
Response of Red-Backed Salamanders (Plethodon Cinereus) to Changes in Hemlock Forest Soil Driven by Invasive Hemlock Woolly Adelgid (Adelges Tsugae)


Abstract
:1. Introduction
2. Materials and Methods
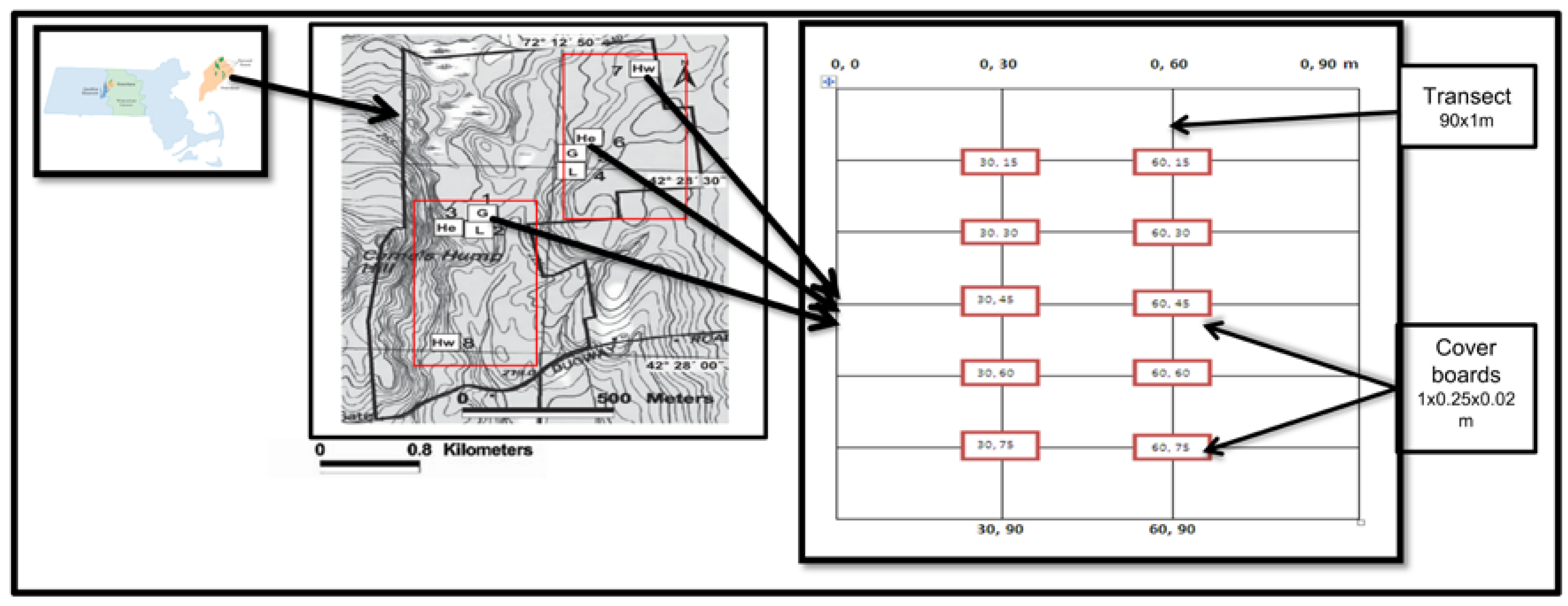
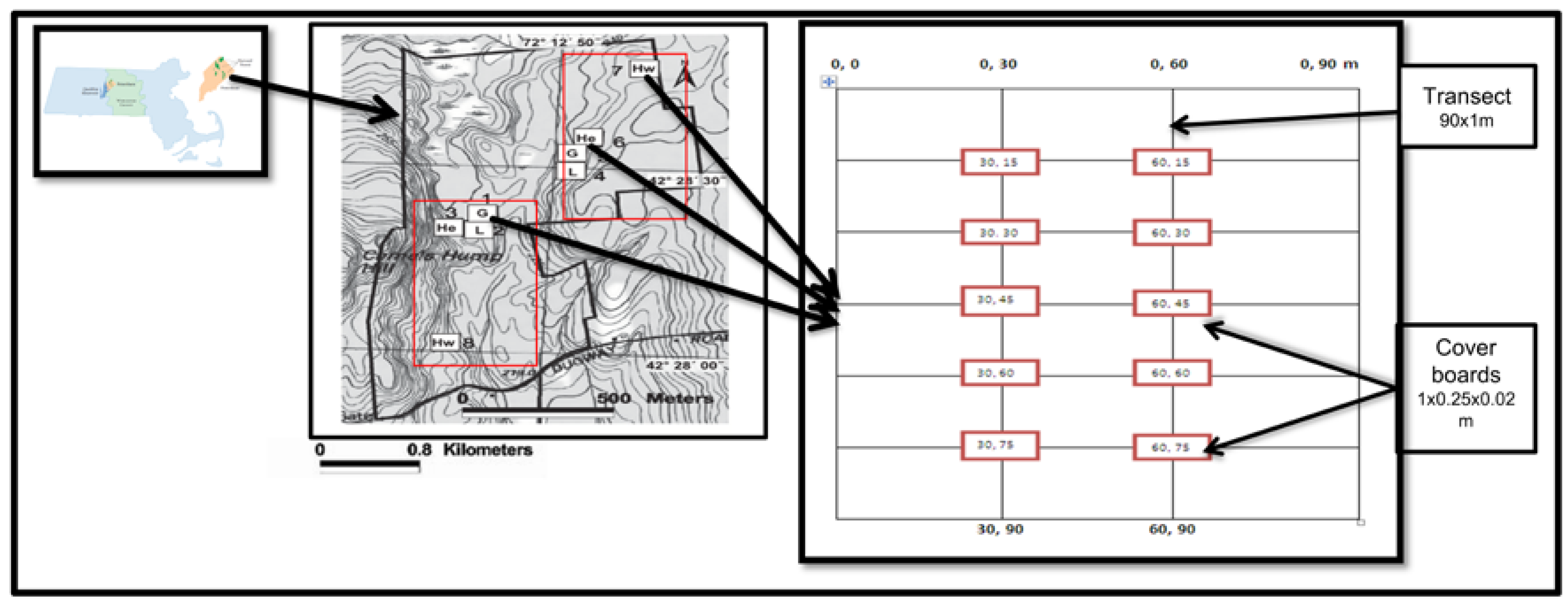
2.1. Study Site and Design
2.2. Salamander Sampling
2.3. Soil Sampling
2.4. Statistical Analysis
3. Results
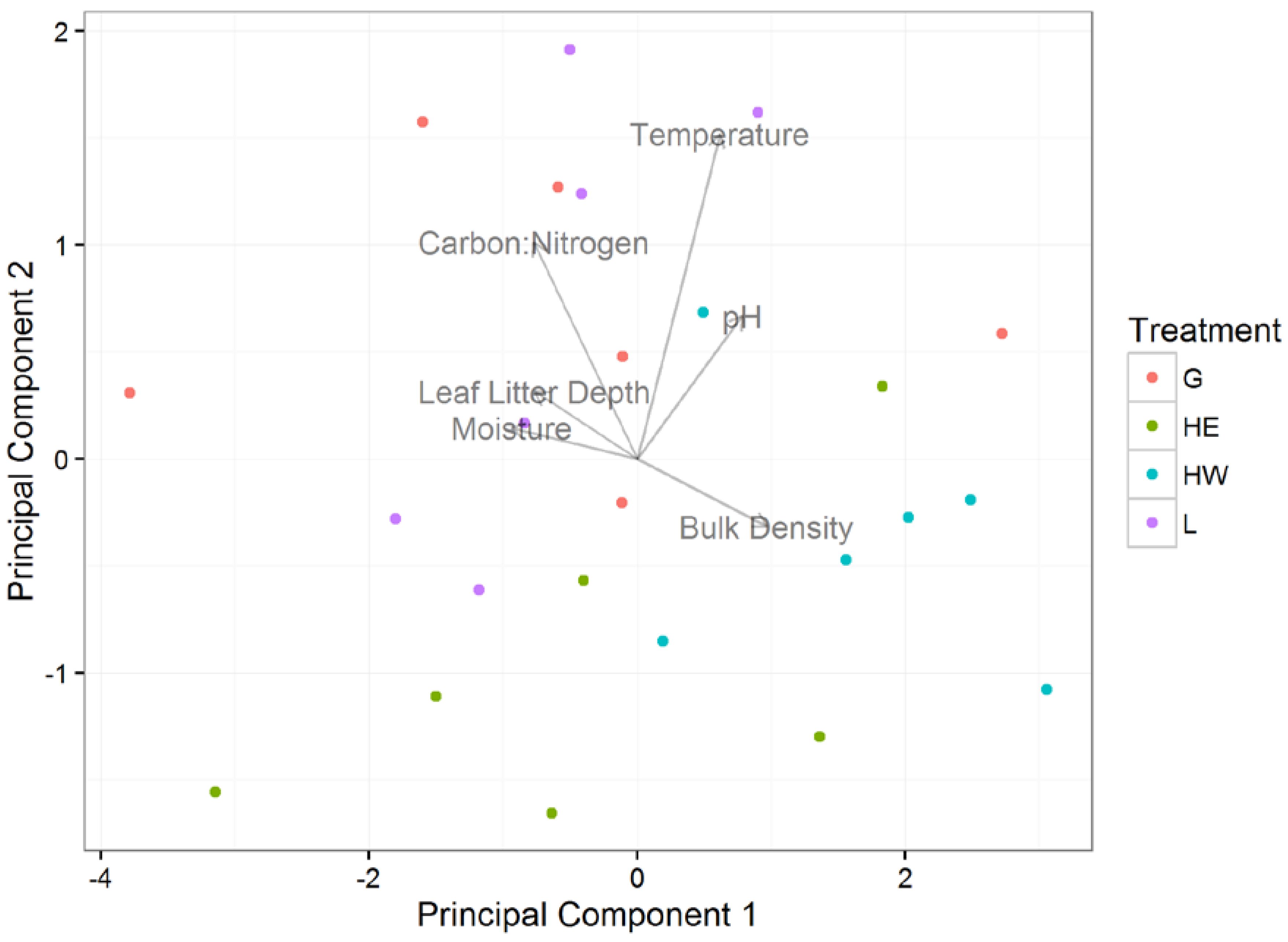
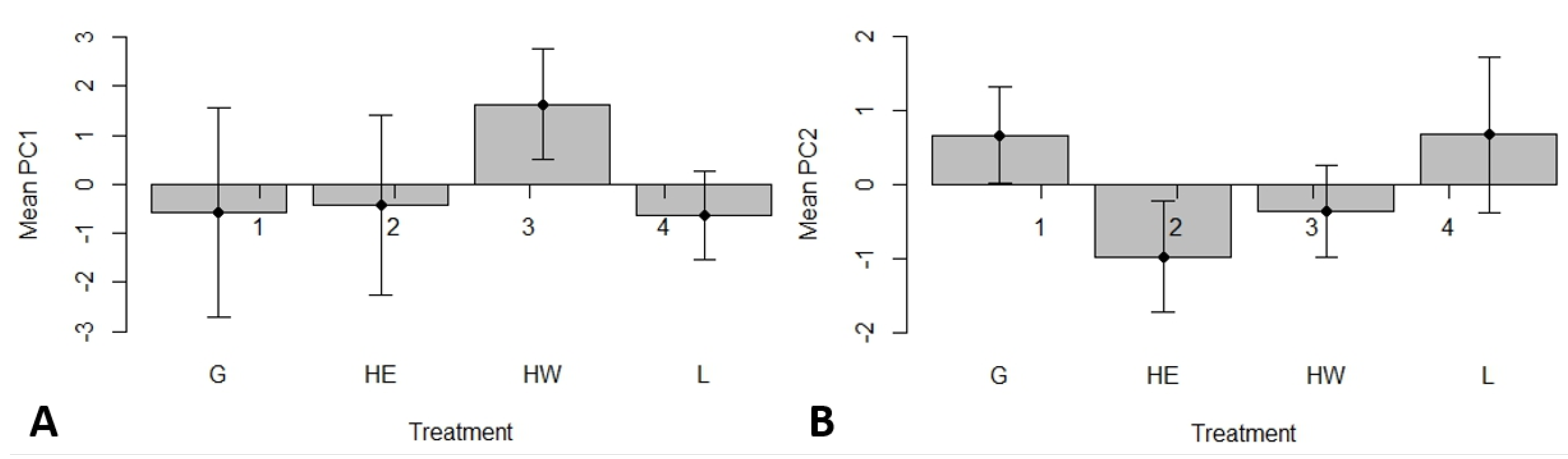
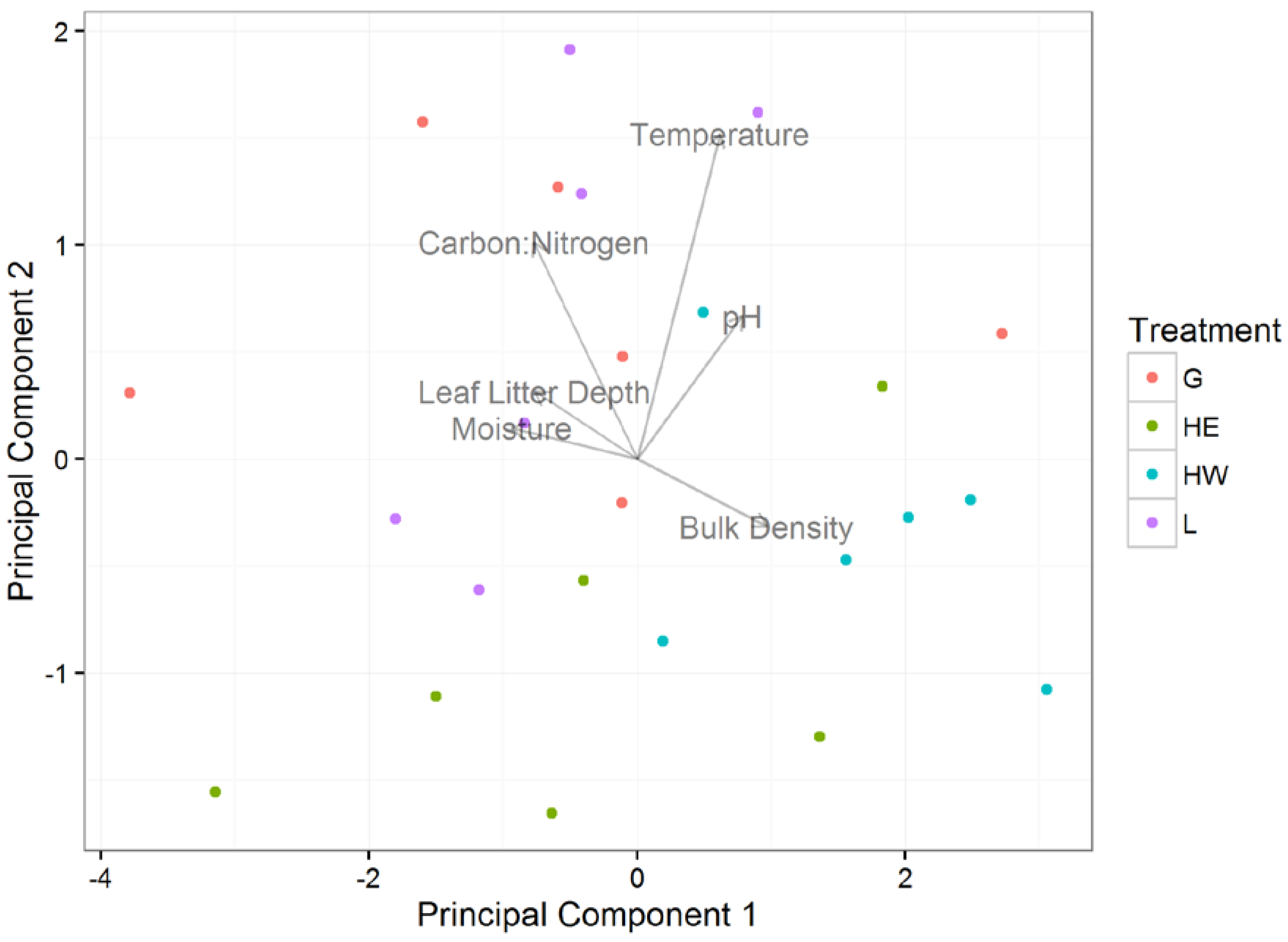
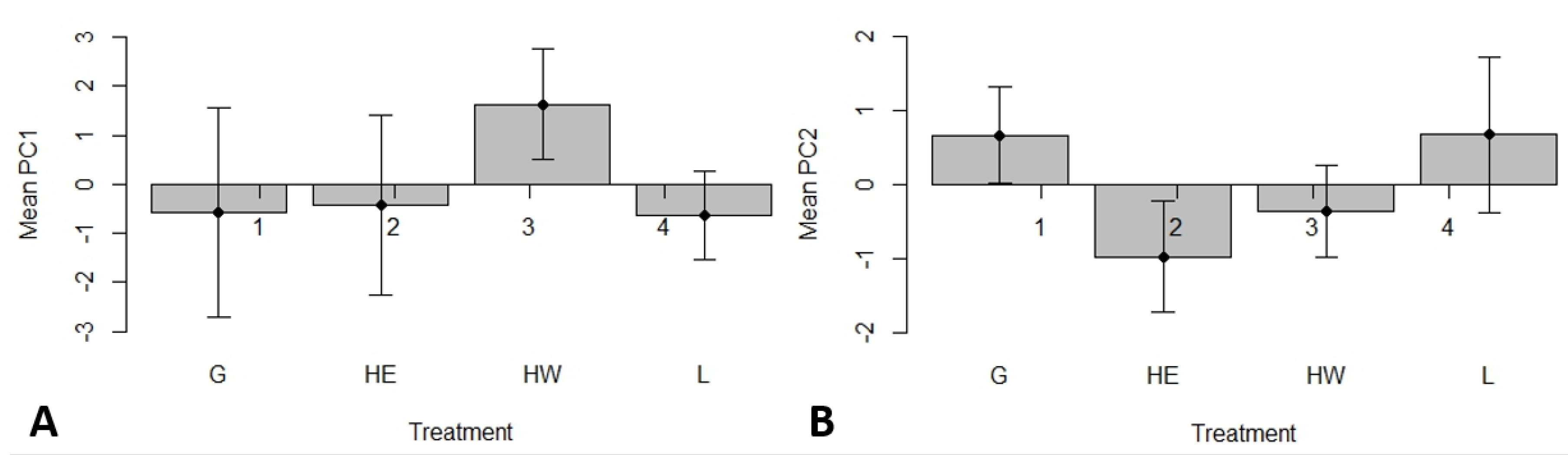
3.1. Effects of Plot Treatment on Soil Characteristics and Salamander Relative Abundance
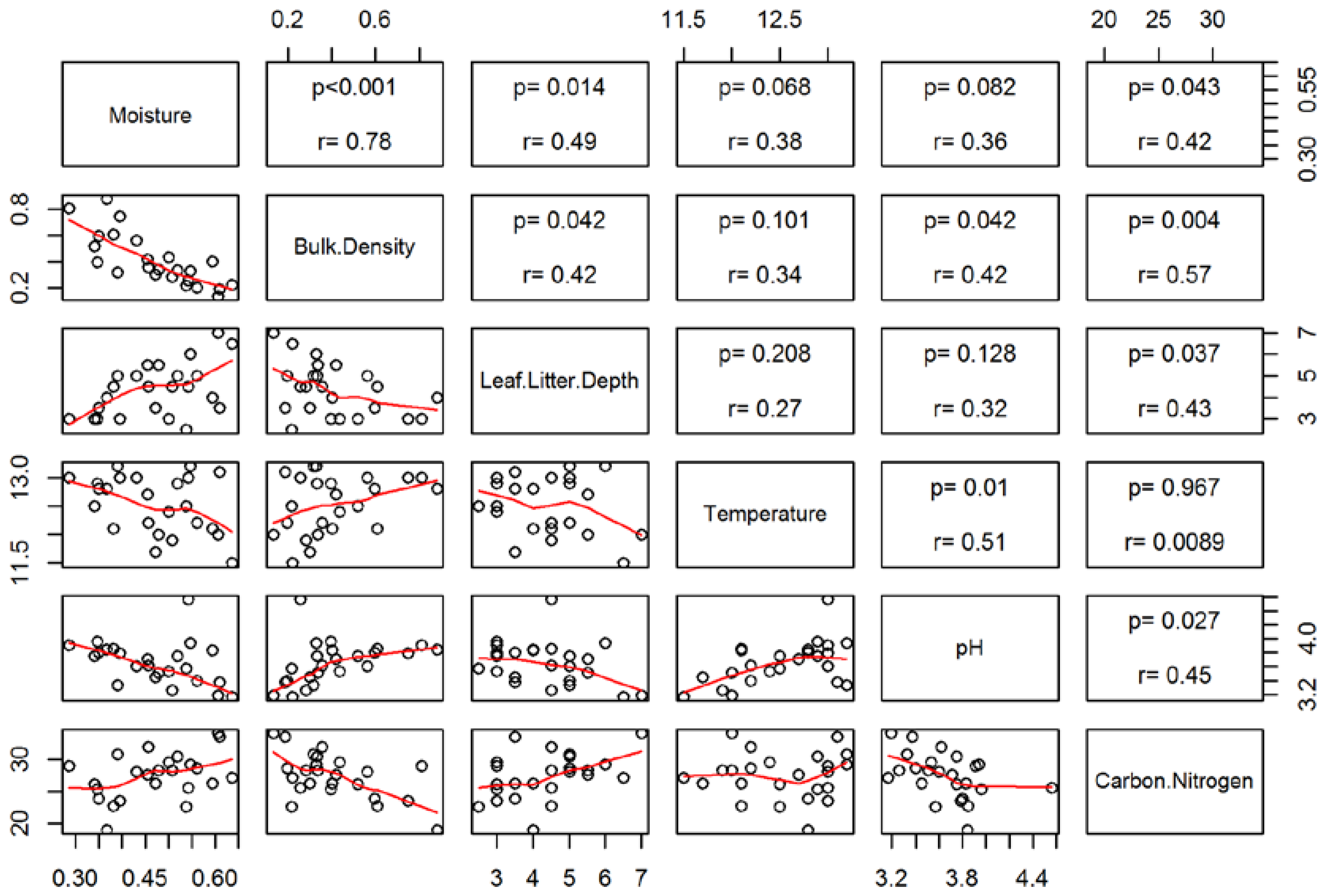
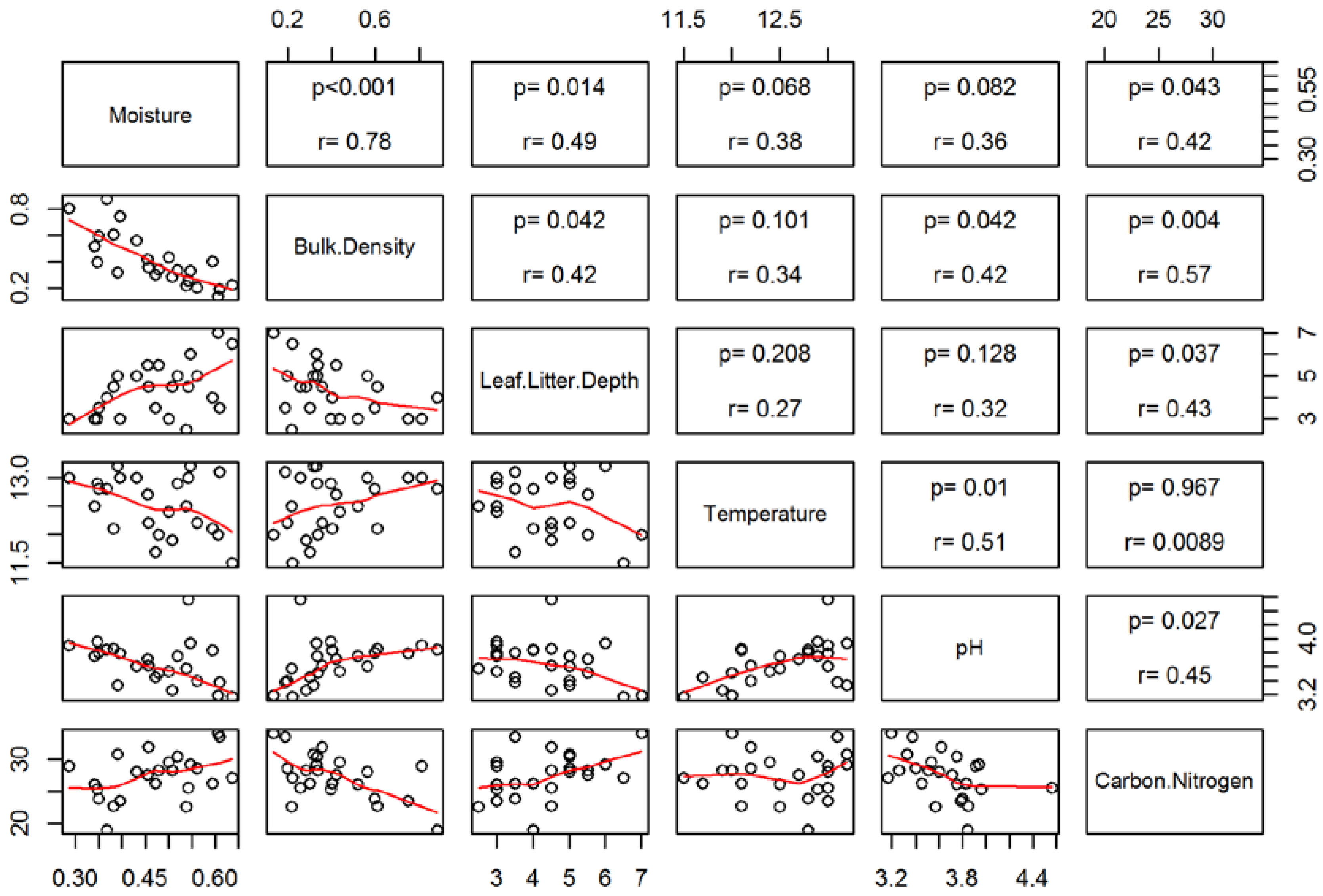
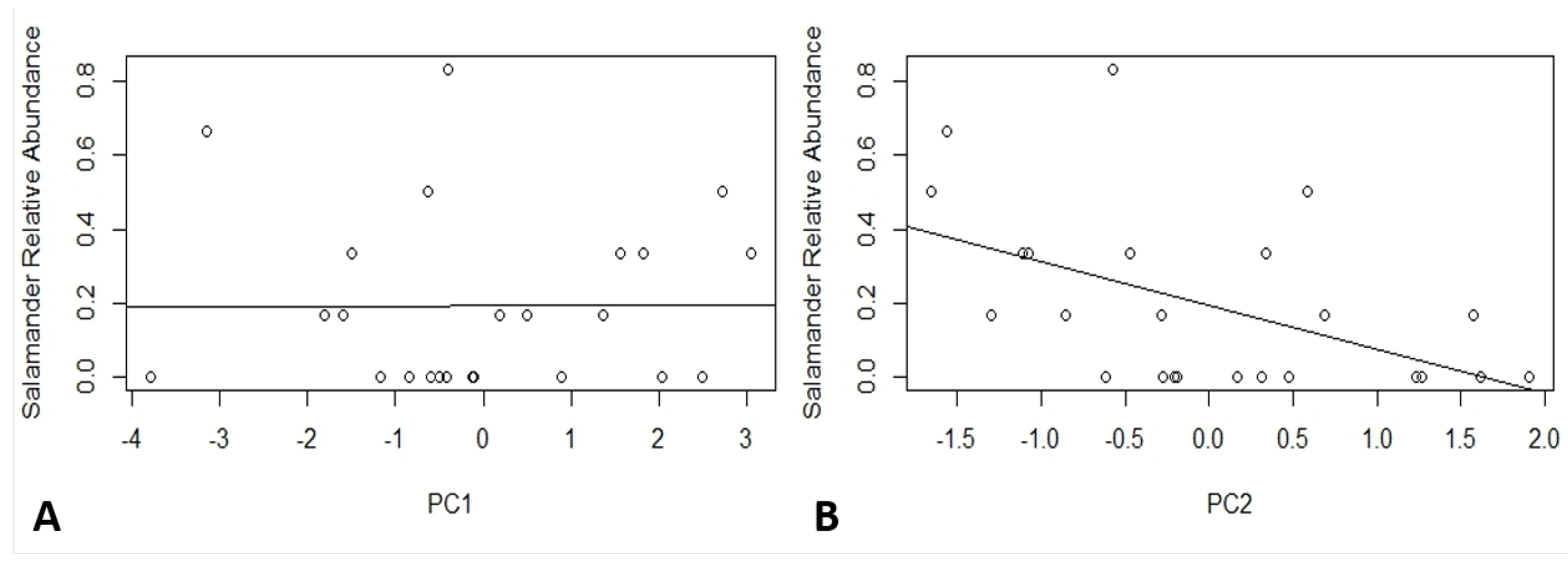
3.2. Soil Characteristics and Salamander Relative Abundance
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Cheah, C.; Montegomery, M.; Salem, S.; Parker, B.; Skinner, M.; Costa, S.; Reardon, R.; Onken, B. Biological Control of Hemlock Woolly Adelgid; Forest Health Technology Enterprise Team: Fort Collins, CO, USA, 2004; p. 22. [Google Scholar]
- Mathewson, B. Salamanders in a changing environment on hemlock hill. Arnoldia 2007, 65, 19–25. [Google Scholar]
- Ellison, A.M.; Bank, M.S.; Clinton, B.D.; Colburn, E.A.; Elliott, K.; Ford, C.R.; Kloeppel, B.D.; Knoepp, J.D.; Lovett, G.M.; Orwig, D.A.; et al. Loss of foundation species: Consequences for the structure and dynamics of forested ecosystems. Front. Ecol. Environ. 2005, 3, 479–486. [Google Scholar] [CrossRef]
- Ellison, A.M.; Barker-Plotkin, A.A.; Foster, D.R.; Orwig, D.A. Experimentally testing the role of foundation species in forests: The Harvard Forest Hemlock Removal Experiment. Methods Ecol. Evol. 2010, 1, 168–179. [Google Scholar] [CrossRef]
- Orwig, D.A.; Barker-Plotkin, A.A.; Davidson, E.A.; Lux, H.; Savage, K.E.; Ellison, A.M. Foundation species loss affects vegetation structure more than ecosystem function in a northeastern USA forest. Peer J. 2013, 1, e41. [Google Scholar] [CrossRef] [Green Version]
- Orwig, D.A. Ecosystem to regional impacts of introduced pests and pathogens: Historical context, questions and issues. J. Biogeogr. 2002, 29, 1471–1474. [Google Scholar] [CrossRef]
- Ushio, M.; Wagai, R.; Balser, T.C.; Kitayama, K. Variations in the soil microbial community composition of a tropical montane forest ecosystem: Does tree species matter? Soil Biol. Biochem. 2008, 40, 2699–2702. [Google Scholar] [CrossRef]
- Hajabbasi, M.A.; Jalalian, A.; Karimzadeh, H.R. Deforestation effects on soil physical and chemical properties, Lordegan, Iran. Plant Soil 1997, 190, 301–308. [Google Scholar] [CrossRef]
- Zeng, X.; Zhang, W.; Cao, J.; Liu, X.; Shen, H.; Zhao, X. Changes in soil organic carbon, nitrogen, phosphorus, and bulk density after afforestation of the “Beijing–Tianjin Sandstorm Source Control” program in China. Catena 2014, 118, 186–194. [Google Scholar] [CrossRef]
- Black, C.A.; Evans, D.D.; Dinauer, R.C. Methods of Soil Analysis, 1st ed.; American Society of Agronomy: Madison, WI, USA, 1965; p. 768. [Google Scholar]
- Grossman, R.B.; Reinsch, T.G. Bulk density and linear extensibility. In Methods of Soil Analysis: Part 4 Physical Methods; Dane, J.H., Topp, C.G., Eds.; Soil Science Society of America: Madison, WI, USA, 2002; pp. 201–228. [Google Scholar]
- Orwig, D.A.; Cobb, R.C.; D’Amato, A.W.; Kizlinski, M.L.; Foster, D.R. Multi-year ecosystem response to hemlock woolly adelgid infestation in southern New England forests. Can. J. For. Res. 2008, 38, 834–843. [Google Scholar] [CrossRef]
- Kizlinski, M.L.; Orwig, D.A.; Cobb, R.C.; Foster, D.R. Direct and indirect ecosystem consequences of an invasive pest on forests dominated by eastern hemlock. J. Biogeogr. 2002, 29, 1489–1503. [Google Scholar] [CrossRef]
- Jenkins, J.C.; Aber, J.D.; Canham, C.D. Hemlock woolly adelgid impacts on community structure and N cycling rates in eastern hemlock forests. Can. J. For. Res. 1999, 29, 630–645. [Google Scholar] [CrossRef]
- Covington, W.W.; Fule, P.Z.; Moore, M.M.; Hart, S.C.; Kolb, T.E.; Mast, J.N.; Sackett, S.S.; Wagner, M.R. Restoring ecosystem health in ponderosa pine forests of the Southwest. J. For. 1997, 95, 23–29. [Google Scholar]
- Scheller, R.M.; Hua, D.; Bolstad, P.V.; Birdsey, R.A.; Mladenoff, D.J. The effects of forest harvest intensity in combination with wind disturbance on carbon dynamics in Lake States Mesic Forests. Ecol. Model. 2011, 222, 144–153. [Google Scholar] [CrossRef]
- Talbot, J.M.; Finzi, A.C. Differential effects of sugar maple, red oak, and hemlock tannins on carbon and nitrogen cycling in temperate forest soils. Oecologia 2008, 155, 583–592. [Google Scholar] [CrossRef] [PubMed]
- Stadler, B.; Muller, T.; Orwig, D. The Ecology of Energy and Nutrient Fluxes in Hemlock Forests Invaded by Hemlock Woolly Adelgid. Ecology 2006, 87, 1792–1804. [Google Scholar] [CrossRef]
- Wyman, R.L. Experimental assessment of salamanders as predators of detrital food webs: Effects on invertebrates, decomposition and the carbon cycle. Biodivers. Conserv. 1998, 7, 641–650. [Google Scholar] [CrossRef]
- Welsh, H.H.; Droege, S. A case for using plethodontid salamanders for monitoring biodiversity and ecosystem integrity of North American Forests. Conserv. Biol. 2001, 15, 558–569. [Google Scholar] [CrossRef]
- Spotila, J.R. Role of temperature and water in the ecology of lungless salamanders. Ecol. Monogr. 1972, 42, 95–125. [Google Scholar] [CrossRef]
- Feder, M.E. Integrating the ecology and physiology of plethodontid salamanders. Herpetologica 1983, 39, 291–310. [Google Scholar]
- Wyman, R.L.; Jancola, J. Degree and scale of terrestrial acidification and amphibian community structure. J. Herpetol. 1992, 26, 392–401. [Google Scholar] [CrossRef]
- Frisbie, M.P.; Wyman, R.L. The effects of soil pH on sodium balance in the red-backed salamander, plethodon cinereus, and three other terrestrial salamanders. Physiol. Zool. 1991, 64, 1050–1068. [Google Scholar] [CrossRef]
- Cameron, A.C.; Hickerson, C.M.; Anthony, C.D. Plethodon cinereus (Eastern Red-Backed Salamander) Not Affected by Long-Term Exposure to Soil Liming. Northeast. Nat. 2016, 23, 88–99. [Google Scholar] [CrossRef]
- Siddig, A.A.; Ellison, A.M.; Jackson, S. Calibrating abundance indices with population size estimators of red back salamanders (Plethodon cinereus) in a New England forest. Peer J. 2015, 14, e952. [Google Scholar] [CrossRef] [PubMed]
- Pankhurst, C.; Doube, B.M.; Gupta, V. Biological Indicators of Soil Health; Cab International: Wallingford, UK, 1997. [Google Scholar]
- Compton, J.; Boone, R.; Motzkin, G.; Foster, D. Soil carbon and nitrogen in a pine-oak sandplain in central Massachusetts: Role of vegetation and land-use history. Oecologia 1998, 116, 536–542. [Google Scholar] [CrossRef]
- Luo, Y.; Durenkamp, M.; De Nobili, M.; Lin, Q.; Brookes, P.C. Short term soil priming effects and the mineralisation of biochar following its incorporation to soils of different pH. Soil Biol. Biochem. 2011, 43, 2304–2314. [Google Scholar] [CrossRef]
- Amacher, M.C.; O’Neill, K.P.; Dresbach, R.; Palmer, C. Forest Inventory and Analysis: Manual of Soil Analysis Methods; US Forest Service: Washington, D.C., USA, 2003.
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Orwig, D.; Foster, D. Forest response to the introduced hemlock woolly adelgid in southern New England, USA. J. Torrey Bot. Soc. 1998, 125, 60–73. [Google Scholar] [CrossRef]
- Lutz, H.; Chandler, R.J. Forest Soils; John Wiley and Sons, Inc.: New York, NY, USA, 1946; p. 514. [Google Scholar]
- D’Amico, M.E.; Bonifacio, E.; Zanini, E. Relationships between serpentine soils and vegetation in a xeric inner-Alpine environment. Plant Soil 2014, 376, 111–128. [Google Scholar] [CrossRef]
- Welsh, H.H.; Hodgson, G.R. Woodland salamanders as metrics of forest ecosystem recovery: A case study from California’s redwoods. Ecosphere 2013, 4, 1–25. [Google Scholar] [CrossRef]
- Highton, R. Declines of eastern North American woodland salamanders (Plethodon). In Amphibian Declines: The Conservation Status United States Species; University of California Press: Berkeley, CA, USA, 2005; pp. 34–46. [Google Scholar]
- Hocking, D.J.; Connette, G.M.; Conner, C.A.; Scheffers, B.R.; Pittman, S.E.; Peterman, W.E.; Semlitsch, R.D. Effects of experimental forest management on a terrestrial, woodland salamander in Missouri. For. Ecol. Manag. 2013, 287, 32–39. [Google Scholar] [CrossRef]
- Siddig, A.A.H.; Ellison, A.M.; Mathewson, B.G. Assessing and predicting the impacts of the decline of eastern hemlock (Tsuga canadensis) stands on the occurrence of terrestrial amphibians in New England Forest. Ecosphere 2016. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Loadings | PC1 | PC2 | PC3 | PC4 | PC5 | PC6 |
| Moisture | −0.46934 | 0.072315 | −0.49328 | 0.214078 | 0.376623 | 0.586082 |
| Bulk Density | 0.483433 | −0.15782 | 0.259876 | −0.40874 | 0.111261 | 0.703138 |
| Leaf Litter Depth | −0.38316 | 0.157665 | −0.14678 | −0.88009 | 0.062283 | −0.16838 |
| Temperature | 0.308939 | 0.759648 | 0.095788 | 0.049976 | 0.545623 | −0.13459 |
| pH | 0.39307 | 0.334458 | −0.65131 | −0.05968 | −0.54448 | 0.097004 |
| Carbon:Nitrogen | −0.38634 | 0.506041 | 0.483951 | 0.08056 | −0.49773 | 0.325926 |
| Importance of Components | PC1 | PC2 | PC3 | PC4 | PC5 | PC6 |
| Standard deviation | 1.7624 | 1.0285 | 0.8641 | 0.7999 | 0.52311 | 0.41958 |
| Proportion of Variance | 0.5177 | 0.1763 | 0.1244 | 0.1066 | 0.04561 | 0.02934 |
| Cumulative Proportion | 0.5177 | 0.694 | 0.8184 | 0.9251 | 0.97066 | 1 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ochs, A.; Siddig, A.A.H. Response of Red-Backed Salamanders (Plethodon Cinereus) to Changes in Hemlock Forest Soil Driven by Invasive Hemlock Woolly Adelgid (Adelges Tsugae). Environments 2017, 4, 8. https://doi.org/10.3390/environments4010008
Ochs A, Siddig AAH. Response of Red-Backed Salamanders (Plethodon Cinereus) to Changes in Hemlock Forest Soil Driven by Invasive Hemlock Woolly Adelgid (Adelges Tsugae). Environments. 2017; 4(1):8. https://doi.org/10.3390/environments4010008
Chicago/Turabian StyleOchs, Alison, and Ahmed A. H. Siddig. 2017. "Response of Red-Backed Salamanders (Plethodon Cinereus) to Changes in Hemlock Forest Soil Driven by Invasive Hemlock Woolly Adelgid (Adelges Tsugae)" Environments 4, no. 1: 8. https://doi.org/10.3390/environments4010008