Mercury in Juvenile Solea senegalensis: Linking Bioaccumulation, Seafood Safety, and Neuro-Oxidative Responses under Climate Change-Related Stressors

 , , ,
, , ,  and
and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Experimental Feeds
2.2. Organisms and Acclimation
2.3. Experimental Design
2.4. Sampling
2.5. Total Hg and MeHg Determination
2.6. Biochemical Analysis
2.7. Data Analysis
3. Results
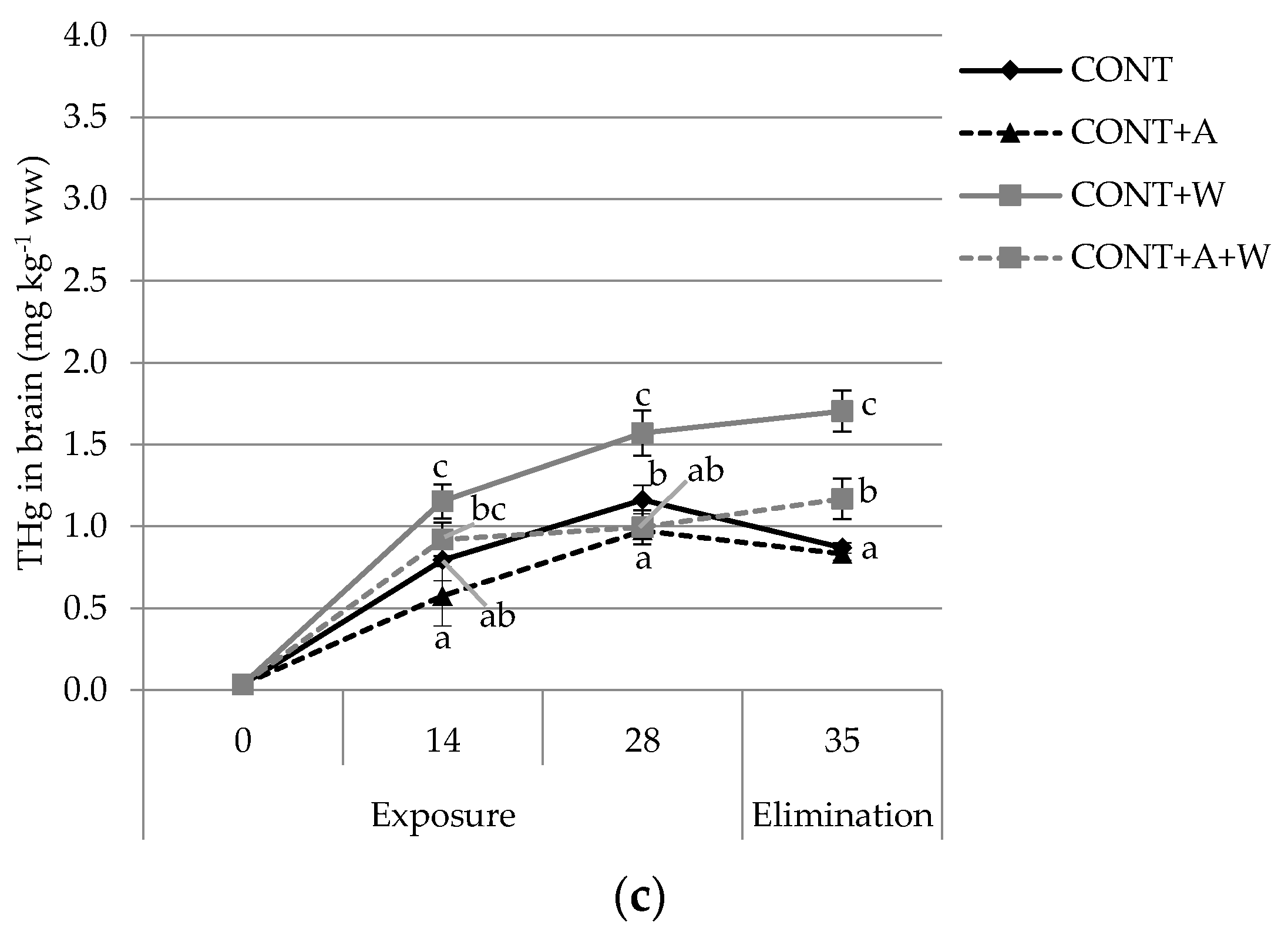
3.1. Mercury Bioaccumulation and Tissue Partioning
3.2. Climate Change-Related Stressors and Potential Impacts on Seafood Safety
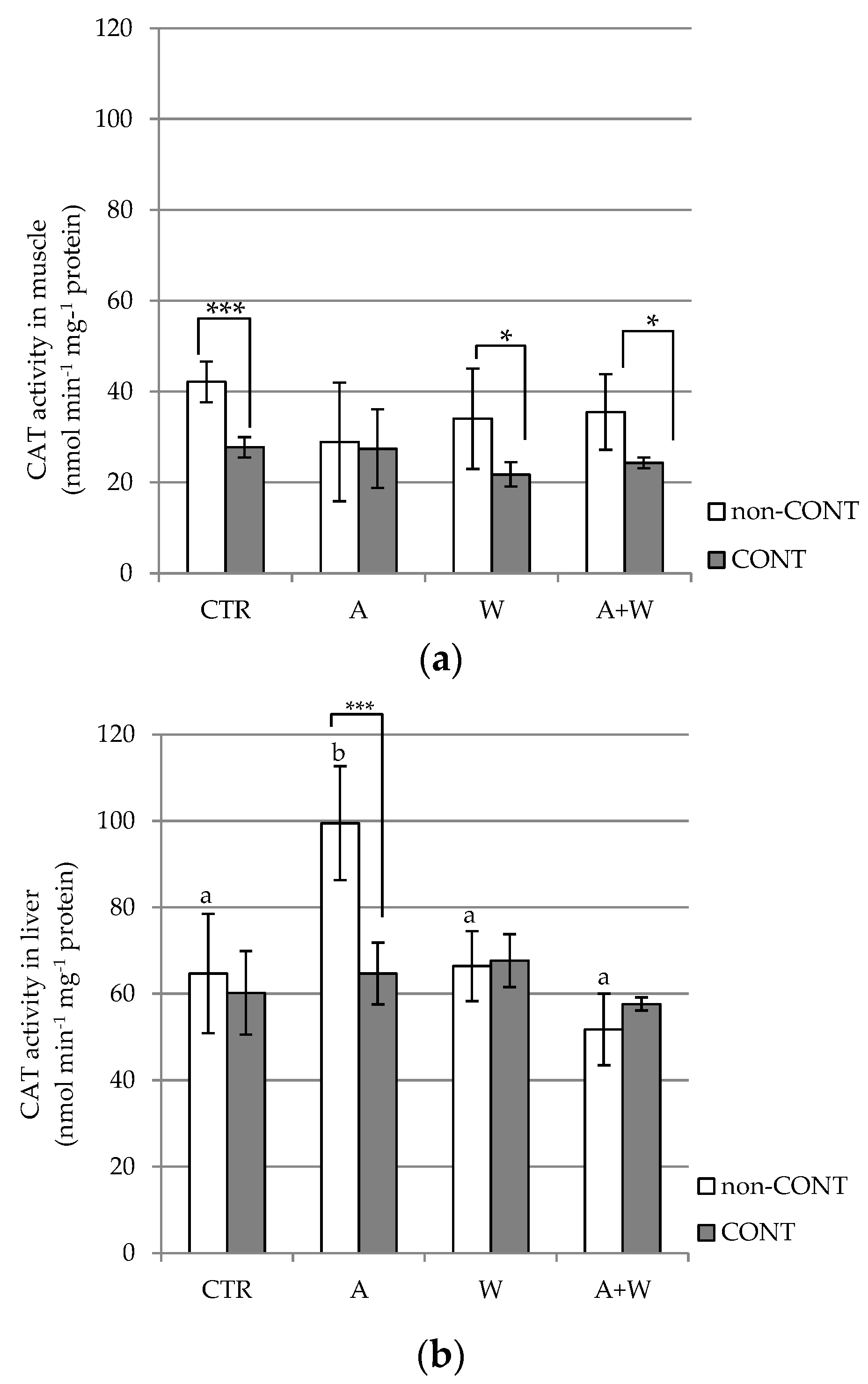
3.3. Animal Condition and Tissue Neuroxidative Responses
4. Discussion
4.1. Mercury Bioaccumulation and Elimination in Tissues with Climate Change
4.2. Linking Hg Contamination, Climate Change Stressors, and Potential Impacts on Seafood Safety
4.3. Interactive Effects of Hg and Climate Change-Related Stressors on Animal Condition and Neuro-Oxidative Responses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Spada, L.; Annicchiarico, C.; Cardellicchio, N.; Giandomenico, S.; Di Leo, A. Mercury and methylmercury concentrations in Mediterranean seafood and surface sediments, intake evaluation and risk for consumers. Int. J. Hyg. Environ. Health 2012, 215, 418–426. [Google Scholar] [CrossRef]
- Maulvault, A.L.; Anacleto, P.; Barbosa, V.; Sloth, J.J.; Rasmussen, R.R.; Tediosi, A.; Fernandez-Tejedor, M.; van den Heuvel, F.H.M.; Kotterman, M.; Marques, A. Toxic elements and speciation in seafood samples from different contaminated sites in Europe. Environ. Res. 2015, 143, 72–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Annibaldi, A.; Truzzi, C.; Carnevali, O.; Pignalosa, P.; Api, M.; Scarponi, G.; Illuminati, S. Determination of Hg in farmed and wild atlantic bluefin tuna (Thunnus thynnus L.) muscle. Molecules 2019, 24, 1273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- EFSA. Panel on Contaminants in the Food Chain (CONTAM). Scientific Opinion on the risk for public health related to the presence of mercury and methylmercury in food. EFSA J. 2012, 10, 2985. [Google Scholar]
- UNEP. Global Mercury Assessment 2013: Sources, Emissions, Releases and Environmental Transport; UNEP: Nairobi, Kenya, 2013. [Google Scholar]
- FAO Fisheries and Aquaculture Department. Cultured Aquatic Species Information Programme Solea spp. Available online: http://www.fao.org/fishery/culturedspecies/Solea_spp/en (accessed on 11 May 2018).
- Kidd, K.; Clayden, M.; Jardine, T. Bioaccumulation and biomagnification of mercury through food webs. In Environmental Chemistry and Toxicology of Mercury; Liu, G., Cai, Y., O’Driscoll, N., Eds.; John Wiley & Sons, Inc: Hoboken, NJ, USA, 2012. [Google Scholar]
- Maulvault, A.L.; Anacleto, P.; Lourenço, H.M.; Carvalho, M.L.; Nunes, M.L.; Marques, A. Nutritional quality and safety of cooked edible crab (Cancer pagurus). Food Chem. 2012, 133, 277–283. [Google Scholar] [CrossRef]
- Newman, M.C.; Xu, X.; Cotton, C.F.; Tom, K.R. High mercury concentrations reflect trophic ecology of three deep-water chondrichthyans. Arch. Environ. Contam. Toxicol. 2011, 60, 618–625. [Google Scholar] [CrossRef]
- Downs, S.G.; MacLeod, C.L.; Lester, J.N. Mercuty in precipitation and its relation to bioaccumulation in fish: A literature review. Water Air Soil Pollut. 1998, 108, 149–187. [Google Scholar] [CrossRef]
- Bradley, M.A.; Barst, B.D.; Basu, N. A review of mercury bioavailability in humans and fish. Int. J. Environ. Res. Public Health 2017, 14, 169. [Google Scholar] [CrossRef] [Green Version]
- Afonso, C.; Lourenço, H.M.; Dias, A.; Nunes, M.L.; Castro, M. Contaminant metals in black scabbard fish (Aphanopus carbo) caught off Madeira and the Azores. Food Chem. 2007, 101, 120–125. [Google Scholar] [CrossRef]
- Maulvault, A.L.; Anacleto, P.; Machado, R.; Amaral, A.; Carvalho, M.L.; Lourenço, H.M.; Nunes, M.L.; Marques, A. Effect of sex, maturation stage and cooking methods on the nutritional quality and safety of black scabbard fish (Aphanopus carbo Lowe, 1839). J. Sci. Food Agric. 2012, 92, 1545–1553. [Google Scholar] [CrossRef]
- Miniero, R.; Beccaloni, E.; Carere, M.; Ubaldi, A.; Mancini, L.; Marchegiani, S.; Cicero, M.R.; Scenati, R.; Lucchetti, D.; Ziemacki, G.; et al. Mercury (Hg) and methyl mercury (MeHg) concentrations in fish from the coastal lagoon of Orbetello, central Italy. Mar. Pollut. Bull. 2013, 76, 365–369. [Google Scholar] [CrossRef] [PubMed]
- Ceccatelli, S.; Bose, R.; Edoff, K.; Onishchenko, N.; Spulber, S. Long-lasting neurotoxic effects of exposure to methylmercury during development. J. Intern. Med. 2013, 273, 490–497. [Google Scholar] [CrossRef] [PubMed]
- Sundseth, K.; Pacyna, J.M.; Pacyna, E.G.; Pirrone, N.; Thorne, R.J. Global sources and pathways of mercury in the context of human health. Int. J. Environ. Res. Public Health 2017, 14, 105. [Google Scholar] [CrossRef] [PubMed]
- Fleeger, J.W.; Carman, K.R.; Nisbet, R.M. Indirect effects of contaminants in aquatic ecosystems. Sci. Total Environ. 2003, 317, 207–233. [Google Scholar] [CrossRef]
- Weis, J.S. Physiological, Developmental and Behavioral Effects of Marine Pollution; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Hammerschmidt, C.R.; Sandheinrich, M.B.; Wiener, J.G.; Rada, R.G. Effects of dietary methylmercury on reproduction of fathead minnows. Environ. Sci. Technol. 2002, 36, 877–883. [Google Scholar] [CrossRef] [PubMed]
- Nunes, M.; Coelho, J.P.; Cardoso, P.G.; Pereira, M.E.; Duarte, A.C.; Pardal, M.A. The macrobenthic community along a mercury contamination in a temperate estuarine system (Ria de Aveiro, Portugal). Sci. Total Environ. 2008, 405, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Berntssen, M.H.G.; Aatland, A.; Handy, R.D. Chronic dietary mercury exposure causes oxidative stress, brain lesions, and altered behaviour in Atlantic salmon (Salmo salar) parr. Aquat. Toxicol. 2003, 65, 55–72. [Google Scholar] [CrossRef]
- Farina, M.; Aschner, M.; Rocha, J.B.T. Oxidative stress in MeHg-induced neurotoxicity. Toxicol. Appl. Pharmacol. 2011, 256, 405–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korbas, M.; MacDonald, T.C.; Pickering, I.J.; George, G.N.; Krone, P.H. Chemical form matters: Differential accumulation of mercury following inorganic and organic mercury exposures in zebrafish larvae. ACS Chem. Biol. 2012, 7, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Maulvault, A.L.; Barbosa, V.; Alves, R.; Custódio, A.; Anacleto, P.; Repolho, T.; Pousão Ferreira, P.; Rosa, R.; Marques, A.; Diniz, M. Ecophysiological responses of juvenile seabass (Dicentrarchus labrax) exposed to increased temperature and dietary methylmercury. Sci. Total Environ. 2017, 586, 551–558. [Google Scholar] [CrossRef]
- Sampaio, E.; Lopes, A.R.; Paula, J.R.; Pimentel, M.; Maulvault, A.L.; Repolho, T.; Grilo, T.F.; Pousão-Ferreira, P.; Marques, A.; Rosa, R. Ocean acidification dampens physiological stress response to warming and contamination in a commercially-important fish (Argyrosomus regius). Sci. Total Environ. 2018, 618, 388–398. [Google Scholar] [CrossRef] [PubMed]
- Sampaio, E.; Maulvault, A.L.; Lopes, V.M.; Paula, J.R.; Barbosa, V.; Alves, R.; Pousão-Ferreira, P.; Repolho, T.; Marques, A.; Rosa, R. Habitat selection disruption and lateralization impairment of cryptic flatfish in a warm, acid, and contaminated ocean. Mar. Biol. 2016, 163, 217. [Google Scholar] [CrossRef]
- Rosa, R.; Lopes, A.R.; Pimentel, M.; Faleiro, F.; Baptista, M.; Trübenbach, K.; Narciso, L.; Dionísio, G.; Pegado, M.R.; Repolho, T. Ocean cleaning stations under a changing climate: Biological responses of tropical and temperate fish-cleaner shrimp to global warming. Glob. Chang. Biol. 2014, 20, 3068–3079. [Google Scholar] [CrossRef] [PubMed]
- Tirado, M.C.; Clarke, R.; Jaykus, L.A.; McQuatters-Gollop, A.; Frank, J.M. Climate change and food safety: A review. Food Res. Int. 2010, 43, 1745–1765. [Google Scholar] [CrossRef]
- IPCC. Summary for Policymakers. In IPCC Special Report on the Ocean and Cryosphere in a Changing Climate; Portner, H.-O., Roberts, D.C., Masson-Delmotte, V., Zhai, P., Tignor, M., Poloczanska, E., Mintenbeck, K., Nicolai, M., Okem, A., Petzold, J., et al., Eds.; IPCC, 2019. Available online: https://report.ipcc.ch/srocc/pdf/SROCC_FinalDraft_FullReport.pdf (accessed on 5 March 2020).
- Gunderson, A.R.; Armstrong, E.J.; Stillman, J.H. Multiple stressors in a changing world: The need for an improved perspective on physiological responses to the dynamic marine environment. Ann. Rev. Mar. Sci. 2016, 8, 357–378. [Google Scholar] [CrossRef]
- Alava, J.J.; Cisneros-Montemayor, A.M.; Sumaila, U.R.; Cheung, W.W.L. Projected amplification of food web bioaccumulation of MeHg and PCBs under climate change in the Northeastern Pacific. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef]
- Booth, S.; Zeller, D. Mercury, food webs, and marine mammals: Implications of diet and climate change for human health. Environ. Health Perspect. 2005, 113, 521–526. [Google Scholar] [CrossRef]
- Marques, A.; Nunes, M.L.; Moore, S.K.; Strom, M.S. Climate change and seafood safety: Human health implications. Food Res. Int. 2010, 43, 1766–1779. [Google Scholar] [CrossRef]
- Rosa, R.; Pimentel, M.S.; Boavida-Portugal, J.; Teixeira, T.; Trübenbach, K.; Diniz, M. Ocean warming enhances malformations, premature hatching, metabolic suppression and oxidative stress in the early life stages of a keystone squid. PLoS ONE 2012, 7, e38282. [Google Scholar] [CrossRef]
- Rosa, R.; Baptista, M.; Lopes, V.M.; Pegado, M.R.; Ricardo Paula, J.; Trübenbach, K.; Leal, M.C.; Calado, R.; Repolho, T. Early-life exposure to climate change impairs tropical shark survival. Proc. R. Soc. B Biol. Sci. 2014, 281, 20141738. [Google Scholar] [CrossRef] [Green Version]
- Rosa, R.; Trubenbach, K.; Pimentel, M.S.; Boavida-Portugal, J.; Faleiro, F.; Baptista, M.; Dionisio, G.; Calado, R.; Portner, H.O.; Repolho, T. Differential impacts of ocean acidification and warming on winter and summer progeny of a coastal squid (Loligo vulgaris). J. Exp. Biol. 2014, 217, 518–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alava, J.J.; Cheung, W.W.L.; Ross, P.S.; Sumaila, U.R. Climate change–contaminant interactions in marine food webs: Toward a conceptual framework. Glob. Chang. Biol. 2017, 23, 3984–4001. [Google Scholar] [CrossRef] [PubMed]
- Alava, J.J. Climate Change and Overfishing are Boosting Toxic Mercury Levels in Fish. Available online: https://theconversation.com/climate-change-and-overfishing-are-boosting-toxic-mercury-levels-in-fish-122748 (accessed on 18 February 2020).
- Siscar, R.; Torreblanca, A.; Palanques, A.; Solé, M. Metal concentrations and detoxification mechanisms in Solea solea and Solea senegalensis from NW Mediterranean fishing grounds. Mar. Pollut. Bull. 2013, 77, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Frapiccini, E.; Annibaldi, A.; Betti, M.; Polidori, P.; Truzzi, C.; Marini, M. Polycyclic aromatic hydrocarbon (PAH) accumulation in different common sole (Solea solea) tissues from the North Adriatic Sea peculiar impacted area. Mar. Pollut. Bull. 2018, 137, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Cheung, W.W.L.; Sarmiento, J.L.; Dunne, J.; Frölicher, T.L.; Lam, V.W.Y.; Palomares, M.L.D.; Watson, R.; Pauly, D. Shrinking of fishes exacerbates impacts of global ocean changes on marine ecosystems. Nat. Clim. Chang. 2013, 3, 254–258. [Google Scholar] [CrossRef]
- Gagné, F. Biochemical Ecotoxicology: Principles and Methods; Academic Press: Cambridge, MA, USA, 2014. [Google Scholar]
- Maulvault, A.L.; Custódio, A.; Anacleto, P.; Repolho, T.; Pousão, P.; Nunes, M.L.; Diniz, M.; Rosa, R.; Marques, A. Bioaccumulation and elimination of mercury in juvenile seabass (Dicentrarchus labrax) in a warmer environment. Environ. Res. 2016, 149, 77–85. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Ellman, G.L.; Courtney, K.D.; Andres, V., Jr.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Dizer, H.; da Silva de Assis, H.C.; Hansen, P.-D. Cholinesterase activity as a bioindicator for monitoring marine pollution in the Baltic Sea and the Mediterranean Sea. In Biomarkers in Marine Organisms; Elsevier: Amsterdam, The Netherlands, 2001. [Google Scholar]
- Johansson, L.H.; Borg, L. A spectrophotometric method for determination of catalase activity in small tissue samples. Anal. Biochem. 1988, 174, 331–336. [Google Scholar] [CrossRef]
- Sun, Y.; Oberley, L.W.; Li, Y. A simple method for clinical assay of superoxide dismutase. Clin. Chem. 1988, 34, 497–500. [Google Scholar] [CrossRef]
- Mihara, M.; Uchiyama, M. Determination of malonaldehyde precursor in tissues by thiobarbituric acid test. Anal. Biochem. 1978, 86, 271–278. [Google Scholar]
- Ricker, W.E. Computation and Interpretation of Biological Statistics of Fish Populations; Bulletin of the Fisheries Research of Canada: Ottawa, ON, Canada, 1975. [Google Scholar]
- Santana, L.M.B.M.; Blasco, J.; Abessa, D.M.S.; Campana, O. Bioaccumulation kinetics of copper in Ruditapes philippinarum exposed to increasing, continuous and pulsed exposure: Implications for growth. Sci. Total Environ. 2017, 595, 920–927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jebali, J.; Chouba, L.; Banni, M.; Boussetta, H. Comparative study of the bioaccumulation and elimination of trace metals (Cd, Pb, Zn, Mn and Fe) in the digestive gland, gills and muscle of bivalve Pinna nobilis during a field transplant experiment. J. Trace Elem. Med. Biol. 2014, 28, 212–217. [Google Scholar] [CrossRef] [PubMed]
- EPA. Guidance of Assessing Chemical Contaminant Data for Use in Fish Advisories; Office of Science and Technology and Office of Water: Washington, WA, USA, 2000.
- FAO. FAO Yearbook. Fishery and Aquaculture Statistics 2017; FAO: Rome, Italy, 2019. [Google Scholar]
- Storelli, M.M.; Normanno, G.; Barone, G.; Dambrosio, A.; Errico, L.; Garofalo, R.; Giacominelli-Stuffler, R. Toxic metals (Hg, Cd, and Pb) in fishery products imported into Italy: Suitability for human consumption. J. Food Prot. 2012, 75, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Bentzen, R.; Castellini, J.M.; Gerlach, R.; Dykstra, C.; O’Hara, T. Mercury concentrations in Alaska Pacific halibut muscle relative to stable isotopes of C and N and other biological variables. Mar. Pollut. Bull. 2016, 113, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Ruus, A.; Hjermann, D.Ø.; Beylich, B.; Schøyen, M.; Øxnevad, S.; Green, N.W. Mercury concentration trend as a possible result of changes in cod population demography. Mar. Environ. Res. 2017, 130, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Frantzen, S.; Maage, A.; Duinker, A.; Julshamn, K.; Iversen, S.A. A baseline study of metals in herring (Clupea harengus) from the Norwegian Sea, with focus on mercury, cadmium, arsenic and lead. Chemosphere 2015, 127, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Chaguri, M.P.; Maulvault, A.L.; Costa, S.; Gonçalves, A.; Nunes, M.L.; Carvalho, M.L.; Sant’ana, L.S.; Bandarra, N.; Marques, A. Chemometrics tools to distinguish wild and farmed meagre (Argyrosomus regius). J. Food Process. Preserv. 2017, 41, e13312. [Google Scholar] [CrossRef]
- Mieiro, C.L.; Pacheco, M.; Duarte, A.C.; Pereira, M.E. Fish consumption and risk of contamination by mercury–Considerations on the definition of edible parts based on the case study of European sea bass. Mar. Pollut. Bull. 2011, 62, 2850–2853. [Google Scholar] [CrossRef]
- Torres-Escribano, S.; Ruiz, A.; Barrios, L.; Vélez, D.; Montoro, R. Influence of mercury bioaccessibility on exposure assessment associated with consumption of cooked predatory fish in Spain. J. Sci. Food Agric. 2011, 91, 981–986. [Google Scholar] [CrossRef]
- Costa, F.; Coelho, J.P.; Baptista, J.; Martinho, F.; Pereira, M.E.; Pardal, M.A. Mercury accumulation in fish species along the Portuguese coast: Are there potential risks to human health? Mar. Pollut. Bull. 2020, 150, 110740. [Google Scholar] [CrossRef] [PubMed]
- EPA, United States Environmental Protection Agency. An Overview of Methods for EPA’s National-Scale Air Toxics Assessment. Available online: https://www.epa.gov/sites/production/files/2015-10/documents/2005-nata-tmd.pdf (accessed on 20 December 2019).
- Wang, R.; Feng, X.-B.; Wang, W.-X. In vivo mercury methylation and demethylation in freshwater tilapia quantified by mercury stable isotopes. Environ. Sci. Technol. 2013, 47, 7949–7957. [Google Scholar] [CrossRef] [PubMed]
- Gentès, S.; Maury-Brachet, R.; Feng, C.; Pedrero, Z.; Tessier, E.; Legeay, A.; Mesmer-Dudons, N.; Baudrimont, M.; Maurice, L.; Amouroux, D.; et al. Specific Effects of Dietary Methylmercury and Inorganic Mercury in Zebrafish (Danio rerio) Determined by Genetic, Histological, and Metallothionein Responses. Environ. Sci. Technol. 2015, 49, 14560–14569. [Google Scholar] [CrossRef] [PubMed]
- Amlund, H.; Lundebye, A.K.; Berntssen, M.H.G. Accumulation and elimination of methylmercury in Atlantic cod (Gadus morhua L.) following dietary exposure. Aquat. Toxicol. 2007, 83, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Bosch, A.C.; O’Neill, B.; Sigge, G.O.; Kerwath, S.E.; Hoffman, L.C. Heavy metals in marine fish meat and consumer health: A review. J. Sci. Food Agric. 2016, 96, 32–48. [Google Scholar] [CrossRef]
- Johnston, I.A.; Dunn, J. Temperature acclimation and metabolism in ectotherms with particular reference to teleost fish. Proc. Symp. Soc. Exp. Biol. 1987, 41, 67–93. [Google Scholar]
- Flynn, E.E.; Bjelde, B.E.; Miller, N.A.; Todgham, A.E. Ocean acidification exerts negative effects during warming conditions in a developing Antarctic fish. Conserv. Physiol. 2015, 3, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Dijkstra, J.A.; Buckman, K.L.; Ward, D.; Evans, D.W.; Dionne, M.; Chen, C.Y. Experimental and Natural Warming Elevates Mercury Concentrations in Estuarine Fish. PLoS ONE 2013, 8, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Wang, W.-X.; Wang, M. Alleviation of mercury toxicity to a marine copepod under multigenerational exposure by ocean acidification. Sci. Rep. 2017, 7, 324. [Google Scholar] [CrossRef] [Green Version]
- Doney, S.C.; Ruckelshaus, M.; Emmett Duffy, J.; Barry, J.P.; Chan, F.; English, C.A.; Galindo, H.M.; Grebmeier, J.M.; Hollowed, A.B.; Knowlton, N.; et al. Climate Change Impacts on Marine Ecosystems. Ann. Rev. Mar. Sci. 2012, 4, 11–37. [Google Scholar] [CrossRef] [Green Version]
- Melzner, F.; Gutowska, M.A.; Langenbuch, M.; Dupont, S.; Lucassen, M.; Thorndyke, M.C.; Bleich, M.; Pörtner, H.-O. Physiological basis for high CO2 tolerance in marine ectothermic animals: Pre-adaptation through lifestyle and ontogeny? Biogeosciences 2009, 6, 2313–2331. [Google Scholar] [CrossRef] [Green Version]
- Heuer, R.M.; Grosell, M. Physiological impacts of elevated carbon dioxide and ocean acidification on fish. Am. J. Physiol. Integr. Comp. Physiol. 2014, 307, R1061–R1084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiedek, D.; Sundelin, B.; Readman, J.W.; Macdonald, R.W. Interactions between climate change and contaminants. Mar. Pollut. Bull. 2007, 54, 1845–1856. [Google Scholar] [CrossRef] [PubMed]
- Byrne, M.; Przeslawski, R. Multistressor impacts of warming and acidification of the ocean on marine invertebrates’ life histories. Integr. Comp. Biol. 2013, 53, 582–596. [Google Scholar] [CrossRef] [Green Version]
- Guzzi, G.; La Porta, C.A.M. Molecular mechanisms triggered by mercury. Toxicology 2008, 244, 1–12. [Google Scholar] [CrossRef]
- Eagles-Smith, C.A.; Ackerman, J.T.; Yee, J.; Adelsbach, T.L. Mercury demethylation in waterbird livers: Dose–response thresholds and differences among species. Environ. Toxicol. Chem. Int. J. 2009, 28, 568–577. [Google Scholar] [CrossRef]
- Maulvault, A.L.; Camacho, C.; Barbosa, V.; Alves, R.; Anacleto, P.; Fogaça, F.; Kwadijk, C.; Kotterman, M.; Cunha, S.C.; Fernandes, J.O.; et al. Assessing the effects of seawater temperature and pH on the bioaccumulation of emerging chemical contaminants in marine bivalves. Environ. Res. 2018, 161, 236–247. [Google Scholar] [CrossRef]
- Maulvault, A.L.; Barbosa, V.; Alves, R.; Anacleto, P.; Camacho, C.; Cunha, S.; Fernandes, J.O.; Ferreira, P.P.; Rosa, R.; Marques, A.; et al. Integrated multi-biomarker responses of juvenile seabass to diclofenac, warming and acidification co-exposure. Aquat. Toxicol. 2018, 202, 65–79. [Google Scholar] [CrossRef]
- Maulvault, A.L.; Anacleto, P.; Marques, A.; Diniz, M.; Rosa, R. Chemical contaminants in a changing ocean. In Ecotoxicology of Marine Organisms; Duarte, B., Caçador, I., Eds.; CRC Press: Boca Raton, FL, USA, 2019. [Google Scholar]
- Stauber, J.L.; Chariton, A.; Apte, S. Global Change. In Marine Ecotoxicology: Current Knowledge and Future Issues; Blasco, J., Chapman, P.M., Campana, O., Hampel, M., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2016. [Google Scholar]
- Madeira, D.; Narciso, L.; Cabral, H.N.; Vinagre, C.; Diniz, M.S. Influence of temperature in thermal and oxidative stress responses in estuarine fish. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2013, 166, 237–243. [Google Scholar] [CrossRef]
- Puga, S.; Pereira, P.; Pinto-Ribeiro, F.; O’Driscoll, N.J.; Mann, E.; Barata, M.; Pousão-Ferreira, P.; Canário, J.; Almeida, A.; Pacheco, M. Unveiling the neurotoxicity of methylmercury in fish (Diplodus sargus) through a regional morphometric analysis of brain and swimming behavior assessment. Aquat. Toxicol. 2016, 180, 320–333. [Google Scholar] [CrossRef]
- Rosa, R.; Paula, J.R.; Sampaio, E.; Pimentel, M.; Lopes, A.R.; Baptista, M.; Guerreiro, M.; Santos, C.; Campos, D.; Almeida-Val, V.M.F.; et al. Neuro-oxidative damage and aerobic potential loss of sharks under elevated CO2 and warming. Mar. Biol. 2016, 163, 119. [Google Scholar] [CrossRef]
- Schmidel, A.J.; Assmann, K.L.; Werlang, C.C.; Bertoncello, K.T.; Francescon, F.; Rambo, C.L.; Beltrame, G.M.; Calegari, D.; Batista, C.B.; Blaser, R.E. Subchronic atrazine exposure changes defensive behaviour profile and disrupts brain acetylcholinesterase activity of zebrafish. Neurotoxicol. Teratol. 2014, 44, 62–69. [Google Scholar] [CrossRef]
- Topal, A.; Alak, G.; Ozkaraca, M.; Yeltekin, A.C.; Comaklı, S.; Acıl, G.; Kokturk, M.; Atamanalp, M. Neurotoxic responses in brain tissues of rainbow trout exposed to imidacloprid pesticide: Assessment of 8-hydroxy-2-deoxyguanosine activity, oxidative stress and acetylcholinesterase activity. Chemosphere 2017, 175, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.J.; Yang, L.; Zhao, Q.; Caen, J.P.; He, H.Y.; Jin, Q.H.; Guo, L.H.; Alemany, M.; Zhang, L.Y.; Shi, Y.F. Induction of acetylcholinesterase expression during apoptosis in various cell types. Cell Death Differ. 2002, 9, 790–800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Rey, M.; Bebianno, M.J. Effects of non-steroidal anti-inflammatory drug (NSAID) diclofenac exposure in mussel Mytilus galloprovincialis. Aquat. Toxicol. 2014, 148, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Lionetto, M.G.; Caricato, R.; Calisi, A.; Giordano, M.E.; Schettino, T. Acetylcholinesterase as a biomarker in environmental and occupational medicine: New insights and future perspectives. Biomed Res. Int. 2013, 2013, 321213. [Google Scholar] [CrossRef]
- Van der Ven, K.; Keil, D.; Moens, L.N.; Van Hummelen, P.; van Remortel, P.; Maras, M.; De Coen, W. Effects of the antidepressant mianserin in zebrafish: Molecular markers of endocrine disruption. Chemosphere 2006, 65, 1836–1845. [Google Scholar] [CrossRef]
- Oliveira, L.D.; Antunes, S.C.; Gonçalves, F.; Rocha, O.; Nunes, B. Evaluation of ecotoxicological effects of drugs on Daphnia magna using different enzymatic biomarkers. Ecotoxicol. Environ. Saf. 2015, 119, 123–131. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tissue | Treatment | NAR during Exposure | EF during Clearance |
|---|---|---|---|
| Muscle | CONT | 53.1 ± 5.8 b | ≤0 |
| CONT + A | 26.5 ± 3.6 a | ≤0 | |
| CONT + W | 73.3 ± 6.3 c | ≤0 | |
| CONT + A + W | 68.0 ± 2.5 c | ≤0 | |
| Liver | CONT | 124.4 ± 8.4 b | 44.0 ± 1.2 b |
| CONT + A | 87.6 ± 4.4 a | 56.8 ± 1.9 c | |
| CONT + W | 150.6 ± 5.5 c | 19.6 ± 4.3 a | |
| CONT + A + W | 118.7 ± 4.4 b | 39.7 ± 4.7 b | |
| Brain | CONT | 40.4 ± 3.3 a | ≤0 |
| CONT + A | 33.6 ± 2.9 a | ≤0 | |
| CONT + W | 54.9 ± 5.0 b | ≤0 | |
| CONT + A + W | 34.3 ± 3.8 a | ≤0 |
| Exposure | S. senegalensis1 | Argyrososmus regius2 | Dicentrachus labrax3 |
|---|---|---|---|
| High Hg contamination + acidification exposure | −47.3% | 0% | n.d. |
| High Hg contamination + warming exposure | 35.9% | 40% | 20.6% |
| High Hg contamination + acidification and warming exposure | 26.4% | 0% | n.d. |
| Species | Hg (µg∙g−1 ww) | Reference | Hazard Quotient (HQ) | |||
|---|---|---|---|---|---|---|
| Actual | W 1 | A 2 | W + A 3 | |||
| Demersal 4 | ||||||
| Sole | 0.15 | [55] | 0.31 | 0.42 | 0.16 | 0.39 |
| Flounder | 0.18 6 | [2] | 0.37 | 0.50 | 0.19 | 0.47 |
| Mullet | 0.04 6 | [2] | 0.07 | 0.10 | 0.04 | 0.09 |
| Halibut | 0.32 | [56] | 0.66 | 0.89 | 0.34 | 0.83 |
| Atlantic cod | 0.20 | [57] | 0.41 | 0.56 | 0.21 | 0.52 |
| Herring | 0.04 | [58] | 0.09 | 0.12 | 0.05 | 0.11 |
| Pelagic 5 | ||||||
| Meagre | 0.12 6 | [59] | 0.24 | 0.33 | 0.24 | 0.24 |
| Seabass | 0.04 | [60] | 0.08 | 0.11 | 0.08 | 0.08 |
| Black scabbard fish | 0.9 | [12] | 1.85 | 2.59 | 1.85 | 1.85 |
| Bluefin tuna | 1.68 | [3] | 3.45 | 4.83 | 3.45 | 3.45 |
| Fresh tuna | 0.94 | [61] | 1.92 | 2.69 | 1.92 | 1.92 |
| Tope shark | 1.13 | [61] | 2.31 | 3.24 | 2.31 | 2.31 |
| Atlantic mackerel | 0.04 | [62] | 0.09 | 0.12 | 0.09 | 0.09 |
| Treatments | W | TL | K |
|---|---|---|---|
| Non-CONT | 10.1 ± 1.3 b | 9.8 ± 0.5 b | 1.08 ± 0.06 |
| Non-CONT + A | 7.3 ± 2.4 a,b | 8.8 ± 1.1 a,b | 1.07 ± 0.12 |
| Non-CONT + W | 8.7 ± 2.0 a,b | 9.3 ± 0.6 a,b | 1.07 ± 0.09 |
| Non-CONT + A + W | 8.9 ± 1.1 a,b | 9.0 ± 0.6 a,b | 1.25 ± 0.25 |
| CONT | 5.9 ± 1.3 a | 8.3 ± 0.6 a | 1.01 ± 0.10 |
| CONT + A | 7.7 ± 2.8 a,b | 8.9 ± 0.8 a,b | 1.07 ± 0.10 |
| CONT + W | 7.1 ± 0.52 a,b | 8.7 ± 1.3 a,b | 1.04 ± 0.19 |
| CONT + A + W | 7.5 ± 1.1 a,b | 8.9 ± 0.4 a,b | 1.07 ± 0.12 |
| Variables (X, Y) | r and associated p-value | ||||
| TL × W | r = 0.840 | p < 0.001 | |||
| K × W | n.s. | ||||
| K × TL | r = −0.432 | p < 0.001 | |||
| Muscle | Liver | Brain | |||
| NAR × W | n.s. | n.s. | ns | ||
| NAR × TL | n.s. | n.s. | ns | ||
| NAR × K | n.s. | n.s. | ns | ||
| NAR × CAT | r = −0.4354 | p = 0.033 | n.s. | r = −0.5073 | p = 0.011 |
| NAR × SOD | n.s. | n.s. | r = −0.4411 | p = 0.031 | |
| NAR × LPO | n.s. | n.s. | r = −0.4354 | p = 0.033 | |
| NAR × AchE | - | - | - | r = −0.4639 | p = 0.022 |
| EF × W | - | - | n.s. | - | - |
| EF × TL | - | - | n.s. | - | - |
| EF × K | - | - | n.s. | - | - |
| EF × CAT | - | - | n.s. | - | - |
| EF × SOD | - | - | n.s. | - | - |
| EF × LPO | - | - | n.s. | - | - |
| K × CAT | n.s. | n.s. | n.s. | ||
| K × SOD | n.s. | n.s. | n.s. | ||
| K × LPO | n.s. | n.s. | n.s. | ||
| K × AchE | n.s. | n.s. | n.s. | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Camacho, C.; Maulvault, A.L.; Santos, M.T.; Barbosa, V.; Fogaça, F.H.S.; Pousão-Ferreira, P.; Nunes, M.L.; Rosa, R.; Marques, A. Mercury in Juvenile Solea senegalensis: Linking Bioaccumulation, Seafood Safety, and Neuro-Oxidative Responses under Climate Change-Related Stressors. Appl. Sci. 2020, 10, 1993. https://doi.org/10.3390/app10061993
Camacho C, Maulvault AL, Santos MT, Barbosa V, Fogaça FHS, Pousão-Ferreira P, Nunes ML, Rosa R, Marques A. Mercury in Juvenile Solea senegalensis: Linking Bioaccumulation, Seafood Safety, and Neuro-Oxidative Responses under Climate Change-Related Stressors. Applied Sciences. 2020; 10(6):1993. https://doi.org/10.3390/app10061993
Chicago/Turabian StyleCamacho, Carolina, Ana Luísa Maulvault, Marta T. Santos, Vera Barbosa, Fabíola H. S. Fogaça, Pedro Pousão-Ferreira, M. Leonor Nunes, Rui Rosa, and António Marques. 2020. "Mercury in Juvenile Solea senegalensis: Linking Bioaccumulation, Seafood Safety, and Neuro-Oxidative Responses under Climate Change-Related Stressors" Applied Sciences 10, no. 6: 1993. https://doi.org/10.3390/app10061993