Abstract
The present study aimed to unveil the phenotypic heterogeneity and heteroresistance of P. aeruginosa to acidic stress and imipenem. Furthermore, the growth, morphology, and potential for biofilm formation of the subject isolates at different pHs were assessed. Isolates of P. aeruginosa were recovered from juice samples and confirmed by molecular analysis. Antibiotics sensitivity was evaluated using the Kirby–Bauer-disk diffusion method, and the MIC for imipenem was determined, followed by a biofilm formation assay and population analysis. Scanning electron microscopy (SEM) was used to visualize biofilm formation. The subject isolates persisted in an acidic environment and adopted a biofilm lifestyle. The population analysis assay indicated the presence of two distinct phenotypes, i.e., a normal colony phenotype (NCP) and slow growing colony phenotype (SGCP). NCP showed visible colonies after 48 h, while SGCP colonies appeared after 72 h of incubation. Both displayed heteroresistance to imipenem and susceptibility to other antibiotics. Biofilm formation at acidic pH was observed in both phenotypes. Interestingly, the recovery of SGCP was increased in an acidic environment. Biofilm consortia were highly resistant to imipenem. The present study indicated that P. aeruginosa persisted for a long time in an acidic environment, through phenotypic alteration. The subject isolates adopted a biofilm lifestyle and reduced metabolism, to neutralize the effects of acidic pH and imipenem toxicity. Interestingly, the biofilm consortia harbored metabolically active (NCP), as well as inactive populations, of (SGCP), to maintain an active growth and persistency. SGCP retained the potential to revert to NCP upon subsequent sub-culturing in plentiful nutrients and optimum conditions.
1. Introduction
Pseudomonas aeruginosa (P. aeruginosa) is a Gram-negative opportunistic pathogen responsible for nosocomial, community-acquired, and food borne infections [1,2]. P. aeruginosa is well known for its adaptability to diverse environments, antibiotic resistance, and production of extracellular and cell-associated virulence factors [2,3]. Most often, P. aeruginosa has been associated with hospital-acquired infection; however, it is one of the major causes of food spoilage and foodborne diseases [4,5]. It has a tendency to grow and survive in a wide range of pHs (4.5 to 9.0) and a high saline environment [6]. Therefore, it is regularly reported in different food items, e.g., vegetables and fruits, dairy products, beef, mutton, fish products, etc. P. aeruginosa also colonizes a wide variety of surfaces and food packaging materials [7,8].
Pseudomonas possesses a strong tendency to develop biofilm, which aids in its persistence on different surfaces [9]. Biofilm formation ability is directly related to the lethality of the organism and offers resistance to the host immune system, chemotherapy, and disinfectants [10]. The biofilm matrix supports the anchoring of these cells to various surfaces and helps them survive in hostile and unfavorable environmental conditions [6,11]. The matrix generally comprises extracellular metabolites, including polysaccharides, environmental DNA, proteins, lipids, and dead cells [12,13]. The composition of this matrix may vary for different environments and surfaces [14]. The major components of biofilm consortia are polysaccharides, which initiate attachment to the surface and cement the surrounding cells together. Furthermore, it has been reported that biofilm consortia harbor different types of cells, including dead bacterial cells, and metabolically active and inactive cells [15]. Planktonic or metabolically active populations are gradually released or dispersed from these consortia, to colonize new surfaces [16,17].
P. aeruginosa shifts between different lifestyles, to survive in toxic and unfavorable conditions [13,18]. It has been reported that P. aeruginosa biofilms harbor small colony variants, rugose small-colony variants, and persister cells [19,20]. Metabolically inactive populations consist of small colony variants, persister cells, and viable but non-cultivable cells. These phenotypes, with slow metabolic rates, have the potential to tolerate and survive different classes of antimicrobial agents [15].
A study conducted by Wilton and collaborators demonstrated that acidic pH is one of the major factors triggering biofilm formation in P. aeruginosa [21]. Accumulation of eDNA in biofilms and infection sites can acidify the local environment, which further promotes the growth of antibiotic-resistant phenotypes of P. aeruginosa. Moreover, low pH may in itself be the factor responsible for the formation of metabolically inactive phenotypes or slow-growing antibiotic-resistant variants having the potential to revert to the wild-type virulent population.
The present study unearthed novel aspects of phenotypic switching of P. aeruginosa in the acidic environment of pulpy juices and under the stress of an antibacterial agent (imipenem), by triggering biofilm formation, consisting of slow growing resistant variants and normal-pigmented phenotypes. The subject isolates showed heteroresistance to imipenem. Slow growing variants outnumbered the normal population under imipenem stress in the adherent biofilms. This phenomenon was found to be reversible when the stress was relieved and cells were supplemented with sufficient nutrients.
2. Materials and Methods
2.1. Bacterial Strains Isolation and Confirmation
The strains of P. aeruginosa used in this study were isolated from an apple juice concentrate sample of pH = 4.5. Fifty grams of the sample was diluted to 1:10 by adding 450 mL of phosphate buffer saline (pH = 7.0). Ten-fold serial dilutions were prepared to 1:1000, and 1.0 mL from each dilution was inoculated with plate count agar (Oxoid, Hampshire, UK) using a pour plate technique and incubated at 35 °C for 48 to 72 h. The calculation was made according to the following formula:
- N = Number of colonies/mL
- Σc = sum of all colonies on all plates counted
- = number of plates in the first dilution counted
- = number of plates in the second dilution counted
- = dilution from which the first counts were obtained
Isolated colonies were picked from plate count agar and inoculated on Pseudomonas Cetrimide agar (OXOID, UK) and Pseudomonas Agar Base with CN-selective supplement (OXOID, UK). Fluorescent green colonies indicated the presence of P. aeruginosa. Polymerase chain reaction (PCR) was carried out for confirmation, using specific primers, as described earlier targeting oprI for Pseudomonas and with oprL for confirmation of P. aeruginosa [22]. The sequences of primers were as follows:
- oprI-F-5′-ATGAACAACGTTCTGAAATTCTCTGCT-3′
- oprI-R-5′-CTTGCGGCTGGCTTTTTCCAG-3′
- oprL-F-5′-ATGGAAATGCTGAAATTCGGC-3′
- oprL-R-5′-CTTCTTCAGCTCGACGCGACG-3′
2.2. Antibiotic Sensitivity Testing (Kirby Bauer-Disk Diffusion Method)
Antibiotic sensitivity was evaluated using commercially acquired antibiotic discs (Hi-Media, Mumbai, India) with the standard P. aeruginosa (ATCC 27853) as a positive control. The Kirby–Bauer method, recommended by CLSI guidelines 2020 [23], was used for antimicrobial susceptibility testing. The subjected P. aeruginosa strains were tested against imipenem, meropenem, levofloxacin, ceftriaxone, amoxycillin, gentamicin, amikacin, ciprofloxacin, ceftazidime, and cefotaxime.
2.3. MIC Determination for Imipenem
Determination of minimum inhibitory concentration of imipenem against P. aeruginosa strains was carried out on Muller Hinton agar with two-fold concentrations, staring from 16 µg/mL to 2 µg/mL. Fresh cultures were prepared after overnight incubation, and concentrations were set with a 0.5 McFarland index. Subsequently, they were applied on Muller Hinton agar supplemented with different concentrations of imipenem and incubated at 35 °C for 48 h. After 48 h, plates were examined on the basis of inhibition (susceptible ≤ 2 μg/mL, intermediate = 4 μg/mL, and resistant ≥ 8 μg/mL) [23].
2.4. Biofilm Formation Assay
Overnight cultures of the subject isolates of P. aeruginosa (NCP) were inoculated (0.1 mL) in 100-mL TSB flasks, having a pH ranging from 2 to 9 and incubated at 35 °C for 48 h. Sterile glass slides were then placed in the respective flasks with the lower half submerged in broth. After every 6 h, till the 48th h of incubation, these slides were removed and washed twice with phosphate buffer saline and stained accordingly with crystal violet, as described earlier by O’Toole [24]. Briefly, slides were placed in 0.1% crystal violet (CV) solution for 25 min. The CV solution was removed and slides were washed twice with 1 mL of sterile saline and dried at room temperature. Finally, the cell-bound crystal violet was dissolved in 33% acetic acid, and the thickness of biofilm was measured in terms of the optical density (OD) at 570 nm. Two glass slides in TSB without P. aeruginosa were used as a negative control.
2.5. Population Analysis Assay
For population analysis, biofilms were detached from glass slides by sonication, using an ultrasound bath with continuous shaking. After sonication, 1mL of biofilm suspension was transferred to a micro-centrifuge tube and vortexed vigorously for 30 s. The cells were diluted in PBS (pH 7.0) to achieve ~1.0 × 102 cfu/mL, and 0.1 mL of thissuspension was inoculated in 2.5-mL TSB tubes with a pH ranging from 4.0 to 9.0 and incubated at 35 °C. After incubation, 100 μL from the TSB was inoculated onto TSA, Pseudomonas Cetrimide agar (USP, EP) and Pseudomonas Agar Base, to study the colony phenotypes of P. aeruginosa [25].
2.6. Small Colony Variant (SCV) Characterization
Respective plate cultures were assessed at 24 to 48 h post-incubation and colonies were categorized based on their size and pigmentation. The colonies that were >1 mm in diameter, pigmented, were recorded as the WT population. SCVs were recorded as being <1 mm in size, with reduced pigmentation, and appeared after 36 h of incubation, as described earlier [25,26].
2.7. Reversion
SCVs were tested for reversion by sub-culturing individual colonies onto TSA plates and incubated overnight at 35 °C. Reversion was defined as the capacity of the SCV colonies to revert to WT cultures following subsequent sub-culturing and growth under optimal conditions. Colonies that displayed WT characteristics (>1 mm and pigmented in comparison to control plates) were recorded as exhibiting reversion [25,26].
2.8. Scanning Electron Microscopy (SEM)
Scanning electron microscopy (SEM) was performed to analyze the production of extracellular matrix material and cell morphology, as described earlier [18]. Briefly, thebiofilm slides were divided into 4-mm sections and washed with distilled water, to remove cellular debris, and negatively stained with 0.2% uranyl acetate for 30 s. The slides were carefully mounted on aluminum stubs using carbon adhesive tape, sputter-coated with gold (JEOL-EC-32010CC), and then carefully settled in a sample chamber of SEM (JEOL JSM-IT 100, Japan).
2.9. Statistical Analysis
The data were analyzed using an IBM SPSS 21 (IBM, Inc., Armonk, NY, USA). A Shapiro-Wilk test and Kolmogorov–Simirnov test were applied to test the normal distribution of the data. The statistics of mean and standard deviation were applied. An unpaired t-test (two tailed) was used to access the significance of the data. A p-value of 0.05 or less was considered statistically significant.
3. Results
Fruit concentrate juices have a high percentage of sugar and low pH, and generally provide unfavorable conditions for the growth of bacterial pathogens. In the present study, apple juice concentrate samples of pH 4.5 were analyzed for their total bacterial load on plate count agar at 35 °C. No growth was observed on any dilution after 24 h of incubation. After 48 h of incubation, five colonies were noticed on the1:10 dilution. The number of colonies reached 13 after 72 h of incubation. All of the colonies were identified as Gram-negative short rods and confirmed as Pseudomonas by molecular characterization. Typical colonies on Pseudomonas Cetrimide agar (USP, EP) (e.g., fluorescent green) and Pseudomonas supplemented Agar Base (e.g., straw-colored colonies with green pigment) were indicative of P. aeruginosa, as later confirmed by oprL gene amplification. Furthermore, on plate count agar, two different morphotypes were noticed. One small pinpointed transparent, non-pigmented, and slow growing (SGCP), which appeared after 72 h of incubation; and other normal pigmented, pin-headed, and translucent (NCP) colony phenotypes, which were noticed on PCA after 48 h of incubation at 35 °C. The PCR results showed that both of these phenotypes were positive for oprI and oprL. Pre-enrichment of the sample in lactose broth was performed for better recovery of P. aeruginosa. The number of total colony forming units (CFU) was increased and reached 51 CFU/mL. Out of 51 colonies, 12 were SGCP and 39 were NCP. In order to study the reversion of SGCP to normal colony phenotypes, a continual enrichment procedure was performed. Five-times pre-enrichment in lactose broth (pH = 7.0) resulted in the reversion of SGCP to NCP. This phenomenon was confirmed and validated by the same procedure for every individual colony phenotype. After the disappearance of SCGPs the recovered NCPs were exposed (2.3 × 105 CFU/mL) to similar conditions to their isolation: the same type of sterile undiluted pulp with pH 4.5, and incubated at 4 °C for a week. The total bacterial count on plate count agar was performed every day for a week. The results showed inhibition of the majority of the P. aeruginosa population and only a few cells were able to grow. On day one and day two, samples showed the recovery of only NCPs (9 CFU/mL). Third-day samples showed a mixed growth of NCPs and SGCPs, i.e., 11 CFU/mL and 3 CFU/mL, respectively. On day seven, the NCPs were counted as 29 CFU/mL and the number of SGCPs reached 8 CFU/mL.
3.1. Effect of pH and Time on P. aeruginosa Biofilm Population
Interestingly, the recovery of SGCP was increased in an acidic environment. The maximum number of NCP colonies was observed at pH 8.0 and 9 (Table 1) after 24 h of incubation. The lowest number of NCP colonies was recorded at pH 4.0. Conversely, the maximum number of SGCP was recorded at pH 5.0 and the lowest at pH 7.0 after 24 h of incubation. Moreover, a drastic change in the pH of the media was recorded after 24 h of incubation (Table 1); the pH of TSB broth adjusted to 9.0 changed to pH 6.3. Furthermore, the number of SGCPs increased after acidification of TSB broth (Table 1).

Table 1.
Effect of incubation time and pH on biofilm population.
3.2. Antibiotic Sensitivity
The results of antibiotic sensitivity testing showed that both of these phenotypes were susceptible to the majority of the antibiotics used, i.e., meropenm, levofloxacin, ceftriaxone, amoxycillin, gentamicin, amikacin, ciprofloxacin, ceftazidime, and cefotaxime. However, the imipenem results showed an interesting anomaly. A major population was susceptible and a minority of slow growing phenotypes (SGCP) showed resistance after 48 h of incubation (Figure 1). The sensitivity assay showed that NCPs were unable to grow in the presence of 4 µg/mL of imipenem. However, few pin-pointed colonies appeared after 48 h of incubation. This heterogeneity was a common factor in both phenotypes (NCP and SGCP) of subject isolates of P. aeruginosa. The number and the size of colonies increased with incubation time.

Figure 1.
Imipenem sensitivity assay showing the presence of two different phenotypes of P. aeruginosa. The majority of the cells (NCP) were susceptible and a minor population (SGCP) was resistant, as shown growing inside the zone of inhibition.
3.3. Biofilm Formation
Biofilm formation was noticed after 48 h of incubation in a flask with pH 4.0 and above. Interestingly, population analysis of biofilm consortia also indicated the presence of two colony phenotypes. However, a difference was noticed in the biofilm of NCP and SGCP. SGCP showed biofilm formation at pH 4.0 and above, whereas NCP showed biofilm formation at pH 5.0 and above (Figure 2a,b). The electron micrographs showed the presence of two types of cells in biofilm consortia: small size rods and large size rods (Figure 2c). This size variation was attributed to colony morphotypes. NCP represented the larger ones nearer to the normal size of wild-type P. aeruginosa, and SGCP were the resistant smaller variants.

Figure 2.
Scanning electron micrographs of biofilm consortia. (a) SGCP vibrant biofilm formation at pH 4.0. (b) NCP dense biofilm formation at pH 5.0. (c) The presence of two types of cell in the biofilm consortia of subject isolates: small size rod shape (SGCP) and large size rods (NCP). This size variation was attributed to colony morphotypes.
Biofilm was also noticed at pH 7.0 and 8.0 in both phenotypes; however, the density of biofilms was much higher at lower pH (Table 1). This was confirmed using crystal violet assay (Figure 3). Established biofilms of both phenotypes were equally resistant to imipenem (16 µg/mL) and no signs of cell lysis or biofilm dispersion were observed. Interestingly, the percentage of SGCP increased with imipenem exposure to the established biofilms (Figure 4). The data is a Gaussian distribution and the difference was statistically significant (p < 0.0001).

Figure 3.
Crystal violet (CV) staining of biofilms formed by P. aeruginosa. Cell-bound crystal violet was dissolved in 33% acetic acid. Biofilm thickness (OD) was measured at 570 nm. (a) Slides were placed in 0.1% CV solution for 25 min. (b) Slides were washed twice with 1 mL of sterile saline and dried at room temperature.
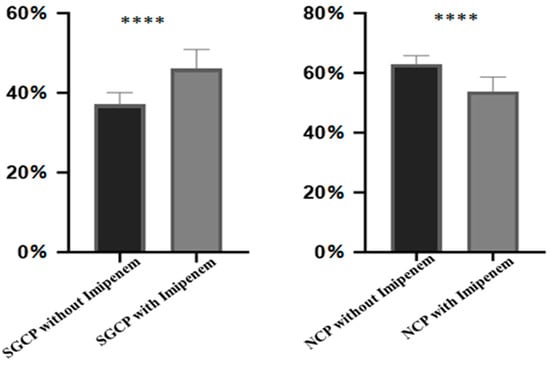
Figure 4.
Bar diagram indicating the percentage of SGCP and NCP under imipenem stress and without imipenem stress on the established biofilms. Statistics of mean and standard deviation were applied to access the degree of shift of phenotype. The level of significance of the data (p < 0.0001) is represented by ****.
4. Discussions
P. aeruginosa is characterized as an opportunistic pathogen and is one of the major causes of food spoilage; especially in fruits and fruit juices, dairy products, fish, and meat products [27,28].Bacteria generally struggle to survive high sugar levels, acidic pH, oxidative stress (Eh), and low water activity (aw) [15,29,30]. However, some pathogens such as P. aeruginosa, not only develop tolerance to such harsh environmental conditions, but also thrive in them [31,32]. They apply different strategies to survive in difficult environmental conditions, and formation of biofilm is one of these tactics. Biofilms are heterogeneous congregations of surface-associated microorganisms, encapsulated within a self-produced polymer matrix, consisting of polysaccharides, protein, and DNA [33,34]. The matrix acts as a physical barrier against antimicrobial compounds and provides a protective ecological niche for the survival of microorganisms. A single species of bacteria or consortia of multispecies microbes can exist within a biofilm. Many microorganisms can build biofilms, e.g., Mycoplasma pneumoniae, Candida albicans, Staphylococcus epidermidis, Mycobacterium tuberculosis, Mycobacterium abscessus, and Staphylococcus aureus [15,35,36,37,38,39]. Biofilms pose serious health concerns, due to the recalcitrant nature of microbes towards antimicrobial drugs and host immune responses. Moreover, biofilms evade the immune system through their exopolymer matrices and arrangement. Immune evasion coupled with multidrug resistance may effectively complement the virulence of a pathogen. Therefore, the prevention and suppression of biofilm formation is of utmost importance [40,41].
The present study exhibited the presence of P. aeruginosa in concentrated fruit juice samples, with a strong potential for biofilm formation, reduced metabolism, and slow growth rate, to survive a low pH and antibiotic stress. They retained the ability for reversion to the normal colony phenotype when the stress was relieved, as reported previously [42,43,44]. P. aeruginosa is also capable of long-term survival without nutrients, by existing in a dormant state [45]. This dormant phenotype not only survived starvation, neutralized toxicity, but also demonstrated resistance against multiple antibacterial agents [46].
The other characteristic feature demonstrated by the subject strain is heterogeneity. This means that a small fraction of the population survived and the majorities of cells either died or were unable to grow in a particular environment [47]. This is a well-established characteristic of pathogens such as methicillin heteroresistant Staphylococcus aureus [48] and Carbapenems heteroresistant Acinetobacter baumannii, Enterobacter cloacae, Klebsiella pneumonia, Escherichia coli, and P. Aeruginosa [49]. Notably, these surviving cells did not produce a noticeable change/deterioration in the quality of the product. This dormancy, a form bet-hedging, is a process used by a wide range of microbial taxa, and refers to an organism’s ability to enter a reversible state of low metabolic activity when faced with unfavorable environmental conditions. Similarly, previous reports showed that starvation, antibiotics, and environmental stressors applied to microbial populations resulted in the induction of persister or dormant cells [45,47,50]. Our study suggested that P. aeruginosa may persist and survive for a long time in the acidic environment of food products. Furthermore, this is a reversible phenotype; they can readopt their normal state in a stress-free environment [48].
The slow growing colony phenotype (SGCP) of P. aeruginosa isolated here exhibited heteroresistance to imipenem; a phenomenon where a phenotype under stress diverged into subpopulations having reduced susceptibility to that particular stress factor as compared with the dominant phenotype [51]. Interestingly, subject isolates of P. aeruginosa were susceptible to all classes of antibiotics, except imipenem. This is an unusual observation, because most antibiotic-resistant P. aeruginosa are multidrug-resistant [2,3,4,7]. We consider this as tolerance to imipenem. In the present situation, the subject population of P. aeruginosa persisted in a dormant state in adherent biofilm until imipenem lost its efficacy or toxicity. Imipenem is one of the most effective antibiotics against P. aeruginosa; however, it is ineffective against SGCP. In the food industry, acidification is a classic method to preserve and protect food products. In addition, high sugar and salt are used to avoid microbial contamination. However, this SGCP survives in all of the above conditions, e.g., acidic pH, high sugar, and also has resistance to imipenem. In all cases, the survival strategy is the same, e.g., slow growth, reduced metabolism, and biofilm formation.
5. Conclusions
The present study confirmed the presence of resistant variants of P. aeruginosa in juice concentrate, with the ability to form dense biofilms on glass surfaces, a reduced metabolism, and slow growth rate, to survive at low pH. These strains have the ability to revert back to their wild type phenotype when stress is relieved. The isolated phenotypes largely depended upon their slow metabolic rate to survive starvation and to neutralize the stress of the environment, i.e., pH 4.0 and high sugar concentration. Both of these phenotypes exhibited heteroresistance to imipenem.
Author Contributions
Conceptualization, F.A., Y.Z., Z.A.M. and M.J.I.; Funding acquisition, Y.Z. and Z.A.M.; Methodology, F.A., Y.Z., Z.A.M., M.J.I., P.N.M., F.Z.K., A.B.K., M.N.K. and Y.L.; Project administration, Y.Z. and Z.A.M.; Resources, Y.Z. and Z.A.M.; Supervision, Y.Z. and Z.A.M.; Writing—original draft, F.A., Y.Z., Z.A.M., M.J.I. and F.Z.K.; Writing—review and editing, M.J.I., P.N.M., A.B.K. and M.N.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was sponsored by the Program of shanghai Academic Research Leader (21XD1401200) and the Agricultural Project of Shanghai Science and Technology Innovation Action Plan, China (19391901600).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors would like to thank FMRRC, Microbiological Analytical Centre, Pakistan Council of Scientific and Industrial Research Laboratories Complex, Karachi, Pakistan, for providing the necessary facilities for the completion of this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bitew, A. High prevalence of multi-drug resistance and extended spectrum beta lactamase production in non-fermenting gram-negative bacilli in Ethiopia. Infect. Dis. Res. Treat. 2019, 12. [Google Scholar] [CrossRef] [PubMed]
- Rehman, A.; Patrick, W.M.; Lamont, I.L. Mechanisms of ciprofloxacin resistance in Pseudomonas aeruginosa: New approaches to an old problem. J. Med. Microbiol. 2019, 68, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Holban, A.-M.; Chifiriuc, M.C.; Cotar, A.I.; Bleotu, C.; Grumezescu, A.M.; Banu, O.; Lazar, V. Virulence markers in Pseudomonas aeruginosa isolates from hospital acquired infections occurred in patients with underlying cardiovascular disease. Rom. Biotechnol. Lett. 2013, 18, 8843–8854. Available online: https://e-repository.org/rbl/vol.18/iss.6/13.pdf (accessed on 8 May 2022).
- Wang, C.; Ye, Q.; Jiang, A.; Zhang, J.; Shang, Y.; Li, F.; Zhou, B.; Xiang, X.; Gu, Q.; Pang, R.; et al. Pseudomonas aeruginosa Detection Using Conventional PCR and Quantitative Real-Time PCR Based on Species-Specific Novel Gene Target Identified by Pangenome Analysis. Front. Microbiol. 2022, 13, 820431. [Google Scholar] [CrossRef]
- Eckmanns, T.; Oppert, M.; Martin, M.; Amorosa, R.; Zuschneid, I.; Frei, U.; Rüden, H.; Weist, K. An outbreak of hospital-acquired Pseudomonas aeruginosa infection caused by contaminated bottled water in intensive care units. Clin. Microbiol. Infect. 2008, 14, 454–458. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.H.; Ahsan, M.; Taylor, W.D.; Kogure, K. Culturability and survival of marine, freshwater and clinical Pseudomonas aeruginosa. Microbes Environ. 2009, 25, 266–274. [Google Scholar] [CrossRef]
- Allydice-Francis, K.; Brown, P.D. Diversity of antimicrobial resistance and virulence determinants in Pseudomonas aeruginosa associated with fresh vegetables. Int. J. Microbiol. 2012, 2012, 426241. [Google Scholar] [CrossRef]
- Haileselassie, M.; Taddele, H.; Adhana, K.; Kalayou, S. Food safety knowledge and practices of abattoir and butchery shops and the microbial profile of meat in Mekelle City, Ethiopia. Asian Pac. J. Trop. Biomed. 2013, 3, 407–412. [Google Scholar] [CrossRef]
- Mikkelsen, H.; Sivaneson, M.; Filloux, A. Key two-component regulatory systems that control biofilm formation in Pseudomonas aeruginosa. Environ. Microbiol. 2011, 13, 1666–1681. [Google Scholar] [CrossRef]
- Mah, T.-F.; Pitts, B.; Pellock, B.; Walker, G.C.; Stewart, P.S.; O’Toole, G.A. A genetic basis for Pseudomonas aeruginosa biofilm antibiotic resistance. Nature 2003, 426, 306–310. [Google Scholar] [CrossRef]
- Ma, L.; Conover, M.; Lu, H.; Parsek, M.R.; Bayles, K.; Wozniak, D.J. Assembly and development of the Pseudomonas aeruginosa biofilm matrix. PLoS Pathog. 2009, 5, e1000354. [Google Scholar] [CrossRef] [PubMed]
- Jennings, L.K.; Storek, K.M.; Ledvina, H.E.; Coulon, C.; Marmont, L.S.; Sadovskaya, I.; Secor, P.R.; Tseng, B.S.; Scian, M.; Filloux, A. Pel is a cationic exopolysaccharide that cross-links extracellular DNA in the Pseudomonas aeruginosa biofilm matrix. Proc. Natl. Acad. Sci. USA 2015, 112, 11353–11358. [Google Scholar] [CrossRef]
- Meirelles, L.A.; Newman, D.K. Both toxic and beneficial effects of pyocyanin contribute to the lifecycle of Pseudomonas aeruginosa. Mol. Microbiol. 2018, 110, 995–1010. [Google Scholar] [CrossRef]
- Wickramasinghe, N.N.; Hlaing, M.M.; Ravensdale, J.T.; Coorey, R.; Chandry, P.S.; Dykes, G.A. Characterization of the biofilm matrix composition of psychrotrophic, meat spoilage pseudomonads. Sci. Rep. 2020, 10, 16457. [Google Scholar] [CrossRef]
- Chan, M.W.H.; Mirani, Z.A.; Khan, M.N.; Ali, A.; Khan, A.B.; Rauf, N. Isolation and characterization of small colony variants of Staphylococcus aureus in various food samples. Biocatal. Agric. Biotechnol. 2021, 35, 102097. [Google Scholar] [CrossRef]
- Berlanga, M.; Guerrero, R. Living together in biofilms: The microbial cell factory and its biotechnological implications. Microb. Cell Factories 2016, 15, 165. [Google Scholar] [CrossRef] [PubMed]
- Costerton, J.W.; Cheng, K.; Geesey, G.G.; Ladd, T.I.; Nickel, J.C.; Dasgupta, M.; Marrie, T.J. Bacterial biofilms in nature and disease. Annu. Rev. Microbiol. 1987, 41, 435–464. [Google Scholar] [CrossRef]
- Moradali, M.F.; Ghods, S.; Rehm, B.H. Pseudomonas aeruginosa lifestyle: A paradigm for adaptation, survival and persistence. Front. Cell. Infect. Microbiol. 2017, 7, 39. [Google Scholar] [CrossRef]
- Häußler, S.; Tümmler, B.; Weißbrodt, H.; Rohde, M.; Steinmetz, I. Small-colony variants of Pseudomonas aeruginosa in cystic fibrosis. Arch. Clin. Infect. Dis. 1999, 29, 621–625. [Google Scholar] [CrossRef]
- Starkey, M.; Hickman, J.H.; Ma, L.; Zhang, N.; De Long, S.; Hinz, A.; Palacios, S.; Manoil, C.; Kirisits, M.J.; Starner, T.D. Pseudomonas aeruginosa rugose small-colony variants have adaptations that likely promote persistence in the cystic fibrosis lung. J. Bacteriol. Res. 2009, 191, 3492–3503. [Google Scholar] [CrossRef]
- Wilton, M.; Charron-Mazenod, L.; Moore, R.; Lewenza, S. Extracellular DNA acidifies biofilms and induces aminoglycoside resistance in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2016, 60, 544–553. [Google Scholar] [CrossRef] [PubMed]
- Gholami, A.; Majidpour, A.; Talebi-Taher, M.; Boustanshenas, M.; Adabi, M. PCR-based assay for the rapid and precise distinction of Pseudomonas aeruginosa from other Pseudomonas species recovered from burns patients. J. Prev. Med. Hyg. 2016, 57, E81. [Google Scholar] [CrossRef] [PubMed]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 30th ed.; CLSI Supplement M100; Clinical and Laboratory Standard Institute: Wayne, PA, USA, 2020. [Google Scholar]
- O’Toole, G.A. Microtiter dish biofilm formation assay. J. Vis. Exp. 2011, 47, e2437. [Google Scholar] [CrossRef] [PubMed]
- Ullah, A.; Yin, X.; Wang, F.; Xu, B.; Mirani, Z.A.; Xu, B.; Chan, M.W.H.; Ali, A.; Usman, M.; Ali, N. Biosynthesis of Selenium Nanoparticles (via Bacillus subtilis BSN313), and Their Isolation, Characterization, and Bioactivities. Molecules 2021, 26, 5559. [Google Scholar] [CrossRef]
- Crompton, M.J.; Dunstan, R.H.; Macdonald, M.M.; Gottfries, J.; von Eiff, C.; Roberts, T.K. Small changes in environmental parameters lead to alterations in antibiotic resistance, cell morphology and membrane fatty acid composition in Staphylococcus lugdunensis. PLoS ONE 2014, 9, e92296. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Xie, J.; Soteyome, T.; Peters, B.M.; Shirtliff, M.E.; Liu, J.; Harro, J.M. Polymicrobial interaction and biofilms between Staphylococcus aureus and Pseudomonas aeruginosa: An underestimated concern in food safety. Curr. Opin. Food Sci. 2019, 26, 57–64. [Google Scholar] [CrossRef]
- Aman, M.; Aneeqha, N.; Bristi, K.; Deeksha, J.; Afza, N.; Sindhuja, V.; Shastry, R.P. Lactic acid bacteria inhibits quorum sensing and biofilm formation of Pseudomonas aeruginosa strain JUPG01 isolated from rancid butter. Biocatal. Agric. Biotechnol. 2021, 36, 102115. [Google Scholar] [CrossRef]
- Broughall, J.; Anslow, P.; Kilsby, D. Hazard analysis applied to microbial growth in foods: Development of mathematical models describing the effect of water activity. J. Appl. Bacteriol. 1983, 55, 101–110. [Google Scholar] [CrossRef]
- Cotter, P.D.; Hill, C. Surviving the acid test: Responses of gram-positive bacteria to low pH. Microbiol. Mol. Biol. Rev. 2003, 67, 429–453. [Google Scholar] [CrossRef]
- Kaprelyants, A.S.; Gottschal, J.C.; Kell, D.B. Dormancy in non-sporulating bacteria. FEMS Microbiol. Lett. 1993, 104, 271–285. [Google Scholar] [CrossRef]
- Møretrø, T.; Langsrud, S. Residential bacteria on surfaces in the food industry and their implications for food safety and quality. Rev. Food Sci. Food Saf. 2017, 16, 1022–1041. [Google Scholar] [CrossRef]
- Jefferson, K.K. What drives bacteria to produce a biofilm? FEMS Microbiol. Lett. 2004, 236, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Alam, A.; Rani, M.; Ehtesham, N.Z.; Hasnain, S.E. Biofilms: Survival and defense strategy for pathogens. Int. J. Med. Microbiol. 2017, 307, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Fennelly, K.P.; Ojano-Dirain, C.; Yang, Q.; Liu, L.; Lu, L.; Progulske-Fox, A.; Wang, G.P.; Antonelli, P.; Schultz, G. Biofilm formation by Mycobacterium abscessus in a lung cavity. Am. J. Respir. Crit. Care Med. 2016, 193, 692–693. [Google Scholar] [CrossRef]
- Fey, P.D.; Olson, M.E. Current concepts in biofilm formation of Staphylococcus epidermidis. Future Microbiol. 2010, 5, 917–933. [Google Scholar] [CrossRef]
- Kornspan, J.D.; Tarshis, M.; Rottem, S. Adhesion and biofilm formation of Mycoplasma pneumoniae on an abiotic surface. Arch. Microbiol. 2011, 193, 833–836. [Google Scholar] [CrossRef]
- Pang, J.M.; Layre, E.; Sweet, L.; Sherrid, A.; Moody, D.B.; Ojha, A.; Sherman, D.R. The polyketide Pks1 contributes to biofilm formation in Mycobacterium tuberculosis. J. Bacteriol. Res. 2012, 194, 715–721. [Google Scholar] [CrossRef] [PubMed]
- Ramage, G.; Walle, K.V.; Wickes, B.L.; Lopez-Ribot, J.L. Characteristics of biofilm formation by Candida albicans. Rev. Iberoam. Micol. 2001, 18, 163–170. Available online: https://pubmed.ncbi.nlm.nih.gov/15496122/ (accessed on 17 August 2022). [PubMed]
- Davey, M.E.; O’Toole, G.A. Microbial biofilms: From ecology to molecular genetics. Microbiol. Mol. Biol. Rev. 2000, 64, 847–867. [Google Scholar] [CrossRef]
- Shevchenko, S.N.; Burkhardt, M.; Sheval, E.V.; Natashina, U.A.; Grosse, C.; Nikolaev, A.L.; Gopin, A.V.; Neugebauer, U.; Kudryavtsev, A.A.; Sivakov, V. Antimicrobial effect of biocompatible silicon nanoparticles activated using therapeutic ultrasound. Langmuir 2017, 33, 2603–2609. [Google Scholar] [CrossRef]
- Frimmersdorf, E.; Horatzek, S.; Pelnikevich, A.; Wiehlmann, L.; Schomburg, D. How Pseudomonas aeruginosa adapts to various environments: A metabolomic approach. Environ. Microbiol. 2010, 12, 1734–1747. [Google Scholar] [CrossRef] [PubMed]
- Gray, D.A.; Dugar, G.; Gamba, P.; Strahl, H.; Jonker, M.J.; Hamoen, L.W. Extreme slow growth as alternative strategy to survive deep starvation in bacteria. Nat. Commun. 2019, 10, 890. [Google Scholar] [CrossRef]
- Moriarty, T.; Elborn, J.; Tunney, M. Effect of pH on the antimicrobial susceptibility of planktonic and biofilm-grown clinical Pseudomonas aeruginosa isolates. Br. J. Biomed. Sci. 2007, 64, 101–104. [Google Scholar] [CrossRef] [PubMed]
- Lewenza, S.; Abboud, J.; Poon, K.; Kobryn, M.; Humplik, I.; Bell, J.R.; Mardan, L.; Reckseidler-Zenteno, S. Pseudomonas aeruginosa displays a dormancy phenotype during long-term survival in water. PLoS ONE 2018, 13, e0198384. [Google Scholar] [CrossRef] [PubMed]
- Workentine, M.L.; Sibley, C.D.; Glezerson, B.; Purighalla, S.; Norgaard-Gron, J.C.; Parkins, M.D.; Rabin, H.R.; Surette, M.G. Phenotypic heterogeneity of Pseudomonas aeruginosa populations in a cystic fibrosis patient. PLoS ONE 2013, 8, e60225. [Google Scholar] [CrossRef]
- Lewis, K. Persister cells, dormancy and infectious disease. Nat. Rev. Microbiol. 2007, 5, 48–56. [Google Scholar] [CrossRef]
- Huang, S.-H.; Chen, Y.-C.; Chuang, Y.-C.; Chiu, S.-K.; Fung, C.-P.; Lu, P.-L.; Wang, L.-S.; Wu, T.-L.; Wang, J.-T. Prevalence of vancomycin-intermediate Staphylococcus aureus (VISA) and heterogeneous VISA among methicillin-resistant S. aureus with high vancomycin minimal inhibitory concentrations in Taiwan: A multicenter surveillance study, 2012–2013. J. Microbiol. Immunol. Infect. 2016, 49, 701–707. [Google Scholar] [CrossRef][Green Version]
- Quale, J.; Shah, N.; Kelly, P.; Babu, E.; Backer, M.; Rosas-Garcia, G.; Salamera, J.; George, A.; Bratu, S.; Landman, D. Activity of polymyxin B and the novel polymyxin analogue CB-182,804 against contemporary Gram-negative pathogens in New York City. Microb. Drug Resist. 2012, 18, 132–136. [Google Scholar] [CrossRef]
- Lennon, J.T.; Jones, S.E. Microbial seed banks: The ecological and evolutionary implications of dormancy. Nat. Rev. Microbiol. 2011, 9, 119–130. [Google Scholar] [CrossRef]
- Xu, Y.; Zheng, X.; Zeng, W.; Chen, T.; Liao, W.; Qian, J.; Lin, J.; Zhou, C.; Tian, X.; Cao, J. Mechanisms of heteroresistance and resistance to imipenem in Pseudomonas aeruginosa. Infect. Drug Resist. 2020, 13, 1419. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).