.; Harris, D.E.; Quail, M.A.; Kieser, H.; Harper, D.; et al. Complete genome sequence of the model actinomycete Streptomyces coelicolor A3(2). Nature 2002, 417, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Hopwood, D.A.; Chater, K.F.; Bibb, M.J. Genetics of antibiotic production in Streptomyces coelicolor A3(2), a model streptomycete. Biotechnology 1995, 28, 65–102. [Google Scholar] [PubMed]
- Hodgson, D.A. Primary metabolism and its control in Streptomycetes. Adv. Microbial. Physiol. 2000, 42, 47–238. [Google Scholar]
- McCarthy, A.J.; Williams, S.T. Actinomycetes as agents of biodegradation in the environment—A review. Gene 1992, 115, 189–192. [Google Scholar] [CrossRef]
- Hopwood, D.A. Soil to genomics: The Streptomyces chromosome. Annu. Rev. Genet. 2006, 40, 1–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karandikar, A.; Sharples, G.P.; Hobbs, G. Differentiation of Streptomyces coelicolor A3(2) under nitrate limited conditions. Microbiology 1997, 143, 3581–3590. [Google Scholar] [CrossRef] [Green Version]
- Wray, L.V., Jr.; Atkinson, M.R.; Fisher, S.H. Identification and cloning of the glnR locus, which is required for transcription of the glnA gene in Streptomyces coelicolor A3(2). J. Bacteriol. 1991, 173, 7351–7360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wray, L.V., Jr.; Fisher, S.H. The Streptomyces coelicolor glnR gene encodes a protein similar to other bacterial response regulators. Gene 1993, 130, 145–150. [Google Scholar] [CrossRef]
- Reuther, J.; Wohlleben, W. Nitrogen Metabolism in Streptomyces coelicolor: Transcriptional and Post-Translational Regulation. J. Mol. Microbiol. Biotechnol. 2007, 12, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Tiffert, Y.; Supra, P.; Wurm, R.; Wohlleben, W.; Wagner, R.; Reuther, J. The Streptomyces coelicolor GlnR regulon: Identification of new GlnR targets and evidence for a central role of GlnR in nitrogen metabolism in actinomycetes. Mol. Microbiol. 2008, 67, 861–880. [Google Scholar] [CrossRef] [PubMed]
- Tiffert, Y.; Franz-Wachtel, M.; Fladerer, C.; Nordheim, A.; Reuther, J.; Wohlleben, W.; Mast, Y. Proteomic analysis of the GlnR-mediated response to nitrogen limitation in Streptomyces coelicolor M145. Appl. Microbiol. Biotechnol. 2011, 89, 1149–1159. [Google Scholar] [CrossRef]
- Fink, D.; Falke, D.; Wohlleben, W.; Engels, A. Nitrogen metabolism in Streptomyces coelicolor A3(2): Modification of glutamine synthetase I by an adenylyltransferase. Microbiology 1999, 145, 2313–2322. [Google Scholar] [CrossRef] [PubMed]
- Fink, D.; Weissschuh, N.; Reuther, J.; Wohlleben, W.; Engels, A. Two transcriptional regulators GlnR and GlnRII are involved in regulation of nitrogen metabolism in Streptomyces coelicolor A3(2). Mol. Microbiol. 2002, 46, 331–347. [Google Scholar] [CrossRef]
- Pullan, S.T.; Chandra, G.; Bibb, M.J.; Merrick, M. Genomewide analysis of the role of GlnR in Streptomyces venezuelae provides new insights into global nitrogen regulation in actinomycetes. BMC Genom. 2011, 12, 175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amin, R.; Franz-Wachtel, M.; Tiffert, Y.; Heberer, M.; Meky, M.; Ahmed, Y.; Matthews, A.; Krysenko, S.; Jakobi, M.; Hinder, M.; et al. Post-translational Serine/Threonine Phosphorylation and Lysine Acetylation: A Novel Regulatory Aspect of the Global Nitrogen Response Regulator GlnR in S. coelicolor M145. Front. Mol. Biosci. 2016, 3, 38. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Zhao, G.-P. GlnR positively regulates nasA transcription in Streptomyces coelicolor. Biochem. Biophys. Res. Commun. 2009, 386, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Amin, R.; Reuther, J.; Bera, A.; Wohlleben, W.; Mast, Y. A novel GlnR target gene, nnaR, is involved in nitrate/nitrite assimilation in Streptomyces coelicolor. Microbiology 2012, 158, 1172–1182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weissschuh, N.; Fink, D.; Vierling, S.; Bibb, M.J.; Wohlleben, W.; Engels, A. Transcriptional analysis of the gene for glutamine synthetase II and two upstream genes in Streptomyces coelicolor A3(2). Mol. Gen. Genet. 2000, 264, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Hindra-Mulder, D.; Yin, C.; Elliot, M.A. Crp is a global regulator of antibiotic production in Streptomyces. mBio 2012, 3, e00407-12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-García, A.; Sola-Landa, A.; Apel, K.; Santos-Beneit, F.; Martín, J.F. Phosphate control over nitrogen metabolism in Streptomyces coelicolor: Direct and indirect negative control of glnR, glnA, glnII and amtB expression by the response regulator PhoP. Nucleic Acids Res. 2009, 37, 3230–3242. [Google Scholar] [CrossRef] [PubMed]
- Sola-Landa, A.; Rodríguez-García, A.; Amin, R.; Wohlleben, W.; Martín, J.F. Competition between the GlnR and PhoP regulators for the glnA and amtB promoters in Streptomyces coelicolor. Nucleic Acids Res. 2013, 41, 1767–1782. [Google Scholar] [CrossRef] [Green Version]
- Perez-Redondo, R.; Rodriguez-Garcia, A.; Botas, A.; Santamarta, I.; Martin, J.F.; Liras, P. ArgR of Streptomyces coelicolor is a versatile regulator. PLoS ONE 2012, 7, e32697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos-Beneit, F.; Rodriguez-Garcia, A.; Martin, J.F. Overlapping binding of PhoP and AfsR to the promoter region of glnR in Streptomyces coelicolor. Microbiol. Res. 2012, 167, 532–535. [Google Scholar] [CrossRef]
- Rigali, S.; Titgemeyer, F.; Barends, S.; Mulder, S.; Thomae, A.W.; Hopwood, D.A.; van Wezel, G.P. Feast or famine: The global regulator DasR links nutrient stress to antibiotic production by Streptomyces. EMBO Rep. 2008, 9, 670–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Wezel, G.P.; McDowall, K.J. The regulation of the secondary metabolism of Streptomyces: New links and experimental advances. Nat. Prod. Rep. 2011, 28, 1311–1333. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Mast, Y.; Wang, J.; Zhang, W.; Zhao, G.; Wohlleben, W.; Lu, Y.; Jiang, W. Identification of two-component system AfsQ1/Q2 regulon and its cross-regulation with GlnR in Streptomyces coelicolor. Mol. Microbiol. 2013, 87, 30–48. [Google Scholar] [CrossRef] [PubMed]
- Lu, F.; Hou, Y.; Zhang, H.; Chu, Y.; Xia, H.; Tian, Y. Regulatory genes and their roles for improvement of antibiotic biosynthesis in Streptomyces. Biotech 2017, 7, 250. [Google Scholar] [CrossRef] [PubMed]
- Waldvogel, E.; Herbig, A.; Battke, F.; Amin, R.; Nentwich, M.; Nieselt, K.; Ellingsen, T.E.; Wentzel, A.; Hodgson, D.A.; Wohlleben, W.; et al. The PII protein GlnK is a pleiotropic regulator for morphological differentiation and secondary metabolism in Streptomyces coelicolor. Appl. Microbiol. Biotechnol. 2011, 92, 1219–1236. [Google Scholar] [CrossRef] [PubMed]
- Wohlleben, W.; Bera, A.; Mast, Y.; Stegmann, E. Regulation of Secondary Metabolites of Actinobacteria. In Biology and Biotechnology of Actinobacteria; Wink, J., Mohammadipanah, F., Hamedi, J., Eds.; Springer International Publishing AG: Berlin/Heidelberg, Germany, 2017; p. 395. [Google Scholar]
- Hesketh, A.; Fink, D.; Gust, B.; Rexer, H.U.; Scheel, B.; Chater, K.; Wohlleben, W.; Engels, A. The GlnD and GlnK homologues of Streptomyces coelicolor A3(2) are functionally dissimilar to their nitrogen regulatory system counterparts from enteric bacteria. Mol. Microbiol. 2002, 46, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Tullius, M.V.; Harth, G.; Horwitz, M.A. Glutamine synthetase GlnA1 is essential for growth of Mycobacterium tuberculosis in human THP-1 macrophages and guinea pigs. Infect. Immun. 2003, 71, 3927–3936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agapova, A.; Serafini, A.; Petridis, M.; Hunt, D.M.; Garza-Garcia, A.; Sohaskey, C.D.; Sório de Carvalho, L.P. Flexible nitrogen utilisation by the metabolic generalist pathogen Mycobacterium tuberculosis. eLife 2019, 8, e41129. [Google Scholar] [CrossRef] [PubMed]
- Gouzy, A.; Poquet, Y.; Neyrolles, O. Nitrogen metabolism in Mycobacterium tuberculosis physiology and virulence. Nat. Rev. Microbiol. 2014, 12, 729–737. [Google Scholar] [CrossRef] [PubMed]
- Harth, G.; Maslesa-Galic, S.; Tullius, M.V.; Horwitz, M.A. All four Mycobacterium tuberculosis glnA genes encode glutamine synthetase activities but only GlnA1 is abundantly expressed and essential for bacterial homeostasis. Mol. Microbiol. 2005, 58, 1157–1172. [Google Scholar] [CrossRef]
- Jenkins, V.A.; Robertson, B.D.; Williams, K.J. Aspartate D48 is essential for the GlnR-mediated transcriptional response to nitrogen limitation in Mycobacterium smegmatis. FEMS Microbiol. Lett. 2012, 330, 38–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, K.J.; Jenkins, V.A.; Barton, G.R.; Bryant, W.A.; Krishnan, N.; Robertson, B.D. Deciphering the metabolic response of Mycobacterium tuberculosis to nitrogen stress. Mol. Microbiol. 2015, 97, 1142–1157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pesole, G.; Gissi, C.; Lanave, C.; Saccone, C. Glutamine synthetase gene evolution in bacteria. Mol. Biol. Evol. 1995, 12, 189–197. [Google Scholar] [PubMed] [Green Version]
- Reitzer, L.; Schneider, B.L. Metabolic context and possible physiological themes of σ54-dependent genes in Escherichia coli. Microbiol. Mol. Biol. Rev. 2001, 65, 422–444. [Google Scholar] [CrossRef] [Green Version]
- Feng, H.; Sun, Y.; Zhi, Y.; Wei, X.; Luo, Y.; Mao, L.; Zhou, P. Identification and characterization of the nitrate assimilation genes in the isolate of Streptomyces griseorubens JSD-1. Microb. Cell Factories 2014, 13, 174. [Google Scholar] [CrossRef] [PubMed]
- Kumada, Y.; Benson, D.R.; Hillemann, D.; Hosted, T.J.; Rochefort, D.A.; Thompson, C.J.; Wohlleben, W.; Tateno, Y. Evolution of the glutamine synthetase gene, one of the oldest existing and functioning genes. Proc. Natl. Acad. Sci. USA 1993, 90, 3009–3013. [Google Scholar] [CrossRef] [Green Version]
- Mathis, R.; Gamas, P.; Meyer, Y.; Cullimore, J.V. The presence of GSI-like genes in higher plants: Support for the paralogous evolution of GSI and GSII genes. J. Mol. Evol. 2000, 50, 116–122. [Google Scholar] [CrossRef]
- Wyatt, K.; White, H.E.; Wang, L.; Bateman, O.A.; Slingsby, C.; Orlova, E.V.; Wistow, G. Lengsin is a survivor of an ancient family of class I glutamine synthetases re-engineered by evolution for a role in the vertebrate lens. Structure 2006, 14, 1823–1834. [Google Scholar] [CrossRef] [Green Version]
- Behrmann, I.; Hillemann, D.; Pühler, A.; Strauch, E.; Wohlleben, W. Overexpression of a Streptomyces viridochromogenes gene (glnII) encoding a glutamine synthetase similar to those of eucaryotes confers resistance against the antibiotic phosphinothricyl-alanyl-alanine. J. Bacteriol. 1990, 172, 5326–5334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, S.L.; Young, J.P. The glutamine synthetases of rhizobia: Phylogenetics and evolutionary implications. Mol. Biol. Evol. 2000, 17, 309–319. [Google Scholar] [PubMed] [Green Version]
- Reyes, J.C.; Florencio, F.J. A mutant lacking the glutamine synthetase gene (glnA) is impaired in the regulation of the nitrate assimilation system in the cyanobacterium Synechocystis sp. strain PCC 6803. J. Bacteriol. 1994, 176, 7516–7523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinoshita, S.; Isu, S.; Kaneko, G.; Yamada, H.; Hara, T.; Itoh, Y.; Watabe, S. The occurrence of eukaryotic type III glutamine synthetase in the marine diatom Chaetoceros compressum. Mar. Genom. 2009, 2, 103–111. [Google Scholar] [CrossRef]
- Chiurazzi, M.; Meza, R.; Lara, M.; Lahm, A.; Defez, R.; Iaccarino, M.; Espín, G. The Rhizobium leguminosarum biovar phaseoli glnT gene, encoding glutamine synthetase III. Gene 1992, 119, 1–8. [Google Scholar] [CrossRef]
- Shatters, R.G.; Liu, Y.; Kahn, M.L. Isolation and characterization of a novel glutamine synthetase from Rhizobium meliloti. J. Biol. Chem. 1993, 268, 469–475. [Google Scholar] [CrossRef]
- Rossbach, S.; Schell, J.; Bruijn, F.J. Cloning and analysis of Agrobacterium tumefaciens C58 loci involved in glutamine biosynthesis: Neither the glnA (GSI) nor the glnII (GSII) gene plays a special role in virulence. Mol. Gen. Genet. 1988, 212, 38–47. [Google Scholar] [CrossRef]
- Eisenberg, D.; Gill, H.S.; Pfluegl, G.M.; Rotstein, S.H. Structure-function relationships of glutamine synthetases. Biochim. Biophys. Acta 2000, 1477, 122–145. [Google Scholar] [CrossRef] [Green Version]
- Robertson, D.L.; Alberte, R.S. Isolation and characterization of glutamine synthetase from the marine diatom Skeletonema costatum. Plant. Physiol. 1996, 111, 1169–1175. [Google Scholar] [CrossRef] [Green Version]
- Unno, H.; Uchida, T.; Sugawara, H.; Kurisu, G.; Sugiyama, T.; Yamaya, T.; Sakakibara, H.; Hase, T.; Kusunoki, M. Atomic structure of plant glutamine synthetase: A key enzyme for plant productivity. J. Biol. Chem. 2006, 281, 29287–29296. [Google Scholar] [CrossRef] [Green Version]
- Seabra, A.R.; Carvalho, H.; Pereira, P.J. Crystallization and preliminary crystallographic characterization of glutamine synthetase from Medicago truncatula. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2009, 65, 1309–1312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krajewski, W.W.; Collins, R.; Holmberg-Schiavone, L.; Jones, T.A.; Karlberg, T.; Mowbray, S.L. Crystal structures of mammalian glutamine synthetases illustrate substrate-induced conformational changes and provide opportunities for drug and herbicide design. J. Mol. Biol. 2008, 375, 217. [Google Scholar] [CrossRef] [Green Version]
- van Rooyen, J.M.; Abratt, V.R.; Belrhali, H.; Sewell, T. Crystal structure of Type III glutamine synthetase: Surprising reversal of the inter-ring interface. Structure 2011, 19, 471–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liaw, S.H.; Pan, C.; Eisenberg, D. Feedback inhibition of fully unadenylylated glutamine synthetase from Salmonella typhimurium by glycine, alanine, and serine. Proc. Natl. Acad. Sci. USA 1993, 90, 4996–5000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gill, H.S.; Eisenberg, D. The crystal structure of phosphinothricin in the active site of glutamine synthetase illuminates the mechanism of enzymatic inhibition. Biochemistry 2001, 40, 1903–1912. [Google Scholar] [CrossRef] [PubMed]
- Joo, H.K.; Park, Y.W.; Jang, Y.Y.; Lee, J.Y. Structural Analysis of Glutamine Synthetase from Helicobacter pylori. Sci. Rep. 2018, 8, 11657. [Google Scholar] [CrossRef] [Green Version]
- Murray, D.S.; Chinnam, N.; Tonthat, N.K.; Whitfill, T.; Wray, L.V., Jr.; Fisher, S.H.; Schumacher, M.A. Structures of the Bacillus subtilis glutamine synthetase dodecamer reveal large intersubunit catalytic conformational changes linked to a unique feedback inhibition mechanism. J. Biol. Chem. 2013, 288, 35801–35811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gill, H.S.; Pfluegl, G.M.; Eisenberg, D. Multicopy crystallographic refinement of a relaxed glutamine synthetase from Mycobacterium tuberculosis highlights flexible loops in the enzymatic mechanism and its regulation. Biochemistry 2002, 41, 9863–9872. [Google Scholar] [CrossRef] [PubMed]
- Wray, L.V.; Fisher, S.H. Cloning and nucleotide sequence of the Streptomyces coelicolor gene encoding glutamine synthetase. Gene 1988, 71, 247–256. [Google Scholar] [CrossRef]
- Hillemann, D.; Dammann, T.; Hillemann, A.; Wohlleben, W. Genetic and biochemical characterization of the two glutamine synthetases GSI and GSII of the phosphinothricyl-alanyl-alanine producer, Streptomyces viridochromeogenes Tu494. J. Gen. Microbiol. 1993, 139, 1773–1783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayward, D.; van Helden, P.D.; Wiid, I.J. Glutamine synthetase sequence evolution in the mycobacteria and their use as molecular markers for Actinobacteria speciation. BMC Evol. Biol. 2009, 9, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rexer, H.U.; Schaberle, T.; Wohlleben, W.; Engels, A. Investigation of the functional properties and regulation of three glutamine synthetase like genes in Streptomyces coelicolor A3(2). Arch. Microbiol. 2006, 186, 447–458. [Google Scholar] [CrossRef] [PubMed]
- Krysenko, S.; Okoniewski, N.; Kulik, A.; Matthews, A.; Grimpo, J.; Wohlleben, W.; Bera, A. Gamma-Glutamylpolyamine Synthetase GlnA3 is involved in the first step of polyamine degradation pathway in Streptomyces coelicolor M145. Front. Microbiol. 2017, 8, 726. [Google Scholar] [CrossRef] [Green Version]
- Krysenko, S.; Matthews, A.; Okoniewski, N.; Kulik, A.; Girbas, M.G.; Tsypik, O.; Meyners, C.S.; Hausch, F.; Wohlleben, W.; Bera, A. Initial metabolic step of a novel ethanolamine utilization pathway and its regulation in Streptomyces coelicolor M145. mBio 2019, 10, e00326-19. [Google Scholar] [CrossRef] [Green Version]
- Krysenko, S.; Okoniewski, N.; Nentwich, M.; Matthews, A.; Bäuerle, M.; Zinser, A.; Busche, T.; Kulik, A.; Gursch, S.; Kemeny, A.; et al. A Second Gamma-Glutamylpolyamine Synthetase, GlnA2, Is Involved in Polyamine Catabolism in Streptomyces coelicolor. Int. J. Mol. Sci. 2022, 23, 3752. [Google Scholar] [CrossRef] [PubMed]
- Yatin, M. Polyamines in living organisms. Mol. Cell. Biol. 2002, 1, 57–67. [Google Scholar]
- Miller-Fleming, L.; Olin-Sandoval, V.; Campbell, K.; Ralser, M. Remaining Mysteries of Molecular Biology: The Role of Polyamines in the Cell. J. Mol. Biol. 2015, 427, 3389–3406. [Google Scholar] [CrossRef] [PubMed]
- Michael, A.J. Polyamine function in archaea and bacteria. J. Biol. Chem. 2018, 293, 18693–18701. [Google Scholar] [CrossRef] [Green Version]
- Oshima, T.; Hamasaki, N.; Uzawa, T. Biochemical Properties of Unusual Polyamines Found in an Extreme Thermophile, Thermus thermophiles. In Progress in Polyamine Research; Zappia, V., Pegg, A.E., Eds.; Springer: Boston, MA, USA, 1988; pp. 633–642. [Google Scholar]
- Hamana, K.; Matsuzaki, S. Distribution of polyamines in actinomycetes. FEMS Microbiol. Lett. 1987, 41, 1574–6968. [Google Scholar] [CrossRef]
- Fukuda, W.; Yamori, Y.; Hamakawa, M.; Osaki, M.; Fukuda, M.; Hidese, R.; Kanesaki, Y.; Okamoto-Kainuma, A.; Kato, S.; Fujiwara, S. Genes regulated by branched-chain polyamine in the hyperthermophilic archaeon Thermococcus kodakarensis. Amino Acids 2020, 52, 287–299. [Google Scholar] [CrossRef] [PubMed]
- Young, C.C.; Chen, L.F. Polyamines in humic acid and their effect on radical growth of lettuce seedlings. Plant Soil 1997, 195, 143–149. [Google Scholar] [CrossRef]
- Lasbury, M.E.; Merali, S.; Durant, P.J.; Tschang, D.; Ray, C.A.; Lee, C.H. Polyamine-mediated apoptosis of alveolar macrophages during Pneumocystis pneumonia. J. Biol. Chem. 2007, 282, 11009–11020. [Google Scholar] [CrossRef] [Green Version]
- Broshtilova, V.; Lozanov, V.; Miteva, L. Comparative analysis of polyamine metabolism in benign and neoplastic keratinocytic proliferations. Acta Derm. Venereol. 2012, 21, 3–5. [Google Scholar]
- Tabor, C.W.; Tabor, H. Polyamines in microorganisms. Microbiol. Rev. 1985, 49, 81–99. [Google Scholar] [CrossRef]
- Grasemann, H.; Shehnaz, D.; Enomoto, M.; Leadley, M.; Belik, J.; Ratjen, F. L-Ornithine Derived Polyamines in Cystic Fibrosis Airways. PLoS ONE 2012, 7, e46618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramakrishna, S.; Guarino, L.; Cohen, S.S. Polyamines of Anacystis nidulans and metabolism of exogenous spermidine and spermine. J. Bacteriol. 1978, 134, 744–750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakamoto, A.; Sahara, J.; Kawai, G.; Yamamoto, K.; Ishihama, A.; Uemura, T.; Igarashi, K.; Kashiwagi, K.; Terui, Y. Cytotoxic Mechanism of Excess Polyamines Functions through Translational Repression of Specific Proteins Encoded by Polyamine Modulon. Int. J. Mol. Sci. 2020, 21, 2406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Igarashi, K.; Kashiwagi, K. Effects of polyamines on protein synthesis and growth of Escherichia coli. J. Biol. Chem. 2018, 293, 18702–18709. [Google Scholar] [CrossRef] [Green Version]
- Hanfrey, C.; Sommer, S.; Mayer, M.J.; Burtin, D.; Michael, A.J. Arabidopsis polyamine biosynthesis: Absence of ornithine decarboxylase and the mechanism of arginine decarboxylase activity. Plant J. 2001, 27, 551–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakada, Y.; Itoh, Y. Identification of the putrescine biosynthetic genes in Pseudomonas aeruginosa and characterization of agmatine deiminase and N-carbamoylputrescine amidohydrolase of the arginine decarboxylase pathway. Microbiology 2003, 149, 707–714. [Google Scholar] [CrossRef] [PubMed]
- Dubin, D.T.; Rosenthal, S.M. The acetylation of polyamines in Escherichia coli. J. Biol. Chem. 1960, 235, 776–782. [Google Scholar] [CrossRef]
- Shah, P.; Swiatlo, E. Immunization with polyamine transport protein PotD protects mice against systemic infection with Streptococcus pneumoniae. Infect. Immun. 2006, 74, 5888–5892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhee, H.J.; Kim, E.J.; Lee, J.K. Physiological polyamines: Simple primordial stress molecules. J. Cell. Mol. Med. 2007, 11, 685–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burrell, M.; Hanfrey, C.C.; Murray, E.J.; Stanley-Wall, N.R.; Michael, A.J. Evolution and multiplicity of arginine decarboxylases in polyamine biosynthesis and essential role in Bacillus subtilis biofilm formation. J. Biol. Chem. 2010, 285, 39224–39238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burrell, M.; Hanfrey, C.C.; Kinch, L.N.; Elliot, K.A.; Michael, J.A. Evolution of a novel lysine decarboxylase in siderophore biosynthesis. Mol. Microbiol. 2012, 86, 485–499. [Google Scholar] [CrossRef] [PubMed]
- Patel, C.N.; Wortham, B.W.; Lines, J.L.; Fetherston, J.D.; Perry, R.D.; Oliveira, M.A. Polyamines are essential for the formation of plague biofilm. J. Bacteriol. 2006, 188, 2355–2363. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Sperandio, V.; Frantz, D.E.; Longgood, J.; Camilli, A.; Phillips, M.A.; Michael, A.J. An alternative polyamine biosynthetic pathway is widespread in bacteria and essential for biofilm formation in Vibrio cholerae. J. Biol. Chem. 2009, 284, 9899–9907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, R.; Hanfrey, C.C.; Elliott, K.A.; McCloskey, D.E.; Wang, X.; Kanugula, S.; Pegg, A.E.; Michael, A.J. Independent evolutionary origins of functional polyamine biosynthetic enzyme fusions catalysing de novo diamine to triamine formation. Mol. Microbiol. 2011, 81, 1109–1124. [Google Scholar] [CrossRef] [Green Version]
- Chattopadhyay, M.K.; Tabor, C.W.; Tabor, H. Polyamines are not required for aerobic growth of Escherichia coli: Preparation of a strain with deletions in all of the genes for polyamine biosynthesis. J. Bacteriol. 2009, 191, 5549–5552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain ATyagi, A.K. Role of polyamines in the synthesis of RNA in mycobacteria. Mol. Cell. Biochem. 1987, 78, 3–8. [Google Scholar]
- Huang, S.C.; Panagiotidis, C.A.; Canellakis, E.S. Transcriptional effects of polyamines on ribosomal proteins and on polyamine-synthesizing enzymes in Escherichia coli. Proc. Natl. Acad. Sci. USA 1990, 87, 3464–3468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarkar, N.K.; Shankar, S.; Tyagi, A.K. Polyamines exert regulatory control on mycobacterial transcription: A study using RNA polymerase from Mycobacterium phlei. Biochem. Mol. Biol. Int. 1995, 35, 1189–1198. [Google Scholar]
- Algranati, I.D.; Echandi, G.; Garcia-Patrone, M.; Gonzalez, N.S.; Goldemberg, S.H. Polyamines, equilibrium between ribosomal particles and protein synthesis in bacteria. Arch. Biol. Med. Exp. 1976, 10, 49–60. [Google Scholar] [PubMed]
- Nastri, H.G.; Fastame, I.G.; Algranati, I.D. Polyamines modulatestreptomycin-induced mistranslation in Escherichia coli. Biochim. Biophys. Acta 1993, 1216, 455–459. [Google Scholar] [CrossRef]
- Higashi, K.; Kashiwagi, K.; Taniguchi, S.; Terui, Y.; Yamamoto, K.; Ishihama, A.; Igarashi, K. Enhancement of C1 frameshift by polyamines during translation of polypeptide release factor 2 in Escherichia coli. J. Biol. Chem. 2006, 281, 9527–9537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terui, Y.; Akiyama, M.; Sakamoto, A.; Tomitori, H.; Yamamoto, K.; Ishihama, A.; Igarashi, K.; Kashiwagi, K. Increase in cell viability by polyamines through stimulation of the synthesis of ppGpp regulatory protein and omega protein of RNA polymerase in Escherichia coli. Int. J. Biochem. Cell. Biol. 2012, 44, 412–422. [Google Scholar] [CrossRef]
- Griffiths, G.L.; Sigel, S.P.; Payne, S.M.; Neilands, J.B. Vibriobactin, a siderophore from Vibrio cholerae. J. Biol. Chem. 1984, 259, 383–385. [Google Scholar] [CrossRef]
- Brickman, T.J.; Armstrong, S.K. The ornithine decarboxylase gene odc is required for alcaligin siderophore biosynthesis in Bordetella spp.: Putrescine is a precursor of alcaligin. J. Bacteriol. 1996, 178, 54–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oves-Costales, D.; Kadi, N.; Fogg, M.J.; Song, L.; Wilson, K.S.; Challis, G.L. Petrobactin biosynthesis: AsbB catalyzes condensation of spermidine with N8-citryl-spermidine and its N1-(3,4-dihydroxybenzoyl) derivative. Chem. Commun. 2008, 34, 4034–4036. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, M.; Kashiwagi, K.; Shigemasa, A.; Taniguchi, S.; Yamamoto, K.; Makinoshima, H.; Ishihama, A.; Igarashi, K. A unifying model for the role of polyamines in bacterial cell growth, the polyamine modulon. J. Biol. Chem. 2004, 279, 46008–46013. [Google Scholar] [CrossRef] [Green Version]
- Igarashi, K.; Kashiwagi, K. Polyamine Modulon in Escherichia coli: Genes involved in the stimulation of cell growth by polyamines. J. Biochem. 2006, 139, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Wortham, B.W.; Patel, C.N.; Oliveira, M.A. Polyamines in bacteria: Pleiotropic effects yet specific mechanisms. Adv. Exp. Med. Biol. 2007, 603, 106–115. [Google Scholar]
- Nesse, L.L.; Berg, K.; Vestby, L.K. Effects of norspermidine and spermidine on biofilm formation by potentially pathogenic Escherichia coli and Salmonella enterica wild-type strains. Appl. Environ. Microbiol. 2015, 81, 2226–2232. [Google Scholar] [CrossRef] [Green Version]
- Jung, I.L.; Kim, I.G. Transcription of ahpC, katG, and katE genes in Escherichia coli is regulated by polyamines: Polyamine-deficient mutant sensitive to H2O2-induced oxidative damage. Biochem. Biophys. Res. Commun. 2003, 301, 915–922. [Google Scholar] [CrossRef]
- Tkachenko, A.G.; Nesterova, L.Y. Polyamines as modulators of gene expression under oxidative stress in Escherichia coli. Biochemistry 2003, 68, 850–856. [Google Scholar]
- Oh, T.J.; Kim, I.G. The expression of Escherichia coli SOS genes recA and uvrA is inducible by polyamines. Biochem. Biophys. Res. Commun. 1999, 264, 584–589. [Google Scholar] [CrossRef]
- Chattopadhyay, M.K.; Tabor, H. Polyamines are critical for the induction of the glutamate decarboxylase-dependent acid resistance system in Escherichia coli. J. Biol. Chem. 2013, 288, 33559–33570. [Google Scholar] [CrossRef] [Green Version]
- Schneider, B.L.; Ruback, S.; Kiupakis, A.K.; Kasbarian, H.; Pybus, C.; Reitzer, L. The Escherichia coli gabDTPC operon: Specific γ-aminobutyrate catabolism and nonspecific induction. J. Bacteriol. 2002, 184, 6976–6986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nastri, H.G.; Algranati, I.D. Effect of polyamines on plasmid-mediated kanamycin resistance and kanamycin phosphotransferase gene expression in Escherichia coli. Cell. Mol. Biol. 1996, 42, 711–717. [Google Scholar] [PubMed]
- Kwon, D.H.; Lu, C.D. Polyamines induce resistance to cationic peptide, aminoglycoside, and quinolone antibiotics in Pseudomonas aeruginosa PAO1. Antimicrob. Agents Chemother. 2006, 50, 1615–1622. [Google Scholar] [CrossRef] [Green Version]
- Tkachenko, A.G.; Pozhidaeva, O.N.; Shumkov, M.S. Role of polyamines in formation of multiple antibiotic resistance of Escherichia coli under stress conditions. Biochemistry 2006, 71, 1042–1049. [Google Scholar] [CrossRef]
- Sarathy, J.P.; Lee, E.; Dartois, V. Polyamines inhibit porin-mediated fluoroquinolone uptake in mycobacteria. PLoS ONE 2013, 8, e65806. [Google Scholar] [CrossRef]
- Van den Bossche, J.; Bogaert, P.; van Hengel, J.; Guérin, C.J.; Berx, G.; Movahedi, K.; Van den Bergh, R.; Pereira-Fernandes, A.; Geuns, J.M.; Pircher, H.; et al. Alternatively activated macrophages engage in homotypic and heterotypic interactions through IL-4 and polyamine-induced E-cadherin/catenin complexes. Blood 2009, 114, 4664–4674. [Google Scholar] [CrossRef] [PubMed]
- Muraille, E.; Leo, O.; Moser, M. TH1/TH2 paradigm extended: Macrophage polarization as an unappreciated pathogen-driven escape mechanism? Front. Immunol. 2014, 5, 603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hesse, M.; Modolell, M.; La Flamme, A.C.; Schito, M.; Fuentes, J.M.; Cheever, A.W.; Pearce, E.J.; Wynn, T.A. Differential regulation of nitric oxide synthase-2 and arginase-1 by type 1/type 2 cytokines in vivo: Granulomatous pathology is shaped by the pattern of L-arginine metabolism. J. Immunol. 2001, 167, 6533–6544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cano, V.; March, C.; Insua, J.L.; Aguiló, N.; Llobet, E.; Moranta, D.; Regueiro, V.; Brennan, G.P.; Millán-Lou, M.I.; Martín, C.; et al. Klebsiella pneumoniae survives within macrophages by avoiding delivery to lysosomes. Cell Microbiol. 2015, 17, 1537–1560. [Google Scholar]
- Monack, D.M.; Bouley, D.M.; Falkow, S. Salmonella typhimurium persists within macrophages in the mesenteric lymph nodes of chronically infected Nramp1+/+ mice and can be reactivated by IFNgamma neutralization. Exp. Med. 2004, 199, 231–241. [Google Scholar] [CrossRef]
- Kerrinnes, T.; Winter, M.G.; Young, B.M.; Diaz-Ochoa, V.E.; Winter, S.E.; Tsolis, R.M. Utilization of host polyamines in alternatively activated macrophages promotes chronic infection by Brucella abortus. Infect. Immun. 2018, 86, e00458-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sycz, G.; Di Venanzio, G.; Distel, J.S.; Sartorio, M.G.; Le, N.H.; Scott, N.E.; Beatty, W.L.; Feldman, M.F. Modern Acinetobacter baumannii clinical isolates replicate inside spacious vacuoles and egress from macrophages. PLoS Pathog. 2021, 17, e1009802. [Google Scholar] [CrossRef] [PubMed]
- Johnson, B.K.; Thomas, S.M.; Olive, A.J.; Abramovitch, R.B. Macrophage Infection Models for Mycobacterium tuberculosis. In Methods in Molecular Biology; Clifton, N.J., Ed.; Humana Press: Totowa, NJ, USA, 2021; pp. 167–182. [Google Scholar]
- Gevrekci, A.Ö. The roles of polyamines in microorganisms. World. J. Microbiol. Biotechnol. 2017, 33, 204. [Google Scholar] [CrossRef] [PubMed]
- Hamana, K.; Tanaka, T.; Hosoya, R.; Niitsu, M.; Itoh, T. Cellular polyamines of the acidophilic, thermophilic and thermoacidophilic archaebacteria, Acidilobus, Ferroplasma, Pyrobaculum, Pyrococcus, Staphylothermus, Thermococcus, Thermodiscus and Vulcanisaeta. J. Gen. Appl. Microbiol. 2003, 49, 287–293. [Google Scholar] [CrossRef] [Green Version]
- Kusano, T.; Berberich, T.; Tateda, C.; Takahashi, Y. Polyamines: Essential factors for growth and survival. Planta 2008, 228, 367–381. [Google Scholar] [CrossRef] [PubMed]
- Nakada, Y.; Itoh, Y. Characterization and regulation of the gbuA gene, encoding guanidinobutyrase in the arginine dehydrogenase pathway of Pseudomonas aeruginosa PAO1. J. Bacteriol. 2002, 184, 3377–3384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wargnies, B.; Lauwers, N.; Stalon, V. Structure and properties of the putrescine carbamoyltransferase of Streptococcus faecalis. Eur. J. Biochem. 1979, 101, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Potter, A.J.; Paton, J.C. Spermidine biosynthesis and transport modulate pneumococcal autolysis. J. Bacteriol. 2014, 196, 3556–3561. [Google Scholar] [CrossRef] [Green Version]
- Tomar, P.C.; Lakra, N.; Mishra, S.N. Cadaverine: A lysine catabolite involved in plant growth and development. Plant Signal. Behav. 2013, 8, e25850. [Google Scholar] [CrossRef] [Green Version]
- Schneider, J.; Wendisch, V.F. Biotechnological production of polyamines by Bacteria: Recent achievements and future perspectives. Appl. Microbiol. Biotechnol. 2011, 91, 17–30. [Google Scholar] [CrossRef] [PubMed]
- Kurihara, S.; Oda, S.; Kato, K.; Kim, H.G.; Koyanagi, T.; Kumagai, H.; Suzuki, H. A novel putrescine utilization pathway involves γ-glutamylated intermediates of Escherichia coli K-12. J. Biol. Chem. 2005, 280, 4602–4608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurihara, S.; Tsuboi, Y.; Oda, S.; Kim, H.G.; Kumagai, H.; Suzuki, H. The putrescine importer PuuP of Escherichia coli K-12. J. Bacteriol. 2009, 191, 2776–2782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kashiwagi, K.; Miyamoto, S.; Suzuki, F.; Kobayashi, H.; Igarashi, K. Excretion of putrescine by the putrescine-ornithine antiporter encoded by the potE gene of Escherichia coli. Proc. Natl. Acad. Sci. USA 1992, 89, 4529–4533. [Google Scholar] [CrossRef] [Green Version]
- Pistocchi, R.; Kashiwagi, K.; Miyamoto, S.; Nukui, E.; Sadakata, Y.; Kobayashi, H.; Igarashi, K. Characteristics of the operon for a putrescine transport system that maps at 19 min on the Escherichia coli chromosome. J. Biol. Chem. 1993, 268, 146–152. [Google Scholar] [CrossRef]
- Kurihara, S.; Suzuki, H.; Oshida, M.; Benno, Y. A novel putrescine importer required for type pili-driven surface motility induced by extracellular putrescine in Escherichia coli K-12. J. Biol. Chem. 2011, 286, 10185–10192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soksawatmaekhin, W.; Kuraishi, A.; Sakata, K.; Kashiwagi, K.; Igarashi, K. Excretion and uptake of cadaverine by CadB and its physiological functions in Escherichia coli. Mol. Microbiol. 2004, 51, 1401–1412. [Google Scholar] [CrossRef]
- Furuchi, T.; Kashiwagi, K.; Kobayashi, H.; Igarashi, K. Characteristics of the gene for a spermidine and putrescine transport-system that maps at 15-min on the Escherichia coli chromosome. J. Biol. Chem. 1991, 266, 20928–20933. [Google Scholar] [CrossRef]
- Igarashi, K.; Kashiwagi, K. Polyamine transport in bacteria and yeast. Biochem. J. 1999, 344, 633–642. [Google Scholar] [CrossRef] [PubMed]
- Higashi, K.; Ishigure, H.; Demizu, R.; Uemura, T.; Nishino, K.; Yamaguchi, A.; Kashiwagi, K.; Igarashi, K. Identification of a spermidine excretion protein complex (MdtJI) in Escherichia coli. J. Bacteriol. 2008, 190, 872–878. [Google Scholar] [CrossRef] [Green Version]
- Karatan, E.; Duncan, T.R.; Watnick, P.I. NspS, a predicted polyamine sensor, mediates activation of Vibrio cholerae biofilm formation by norspermidine. J. Bacteriol. 2005, 187, 7434–7443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGinnis, M.W.; Parker, Z.M.; Walter, N.E.; Rutkovsky, A.C.; Cartaya-Marin, C.; Karatan, E. Spermidine regulates Vibrio cholerae biofilm formation via transport and signaling pathways. FEMS Microbiol. Lett. 2009, 299, 166–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zijnge, V.; Kieselbach, T.; Oscarsson, J. Proteomics of protein secretion by Aggregatibacter actinomycetemcomitans. PLoS ONE 2012, 7, e41662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, P.; Swiatlo, E. A multifaceted role for polyamines in bacterial pathogens. Mol. Microbiol. 2008, 68, 4–16. [Google Scholar] [CrossRef] [PubMed]
- Kurihara, S.; Sakai, Y.; Suzuki, H.; Muth, A.; Phanstiel, O., IV; Rather, P.N. Putrescine importer PlaP contributes to swarming motility and urothelial cell invasion in Proteus mirabilis. J. Biol. Chem. 2013, 288, 15668–15676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Maezato, Y.; Kim, S.H.; Kurihara, S.; Liang, J.; Michael, A.J. Polyamine-independent growth and biofilm formation, and functional spermidine/spermine N-acetyltransferases in Staphylococcus aureus and Enterococcus faecalis. Mol. Microbiol. 2019, 111, 159–175. [Google Scholar] [CrossRef] [Green Version]
- Yao, X.; He, W.; Lu, C.-D. Functional characterization of seven Glutamylpolyamine synthetase genes and the bauRABCD locus for polyamine and Alanine utilization in Pseudomonas aeruginosa PAO1. J. Bacteriol. 2011, 193, 3923–3930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, X.; Li, C.; Zhang, J.; Lu, C.-D. γ-glutamyl Spermine Synthetase PauA2 as a potential target of antibiotic development against Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2012, 56, 5309–5314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joshi, G.S.; Spontak, J.S.; Klapper, D.G.; Richardson, A.R. Arginine catabolic mobile element encoded speG abrogates the unique hyper-sensitivity of Staphylococcus aureus to exogenous polyamines. Mol. Microbiol. 2011, 82, 9–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, A.Q.; Schneider, J.; Wendisch, V.F. Elimination of polyamine N-acetylation and regulatory engineering improved putrescine production by Corynebacterium glutamicum. J. Biotechnol. 2015, 201, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Kurihara, S.; Oda, S.; Kumagai, H.; Suzuki, H. γ-Glutamyl-γ-aminobutyrate hydrolase in the putrescine utilization pathway of Escherichia coli K-12. FEMS Microbiol. Lett. 2006, 256, 318–323. [Google Scholar] [CrossRef] [PubMed]
- Kurihara, S.; Oda, S.; Tsuboi, Y.; Kim, H.G.; Oshida, M.; Kumagai, H.; Suzuki, H. γ-Glutamylputrescine synthetase in the putrescine utilization pathway of Escherichia coli K-12. J. Biol. Chem. 2008, 283, 19981–19990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, B.L.; Reitzer, L. Pathway and enzyme redundancy in putrescine catabolism in Escherichia coli. J. Bacteriol. 2012, 194, 4080–4088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurihara, S.; Kato, K.; Asada, K.; Kumagai, H.; Suzuki, H. A putrescine-inducible pathway comprising PuuE-YneI in which γ-aminobutyrate is degraded into succinate in Escherichia coli K-12. J. Bacteriol. 2010, 192, 4582–4591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samsonova, N.N.; Smirnov, S.V.; Altman, I.B.; Ptitsyn, L.R. Molecular cloning and characterization of Escherichia coli K12 ygjG gene. BMC Microbiol. 2003, 3, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaibe, E.; Metzer, E.; Halpern, Y.S. Metabolic pathway for the utilization of Larginine, L-ornithine, agmatine, and putrescine as nitrogen sources in Escherichia coli K-12. J. Bacteriol. 1985, 163, 933–937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaibe, E.; Metzer, E.; Halpern, Y.S. Control of utilization of L-arginine, L-ornithine, agmatine, and putrescine as nitrogen sources in Escherichia coli K-12. J. Bacteriol. 1985, 163, 938–942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forouhar, F.; Lee, I.S.; Vujcic, J.; Vujcic, S.; Shen, J.; Vorobiev, S.M.; Xiao, R.; Acton, T.B.; Montelione, G.T.; Porter, C.W.; et al. Structural and functional evidence for Bacillus subtilis PaiA as a novel N1- spermidine/spermine acetyltransferase. J. Biol. Chem. 2005, 280, 40328–40336. [Google Scholar] [CrossRef] [Green Version]
- Foster, A.; Barnes, N.; Speight, R.; Keane, M.A. Genomic organisation, activity and distribution analysis of the microbial putrescine oxidase degradation pathway. Syst. Appl. Microbiol. 2013, 36, 457–466. [Google Scholar] [CrossRef]
- Campilongo, R.; Di Martino, M.L.; Marcocci, L.; Pietrangeli, P.; Leuzzi, A.; Grossi, M.; Casalino, M.; Nicoletti, M.; Micheli, G.; Colonna, B.; et al. Molecular and functional profiling of the polyamine content in enteroinvasive E. coli: Looking into the gap between commensal E. coli and harmful Shigella. PLoS ONE 2014, 9, e106589. [Google Scholar] [CrossRef]
- Fukuchi, J.; Kashiwagi, K.; Takio, K.; Igarashi, K. Properties and structure of spermidine acetyltransferase in Escherichia coli. J. Biol. Chem. 1994, 269, 22581–22585. [Google Scholar] [CrossRef]
- Bollinger, J.M., Jr.; Kwon, D.S.; Huisman, G.W.; Kolter, R.; Walsh, C.T. Glutathionylspermidine metabolism in Escherichia coli. Purification, cloning, overproduction, and characterization of a bifunctional glutathionylspermidine synthetase/amidase. J. Biol. Chem. 1995, 270, 14031–14041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, D.S.; Lin, C.H.; Chen, S.; Coward, J.K.; Walsh, C.T.; Bollinger, J.M., Jr. Dissection of glutathionylspermidine synthetase/amidase from Escherichia coli into autonomously folding and functional synthetase and amidase domains. J. Biol. Chem. 1997, 272, 2429–2436. [Google Scholar] [CrossRef] [Green Version]
- Cardona-Cardona, Y.V.; Regla, I.; Juárez-Díaz, J.A.; Carrillo-Campos, J.; López-Ortiz, M.; Aguilera-Cruz, A.; Mújica-Jiménez, C.; Muñoz-Clares, R.A. The critical role of the aldehyde dehydrogenase PauC in spermine, spermidine, and diaminopropane toxicity in Pseudomonas aeruginosa: Its possible use as a drug target. FEBS J. 2022, 289, 2685–2705. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.D.; Itoh, Y.; Nakada, Y.; Jiang, Y. Functional analysis and regulation of the divergent spuABCDEFGH-spuI operons for polyamine uptake and utilization in Pseudomonas aeruginosa PAO1. J. Bacteriol. 2002, 184, 3765–3773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chou, H.T.; Li, J.Y.; Peng, Y.C.; Lu, C.D. Molecular characterization of PauR and its role in control of putrescine and cadaverine catabolism through the γ-glutamylation pathway in Pseudomonas aeruginosa PAO1. J. Bacteriol. 2013, 195, 3906–3913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, X.; Lu, C.-D. Characterization of Staphylococcus aureus responses to spermine stress. Curr. Microbiol. 2014, 69, 394–403. [Google Scholar] [CrossRef] [PubMed]
- Che, S.; Liang, Y.; Chen, Y.; Wu, W.; Liu, R.; Zhang, Q.; Bartlam, M. Structure of Pseudomonas aeruginosa spermidine dehydrogenase: A polyamine oxidase with a novel heme-binding fold. FEBS J. 2022, 289, 1911–1928. [Google Scholar] [CrossRef] [PubMed]
- Kakegawa, T.; Guo, Y.; Chiba, Y.; Miyazaki, T.; Nakamura, M.; Hirose, S.; Canellakis, Z.N.; Igarashi, K. Effect of acetylpolyamines on in vitro protein synthesis and on the growth of a polyamine-requiring mutant of Escherichia coli. J. Biochem. 1991, 109, 627–631. [Google Scholar] [CrossRef]
- Bai, F.; Xu, H.; Zhang, Q.; Qi, X.; Mou, R.; Bai, G.; Qiao, M. Functional characterization of pfm in protein secretion and lung infection of Pseudomonas aeruginosa. Can. J. Microbiol. 2011, 57, 829–837. [Google Scholar] [CrossRef] [PubMed]
- Planet, P.J.; Larussa, S.J.; Dana, A.; Smith, H.; Xu, A.; Ryan, C.; Uhlemann, A.C.; Boundy, S.; Goldberg, J.; Narechania, A.; et al. Emergence of the epidemic methicillin-resistant Staphylococcus aureus strain USA300 coincides with horizontal transfer of the arginine catabolic mobile element and speG-mediated adaptations for survival on skin. mBio 2013, 4, e00889-13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmer, D.P.; Soupene, E.; Lee, H.L.; Wendisch, V.F.; Khodursky, A.B.; Peter, B.J.; Bender, R.A.; Kustu, S. Nitrogen regulatory protein C-controlled genes of Escherichia coli: Scavenging as a defense against nitrogen limitation. Proc. Natl. Acad. Sci. USA 2000, 97, 14674–14679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maciag, A.; Peano, C.; Pietrelli, A.; Egli, T.; De Bellis, G.; Landini, P. In vitro transcription profiling of the σ S subunit of bacterial RNA polymerase: Redefinition of the σ S regulon and identification of σ S-specific promoter sequence elements. Nucleic Acids Res. 2011, 39, 5338–5355. [Google Scholar] [CrossRef] [Green Version]
- Zhao, K.; Liu, M.; Burgess, R.R. Promoter and regulon analysis of nitrogen assimilation factor, σ54, reveal alternative strategy for E. coli MG1655 flagellar biosynthesis. Nucleic Acids Res. 2010, 38, 1273–1283. [Google Scholar] [CrossRef] [Green Version]
- Nemoto, N.; Kurihara, S.; Kitahara, Y.; Asada, K.; Kato, K.; Suzuki, H. Mechanism for regulation of the putrescine utilization pathway by the transcription factor PuuR in Escherichia coli K-12. J. Bacteriol. 2012, 194, 3437–3447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Partridge, J.D.; Scott, C.; Tang, Y.; Poole, R.K.; Green, J. Escherichia coli transcriptome dynamics during the transition from anaerobic to aerobic conditions. J. Biol. Chem. 2006, 281, 27806–27815. [Google Scholar] [CrossRef] [Green Version]
- Randle, C.L.; Albro, P.W.; Dittmer, J.C. The phosphoglyceride composition of gram-negative bacteria and the changes in composition during growth. Biochim. Biophys. Acta (BBA)-Lipids Lipid Metab. 1969, 187, 214–220. [Google Scholar] [CrossRef]
- White, D.A. The phospholipids composition of mammalian tissues. In Form and Function of Phospholipids, 2nd ed.; Ansel, G.B., Hawthorne, J.N., Dawson, R.M.C., Eds.; Elsevier: Amsterdam, The Netherlands, 1973; pp. 441–483. [Google Scholar]
- Knaak, J.B.; Leung, H.-W.; Stott, W.T.; Busch, J.; Bilsky, J. Toxicology of mono-, di-, and triethanolamine. Rev. Environ. Contam. Toxicol. 1997, 149, 1–86. [Google Scholar] [PubMed]
- Vance, J.E.; Tasseva, G. Formation and function of phosphatidylserine and phosphatidylethanolamine in mammalian cells. Biochim. Biophys. Acta 2012, 1831, 543–554. [Google Scholar] [CrossRef] [PubMed]
- Bingham, E.; Cohrssen, B.; Powell, C.H. Patty’s Toxicology, 5th ed.; John Wiley & Sons: New York, NY, USA, 2001; Volume 1–9, p. 782. [Google Scholar]
- Anderson, J.C.; Kendall, M.M. Location, location, location. Salmonella senses ethanolamine to gauge distinct host environments and coordinate gene expression. Microb. Cell 2016, 3, 89–91. [Google Scholar] [CrossRef]
- Kofoid, E.; Rappleye, C.; Stojiljkovic, I.; Roth, J. The 17-gene ethanolamine (eut) operon of Salmonella typhimurium encodes five homologues of carboxysome shell proteins. J. Bacteriol. 1999, 181, 5317–5329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larson, T.J.; Ehrmann, M.; Boos, W. Periplasmic glycerophosphodiester phosphodiesterase of Escherichia coli, a new enzyme of the glp regulon. J. Biol. Chem. 1983, 258, 5428–5432. [Google Scholar] [CrossRef]
- Proulx, P.; Fung, C.K. Metabolism of phosphoglycerides in E. coli. IV. The positional specificity and properties of phospholipase A. Can. J. Biochem. 1969, 47, 1125–1128. [Google Scholar]
- Nandedkar, A.K. Report on the utilization of ethanolamine-1-14C by Mycobacterium 607. Biochem. Med. 1974, 11, 67–70. [Google Scholar] [CrossRef]
- Tsoy, O.; Ravcheev, D.; Mushegian, A. Comparative genomics of ethanolamine utilization. J. Bacteriol. 2009, 191, 7157–7164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garsin, D.A. Ethanolamine: A Signal to Commence a Host-Associated Lifestyle? mBio 2012, 1, e00066-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerlt, J.A. Tools and strategies for discovering novel enzymes and metabolic pathways. Perspect. Sci. 2016, 9, 24–32. [Google Scholar] [CrossRef] [Green Version]
- Korbel, J.O.; Doerks, T.; Jensen, L.J.; Perez-Iratxeta, C.; Kaczanowski, S.; Hooper, S.D.; Andrade, M.A.; Bork, P. Systematic association of genes to phenotypes by genome and literature mining. PLoS Biol. 2005, 3, e134. [Google Scholar] [CrossRef] [Green Version]
- Cronan, C.E., Jr.; Rock, C.O. Biosynthesis of membrane lipids. In Escherichia coli and Salmonella: Cellular and Molecular Biology; Neidhardt, F.C., Ed.; ASM Press: Washington, DC, USA, 1996; pp. 612–636. [Google Scholar]
- Gibellini, F.; Smith, T.K. The Kennedy pathway—De novo synthesis of phosphatidylethanolamine and phosphatidylcholine. IUBMB Life 2010, 62, 414–428. [Google Scholar] [CrossRef]
- Penrod, J.T.; Mace, C.C.; Roth, J.R. A pH-sensitive function and phenotype: Evidence that EutH facilitates diffusion of uncharged ethanolamine in Salmonella enterica. J. Bacteriol. 2004, 186, 6885–6890. [Google Scholar] [CrossRef] [Green Version]
- Stojiljkovic, I.; Baumler, A.J.; Heffron, F. Ethanolamine utilization in Salmonella typhimurium: Nucleotide sequence, protein expression, and mutational analysis of the cchA, cchB, eutE, eutJ, eutG, eutH gene cluster. J. Bacteriol. 1995, 177, 1357–1366. [Google Scholar] [CrossRef] [Green Version]
- Garsin, D.A. Ethanolamine Utilization in Bacterial Pathogens: Roles and Regulation. Nat. Rev. Microbiol. 2010, 8, 290–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, G.W.; Chang, J.T. Evidence for the B12-dependent enzyme ethanolamine deaminase in Salmonella. Nature 1975, 254, 150–151. [Google Scholar] [CrossRef]
- Del Papa, M.F.; Perego, M. Ethanolamine activates a sensor histidine kinase regulating its utilization in Enterococcus faecalis. J. Bacteriol. 2008, 190, 7147–7156. [Google Scholar] [CrossRef] [Green Version]
- Blackwell, C.M.; Scarlett, F.A.; Turner, J.M. Ethanolamine catabolism by bacteria, including Escherichia coli. Biochem. Soc. Trans. 1976, 4, 495–497. [Google Scholar] [CrossRef] [PubMed]
- Blackwell, C.M.; Turner, J.M. Microbial metabolism of amino alcohols. Formation of coenzyme B12-dependent ethanolamine ammonia-lyase and its concerted induction in Escherichia coli. Biochem. J. 1978, 176, 751–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khatri, N.; Khatri, I.; Subramanian, S.; Raychaudhuri, S. Ethanolamine utilization in Vibrio alginolyticus. Biol. Direct 2012, 7, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldman, D.S.; Wagner, M.J. Enzyme systems in the mycobacteria. XIII. Glycine dehydrogenase and the glyoxylic acid cycle. Biochim. Biophys. Acta 1962, 65, 297–306. [Google Scholar] [CrossRef]
- Shukla, S.D.; Turner, J.M. Biosynthetic utilization of ethanolamine for lipid synthesis by bacteria. Biochem. J. 1980, 186, 13–19. [Google Scholar] [CrossRef] [Green Version]
- San Francisco, B.; Zhang, X.; Whalen, K.; Gerlt, K. A novel pathway for bacterial ethanolamine metabolism. FASEB J. 2015, 29, 573.45. [Google Scholar] [CrossRef]
- Roof, D.M.; Roth, J.R. Ethanolamine utilization in Salmonella typhimurium. J. Bacteriol. 1988, 170, 3855–3863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheppard, D.E.; Penrod, J.T.; Bobik, T.; Kofoid, E.; Roth, J.R. Evidence that a B12-adenosyl transferase is encoded within the ethanolamine operon of Salmonella enterica. J. Bacteriol. 2004, 186, 7635–7644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaval, K.G.; Garsin, D.A. Ethanolamine Utilization in Bacteria. mBio 2018, 3, e00172-12. [Google Scholar] [CrossRef] [Green Version]
- Yeates, T.O.; Kerfeld, C.A.; Heinhorst, S.; Cannon, G.C.; Shively, J.M. Proteinbased organelles in bacteria: Carboxysomes and related microcompartments. Nat. Rev. Microbiol. 2008, 6, 681–691. [Google Scholar] [CrossRef] [PubMed]
- Brinsmade, S.R.; Escalante-Semerena, J.C. The eutD gene of Salmonella enterica encodes a protein with phosphotransacetylase enzyme activity. J. Bacteriol. 2004, 186, 1890–1892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penrod, J.T.; Roth, J.R. Conserving a volatile metabolite: A role for carboxysome-like organelles in Salmonella enterica. J. Bacteriol. 2006, 188, 2865–2874. [Google Scholar] [CrossRef] [Green Version]
- Bradbeer, C. The clostridial fermentations of choline and ethanolamine. II. Requirement for a cobamide coenzyme by an ethanolamine deaminase. J. Biol. Chem. 1965, 240, 4675–4681. [Google Scholar] [CrossRef]
- Bologna, F.P.; Campos-Bermudez, V.A.; Saavedra, D.D.; Andreo, C.S.; Drincovich, M.F. Characterization of Escherichia coli EutD: A phosphotransacetylase of the ethanolamine operon. J. Microbiol. 2010, 48, 629–636. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.W.; Turner, J.M. Interrelationships between the enzymes of ethanolamine metabolism in Escherichia coli. J. Gen. Microbiol. 1984, 130, 299–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brinsmade, S.R.; Paldon, T.; Escalante-Semerena, J.C. Minimal functions and physiological conditions required for growth of Salmonella enterica on ethanolamine in the absence of the metabolosome. J. Bacteriol. 2005, 187, 8039–8046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Starai, V.J.; Garrity, J.; Escalante-Semerena, J.C. Acetate excretion during growth of Salmonella enterica on ethanolamine requires phosphotransacetylase (EutD) activity, and acetate recapture requires acetyl-CoA synthetase (Acs) and phosphotransacetylase (Pta) activities. Microbiology 2005, 151, 3793–3801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barisch, C.; Soldati, T. Mycobacterium marinum Degrades Both Triacylglycerols and Phospholipids from Its Dictyostelium Host to Synthesise Its Own Triacylglycerols and Generate Lipid Inclusions. PLoS Pathog. 2017, 13, e1006095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larrouy-Maumus, G.; Biswas, T.; Hunt, D.M.; Kelly, G.; Tsodikov, O.V.; Sório de Carvalho, L.P. Discovery of a glycerol 3-phosphate phosphatase reveals glycerophospholipid polar head recycling in Mycobacterium tuberculosis. Proc. Natl. Acad. Sci. USA 2013, 110, 11320–11325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nandedkar, A.K.N. Biosynthesis of phosphatidyl ethanolamine in Mycobacterium 607. Biochem. Med. 1975, 12, 116–122. [Google Scholar] [CrossRef]
- Kaval, K.G.; Gebbie, M.; Goodson, J.R.; Cruz, M.R.; Winkler, W.C.; Garsin, D.A. Ethanolamine Utilization and Bacterial Microcompartment Formation Are Subject to Carbon Catabolite Repression. J. Bacteriol. 2019, 201, e00703-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roof, D.M.; Roth, J.R. Autogenous regulation of ethanolamine utilization by a transcriptional activator of the eut operon in Salmonella typhimurium. J. Bacteriol. 1992, 174, 6634–6643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luzader, D.H.; Clark, D.E.; Gonyar, L.A.; Kendall, M.M. EutR is a direct regulator of genes that contribute to metabolism and virulence in enterohemorrhagic Escherichia coli O157:H7. J. Bacteriol. 2013, 195, 4947–4953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fox, K.A.; Ramesh, A.; Stearns, J.E.; Bourgogne, A.; Reyes-Jara, A.; Winkler, W.C.; Garsin, D.A. Multiple posttranscriptional regulatory mechanisms partner to control ethanolamine utilization in Enterococcus faecalis. Proc. Natl. Acad. Sci. USA 2009, 106, 4435–4440. [Google Scholar] [CrossRef] [Green Version]
- Mowbray, S.L.; Kathiravan, M.K.; Pandey, A.A.; Odell, L.R. Inhibition of glutamine synthetase: A potential drug target in Mycobacterium tuberculosis. Molecules 2014, 19, 13161–13176. [Google Scholar] [CrossRef] [Green Version]
- Gallant, J.L.; Viljoen, A.J.; van Helden, P.D.; Wiid, I.J. Glutamate Dehydrogenase Is Required by Mycobacterium bovis BCG for Resistance to Cellular Stress. PLoS ONE 2016, 11, e0147706. [Google Scholar] [CrossRef] [PubMed]
- Somani, R.R.; Rai, P.R.; Kandpile, P.S. Ornithine Decarboxylase Inhibition: A Strategy to Combat Various Diseases. Mini Rev. Med. Chem. 2018, 18, 1008–1021. [Google Scholar] [CrossRef] [PubMed]
- Carter, N.S.; Kawasaki, Y.; Nahata, S.S.; Elikaee, S.; Rajab, S.; Salam, L.; Alabdulal, M.Y.; Broessel, K.K.; Foroghi, F.; Abbas, A.; et al. Polyamine Metabolism in Leishmania Parasites: A Promising Therapeutic Target. Med. Sci. 2022, 10, 24. [Google Scholar] [CrossRef] [PubMed]
- Ayoola, M.B.; Shack, L.A.; Lee, J.H.; Lim, J.; Eoh, H.; Swiatlo, E.; Phanstiel, O., IV; Nanduri, B. Difluoromethylornithine (DFMO) and AMXT 1501 inhibit capsule biosynthesis in pneumococci. Sci. Rep. 2022, 12, 11804. [Google Scholar] [CrossRef] [PubMed]
- Reigada, C.; Sayé, M.; Phanstiel, O., IV; Valera-Vera, E.; Miranda, M.R.; Pereira, C.A. Identification of Trypanosoma cruzi Polyamine Transport Inhibitors by Computational Drug Repurposing. Front. Med. (Lausanne) 2019, 6, 256. [Google Scholar] [CrossRef] [Green Version]
- Shah, P.; Nanduri, B.; Swiatlo, E.; Ma, Y.; Pendarvis, K. Polyamine biosynthesis and transport mechanisms are crucial for fitness and pathogenesis of Streptococcus pneumoniae. Microbiology 2011, 157, 504–515. [Google Scholar] [CrossRef] [Green Version]
- Jelsbak, L.; Thomsen, L.E.; Wallrodt, I.; Jensen, P.R.; Olsen, J.E. Polyamines are required for virulence in Salmonella enterica serovar Typhimurium. PLoS ONE 2012, 7, e36149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rifkin, M.R.; Strobos, C.A.; Fairlamb, A.H. Specificity of ethanolamine transport and its further metabolism in Trypanosoma brucei. J. Biol. Chem. 1995, 270, 16160–16166. [Google Scholar] [CrossRef] [Green Version]
- Roberts, S.; Ullman, B. Parasite Polyamines as Pharmaceutical Targets. Curr. Pharm. Des. 2017, 23, 3325–3341. [Google Scholar] [CrossRef]
- Mann, S.; Frasca, K.; Scherrer, S.; Henao-Martínez, A.F.; Newman, S.; Ramanan, P.; Suarez, J.A. A Review of Leishmaniasis: Current Knowledge and Future Directions. Curr. Trop. Med. Rep. 2021, 8, 121–132. [Google Scholar] [CrossRef]
- Thiriot, J.D.; Martinez-Martinez, Y.B.; Endsley, J.J.; Torres, A.G. Hacking the host: Exploitation of macrophage polarization by intracellular bacterial pathogens. Pathog. Dis. 2020, 78, ftaa009. [Google Scholar] [CrossRef] [Green Version]
- Perdeh, J.; Berioso, B.; Love, Q.; LoGiudice, N.; Le, T.L.; Harrelson, J.P.; Roberts, S.C. Critical functions of the polyamine putrescine for proliferation and viability of Leishmania donovani parasites. Amino Acids 2020, 52, 261–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boitz, J.M.; Yates, P.A.; Kline, C.; Gaur, U.; Wilson, M.E.; Ullman, B.; Roberts, S.C. Leishmania donovani ornithine decarboxylase is indispensable for parasite survival in the mammalian host. Infect. Immun. 2009, 77, 756–763. [Google Scholar] [CrossRef] [Green Version]
- Samal, K.; Zhao, P.; Kendzicky, A.; Yco, L.P.; McClung, H.; Gerner, E.; Burns, M.; Bachmann, A.S.; Sholler, G. AMXT-1501, a novel polyamine transport inhibitor, synergizes with DFMO in inhibiting neuroblastoma cell proliferation by targeting both ornithine decarboxylase and polyamine transport. Int. J. Cancer 2013, 133, 1323–1333. [Google Scholar] [CrossRef] [PubMed]
- Muth, A.; Kamel, J.; Kaur, N.; Shicora, A.C.; Ayene, I.S.; Gilmour, S.K.; Phanstiel, O., IV. Development of polyamine transport ligands with improved metabolic stability and selectivity against specific human cancers. J. Med. Chem. 2013, 56, 5819–5828. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Yamada, C.; Kato, K. Gamma-glutamyl compounds and their enzymatic production using bacterial gamma-glutamyltranspeptidase. Amino Acids 2007, 32, 333–340. [Google Scholar] [CrossRef]
- Kruh, N.A.; Troudt, J.; Izzo, A.; Prenni, J.; Dobos, K.M. Portrait of a pathogen: The Mycobacterium tuberculosis proteome in vivo. PLoS ONE 2010, 5, e13938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Schematic illustration of bacterial nitrogen metabolism. The main nitrogen sources, their uptake, and assimilation routes are depicted, as well as representative example of proteins involved in these processes. Enzyme systems for ammonia assimilation are shown in detail. GS: glutamine synthetase; GOGAT: glutamine-2-oxoglutarate-aminotransferase; GDH: glutamate dehydrogenase. Red arrows: GS/GOGAT pathway; light gray arrow: GDH pathway; black arrows: other metabolic routes; dark gray cylinders: transporters [5,6,16].
Figure 1.
Schematic illustration of bacterial nitrogen metabolism. The main nitrogen sources, their uptake, and assimilation routes are depicted, as well as representative example of proteins involved in these processes. Enzyme systems for ammonia assimilation are shown in detail. GS: glutamine synthetase; GOGAT: glutamine-2-oxoglutarate-aminotransferase; GDH: glutamate dehydrogenase. Red arrows: GS/GOGAT pathway; light gray arrow: GDH pathway; black arrows: other metabolic routes; dark gray cylinders: transporters [5,6,16].

Figure 2.
Combined polyamine biosynthetic pathways in bacteria. Enzymes that catalyze central reactions of polyamine biosynthesis as well as substrates and products of reactions are shown. Both ODC and ADC pathways and also LDC-dependent cadaverine production are depicted. Polyamine products are marked in boxes. SAM: S-adenosylmethionine; dSAM: decarboxylated S-adenosylmethionine [14,117].
Figure 2.
Combined polyamine biosynthetic pathways in bacteria. Enzymes that catalyze central reactions of polyamine biosynthesis as well as substrates and products of reactions are shown. Both ODC and ADC pathways and also LDC-dependent cadaverine production are depicted. Polyamine products are marked in boxes. SAM: S-adenosylmethionine; dSAM: decarboxylated S-adenosylmethionine [14,117].

Figure 4.
Combined schematic model of ethanolamine and phosphatidylethanolamine biosynthesis in bacteria [229,232,240]. Biosynthetic reactions with the respective substrates, products, and involved enzymes are shown. The bacteria indirectly biosynthesise of ethanolamine from serine via the phosphatidylserine synthase and the phosphatidylserine decarboxylase is depicted. CDP: cytidine diphosphate.
Figure 4.
Combined schematic model of ethanolamine and phosphatidylethanolamine biosynthesis in bacteria [229,232,240]. Biosynthetic reactions with the respective substrates, products, and involved enzymes are shown. The bacteria indirectly biosynthesise of ethanolamine from serine via the phosphatidylserine synthase and the phosphatidylserine decarboxylase is depicted. CDP: cytidine diphosphate.


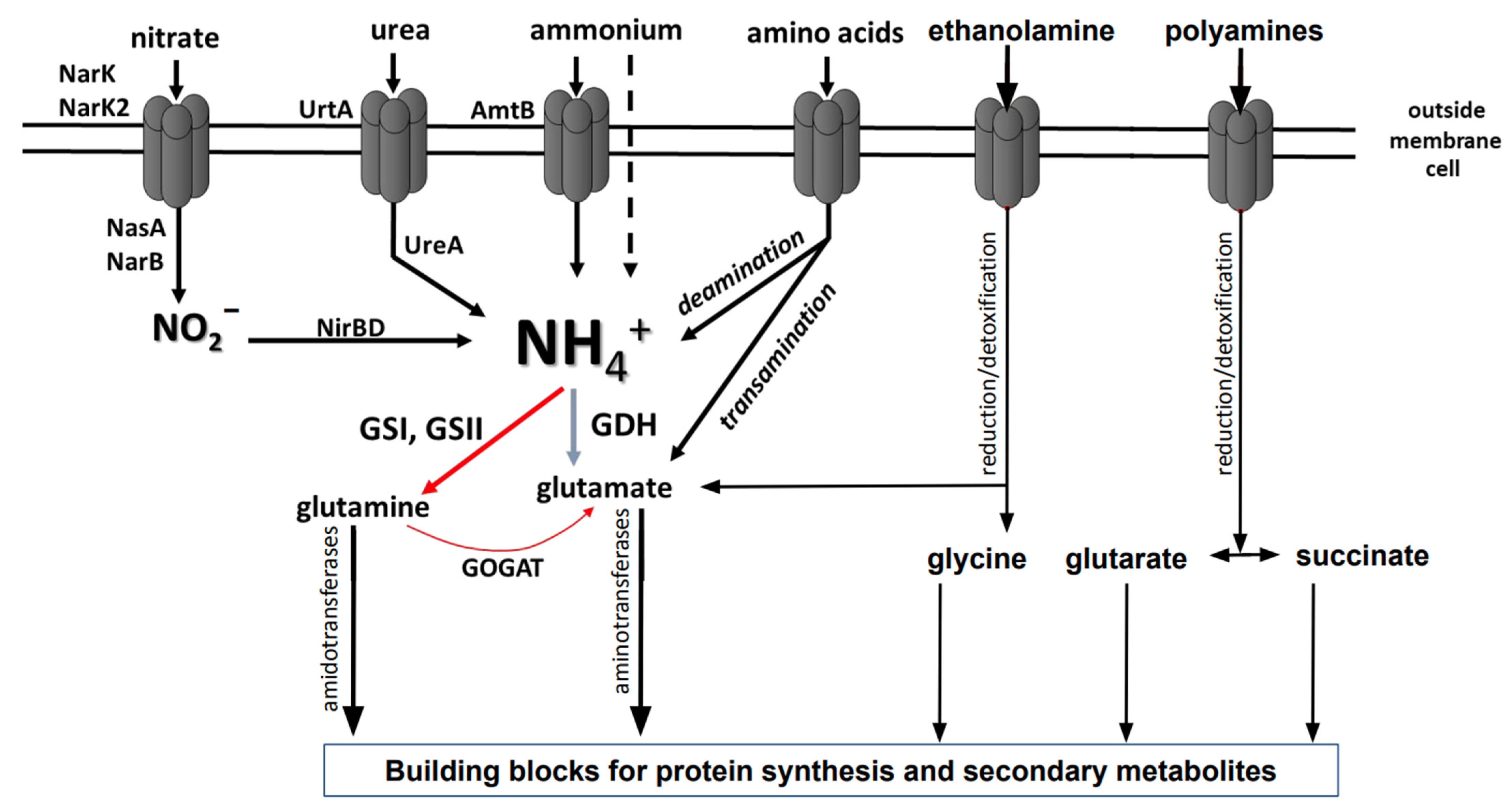{kind=link}
{kind=link}
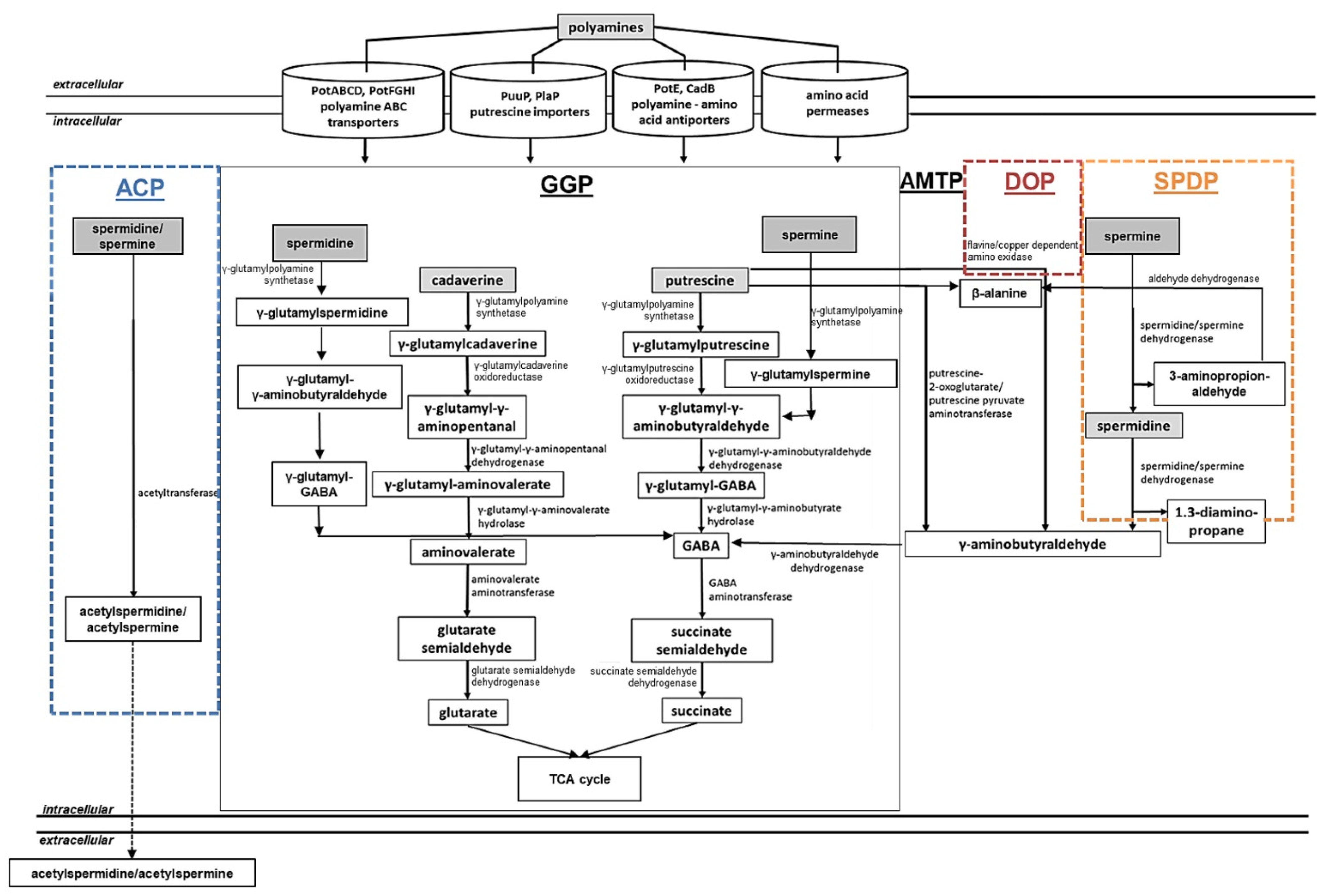{kind=link}
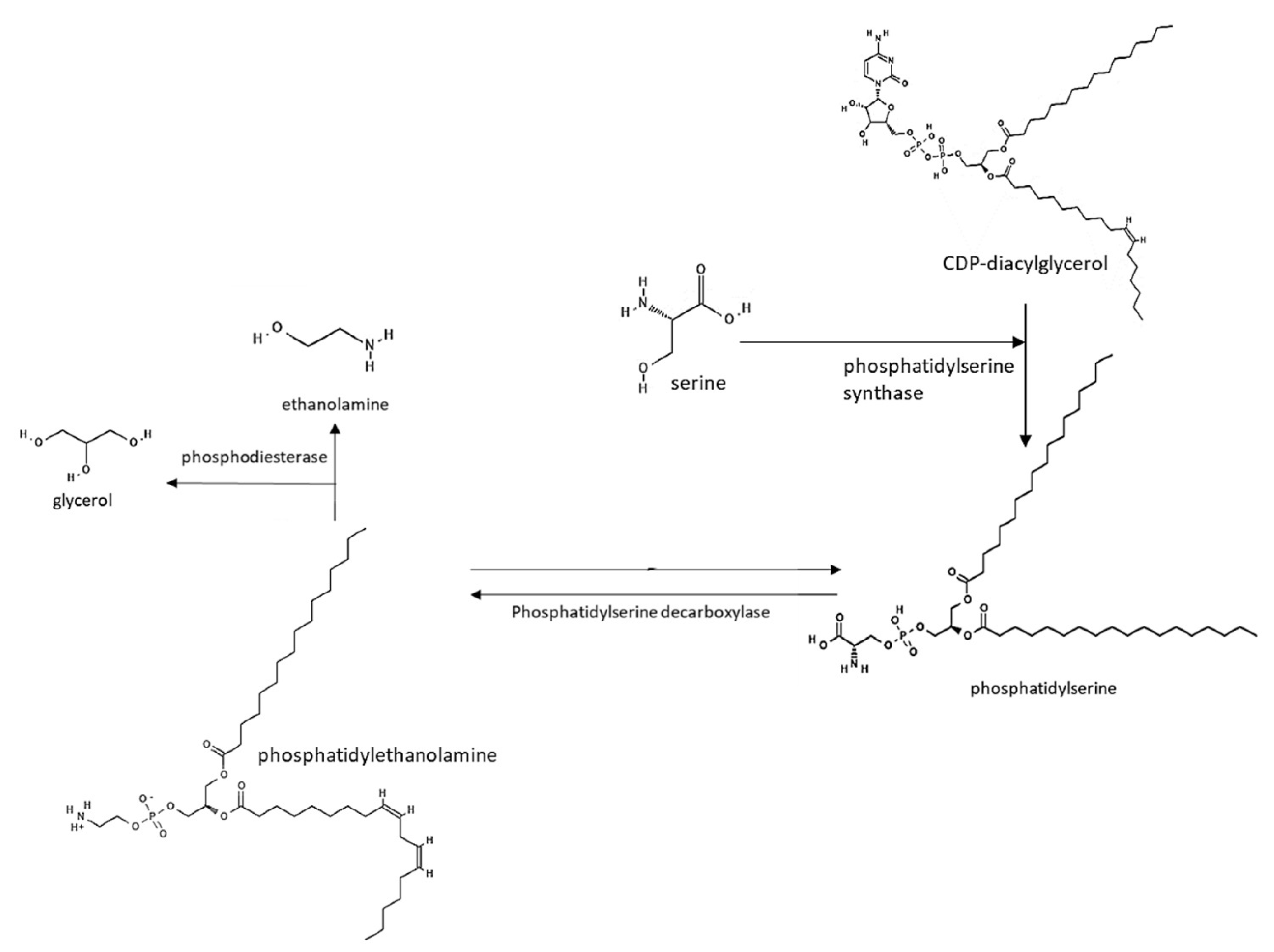{kind=link}
{kind=link}
Table 1.
List of proteins involved in polyamine utilization in E. coli, P. aeruginosa, and S. coelicolor [113,115].
| Annotated Function | Homologue in E. coli | Homologue in P. aeruginosa | Homologue in S. coelicolor |
|---|---|---|---|
| Polyamine ABC transporter ATP-binding protein PotA-like | PotA (b1126)/YdcT (b1441) | PAO603/PAO326 | SCO3453 |
| Polyamine ABC transporter ATP-binding protein PotC-like | PotC (b1124)/YdcV (b1443) | PAO324/PotC (PA3609) | SCO3454 |
| Polyamine ABC transporter protein | PotB (b1125)/YdcU (b1442) | PotB (PA0205)/PA3252 | SCO3455 |
| Polyamine ABC transporter protein—substrate binding protein | YnjB (b1754) | PA0203 | SCO3456 |
| Amino acid/polyamine permease | PuuP (b1296)/PlaP (b2014) | PA5510 | SCO5057 |
| Lysine/ornithine decarboxylase-like enzyme | - | - | SCO5651 |
| Pyruvate-polyamine aminotransferase | PatA (b3073) | SpuC (PA0299) | SCO5655 |
| Lrp/AsnC family transcriptional regulator | - | - | SCO5656 |
| γ-aminobutyraldehyde or γ-glutamyl-γ-amino-butyraldehyde dehydrogenase | PatD (b1444)/PuuC (b1300) | BetB (PA5373)/PAO219 | SCO5657 |
| Polyamine-binding lipoprotein | PotF (b0854) | SpuD (PA0300) | SCO5658 |
| γ-aminobutyraldehyde dehydrogenase or 4-guanidino-butyraldehyde dehydrogenase | PatD (b1444) PuuC (b1300) | PauC/KauB (PA5312) | SCO5666 |
| Polyamine ABC transporter substrate-binding protein | PotF (b0854) | SpuE (PA0301) | SCO5667 |
| Polyamine ABC transporter substrate-binding protein | PotG (b0855) | SpuF (PA0302) | SCO5668 |
| Polyamine ABC-transporter integral membrane protein | PotH (b0856) | SpuG (PA0303) | SCO5669 |
| Polyamine ABC-transporter integral membrane protein | PotI (b0857) | SpuH (PA0304) | SCO5670 |
| γ-glutamyl-polyamine oxidoreductase | PuuB (b1301) | PauB3 (PA2776) | SCO5671 |
| γ-aminobutyrate aminotransferase gabT-like or puuE-like | GabT (b2662)/PuuE (b1302) | GabT (PA266) | SCO5676 |
| Succinate-semialdehyde dehydrogenase gabD-like | GabD (b2661) | GabD (PA0265) | SCO5679 |
| Amino acids/polyamine permease | PuuP (b1296) | PA5510/PAO322 | SCO5977 |
| Hydrolase | - | - | SCO6960 |
| Amidohydrolase | - | - | SCO6961 |
| γ-glutamyl-polyamine synthetase | PuuA (b1297) | PauA7 (PA5508)/SpuI (PA0296) | SCO6962 |
Table 2.
Combined list of enzymes involved in ethanolamine utilization in C. salexigens and S. coelicolor [114,237].
| Annotated Function | Orthologues in C. salexigens (Protein Family) | Orthologues in S. coelicolor |
|---|---|---|
| γ-glutamyl ethanolamine synthetase/ethanolamine γ-glutamylase | PF00120 | SCO1613 |
| γ-glutamyl ethanolamine dehydrogenase/iron-dependent dehydrogenase | PF00465 | SCO1611 |
| γ-glutamyl aldehyde dehydrogenase | PF00171 | SCO1612 |
| γ-glutamyl glycine amidohydrolase /formylglutamate amidohydrolase | PF05013 | SCO1615 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Krysenko, S.; Wohlleben, W. Polyamine and Ethanolamine Metabolism in Bacteria as an Important Component of Nitrogen Assimilation for Survival and Pathogenicity. Med. Sci. 2022, 10, 40. https://doi.org/10.3390/medsci10030040
AMA Style
Krysenko S, Wohlleben W. Polyamine and Ethanolamine Metabolism in Bacteria as an Important Component of Nitrogen Assimilation for Survival and Pathogenicity. Medical Sciences. 2022; 10(3):40. https://doi.org/10.3390/medsci10030040
Chicago/Turabian StyleKrysenko, Sergii, and Wolfgang Wohlleben. 2022. "Polyamine and Ethanolamine Metabolism in Bacteria as an Important Component of Nitrogen Assimilation for Survival and Pathogenicity" Medical Sciences 10, no. 3: 40. https://doi.org/10.3390/medsci10030040
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.