
Antidiabetic, Anticholesterol, and Antioxidant Activity of Gryllusbimaculatus Fermented by Bacillus and Lactobacillus Strains


Department of Food Science and Nutrition, Dankook University, Cheonan 31116, Korea
*
Author to whom correspondence should be addressed.
Appl. Sci. 2021, 11(5), 2090; https://doi.org/10.3390/app11052090
Submission received: 2 February 2021
/
Revised: 21 February 2021
/
Accepted: 23 February 2021
/
Published: 26 February 2021
(This article belongs to the Section Food Science and Technology)
Abstract
:In this study, functionality of Gryllus bimaculatus (GB) fermented by Bacillus and Lactobacillus strain was investigated. GB was fermented by each of the following strains: probiotic Bacillus amyloliquefaciens MKSK-J1 (SKGB), probiotic Lactobacillus plantarum MKHA15 (HAGB), Bacillus amyloliquefaciens MKSE (SEGB), and Lactobacillus plantarum KCTC 3103 (LPGB). Fermentation was carried out at 35 °C for 24 h. In HAGB, complete inhibition of α-glucosidase and 3-hydroxy-methyl glutaryl-coenzyme A (HMG-CoA) reductase occurred (101.94% and 120.89%, respectively), and superoxide dismutase (SOD)-like activity (IC50) was significantly low (22.37 mg/mL). After in vitro digestion, SOD-like activity was the highest in HAGB (21.18%). In SKGB, reducing power (EC50) was significantly low (0.29 mg/mL). After in vitro digestion, the reducing was also highest in SKGB (86.06%). Fermentation enhanced the bioactivity of GB; in particular, MKHA15 was found to have great potential as a starter in the production of fermented GB, as it would offer multi-health functions, including antidiabetic, anticholesterol, and antioxidant activity.
1. Introduction
According to the Food and Agriculture Organization of the United Nations (FAO, Rome, Italy), the world’s population will surpass 9 billion in 2025, which will result in a food crisis. Edible insects have attracted attention as an important alternative food source to overcome the scarcity of food sources and limitations of animal protein sources such as livestock. Insects can grow sufficiently with a small amount of feed and water and can grow by consuming plant leaves and organic by-products; therefore, insects are a low-cost and high-protein food source compared to livestock [1]. Moreover, insects are a valuable source of amino acids, protein, vital lipids, and micronutrients [2]. Therefore, research on the excellent nutritional value of insects as a new biological resource is actively being conducted and is expected to be used in various fields such as alternative food and new biological resources [3]. Almost 2000 species of insects are known to be edible [4], and the representative examples are Tenebrio molitor, Gryllodes sigillatus, Allomyrina dichotoma, and Gryllus bimaculatus [5,6]. Among them, Gryllus bimaculatus (GB) is particularly high in unsaturated fatty acids and protein. Research on various edible insects has been carried out with the aim to develop health supplements [6].
According to the World Health Organization (WHO), the number of diabetic patients worldwide more than doubled from 180 million in 1980 to 422 million in 2014 [7]. One of the current treatments for Type 2 diabetes mellitus (T2DM) is the inhibition of carbohydrate-hydrolyzing enzymes, including α-amylase and α-glucosidase. By inhibiting these enzymes, glucose absorption can be delayed, and in this way, plasma glucose levels can be reduced [8]. Likewise, in most hyperlipidemia treatments, statin competitively inhibits 3-hydroxy-methyl glutaryl-coenzyme A (HMG-CoA) reductase, an important rate-limiting enzyme in the cholesterol biosynthesis process, which ultimately reduces blood cholesterol levels [9]. Long-term treatment is needed to improve the prognosis for chronic diseases, but side-effects such as liver damage, abdominal distention, diarrhea, and muscle weakness have been reported due to long-term use of drugs such as acarbose and statins. Therefore, this has increased the demand for novel enzyme inhibitors of non-toxic natural materials [10,11].
Probiotics are defined as “Live microorganisms which, when administered in adequate amounts, confer a health benefit to the host” [12], and are generally categorized as lactic acid bacteria (LAB), spore-forming Bacillus spp., or yeast. In particular, LAB and Bacillus spp. can be utilized as starter culture during the fermentation process. Their metabolites such as organic acids and peptides exhibit diverse physiological activities, such as antioxidant activity and improved nutrient utilization, as well as sensory improvements [1,13]. Research on fermented foods using different starters has been conducted with the aim to indicate various health functional properties [14,15]. MKHA15 was isolated from kimchi [16], and MKSK-J1 was isolated from ganjang [17]. Both probiotic MKHA 15 and MKSK-J1 have tolerance to acid and bile salt and are safe to use. In addition, MKHA15 has functional properties, such as antioxidant, antidiabetic activity, and anticholesterol activity, whereas MKSK-J1 has amylase and protease activity.
The main objective of this study was to compare the health functionality factors of fermented GB using the probiotics MKHA15 and MKSK-J1, which were isolated from fermented food in previous studies [18,19], Lactobacillus plantarum KCTC 3103, and non-probiotic Bacillus amyloliquefaciens MKSE. Ultimately, the development of functional fermented GBs that possess antidiabetic, anticholesterol, and antioxidant activities was intended.
2. Materials and Methods
2.1. Raw Materials and Chemicals
Dried GB was purchased from Cricket Farm (Korea). The starters used in this study, L. plantarum MKHA15 (MKHA15), B. amyloliquefaciens MKSK-J1 (MKSK-J1), and B. amyloliquefaciens MKSE (MKSE) were already possessed by our laboratory, and L. plantarum KCTC 3103 was distributed from KCTC. These strains were cultured in De Man, Rogosa, and Sharp (MRS) broth and nutrient broth (NB) medium. Diphenyl-1-picrylhydrazyl (DPPH), pyrogallol, 2,4,6-tris(2-pridyl)-s-triazine, bile salt, α-glucosidase from Bacillus stearothermophilus, 2,2-azino-bis (3-ethylbenzothiazoline-6-sulphonic acid), and an HMG-CoA reductase assay kit were all purchased from Sigma-Aldrich (St. Louis, MO, USA). Ascorbic acid was purchased from Duksan Pure Chemicals Co., Ltd. (Ansan, Korea). All materials were used in the original purchased form.
2.2. Fermentation
MKHA15 (KCTC 13928BP) and L. plantarum KCTC 3103 were grown in MRS, and MKSK-J1 (KCTC 18452P) and MKSE (KCTC 18753P) were grown in NB at 35 °C for 24 h before inoculation. Next, 100 g dried GB powder was mixed with 2.5% (w/v) glucose, and distilled water was added so that the total solid and liquid ratio was 1:5. Then, the mixture was autoclaved at 121 °C and 15 psi for 20 min. After cooling to 35 °C, 5% (v/v) starter culture was inoculated (Bacillus amyloliquefaciens MKSK-J1 (SKGB), Bacillus amyloliquefaciens MKSE (SEGB), Lactobacillus plantarum MKHA15 (HAGB), L. plantarum KCTC 3103 (LPGB), and no starter—CON). The mixture was incubated at 35 °C and shaken at 130 rpm for 24 h. Thereafter, the fermented product was centrifuged at 4000× g for 5 min (Beckman Coulter, Inc., Brea, CA, USA), and the supernatant was freeze-dried at −80 °C (Ilshin Lab Co., Ltd., Seoul, Korea). The supernatant and freeze-dried powder of fermented products were stored at −20 °C until use.
2.3. Microbiological Analysis
The sample (0.1 mL) was mixed thoroughly with 0.9 mL 0.1% peptone water, and serial dilutions ranging from 10−1 to 10−7 were performed using peptone water as the diluents. Each diluted sample was spread on MRS agar and nutrient agar (NA), and incubated at 35 °C for 24 h. The visible colonies were counted for viable cell counts (colony forming unit (CFU)/mL).
2.4. pH Measurement
The sample was diluted 10 times using distilled water, and the pH was measured with a pH meter (Fisher Science Education, Waltham, MA, USA).
2.5. In Vitro Digestion
In vitro digestion of fermented GB was produced using the modified method by Li et al. (2019) [18]. The first step was the oral phase, in which 5 mL of sample was incubated at 37 °C for 10 min in the presence of artificial saliva (2.38 g/L Na2HPO4, 0.19 g/L KH2PO4, and 8 g/L NaCl in 0.1 mol/L phosphate buffer (pH 6.9)) containing α-amylase (75 units/mL). After the pH was adjusted to 1.5 by adding 1.0 mol/L HCl, the mixture was incubated under gastric conditions. For this, digesta was mixed with 15 mL of an artificial gastric solution (3.2 g/L pepsin and 17.55 g/L NaCl adjusted to pH 1.2 with 1 mol/L HCl in distilled water) and incubated at 37 °C for 2 h in shaking water bath at 120 rpm. Then, to imitate intestinal digestion, the pH of the gastric digesta was adjusted to 7.0 with 0.1 mol/L NaHCO3, and 40 mL artificial intestinal solution (7.5 g/L bile salt, 0.94 g/L pancreatin, and 8.4 g/L NaHCO3 in distilled water) was added. The mixture was incubated at 37 °C for 2 h. The final reaction solution was collected and stored at −20 °C for use in a later experiment to assess the antioxidant activity and total phenolic compounds (TPC).
2.6. α-Glucosidase Inhibitory (AGI) Activity
The AGI activity was analyzed using a modified method by Chen et al. (2014) [19]. The reaction mixture, containing 50 μL of 0.02 mol/L sodium phosphate buffer (pH 6.8), 50 μL of sample, 200 μL of 2 mmol/L p-nitrophenyl α-D-glucopyranoside solution, and 50 μL α-glucosidase enzyme solution, was incubated at 37 °C for 40 min. The reaction was terminated by adding 1.5 mL of 0.1 mol/L Na2CO3, and the inhibitory activity was measured by determining the absorbance at 405 nm. The inhibitory activity was defined as follows:
Abssample = absorbance of the experimental sample
Absblank = absorbance of the blank
Abscontrol = absorbance of the control
2.7. α-Amylase Inhibitory (AAI) Activity
The AAI activity was analyzed using a modified method reported by Dilna et al. (2015) [20]. The reaction mixture, containing 50 μL of sample added to 50 μL of 0.02 mol/L SPB (pH 7.0), 50 μL porcine α-amylase enzyme solution, and 100 μL soluble starch solution (0.01 g/mL), was incubated at 37 °C for 40 min. Then, 1 mL of 3,5-dinitrosalicylic acid solution was added and incubated in a 95 °C water bath for 5 min, followed by the addition of 3 mL sterile distilled water. The absorbance was measured at 540 nm. The inhibitory activity was defined as follows:
Abssample = absorbance of the experimental sample
Absblank = absorbance of the blank
Abscontrol = absorbance of the control
2.8. HMG-CoA Reductase Inhibitory Activity
The HMG-CoA reductase inhibitory (HMGRI) activity was determined on the basis of spectrophotometric measurements. A total of 1 μL of sample, 182 μL of 1× assay buffer, 12 μL HMG-CoA, 4 μL nicotinamide adenine dinucleotide phosphate (reduced form), and 2 μL HMG-CoA reductase were added in order and mixed. The reaction was incubated at 37 °C, and the absorbance was measured at 340 nm within 10 min. Atorvastatin (100 mmol/L) was used as a positive control. The inhibitory activity was defined as follows:
Δ Abssample = change in absorbance of the experimental sample
Δ Abscontrol = change in absorbance of the control
2.9. DPPH Radical Scavenging Activity
The scavenging activity of the DPPH radicals was analyzed using a modified method by Chen et al. (2014) [17]. A total of 100 μL of sample and 1.2 mL of 0.2 mmol/L DPPH solution were mixed and reacted for 30 min in the dark at room temperature. The DPPH radical scavenging activity was measured by the decrease in absorbance at 517 nm. Positive control samples contained ascorbic acid (1 mg/mL) as the standard antioxidant. The EC50 (mg/mL) value was determined from a linear regression analysis of scavenging activity versus the sample concentration, which is defined as the amount of antioxidant needed to decrease the initial DPPH radical concentration by 50%. Moreover, the DPPH radical scavenging activity expressed as percentage value was defined as follows:
DPPH radical scavenging activity (%)
Abssample = absorbance of the experimental sample
Absblank = absorbance of the blank
Abscontrol = absorbance of the control
2.10. The SOD-Like Activity
The superoxide dismutase (SOD)-like activity was analyzed using a modified method by Lee et al. [21]. A total of 100 μL of sample was mixed with 1.5 mL tris-HCl buffer (50 mmol/L tris (hydroxymethyl) aminomethane + 10 mmol/L ethylenediaminetetraacetic acid, pH 8.0) and 100 μL of 7.2 mmol/L pyrogallol. The mixture reacted for 10 min at room temperature. The SOD-like activity was measured the absorbance at 450 nm. Ascorbic acid (1 mg/mL) was used as a positive control. The IC50 (mg/mL) value was defined as the number of antioxidants that inhibited pyrogallol by 50% and was determined by linear regression of the inhibitory activity of pyrogallol versus the sample concentration. The SOD-like activity expressed as percentage value was defined as follows
Abssample = absorbance of the experimental sample
Absblank = absorbance of the blank
Abscontrol = absorbance of the control
2.11. Reducing Power
The reducing power was analyzed using a modified method by Lin and Yen (1999) [22]. A mixture of 250 μL of sample, 250 μL of 0.2 mol/L SPB (pH 6.6), and 250 μL potassium ferricyanide (10 mg/mL) was mixed and reacted for 20 min in a 50 °C water bath. After reacting, 250 μL trichloroacetic acid (TCA) was added and centrifuged at 12,000× g for 5 min. Then, 500 μL of the supernatant, 500 μL of distilled water, and 100 μL ferric chloride (1 mg/mL) was mixed. The absorbance was measured at 700 nm. Ascorbic acid (1 mg/mL) was used as a positive control. The EC50 (mg/mL) value was defined as the concentration that yields an absorbance of 0.5 for reducing power. The reducing power expressed as percentage value was defined as follows:
Abssample = absorbance of the experimental sample
Abspositive control = absorbance of the positive control (ascorbic acid)
2.12. Total Polyphenol Contents
The TPC was determined using the Folin–Ciocaltue method with some modifications (Obanda and Owuor, 1997) [23]. A gallic acid standard curve ranging from 0.1 to 0.75 mg/mL was prepared, and the results that were determined from the regression equation of the standard curve were expressed as the milligram gallic acid equivalents (GAE)/mL and milligram GAE/g in fermented GB. Then, 100 μL of sample was mixed with 500 μL of a 10-fold-diluted Folin–Ciocalteu phenol reagent, followed by the addition of 400 μL of Na2CO3 (75 mg/mL) solution. The mixture reacted for 1 h at room temperature, and the absorbance was measured at 765 nm.
2.13. Statistical Analysis
The data are expressed as the mean ± standard deviation of duplicates. Analysis of variance (ANOVA) and Fisher’s means comparison tests were performed to determine significant differences (p < 0.05) in the results, using Minitab 16.0 software (Minitab Inc., State College, USA).
3. Results and Discussion
3.1. Microbiological and Physicochemical Change during Fermentation
To assess availability of starters we used in fermented GB, we measured changes in the microbial cell counts and pH. During fermentation, viable cell counts of all fermented GB significantly increased more than 1.3 log CFU/mL (Table 1). Even CON, in which the starter was not inoculated, showed microbial growth up to 8.53 log CFU/mL after 24 h fermentation. A morphology analysis confirmed that the Bacillus-like strains had been detected. These Bacillus strains may have survived in spore form during the autoclave process and then grew during fermentation. It was also reported by An et al. [24] that Bacillus spp. grew when the mealworm was fermented without a starter. All Bacillus fermentation GB slightly decreased in pH. According to various studies, some Bacillus strains can lower pH by producing acidic metabolite during the initial stage of fermentation, and some Bacillus strains can increase pH by decomposing protein during fermentation to produce ammonia or amine [25,26]. Fermentation is the process of transforming the raw materials into a useful physiologically active substance by various microorganisms [3]; thus, the fermentation pattern varied with a starter or components such as the protein or carbohydrate of the raw materials. Therefore, it is thought that the combined effect of producing alkaline substances or acidic metabolites slightly lowered the pH of CON, SKGB, and SEGB, whereas the pH of HAGB and LPGB, which inoculated LAB, distinctly decreased about 2.1−2.7 during fermentation. The decrease in pH was mainly caused by organic acids containing lactic acid produced by LAB, MKHA15, and L. plantarum KCTC 3103.
In probiotic foods, probiotics are considered to provide concentrations of at least 106 CFU/mL to achieve therapeutic effects, and higher viable cell counts are required in order to promote a functional effect [27]. In this study, all starters we used were significantly grown in GB for 24 h. In particular, MKHA15 and MKSK-J1, which survived more than 8.5 log CFU/mL in fermented GB, were confirmed to be suitable strains for probiotic foods.
3.2. Inhibitory Activities of the Carbohydrate-Hydrolyzing Enzyme
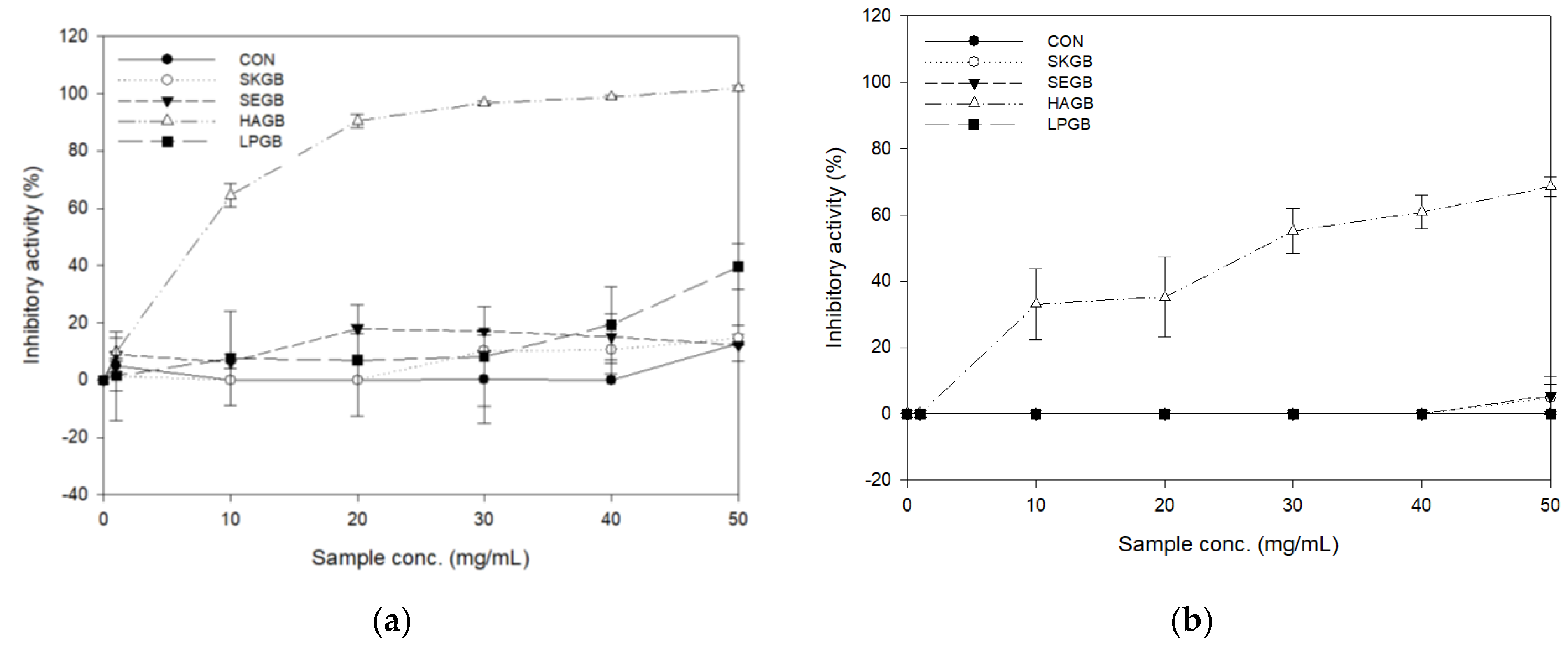
Inhibition of the carbohydrate-hydrolyzing enzyme is one of the ways to treat T2DM. The carbohydrate-hydrolyzing enzyme inhibitory activities of the fermented GB are shown in Figure 1.
The AGI activity of HAGB was significantly increased in a concentration-dependent manner and showed a strong inhibitory activity (> 96% at 30 mg/mL). The AGI activity of LPGB increased up to 39.61% at 50 mg/mL, whereas CON, SKGB, and SEGB were also very low, even at 50 mg/mL (12.36–14.64%). The AAI activity of fermented GB was also significantly high in HAGB alone. As the concentration of HAGB increased, the AAI activity also increased, reaching 68.48% at 50 mg/mL. On the other hand, other fermented GBs had no effect on inhibition of α-amylase. The AGI and AAI activity of MKHA15 were 99.25% and 69.68%, respectively (Table A1). The AGI activity and AAI activity of MKHA15 were strongly maintained after fermentation, which resulted in HAGB having excellent antidiabetic activity. The AGI activity was also strongly maintained in the study by Jang and Kim [28], who fermented onion, tomato, and cabbage juice with MKHA15. LPGB did not exhibit powerful antidiabetic activity like HAGB; it showed 39.61% of AGI and no AAI activity at 50 mg/mL. As reported in several studies, AGI activity is exhibited by physiologically active substances, such as an exopolysaccharides (EPS) or inulin, which are produced by some LAB [19,29]. According to the study by Chen et al. (2014), which measured the AGI activity for 12 LAB species, 12 strains showed of approximately 3.42%−29.57% AGI activity [19], whereas the AGI activity for MKHA15 was much stronger at 99.25%. Notably, HAGB was much higher than LPGB fermented with the same L. plantarum species. According to the study by Ramchandran and Shah [30], there is a difference in EPS productivity, even among the same species, and there is a difference in the AGI activity of fermented yogurt that use those species. This is consistent with the goal of antidiabetes, which exhibits strong AGI and moderate AAI activity; thus, HAGB exhibits excellent antidiabetic activity.
3.3. Anticholesterol Activity
The HMGRI activity of fermented GB is shown in Table 2. The HMGRI activity of all GB fermented with a starter culture was significantly higher than that of CON (2.91%). In particular, HAGB showed significantly strong HMGRI activity (120.89%), even more than that of atorvastatin (111.24%), which was used as the positive control, and LPGB (103.24%). SKGB (45.50%) and SEGB (37.90%) showed similar HMGRI activity. The MKSK-J1 had no HMGRI activity (Table A1), but after GB fermentation, it increased to 45.5%. This was thought to be caused by various bioactive substances produced by interactions between GB and the starter during the fermentation process. Several studies have reported that some phenolic compounds, polyglutamic acids, or viscous substances produced during fermentation may exhibit HMGRI activity [31,32]. On the other hand, MKHA15, which had 65.81% of HMGRI activity, increased to 120.89% after GB fermentation. Several researchers have reported that during fermentation, some organic acids such as ferulic acid and propionic acid, or peptides inhibit HMGR activity [33,34]. Therefore, in our results, it was predicted that HAGB showed significantly high HMGRI activity as the combined effect from various active substances such as organic acids and peptides increased during fermentation as well as the enzyme inhibitory activity of the strain itself.
Another point to note about the result of HMGRI activity is that there was no inhibitory activity for CON. Even in CON, unknown Bacillus strains proliferated during the 24 h of fermentation, although it was difficult to achieve the desired functionality through natural fermentation.
3.4. Antioxidant Activity and the TPC of Fermented GB
The antioxidant activities and TPC of fermented GB are shown in Table 3. The antioxidant activities of fermented GB were measured by three methods—DPPH radical scavenging activity is a measure of the degree to which stable DPPH radical is reduced by antioxidants with the electron-donating ability [35], and reducing power is a measurement the ability to reduce of the Fe3+/ferricyanide complex to the Fe2+/ferrous form in the presence of the reducer (i.e., antioxidants) [36]. DPPH radical scavenging activity expressed as EC50 was significantly low in SKGB and SEGB (8.21 and 8.88 mg/mL), followed by that of LPGB (12.71 mg/mL), HAGB (17.05 mg/mL), and CON (19.60 mg/mL). Furthermore, the reducing power expressed as EC50 was also significantly low in SKGB (0.29 mg/mL), followed by that of HAGB (0.69 mg/mL), SEGB (0.88 mg/mL), LPGB (0.94 mg/mL), and CON (2.06 mg/mL). The SOD-like activity is a measurement the inhibition of pyrogallol auto-oxidation, which is catalyzed by the superoxide radicals [37]. The SOD-like activity, expressed as IC50, was significantly low in HAGB (22.37 mg/mL), followed by that of LPGB (30.51 mg/mL), SEGB (46.41 mg/mL), SKGB (49.79 mg/mL), and CON (67.34 mg/mL). During fermentation, the starter could enhance certain functionalities, such as the antioxidant activity, by degrading proteins, carbohydrates, and polyphenols, etc. from raw materials through microbial bioconversion [3]. Bacillus spp. has an excellent ability to produce low-molecular weight functional compounds or peptides by decomposing high-molecular weight substances by secreting various hydrolysis enzymes [3]. On the other hand, facultative anaerobes such as Lactobacillus spp. that grow in the presence of oxygen have enzymes such as SOD to remove free radicals produced by a reaction with oxygen [38]. Additionally, it has been reported that EPS produced by LAB has antioxidant activity like SOD [39]. For such a reason, in measuring the antioxidant activity of a mixed substance, it is important to evaluate the different mechanisms using several assays, rather than using only a single assay for the evaluation [40]. Some differences depend on the starter, but the overall fermented GB inoculated starter culture, especially SKGB and HAGB, exhibited good antioxidant activity compared to that of natural fermentation (CON).
The TPC of fermented GB was also higher in the fermented GB inoculated starter culture (329.40−409.00 mg GAE/g) than in the CON (91.30 mg GAE/g). Polyphenols are known to be major indicators of antioxidant activity in soybean, fruits, vegetables, cereals, etc. According to Mattia et al. [41], the antioxidants present in edible insects are phenolic compounds, peptides, and un-identified components. Therefore, it can be predicted that the excellent antioxidant activity of the fermented GB combined with the starter is caused by high TPC, peptides, or uncertain components, which are increased by the starter.
3.5. Antioxidant Activity and TPC After In Vitro Digestion
The antioxidant activities and TPC of fermented GB before and after in vitro digestion are shown in Table 4. In SKGB and HAGB, TPC greatly increased after in vitro digestion by more than sevenfold. As for the trend in the TPC change, the reducing power also increased significantly after in vitro digestion in SKGB and HAGB. Several studies have reported that the digestive process can exert beneficial effects by releasing bioactive compounds, and the antioxidant activity also increases due to the phenolic compounds of extracts, fermented products, or peptides obtained by the enzymatic digestion of proteins [42,43].
The SOD-like activity significantly increased in LAB-fermented GB (HAGB and LPGB). As explained in Section 3.4, LAB may have SOD for survival, and they may produce EPS during fermentation by the carbohydrate metabolism, and thus they can exhibit SOD-like activity for such reasons. We confirmed that the excellent SOD-like activities of HAGB and LPGB are strongly sustained even after in vitro digestion. Except for DPPH radical scavenging activity, GB fermented with a probiotic starter had higher antioxidant activity after in vitro digestion than other fermented GB. The DPPH radical scavenging activity showed an opposite trend in other antioxidant experiments, increasing in SEGB and LPGB, and decreasing in CON, SKGB, and HAGB. According to Chiang et al. [44], in vitro digestion could enhance the antioxidant components, but the antioxidant activity may differ among experiments due to differences in the mechanism of the antioxidants. It is reported that decrease or absence of change in antioxidant activity after in vitro digestion was due to resistance to hydrolysis enzyme or pH change [45]. LPGB and SEGB decreased in TPC but increased in DPPH radical scavenging activity. Through this, we can assume that the DPPH radical scavenging activity of fermented GB was due not only to TPC but also other antioxidants. A study by Pavan et al. [45], which measured the change in antioxidant activity of Annona marcgravii according to in vitro digestion, showed that the TPC decreased, but the antioxidant activity increased, which is similar to the result of this study. Furthermore, they predicted that non-phenolic substances, amino acids, and peptides involved in the increase of antioxidant activity may have been activated during in vitro digestion.
In this study, the GB that was fermented using a probiotic starter (MKHA15 and MKSK-J1) indicated overall high antioxidant activities after in vitro digestion, which is thought to be the result of the MKHA15 and MKSK-J1 playing an excellent role in increasing TPC or other unknown antioxidants in GB fermented by the starter and digestive enzymes.
4. Conclusions
GB was fermented with Lactobacillus and Bacillus starter to investigate the bioactive properties. The inhibitory activities of α-glucosidase, α-amylase, and HMG-CoA reductase were significantly higher for HAGB than for the other GB samples. The antioxidant activity and TPC showed excellent performance in the fermented GB-inoculated starter culture compared to those of the CON. After in vitro digestion, the antioxidant activities and TPC were enhanced by fermented GB inoculated with probiotic starter. In this study, we developed GB fermented with MKHA15, which also possesses excellent antioxidant, anti-diabetic, and anti-cholesterol activities. This study could be used as valuable research data to supplement existing fermentation studies of edible insects where data were insufficient.
Author Contributions
Formal analysis, data curation, and writing—original draft, H.J.; funding acquisition, conceptualization, supervision, and writing—review and editing, M.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by National Research Foundation (NRF) of Korea, 2020R1I1A3072127 and Korea Institute of Planning and Evaluation for Technology in Food, Agriculture, and Forestry (IPET), 119019-02-1-HD020.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
This article is a part of Hyunah Jang’s master’s thesis at the Department of Food Science and Nutrition, Dankook University, Korea, under the supervision of Misook Kim.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A

{kind=link}
Table A1.
Probiotic and physiological characteristics of MKHA15 and MKSK-J1.
| Characteristics | MKHA15 1,2 | MKSK-J1 3,4 | |
|---|---|---|---|
| Acid and bile salt tolerance | Survival rate to pepsin (%) | 96.30 ± 0.10 5 | 91.85 ± 4.98 |
| Survival rate to bile salt (%) | 53.56 ± 5.86 | 90.31 ± 0.44 | |
| Safety test | γ-hemolysis | γ-hemolysis | |
| Enzymatic characteristics | Amylase | β-Glucuronidase negative | N.D.4 |
| Protease | Negative (qualitative) | 1.37 ± 0.63 (U/mL) | |
| Antioxidant activity | DPPH radical scavenging activity (%) | N.D.6 | 23.22 ± 0.21 (U/mL) |
| SOD-like activity (%) | 100.53 ± 6.91 | 7.69 ± 2.71 | |
| Reducing power (% ascorbic acid) | 56.62 ± 1.77 | 28.10 ± 2.04 | |
| Antidiabetic activity | α-Glucosidase inhibitory activity (%) | 98.76 ± 7.12 | 41.36 ± 0.63 |
| α-Amylase inhibitory activity (%) | 69.68 ± 2.38 | None | |
| Anticholesterol activity | HMG-CoA reductase inhibitory activity (%) | 65.81 ± 6.50 | −6.04 ± 7.70 |
1Lactobacillus plantarum MKHA15. 2 Source: Kim, M.S.; Seong, H.Y.; Jang, H.A.; Jeong, Y.H.; and Choi, J.A. (2019). Novel Lactobacillus plantarum MKHA15 and uses thereof. Korea patent pending. Invertor: Industry-Academic cooperation Foundation of Dankook University Cheonan Campus, application number: Korea 10-2019-0112122 (10 September 2019). 3 Bacillus amyloliquefaciens MKSK-J1. 4 Source: Lee, S.K.; Lee, J.J.; Jin, Y.I.; Jeong, J.C.; Chang, Y.H.; Lee, Y.S.; Jeong, Y.H.; and Kim, M.S. (2017). Probiotic characteristics of Bacillus strains isolated from Korean traditional soy sauce. LWT-Food Science and Technology, 79, 518–524. 5 Values are mean ± standard error (n = 2). 6 N.D. = not determined. N.D.4 = not determined.
References
- Zhao, H.L.; Cho, J.H.; Chung, C.H. Characteristics of seasoning pastes fermented by Aspergillus oryzae and Bacillus sutilis using edible insects. Korean J. Food Sci. Technol. 2018, 50, 297–307. [Google Scholar]
- Dobermann, D.; Field, L.N.; Michaelson, L.V. Impact of heat processing on the nutritional content of Gryllus bimaculatus (black cricket). Nutr. Bull. 2019, 44, 116–122. [Google Scholar] [CrossRef] [Green Version]
- Sim, S.Y.; Jang, S.H.; Ahn, H.Y.; Cho, H.D.; Seo, K.I.; Cho, Y.S. Optimization of fermentation conditions Protaetia brevitarsis seulensis larvae using Bacillus subtilis. Korean J. Food Preserv. 2019, 26, 123–133. [Google Scholar] [CrossRef] [Green Version]
- Udomsil, N.; Imsoonthornruksa, S.; Gosalawit, C.; Ketudat-cairns, M. Nutritional Values and Functional Properties of House Cricket (Acheta domesticus) and Field Cricket (Gryllus bimaculatus). Food Sci. Technol. Res. 2019, 25, 597–605. [Google Scholar] [CrossRef]
- Zielinska, E.; Baraniak, B.; Karas, M.; Rybczynska, K.; Jakubczyk, A. Selected species of edible insects as a source of nutrient composition. Food Res. Int. 2015, 77, 460–466. [Google Scholar] [CrossRef]
- Lee, D.G.; Kim, K.B.; Choi, S.K. Quality Characteristics of Two-spotted Cricket (Gryllus bimaculatus) Brown Stock by High Pressure Cooking. Culi. Sci. Hos. Res. 2017, 23, 163–174. [Google Scholar]
- Roglic, G. WHO Global report on diabetes: A summary. Int. J. Noncommun. Dis. 2016, 1, 3–8. [Google Scholar] [CrossRef]
- Taslimi, P.; Koksal, E.; Goren, A.C.; Bursal, E.; Aras, A.; Kilic, O.; Alwasel, S.; Gulcin, I. Anti-Alzheimer, antidiabetic and antioxidant potential of Satureja cuneifolia and analysis of its phenolic contents by LC-MS/MS. Arab. J. Chem. 2020, 13, 4528–4537. [Google Scholar] [CrossRef]
- Erejuwa, O.O.; Akpan, J.L.; Uwaezuoke, N.J.; Nwobodo, N.N.; Ezeokpo, B.C.; Erhiano, E.; Araromi, E.J.; Ude, U.N.; Abdul, W.M.; Sulaiman, S.A. Effects of Honey on Postprandial Hyperlipidemia and Oxidative Stress in Wistar Rats: Role of HMG-CoA Reductase Inhibition and Antioxidant Effect. Niger. J. Physiol. Sci. 2018, 33, 129–138. [Google Scholar]
- Beatrice, A.G.; Marcella, A.E. Statin adverse effects: A review of the literature and evidence for a mitochondrial mechanism. Am. J. Cardiovasc. Drugs 2008, 8, 373–418. [Google Scholar]
- Wu, H.; Xu, B. Inhibitory effects of onion against α-glucosidase activity and its correlation with phenolic antioxidants. Int. J. Food Prop. 2014, 17, 599–609. [Google Scholar] [CrossRef]
- FAO/WHO. Joint FAO/WHO Working Group Report on Drafting Guidelines for the Evaluation of Probiotics in Food; FAO/WHO: London, ON, Canada, 2002. [Google Scholar]
- Smid, E.J.; Kleerebezem, M. Production of Aroma Compounds in Lactic Fermentations. Annu. Rev. Food Sci. Technol. 2014, 5, 313–326. [Google Scholar] [CrossRef] [PubMed]
- Frederic, L.; Jurgen, V.; Luc, D.V. Functional meat starter cultures for improved sausage fermentation. Int. J. Food Microbiol. 2006, 106, 270–285. [Google Scholar]
- Omafuvbe, B.O.; Abiose, S.H.; Shonukan, O.O. Fermentation of soybean (Glycine max) for soy-daddawa production by starter cultures of Bacillus. Food Microbiol. 2002, 19, 561–566. [Google Scholar] [CrossRef]
- Kim, M.S.; Seong, H.Y.; Jang, H.A.; Jeong, Y.H.; Choi, J.A. Novel Lactobacillus plantarum MKHA15 and Uses thereof. Korea Granted Patent. Invertor: Industry-Academic Cooperation Foundation of Dankook University Cheonan Campus. Application number Korea 10-2019-0112122, 10 September 2019. [Google Scholar]
- Lee, S.K.; Lee, J.J.; Jin, Y.I.; Jeong, J.C.; Chang, Y.H.; Lee, Y.S.; Jeong, Y.H.; Kim, M.S. Probiotic characteristics of Bacillus strains isolated from Korean traditional soy sauce. LWT-Food Sci. Technol. 2017, 79, 518–524. [Google Scholar] [CrossRef]
- Li, E.; Yang, H.; Zou, Y.; Wang, H.; Hu, T.; Li, Q.; Liao, S. In-Vitro digestion by simulated gastrointestinal juices of Lactobacillus rhamnosus cultured with mulberry oligosaccharides and subsequent fermentation with human fecal inocula. LWT-Food Sci. Technol. 2019, 101, 61–68. [Google Scholar] [CrossRef]
- Chen, P.; Zhang, Q.; Dang, H.; Liu, X.; Tian, F.; Zhao, J.; Chen, Y.; Zhang, H.; Chen, W. Screening for potential new probiotic based on probiotic properties and α-glucosidase inhibitory activity. Food Control 2014, 35, 65–72. [Google Scholar] [CrossRef]
- Dilna, S.V.; Surya, H.; Aswathy, R.G.; Varsha, K.K.; Sakthikumar, D.N.; Pandey, A.; Nampoothiri, K.M. Characterization of an exopolysaccharide with potential health-benefit properties from a probiotic Lactobacillus plantarum RJF4. LWT-Food Sci. Technol. 2015, 64, 1179–1186. [Google Scholar] [CrossRef]
- Lee, H.J.; Do, J.R.; Jung, S.K.; Kim, H.K. Physiological Properties of Sarcodon aspratus Extracts by Ethanol Concentration. J. Korean Soc. Food Sci. Nutr. 2014, 43, 656–660. [Google Scholar] [CrossRef]
- Lin, M.Y.; Yen, C.L. Antioxidative Ability of Lactic Acid Bacteria. J. Agric. Food Chem. 1999, 47, 1460–1466. [Google Scholar] [CrossRef]
- Obanda, M.; Owuor, P.O. Flavanol Composition and Caffeine Content of Green Leaf as Quality Potential Indicators of Kenyan Black Teas. J. Sci. Food Agric. 1997, 74, 209–215. [Google Scholar] [CrossRef]
- An, B.; Sam, C.; Dries, V.; Ruben, S.; Christel, V.; Mik, V.D.B.; Bart, L.; Leen, V.C. Comparison of Six Commercial Meat Starter Cultures for the Fermentation of Yellow Mealworm (Tenebrio molitor) Paste. Microorganisms 2019, 7, 540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, M.J.; Lee, J.Y.; Kim, J.H. Microbial and Physiochemical Properties of Cheonggukjang Fermented Using Bacillus strains with Antibacterial or Antifungal Activities. Food Sci. Biotechnol. 2014, 23, 1525–1532. [Google Scholar] [CrossRef]
- Lee, J.Y.; Shim, J.M.; Liu, X.; Yao, Z.; Lee, K.W.; Cho, K.M.; Kim, G.M.; Shin, J.H.; Kim, J.S.; Kim, J.H. Inhibition of Bacillus cereus in Cheonggukjang Fermentation with Bacillus Starters with Antimicrobial Activities. J. Korean Soc. Food Sci. Nutr. 2016, 45, 736–745. [Google Scholar] [CrossRef]
- Xiao, Y.; Wang, L.; Rui, X.; Li, W.; Chen, X.; Jiang, M.; Dong, M. Enhancement of the antioxidant capacity of soy whey by fermentation with Lactobacillus plantarum B1–6. J. Funct. Foods 2015, 12, 33–44. [Google Scholar] [CrossRef]
- Jang, H.A.; Kim, M.S. Characteristics of Vegetable Juice Fermented with Lactobacillus plantarum MKHA15 and Leuconostoc mesenteroides MKSR. J. Korean Diet. Assoc. 2019, 25, 281–294. [Google Scholar]
- Nurhayati, R.; Miftakhussolikhah; Frediansyah, A.; Rachmah, D.L. Lactic acid bacteria producing inhibitor of alpha glucosidase isolated from Ganyong (Canna Edulis) and Kimpul (Xanthosoma sagittifolium). Earth Environ. Sci. 2017, 101, 1–5. [Google Scholar] [CrossRef]
- Ramchandran, L.; Shah, N.P. Effect of exopolysaccharides and inulin on the proteolytic, angiotensin-I-converting enzyme- and α-glucosidase-inhibitory activities as well as on textural and rheological properties of low-fat yogurt during refrigerated storage. Dairy Sci. Technol. 2009, 89, 583–600. [Google Scholar] [CrossRef]
- Son, K.H.; Lee, J.Y.; Lee, J.S.; Kang, S.S.; Sohn, H.Y.; Kwon, C.S. Screening of Phenolic Compounds with Inhibitory Activities against HMG-CoA Reductase. J. Life Sci. 2017, 27, 325–333. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.G.; Kim, H.J.; Im, N.K.; Lee, E.J.; Lee, S.P.; Lee, I.S. Antithrombotic and Cholesterol Reduction Effects of Defatted Soybean Grits Fermented by Bacillus subtilis NUC1. Korean J. Food Sci. Technol. 2009, 41, 423–427. [Google Scholar] [CrossRef] [Green Version]
- Tomaro-Duchesneau, C.; Jones, M.; Shah, D.; Jain, P.; Saha, S.; Prakash, S. Cholesterol Assimilation by Lactobacillus Probiotic Bacteria: An In Vitro Investigation. BioMed Res. Int. 2014, 1, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Oh, N.S.; Kim, K.M.; Oh, S.N.; Kim, Y.H. Enhanced Production of Galactooligosaccharides Enriched Skim Milk and Applied to Potentially Synbiotic Fermented Milk with Lactobacillus rhamnosus 4B15. Food Sci. Anim. Resour. 2019, 39, 725–741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.J.; Kim, J.H.; Lee, S.H.; Cho, E.J.; Kim, H.Y. Determination of Radical Scavenging Activity of Aster yomena (Kitam.) Honda. J. Korean Acad. Industr. Coop. Soc. 2018, 19, 402–407. [Google Scholar]
- Ferreira, I.C.; Baptista, P.; Vilas-Boas, M.; Barros, L. Free-radical scavenging capacity and reducing power of wild edible mushrooms from northeast Portugal: Individual cap and stipe activity. Food Chem. 2007, 100, 1511–1516. [Google Scholar] [CrossRef]
- Kim, S.J.; Han, D.S.; Moon, K.D.; Rhee, J.S. Measurement of Superoxide Dismutase-like Activity of Natural Antioxidants. Biosci. Biotechnol. Biochem. 1995, 59, 822–826. [Google Scholar] [CrossRef] [Green Version]
- Brioukhanov, A.L.; Thauer, R.K.; Netrusov, A.I. Catalase and superoxide dismutase in the cells of strictly anaerobic microorganisms. Microbiology 2002, 71, 281–285. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, C.; Li, D.; Zhao, Y.; Zhang, X.; Zeng, X. Antioxidant activity of an exopolysaccharide isolated from Lactobacillus plantarum C88. Int. J. Biol. Macromol. 2013, 54, 270–275. [Google Scholar] [CrossRef] [PubMed]
- Nissen, L.; Samaei, S.P.; Babini, E.; Gianotti, A. Gluten free sourdough bread enriched with cricket flour for protein fortification: Antioxidant improvement and Volatilome characterization. Food Chem. 2020, 333, 1–13. [Google Scholar] [CrossRef]
- Mattia, C.D.; Battista, N.; Sacchetti, G.; Serafini, M. Antioxidant Activities In Vitro of Water and Liposoluble Extracts Obtained by Different Species of Edible Insects and Invertebrates. Front. Nutr. 2019, 1, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.K.; Choi, Y.R.; Park, P.J.; Choi, J.H.; Moon, S.H. Purification and characterization of antioxidative peptides from enzymatic hydrolysate of Cod Teiset protein. J. Korean Fish. Soc. 2000, 33, 198–204. [Google Scholar]
- Hall, F.; Johnson, P.E.; Liceaga, A. Effect of enzymatic hydrolysis on bioactive properties and allergenicity of cricket (Gryllodes sigillatus) protein. Food Chem. 2018, 262, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Chiang, C.J.; Kadouh, H.; Zhou, K. Phenolic compounds and antioxidant properties of gooseberry as affected by In Vitro digestion. LWT-Food Sci. Technol. 2013, 51, 417–422. [Google Scholar] [CrossRef]
- Pavan, V.; Sancho, R.A.S.; Pastore, G.M. The effect of In Vitro digestion on the antioxidant activity of fruit extracts (Carica papaya, Artocarpus heterophillus and Annona marcgravii). LWT-Food Sci. Technol. 2014, 59, 1247–1251. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
(a) α-Glucosidase inhibitory activity of fermented GB; (b) α-amylase inhibitory activity of fermented GB.
Figure 1.
(a) α-Glucosidase inhibitory activity of fermented GB; (b) α-amylase inhibitory activity of fermented GB.
Table 1.
Change of viable cell counts and pH during Gryllus bimaculatus (GB) fermentation.
| Viable Cell Counts (log CFU/mL) | ||
| 0 h 1 | 24 h | |
| CON | 0.00 ± 0.00 b | 8.53 ± 0.19 a |
| SKGB | 5.40 ± 0.14 b | 8.88 ± 0.17 a |
| SEGB | 6.11 ± 0.00 b | 8.72 ± 0.17 a |
| HAGB | 8.22 ± 0.13 b | 9.52 ± 0.17 a |
| LPGB | 7.85 ± 0.00 b | 9.25 ± 0.03 a |
| pH | ||
| 0 h | 24 h | |
| CON | 7.08 a | 6.32 ± 0.21 b |
| SKGB | 7.05 a | 6.27 ± 0.01 b |
| SEGB | 6.72 a | 5.72 ± 0.00 b |
| HAGB | 6.67 a | 3.97 ± 0.02 b |
| LPGB | 6.37 a | 4.27 ± 0.00 b |
1 Fermentation time. Values are mean ± standard error (n = 2). Means with different superscripts in a row (a,b) are significantly different by Fisher’s least significant difference test (p < 0.05).
Table 2.
3-Hydroxy-methyl glutaryl-coenzyme A (HMG-CoA) reductase inhibitory activity of fermented GB.
Table 2.
3-Hydroxy-methyl glutaryl-coenzyme A (HMG-CoA) reductase inhibitory activity of fermented GB.
| HMG-CoA Reductase Inhibitory Activity (%) | |
|---|---|
| CON | 2.91 ± 6.02 d |
| SKGB | 45.50 ± 5.99 c |
| SEGB | 37.90 ± 2.88 c |
| HAGB | 120.89 ± 0.42 a |
| LPGB | 103.24 ± 9.11 b |
| Atorvastatin (100 mM) | 111.24 ± 8.60 |
Values are mean ± standard error (n = 2). Means with different superscripts in a column (a–d) are significantly different in terms of Fisher’s least significant difference test (p < 0.05).
Table 3.
Antioxidant activity and total phenolic compounds of fermented GB.
| DPPH Radical Scavenging Activity (EC50 mg/mL) | SOD-Like Activity (IC50 mg/mL) | Reducing Power (EC50 mg/mL) | TPC (mg GAE/g) | |
|---|---|---|---|---|
| CON | 19.60 ± 3.94 a | 67.34 ± 8.22 a | 2.06 ± 0.30 a | 91.30 ± 24.18 c |
| SKGB | 8.21 ± 0.24 c | 49.79 ± 6.85 a,b | 0.29 ± 0.02 c | 367.50 ± 3.13 a,b |
| SEGB | 8.88 ± 1.80 c | 46.41 ± 2.64 b,c | 0.88 ± 0.03 b | 393.21 ± 30.81 a |
| HAGB | 17.05 ± 1.10 a,b | 22.37 ± 0.54 d | 0.69 ± 0.04 b | 329.40 ± 7.64 b |
| LPGB | 12.71 ± 0.82 b,c | 30.51 ± 3.06 c,d | 0.94 ± 0.00 b | 409.00 ± 16.00 a |
Values are mean ± standard error (n = 2). Means with different superscripts in a column (a–d) are significantly different in terms of Fisher’s least significant difference test (p < 0.05).
Table 4.
Antioxidant activity and TPC of fermented GB before and after in-vitro digestion.
| DPPH Radical Scavenging Activity (%) | SOD-Like Activity (%) | Reducing Power (% Ascorbic Acid) | TPC (mg GAE/mL) | |||||
|---|---|---|---|---|---|---|---|---|
| CON | 28.88 ± 4.73 A,a | 13.58 ± 3.53 A,b | 9.61 ± 1.94 A,b | 13.84 ± 2.40 A,c | 39.15 ± 2.78 B,b | 60.33 ± 0.71 A,b | 9.13 ± 2.42 A,a | 10.54 ± 0.86 A,b |
| SKGB | 28.55 ± 4.20 A,a | 17.77 ± 0.33 A,b | 17.13 ± 3.34 A,a | 15.34 ± 1.40 A,b,c | 46.47 ± 0.68 B,a | 86.06 ± 0.91 A,a | 1.28 ± 0.03 B,c | 12.48 ± 0.00 A,a |
| SEGB | 1.92 ± 0.87 B,c | 30.49 ± 1.15 A,a | 9.75 ± 3.63 A,b | 7.00 ± 1.79 A,d | 37.71 ± 0.07 A,b | 37.36 ± 0.04 B,d | 5.51 ± 0.24 A,b | 2.38 ± 0.27 B,d |
| HAGB | 12.42 ± 1.32 A,b | 5.62 ± 1.39 B,c | 9.60 ± 2.15 B,b | 21.18 ± 2.47 A,a | 29.28 ± 0.08 B,c | 43.08 ± 0.19 A,c | 0.64 ± 0.02 B,c | 7.99 ± 0.79 A,c |
| LPGB | 2.36 ± 1.76 B,c | 15.97 ± 0.78 A,b | 10.70 ± 2.11 B,b | 19.11 ± 0.50 A,a,b | 23.65 ± 0.56 A,c | 24.73 ± 0.17 A,e | 4.73 ± 0.04 A,b | 1.10 ± 0.07 B,d |
| Ascorbic acid (1 mg/mL) | 97.41 ± 1.94 | 72.15 ± 4.98 | 100 | N.D.4) | ||||
Values are mean ± standard error (n = 2). Means with different superscripts in a row (A,B) and a column (a–e) are significantly different in terms of Fisher’s least significant difference test (p < 0.05). N.D.4) = not determined.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Jang, H.; Kim, M. Antidiabetic, Anticholesterol, and Antioxidant Activity of Gryllusbimaculatus Fermented by Bacillus and Lactobacillus Strains. Appl. Sci. 2021, 11, 2090. https://doi.org/10.3390/app11052090
AMA Style
Jang H, Kim M. Antidiabetic, Anticholesterol, and Antioxidant Activity of Gryllusbimaculatus Fermented by Bacillus and Lactobacillus Strains. Applied Sciences. 2021; 11(5):2090. https://doi.org/10.3390/app11052090
Chicago/Turabian StyleJang, Hyunah, and Misook Kim. 2021. "Antidiabetic, Anticholesterol, and Antioxidant Activity of Gryllusbimaculatus Fermented by Bacillus and Lactobacillus Strains" Applied Sciences 11, no. 5: 2090. https://doi.org/10.3390/app11052090
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.