
Cortical Activation in Response to Speech Differs between Prelingually Deafened Cochlear Implant Users with Good or Poor Speech-in-Noise Understanding: An fNIRS Study


Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Sentence-in-Noise Test
2.3. CI simulation
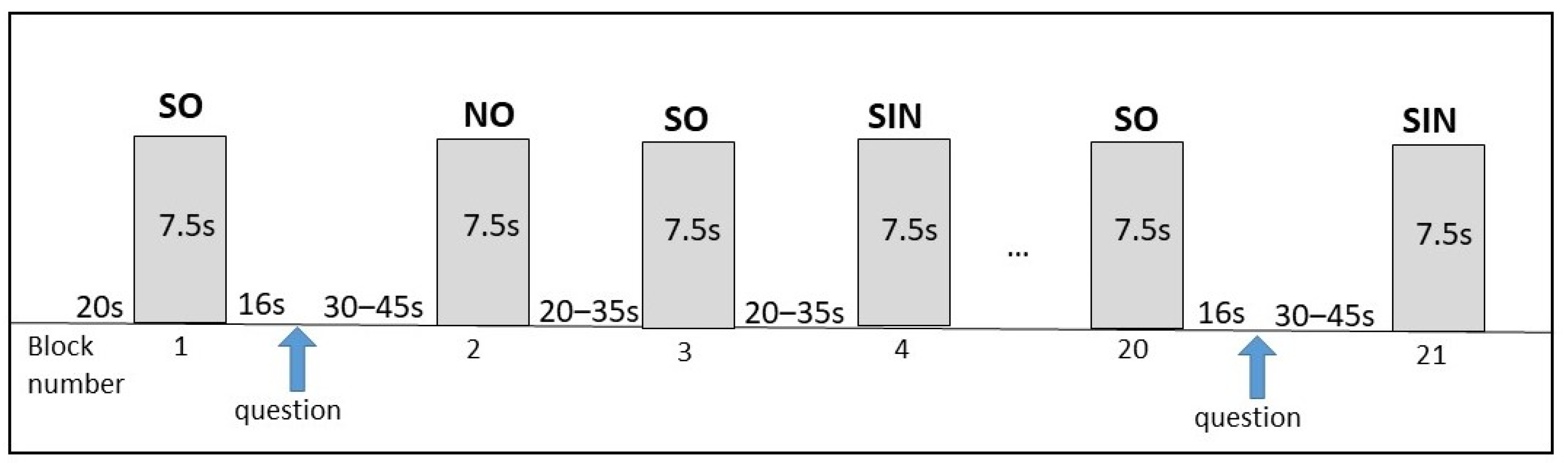
2.4. fNIRS Paradigm
2.5. fNIRS Data Collection
2.6. Study Design
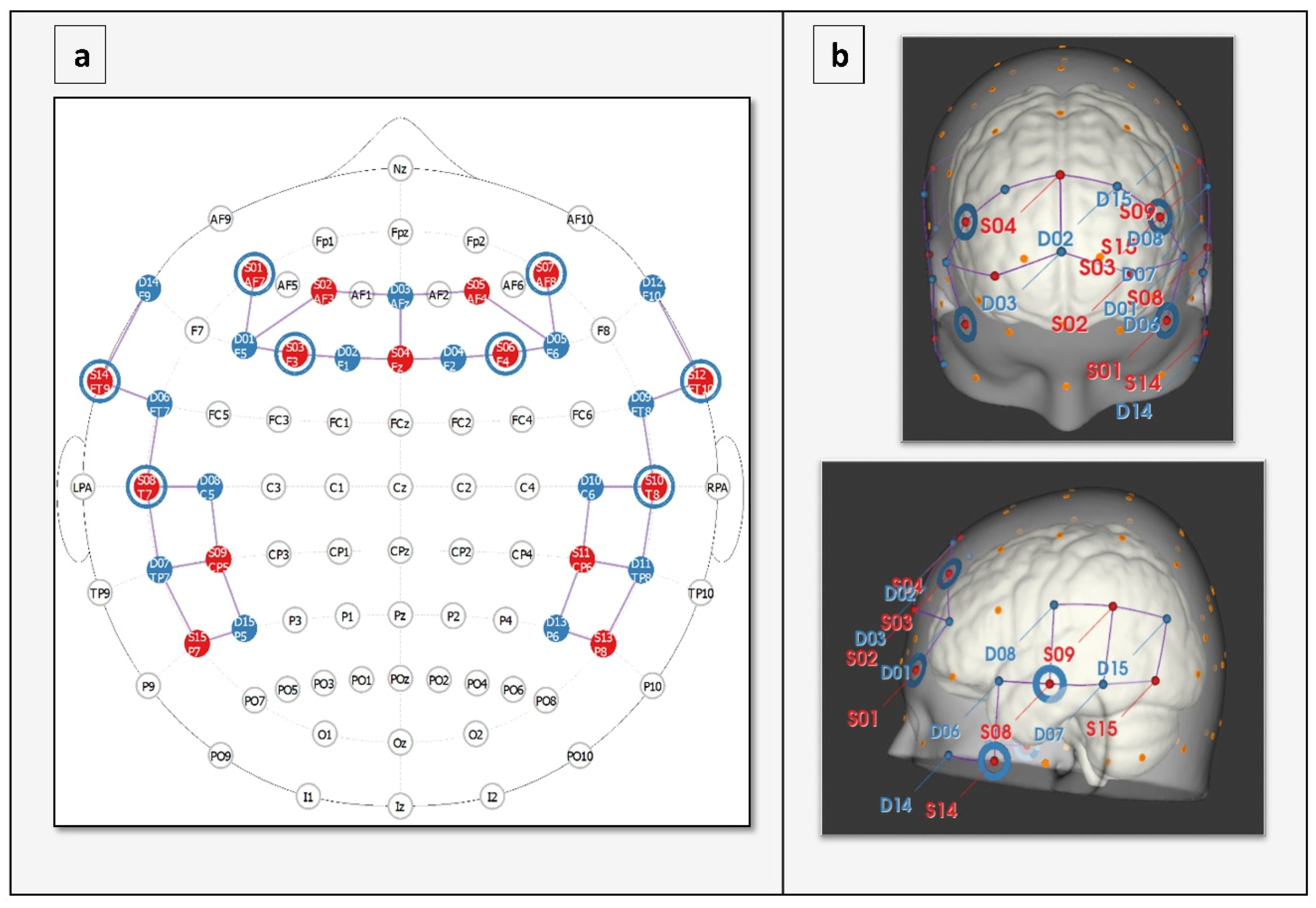
2.7. Apparatus
2.8. fNIRS Data Analysis
Signal Processing
2.9. Statistical Analysis
3. Results
3.1. Behavioral Data
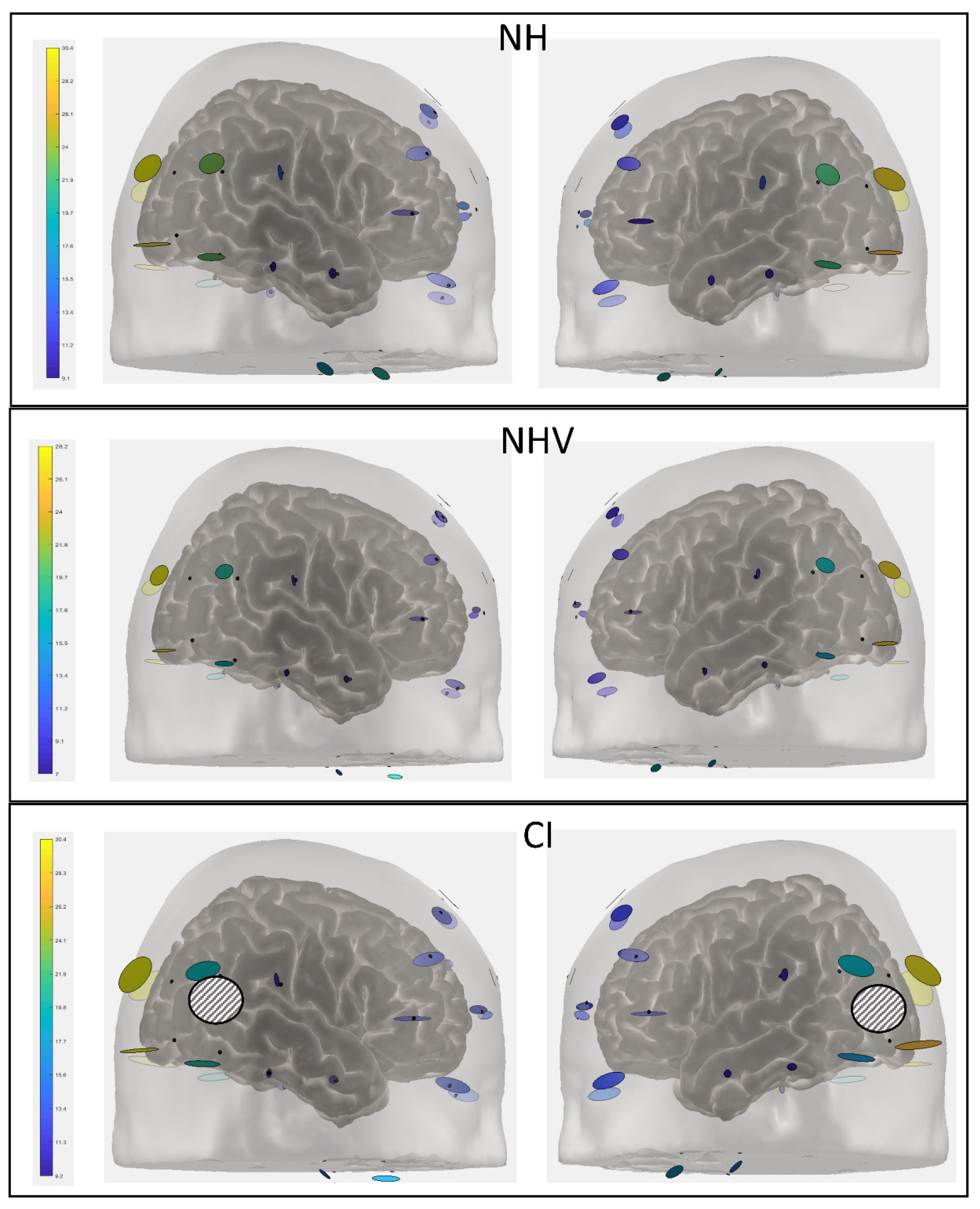
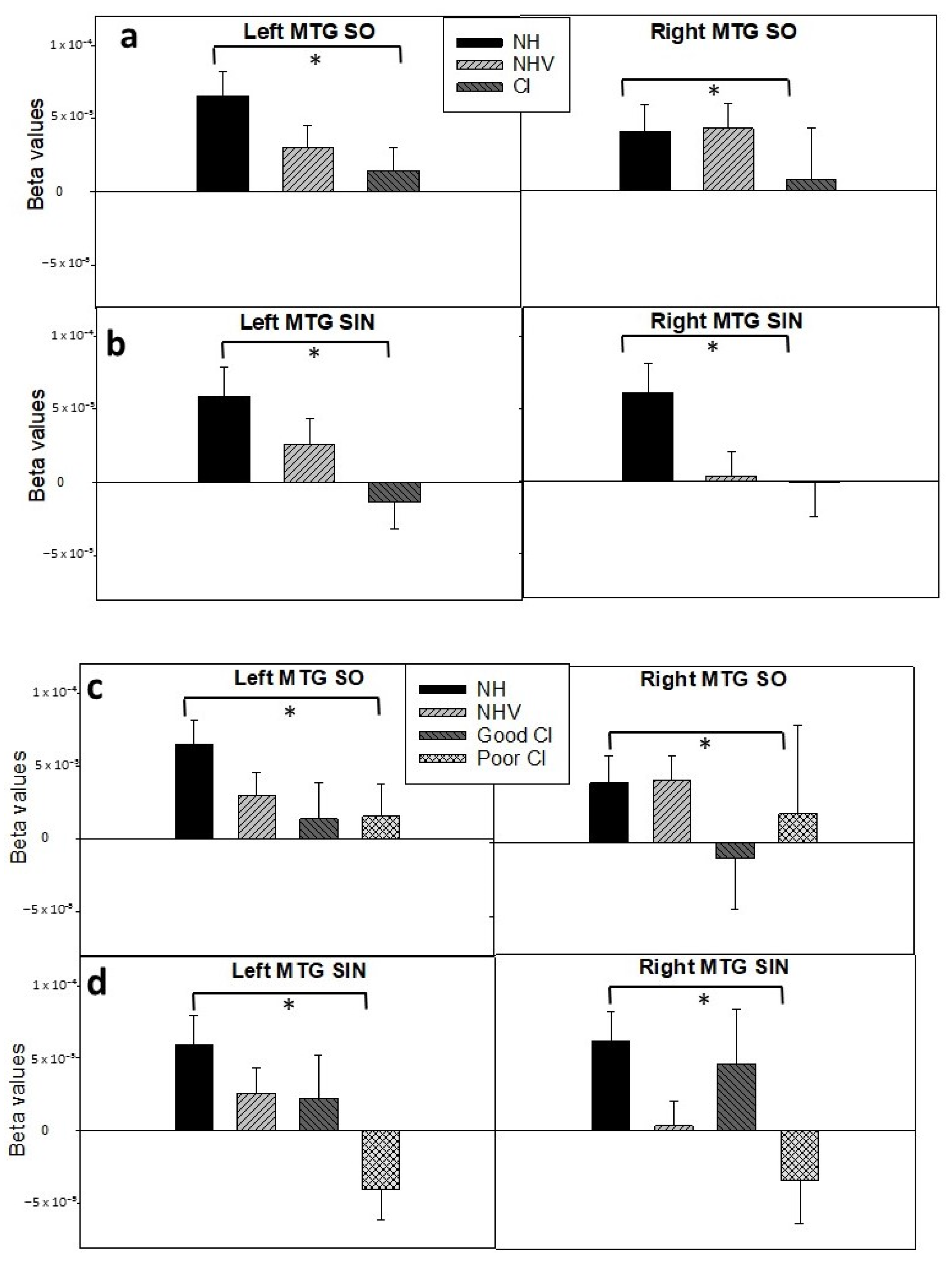
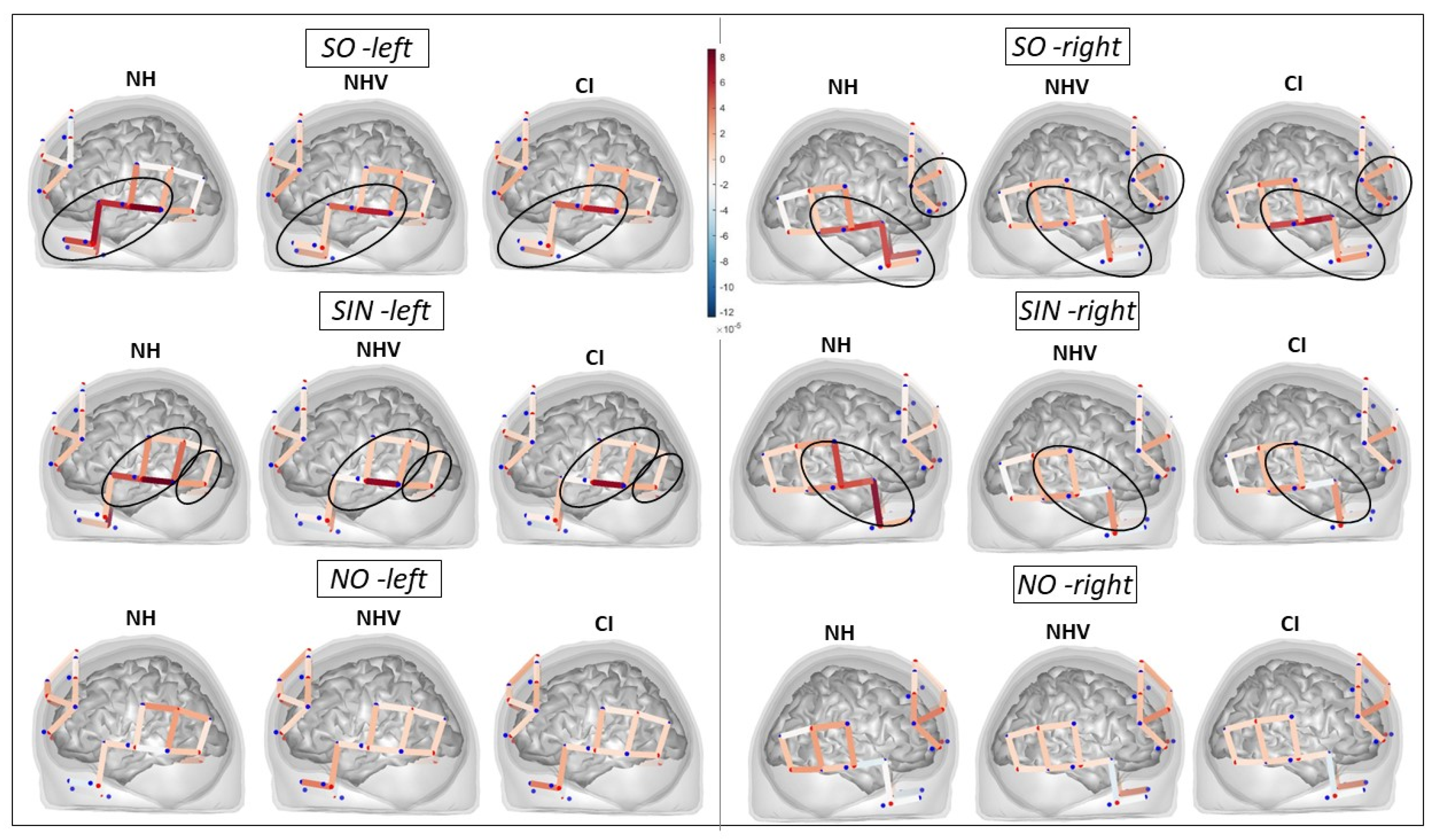
3.2. fNIRS: HbO Data
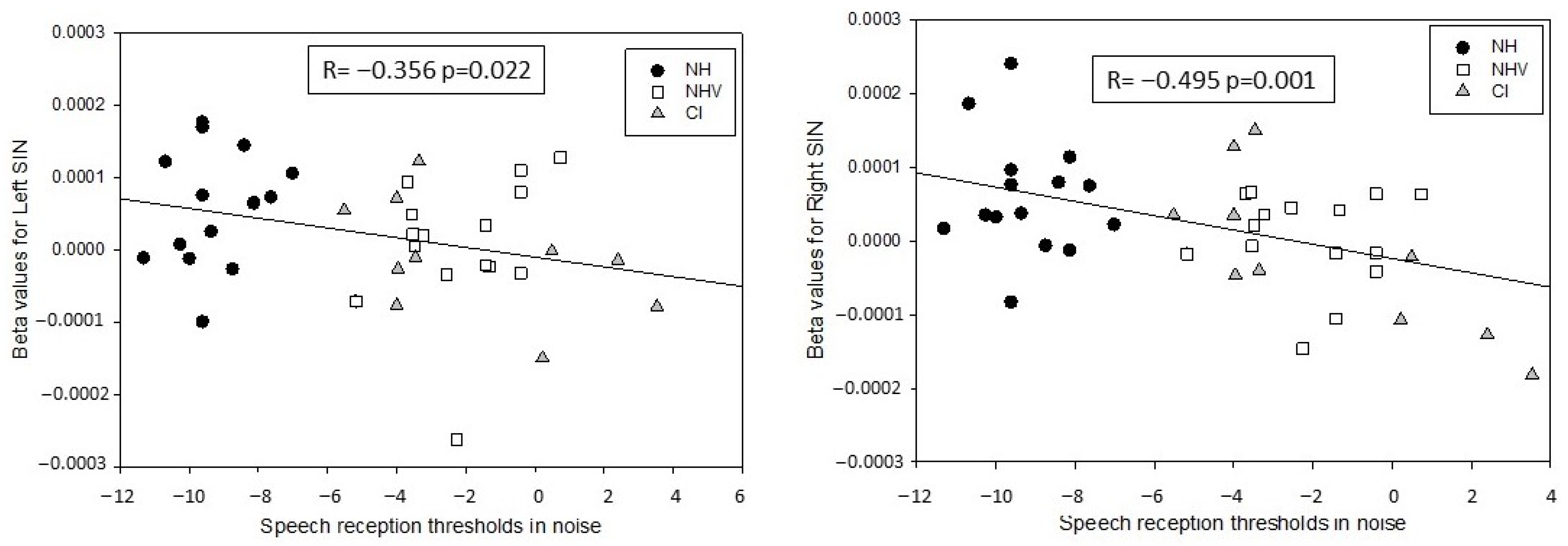
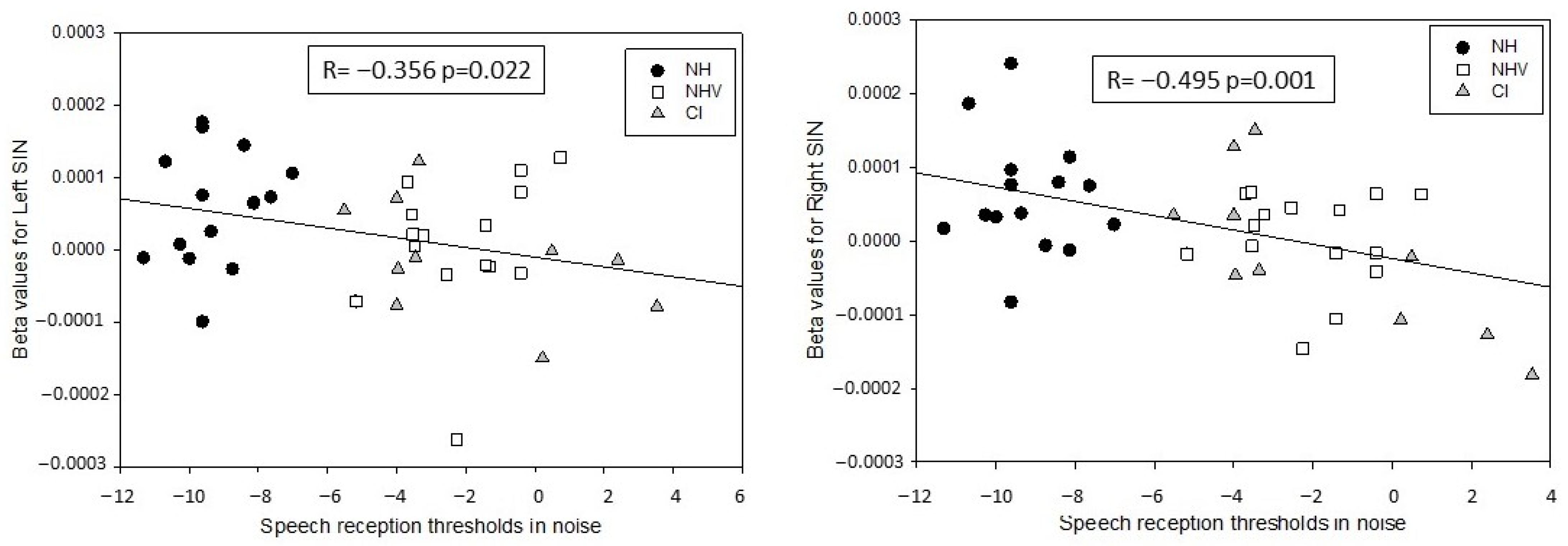
3.3. Explaining Factors for the Results
3.4. fNIRS: HbR Data
4. Discussion
Limitations and Suggestions for Future Studies
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hochmair-Desoyer, I.; Schulz, E.; Moser, L.; Schmidt, M. The HSM sentence test as a tool for evaluating the speech understanding in noise of cochlear implant users. Am. J. Otolaryngol. 1997, 18, 83. [Google Scholar]
- Caldwell, A.; Nittrouer, S. Speech perception in noise by children with cochlear implants. J. Speech Lang. Hear. Res. 2013, 56, 13–30. [Google Scholar] [CrossRef] [Green Version]
- Eisenberg, L.S.; Fisher, L.M.; Johnson, K.C.; Ganguly, D.H.; Grace, T.; Niparko, J.K.; Team, C.I. Sentence recognition in quiet and noise by pediatric cochlear implant users: Relationships to spoken language. Otol. Neurotol. 2016, 37, e75–e81. [Google Scholar] [CrossRef] [Green Version]
- Ching, T.Y.; Zhang, V.W.; Flynn, C.; Burns, L.; Button, L.; Hou, S.; McGhie, K.; Van Buynder, P. Factors influencing speech perception in noise for 5-year-old children using hearing aids or cochlear implants. Int. J. Audiol. 2018, 57, S70–S80. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.K.; Boddupally, S.P. Auditory cognitive training for pediatric cochlear implant recipients. Ear Hear. 2018, 39, 48–59. [Google Scholar] [CrossRef] [PubMed]
- Bugannim, Y.; Roth, D.A.E.; Zechoval, D.; Kishon-Rabin, L. Training of speech perception in noise in pre-lingual hearing impaired adults with cochlear implants compared to normal hearing adults. Otol. Neurotol. 2019, 40, e316–e325. [Google Scholar] [CrossRef] [PubMed]
- Zaltz, Y.; Bugannim, Y.; Zechoval, D.; Kishon-Rabin, L.; Perez, R. Listening in noise remains a significant challenge for cochlear implant users: Evidence from early deafened and those with progressive hearing loss compared to peers with normal hearing. J. Clin. Med. 2020, 9, 1381. [Google Scholar] [CrossRef] [PubMed]
- Fu, Q.J.; Galvin, J.J. Maximizing cochlear implant patients’ performance with advanced speech training procedures. Hear. Res. 2008, 242, 198–208. [Google Scholar] [CrossRef] [Green Version]
- Kronenberger, W.G.; Colson, B.G.; Henning, S.C.; Pisoni, D.B. Executive functioning and speech-language skills following long-term use of cochlear implants. J. Deaf Stud. Deaf Educ. 2014, 19, 456–470. [Google Scholar] [CrossRef] [Green Version]
- Dorman, M.F.; Gifford, R.H. Speech understanding in complex listening environments by listeners fit with cochlear implants. J. Speech Lang. Hear. Res. 2017, 60, 3019–3026. [Google Scholar] [CrossRef] [Green Version]
- Geers, A.E. Speech, language, and reading skills after early cochlear implantation. Arch. Otolaryngol. Head Neck Surg. 2004, 130, 634–638. [Google Scholar] [CrossRef]
- Svirsky, M.A.; Teoh, S.W.; Neuburger, H. Development of language and speech perception in congenitally, profoundly deaf children as a function of age at cochlear implantation. Audiol. Neurootol. 2004, 9, 224–233. [Google Scholar] [CrossRef] [PubMed]
- Akeroyd, M.A. Are individual differences in speech reception related to individual differences in cognitive ability? A survey of twenty experimental studies with normal and hearing-impaired adults. Int. J. Audiol. 2008, 47, S53–S71. [Google Scholar] [CrossRef] [PubMed]
- Rudner, M.; Foo, C.; Sundewall-Thorén, E.; Lunner, T.; Rönnberg, J. Phonological mismatch and explicit cognitive processing in a sample of 102 hearing-aid users. Int. J. Audiol. 2008, 47, S91–S98. [Google Scholar] [CrossRef] [PubMed]
- Lunner, T.; Rudner, M.; Rönnberg, J. Cognition and hearing aids. Scand. J. Psychol. 2009, 50, 395–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moberly, A.C.; Bates, C.; Harris, M.S.; Pisoni, D.B. The Enigma of Poor Performance by Adults with Cochlear Implants. Otol. Neurotol. 2016, 37, 1522–1528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davidson, L.S.; Geers, A.E.; Uchanski, R.M.; Firszt, J.B. Effects of early acoustic hearing on speech perception and language for pediatric cochlear implant recipients. J. Speech Lang. Hear. Res. 2019, 62, 3620–3637. [Google Scholar] [CrossRef] [PubMed]
- Pasley, B.N.; David, S.V.; Mesgarani, N.; Flinker, A.; Shamma, S.A.; Crone, N.E.; Knight, R.T.; Chang, E.F. Reconstructing speech from human auditory cortex. PLoS Biol. 2012, 10, e1001251. [Google Scholar] [CrossRef] [Green Version]
- Steinschneider, M.; Nourski, K.V.; Rhone, A.E.; Kawasaki, H.; Oya, H.; Howard, M.A., III. Differential activation of human core, non-core and auditory-related cortex during speech categorization tasks as revealed by intracranial recordings. Front. Neurosci. 2014, 8, 240. [Google Scholar] [CrossRef] [Green Version]
- Gaab, N.; Gabrieli, J.D.; Glover, G.H. Assessing the influence of scanner background noise on auditory processing. I. An fmri study comparing three experimental designs with varying degrees of scanner noise. Hum. Brain Mapp. 2007, 28, 703–720. [Google Scholar] [CrossRef] [PubMed]
- Scarff, C.J.; Dort, J.C.; Eggermont, J.J.; Goodyear, B.G. The effect of mr scanner noise on auditory cortex activity using fMRI. Hum. Brain Mapp. 2004, 22, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Deprez, H.; Gransier, R.; Hofmann, M.; van Wieringen, A.; Wouters, J.; Moonen, M. Independent component analysis for cochlear implant artifacts attenuation from electrically evoked auditory steady-state response measurements. J. Neural Eng. 2017, 15, 016006. [Google Scholar] [CrossRef]
- BinKhamis, G.; Perugia, E.; O’Driscoll, M.; Kluk, K. Speech-abrs in cochlear implant recipients: Feasibility study. Int. J. Audiol. 2019, 58, 678–684. [Google Scholar] [CrossRef] [Green Version]
- Fujiki, N.; Naito, Y.; Hirano, S.; Kojima, H.; Shiomi, Y.; Nishizawa, S.; Konishi, J.; Honjo, I. Correlation between rcbf and speech perception in cochlear implant users. Auris Nasus Larynx 1999, 26, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Green, K.M.J.; Julyan, P.J.; Hastings, D.L.; Ramsden, R.T. Auditory cortical activation and speech perception in cochlear implant users: Effects of implant experience and duration of deafness. Hear. Res. 2005, 205, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Mortensen, M.V.; Mirz, F.; Gjedde, A. Restored speech comprehension linked to activity in left inferior prefrontal and right temporal cortices in postlingual deafness. Neuroimage 2006, 31, 842–852. [Google Scholar] [CrossRef]
- Boas, D.A.; Elwell, C.E.; Ferrari, M.; Taga, G. Twenty years of functional near-infrared spectroscopy: Introduction for the special issue. NeuroImage 2014, 85, 1–5. [Google Scholar] [CrossRef]
- Wiggins, I.M.; Anderson, C.A.; Kitterick, P.T.; Hartley, D.E. Speech-evoked activation in adult temporal cortex measured using functional near-infrared spectroscopy (fNIRS): Are the measurements reliable? Hear. Res. 2016, 339, 142–154. [Google Scholar] [CrossRef]
- Pinti, P.; Tachtsidis, I.; Hamilton, A.; Hirsch, J.; Aichelburg, C.; Gilbert, S.; Burgess, P.W. The present and future use of functional near-infrared spectroscopy (fNIRS) for cognitive neuroscience. Ann. N. Y. Acad. Sci. 2020, 1464, 5–29. [Google Scholar] [CrossRef] [PubMed]
- Pollonini, L.; Olds, C.; Abaya, H.; Bortfeld, H.; Beauchamp, M.S.; Oghalai, J.S. Auditory cortex activation to natural speech and simulated cochlear implant speech measured with functional near-infrared spectroscopy. Hear. Res. 2014, 309, 84–93. [Google Scholar] [CrossRef] [Green Version]
- Olds, C.; Pollonini, L.; Abaya, H.; Larky, J.; Loy, M.; Bortfeld, H.; Beauchamp, M.S.; Oghalai, J.S. Cortical Activation Patterns Correlate with Speech Understanding After Cochlear Implantation. Ear Hear. 2016, 37, e160–e172. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Seghouane, A.K.; Shah, A.; Innes-Brown, H.; Cross, W.; Litovsky, R.; McKay, C.M. Cortical speech processing in postlingually deaf adult cochlear implant users, as revealed by functional near-infrared spectroscopy. Trends Hear. 2018, 22, 2331216518786850. [Google Scholar] [CrossRef]
- Butler, L.K.; Kiran, S.; Tager-Flusberg, H. Functional near-infrared spectroscopy in the study of speech and language impairment across the life span: A systematic review. Am. J. Speech Lang. Pathol. 2020, 29, 1674–1701. [Google Scholar] [CrossRef]
- Saliba, J.; Bortfeld, H.; Levitin, D.J.; Oghalai, J.S. Functional near-infrared spectroscopy for neuroimaging in cochlear implant recipients. Hear. Res. 2016, 338, 64–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, C.A.; Lazard, D.S.; Hartley, D.E. Plasticity in bilateral superior temporal cortex: Effects of deafness and cochlear implantation on auditory and visual speech processing. Hear. Res. 2017, 343, 138–149. [Google Scholar] [CrossRef] [PubMed]
- Basura, G.J.; Hu, X.S.; Juan, J.S.; Tessier, A.M.; Kovelman, I. Human central auditory plasticity: A review of functional near-infrared spectroscopy (fNIRS) to measure cochlear implant performance and tinnitus perception. Laryngoscope Investig. Otolaryngol. 2018, 3, 463–472. [Google Scholar] [CrossRef] [Green Version]
- Bortfeld, H. Functional near-infrared spectroscopy as a tool for assessing speech and spoken language processing in pediatric and adult cochlear implant users. Dev. Psychobiol. 2019, 61, 430–443. [Google Scholar] [CrossRef]
- Mushtaq, F.; Wiggins, I.M.; Kitterick, P.T.; Anderson, C.A.; Hartley, D.E.H. The Benefit of Cross-Modal Reorganization on Speech Perception in Pediatric Cochlear Implant Recipients Revealed Using Functional Near-Infrared Spectroscopy. Front. Hum. Neurosci. 2020, 14, 308. [Google Scholar] [CrossRef]
- Anderson, C.A.; Wiggins, I.M.; Kitterick, P.T.; Hartley, D.E.H. Pre-operative Brain Imaging Using Functional Near-Infrared Spectroscopy Helps Predict Cochlear Implant Outcome in Deaf Adults. J. Assoc. Res. Otolaryngol. 2019, 20, 511–528. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.-C.; Sandmann, P.; Thorne, J.D.; Bleichner, M.G.; Debener, S. Cross-modal functional reorganization of visual and auditory cortex in adult cochlear implant users identified with fnirs. Neural Plast. 2016, 2016, 4382656. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.-C.; Puschmann, S.; Debener, S. Increased cross-modal functional connectivity in cochlear implant users. Sci. Rep. 2017, 7, 10043. [Google Scholar] [CrossRef]
- Defenderfer, J.; Kerr-German, A.; Hedrick, M.; Buss, A.T. Investigating the role of temporal lobe activation in speech perception accuracy with normal hearing adults: An event-related fNIRS study. Neuropsychologia 2017, 106, 31–41. [Google Scholar] [CrossRef]
- Lawrence, R.J.; Wiggins, I.M.; Anderson, C.A.; Davies-Thompson, J.; Hartley, D.E.H. Cortical correlates of speech intelligibility measured using functional nearinfrared spectroscopy (fNIRS). Hear. Res. 2018, 370, 53–64. [Google Scholar] [CrossRef]
- Lawrence, R.J.; Wiggins, I.M.; Hodgson, J.C.; Hartley, D.E.H. Evaluating cortical responses to speech in children: A functional near-infrared spectroscopy (fNIRS) study. Hear. Res. 2021, 401, 108155. [Google Scholar] [CrossRef] [PubMed]
- Wijayasiri, P.; Hartley, D.E.H.; Wiggins, I.M. Brain activity underlying the recovery of meaning from degraded speech: A functional near-infrared spectroscopy (fNIRS) study. Hear. Res. 2017, 351, 55–67. [Google Scholar] [CrossRef]
- Zhou, X.; Sobczak, G.S.; McKay, C.M.; Litovsky, R.Y. Effects of degraded speech processing and binaural unmasking investigated using functional near-infrared spectroscopy (fNIRS). PLoS ONE 2022, 17, e0267588. [Google Scholar] [CrossRef]
- Defenderfer, J.; Forbes, S.; Wijeakumar, S.; Hedrick, M.; Plyler, P.; Buss, A.T. Frontotemporal activation differs between perception of simulated cochlear implant speech and speech in background noise: An image-based fNIRS study. Neuroimage 2021, 240, 118385. [Google Scholar] [CrossRef]
- Bisconti, S.; Shulkin, M.; Hu, X.; Basura, G.J.; Kileny, P.R.; Kovelman, I. Functional near-infrared spectroscopy brain imaging investigation of phonological awareness and passage comprehension abilities in adult recipients of cochlear implants. J. Speech Lang. Hear. Res. 2016, 59, 239–253. [Google Scholar] [CrossRef] [PubMed]
- Van de Rijt, L.P.; van Opstal, A.J.; Mylanus, E.A.; Straatman, L.V.; Hu, H.Y.; Snik, A.F.; van Wanrooij, M.M. Temporal cortex activation to audiovisual speech in normal-hearing and cochlear implant users measured with functional near-infrared spectroscopy. Front. Hum. Neurosci. 2016, 10, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.-C.; Stropahl, M.; Schönwiesner, M.; Debener, S. Enhanced visual adaptation in cochlear implant users revealed by concurrent eeg-fnirs. Neuroimage 2017, 146, 600–608. [Google Scholar] [CrossRef]
- Anderson, S.; Kraus, N. Sensory-cognitive interaction in the neural encoding of speech in noise: A review. J. Am. Acad. Audiol. 2010, 21, 575–585. [Google Scholar] [CrossRef]
- ANSI, S 3.6; 2018 Specification for Audiometers. ANSI: New York, NY, USA, 2018.
- Levitt, H. Transformed up-down methods in psychoacoustics. J. Acoust. Soc. Am. 1971, 49, 467–477. [Google Scholar] [CrossRef]
- Zaltz, Y.; Goldsworthy, R.L.; Kishon-Rabin, L.; Eisenberg, L.S. Voice discrimination by adults with cochlear implants: The benefits of early implantation for vocal-tract length perception. J. Assoc. Res. Otolaryngol. JARO 2018, 19, 193–209. [Google Scholar] [CrossRef]
- Zaltz, Y.; Goldsworthy, R.L.; Eisenberg, L.S.; Kishon-Rabin, L. Children with normal hearing are efficient users of fundamental frequency and vocal tract length cues for voice discrimination. Ear Hear. 2020, 41, 182–193. [Google Scholar] [CrossRef] [PubMed]
- Saager, R.B.; Telleri, N.L.; Berger, A.J. Two-detector corrected near infrared spectroscopy (C-NIRS) detects hemodynamic activation responses more robustly than single-detector NIRS. Neuroimage 2011, 55, 1679–1685. [Google Scholar] [CrossRef] [PubMed]
- American Clinical Neurophysiology Society. Guideline 5: Guidelines for standard electrode position nomenclature. Am. J. Electroneurodiagnostic. Technol. 2006, 46, 222–225. [Google Scholar] [CrossRef]
- Jurcak, V.; Tsuzuki, D.; Dan, I. 10/20, 10/10, and 10/5systems revisited: Their validity as relative head-surfacebased positioning systems. NeuroImage 2007, 34, 1600–1611. [Google Scholar] [CrossRef]
- Zimeo Morais, G.A.; Balardin, J.B.; Sato, J.R. fNIRS Optodes’ Location Decider (fOLD): A toolbox for probe arrangement guided by brain regions-of-interest. Sci. Rep. 2018, 8, 3341. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.L.; Wagner, J.; Heugel, N.; Sugar, J.; Lee, Y.W.; Conant, L.; Malloy, M.; Heffernan, J.; Quirk, B.; Zinos, A.; et al. Functional Near-Infrared Spectroscopy and Its Clinical Application in the Field of Neuroscience: Advances and Future Directions. Front. Neurosci. 2020, 14, 724. [Google Scholar] [CrossRef] [PubMed]
- Aasted, C.M.; Yücel, M.A.; Cooper, R.J.; Dubb, J.; Tsuzuki, D.; Becerra, L.; Petkov, M.P.; Borsook, D.; Dan, I.; Boas, D.A. Anatomical guidance for functional near-infrared spectroscopy: AtlasViewer tutorial. Neurophotonics 2015, 2, 020801. [Google Scholar] [CrossRef] [Green Version]
- Huppert, T.J.; Diamond, S.G.; Franceschini, M.A.; Boas, D.A. HomER: A review of time-series analysis methods for near-infrared spectroscopy of the brain. Appl. Opt. 2009, 48, D280–D298. [Google Scholar] [CrossRef] [PubMed]
- Pollonini, L.; Bortfeld, H.; Oghalai, J.S. PHOEBE: A method for real time mapping of optodes-scalp coupling in functional near-infrared spectroscopy. Biomed. Opt. Express 2016, 7, 5104–5119. [Google Scholar] [CrossRef] [Green Version]
- Themelis, G.; Selb, J.; Thaker, S.; Stott, J.J.; Custo, A.; Boas, D.A.; Franceschini, M.-A. Depth of Arterial Oscillation Resolved with NIRS Time and Frequency Domain; Optical Society of America: Washington, DC, USA, 2004. [Google Scholar]
- Scholkmann, F.; Spichtig, S.; Muehlemann, T.; Wolf, M. How to detect and reduce movement artifacts in near-infrared imaging using moving standard deviation and spline interpolation. Physiol. Meas. 2010, 31, 649–662. [Google Scholar] [CrossRef] [Green Version]
- Molavi, B.; Dumont, G.A. Wavelet-based motion artifact removal for functional near-infrared spectroscopy. Physiol. Meas. 2012, 33, 259. [Google Scholar] [CrossRef]
- Brigadoi, S.; Ceccherini, L.; Cutini, S.; Scarpa, F.; Scatturin, P.; Selb, J.; Gagnon, L.; Boas, D.A.; Cooper, R.J. Motion artifacts in functional near-infrared spectroscopy: A comparison of motion correction techniques applied to real cognitive data. Neuroimage 2014, 85, 181–191. [Google Scholar] [CrossRef] [Green Version]
- Delpy, D.T.; Cope, M.; van der Zee, P.; Arridge, S.; Wray, S.; Wyatt, J. Estimation of optical pathlength through tissue from direct time of flight measurement. Phys. Med. Biol. 1988, 33, 1433–1442. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.C.; Tak, S.; Jang, K.E.; Jung, J.; Jang, J. NIRS-SPM: Statistical parametric mapping for near-infrared spectroscopy. Neuroimage 2009, 44, 428–447. [Google Scholar] [CrossRef] [PubMed]
- Uga, M.; Dan, I.; Sano, T.; Dan, H.; Watanabe, E. Optimizing the general linear model for functional near-infrared spectroscopy: An adaptive hemodynamic response function approach. Neurophotonics 2014, 1, 015004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glick, H.; Sharma, A. Cross-modal plasticity in developmental and age-related hearing loss: Clinical implications. Hear. Res. 2017, 343, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Bornstein, M.H. Sensitive periods in development: Interdisciplinary Perspective. In Lawrence Erlbaum Associative; New York University Press: New York, NY, USA, 1987. [Google Scholar]
- Bornstein, M.H. Sensitive periods in development: Structural characteristics and causal interpretations. Psychol. Bull. 1989, 105, 179–197. [Google Scholar] [CrossRef]
- Gordon, K.A.; Papsin, B.C.; Harrison, R.V. Effects of cochlear implant use on the electrically evoked middle latency response in children. Hear. Res. 2005, 204, 78–89. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Dorman, M.F.; Kral, A. The influence of a sensitive period on central auditory development in children with unilateral and bilateral cochlear implants. Hear. Res. 2005, 203, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Kral, A.; Tillein, J. Brain plasticity under cochlear implant stimulation. Adv. Otorhinolaryngol. 2006, 64, 89–108. [Google Scholar] [PubMed]
- Barone, P.; Lacassagne, L.; Kral, A. Reorganization of the connectivity of cortical field DZ in congenitally deaf cat. PLoS ONE 2013, 8, e60093. [Google Scholar] [CrossRef] [PubMed]
- Hickok, G.; Poeppel, D. The cortical organization of speech processing. Nat. Rev. Neurosci. 2007, 8, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Suarez, A.; Valdes-Hernandez, P.A.; Moshkforoush, A.; Tsoukias, N.; Riera, J. Arterial blood stealing as a mechanism of negative BOLD response: From the steady-flow with nonlinear phase separation to a windkessel-based model. J. Theor. Biol. 2021, 529, 110856. [Google Scholar] [CrossRef]
- Mayhew, S.D.; Coleman, S.C.; Mullinger, K.J.; Can, C. Across the adult lifespan the ipsilateral sensorimotor cortex negative BOLD response exhibits decreases in magnitude and spatial extent suggesting declining inhibitory control. NeuroImage 2022, 253, 119081. [Google Scholar] [CrossRef]
- Steinmetzger, K.; Shen, Z.; Riedel, H.; Rupp, A. Auditory cortex activity measured using functional near-infrared spectroscopy (fNIRS) appears to be susceptible to masking by cortical blood stealing. Hear. Res. 2020, 396, 108069. [Google Scholar] [CrossRef]
- Majerus, S.; Van Der Linden, M.; Collette, F.; Laureys, S.; Poncelet, M.; Degueldre, C.; Delfiore, G.; Luxen, A.; Salmon, E. Modulation of brain activity during phonological familiarization. Brain Lang. 2005, 92, 320–331. [Google Scholar] [CrossRef]
- Graves, W.W.; Grabowski, T.J.; Mehta, S.; Gupta, P. The left posterior superior temporal gyrus participates specifically in accessing lexical phonology. J. Cogn. Neurosci. 2008, 20, 1698–1710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gow, D., Jr. The cortical organization of lexical knowledge: A dual lexicon model of spoken language processing. Brain Lang. 2012, 121, 273–288. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.; Goswami, U. Phonological awareness, vocabulary, and reading in deaf children with cochlear implants. J. Speech Lang. Hear. Res. 2010, 53, 237–261. [Google Scholar] [CrossRef] [PubMed]
- Niparko, J.K.; Tobey, E.A.; Thal, D.J.; Eisenberg, L.S.; Wang, N.Y.; Quittner, A.L.; Fink, N.E.; Team, C.I. Spoken language development in children following cochlear implantation. JAMA 2010, 303, 1498–1506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geers, A.E.; Hayes, H. Reading, writing, and phonological processing skills of adolescents with 10 or more years of cochlear implant experience. Ear. Hear. 2011, 32, 49S–59S. [Google Scholar] [CrossRef] [Green Version]
- Chandramouli, S.H.; Kronenberger, W.G.; Pisoni, D.B. Verbal Learning and Memory in Early-Implanted, Prelingually Deaf Adolescent and Adult Cochlear Implant Users. J. Speech Lang. Hear. Res. 2019, 62, 1033–1050. [Google Scholar] [CrossRef]
- Naito, Y.; Tateya, I.; Fujiki, N.; Hirano, S.; Ishizu, K.; Nagahama, Y.; Fukuyama, H.; Kojima, H. Increased cortical activation during hearing of speech in cochlear implant users. Hear. Res. 2000, 143, 139–146. [Google Scholar] [CrossRef]
- Rowland, S.C.; Hartley, D.; Wiggins, I.M. Listening in naturalistic scenes: What can functional Near-Infrared Spectroscopy and intersubject correlation analysis tell us about the underlying brain activity? Trends Hear. 2018, 22, 2331216518804116. [Google Scholar] [CrossRef] [Green Version]
- Evans, S.; McGettigan, C.; Agnew, Z.K.; Rosen, S.; Scott, S.K. Getting the Cocktail Party Started: Masking Effects in Speech Perception. J. Cogn. Neurosci. 2016, 28, 483–500. [Google Scholar] [CrossRef] [Green Version]
- Çukur, T.; Huth, A.G.; Nishimoto, S.; Gallant, J.L. Functional subdomains within human FFA. J. Neurosci. Off. J. Soc. Neurosci. 2013, 33, 16748–16766. [Google Scholar] [CrossRef] [Green Version]
- Weiner, K.S.; Zilles, K. The anatomical and functional specialization of the fusiform gyrus. Neuropsychologia 2016, 83, 48–62. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Wang, J.; Fan, L.; Zhang, Y.; Fox, P.T.; Eickhoff, S.B.; Yu, C.; Jiang, T. Functional organization of the fusiform gyrus revealed with connectivity profiles. Hum. Brain Mapp. 2016, 37, 3003–3016. [Google Scholar] [CrossRef] [PubMed]
- Forseth, K.J.; Kadipasaoglu, C.M.; Conner, C.R.; Hickok, G.; Knight, R.T.; Tandon, N. A lexical semantic hub for heteromodal naming in middle fusiform gyrus. Brain J. Neurol. 2018, 141, 2112–2126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, X.; Sobczak, G.; McKay, C.M.; Litovsky, R.Y. Comparing fNIRS signal qualities between approaches with and without short channels. PLoS ONE 2020, 15, e0244186. [Google Scholar] [CrossRef] [PubMed]
- Luke, R.; Larson, E.; Shader, M.J.; Innes-Brown, H.; Van Yper, L.; Lee, A.K.C.; Sowman, P.F.; McAlpine, D. Analysis methods for measuring passive auditory fNIRS responses generated by a block-design paradigm. Neurophotonics 2021, 8, 025008. [Google Scholar] [CrossRef]
- Yücel, M.A.; Lühmann, A.V.; Scholkmann, F.; Gervain, J.; Dan, I.; Ayaz, H.; Boas, D.; Cooper, R.J.; Culver, J.; Elwell, C.E.; et al. Best practices for fNIRS publications. Neurophotonics 2021, 8, 012101. [Google Scholar] [CrossRef]
- Gagnon, L.; Yücel, M.A.; Boas, D.A.; Cooper, R.J. Further improvement in reducing superficial contamination in NIRS using double short separation measurements. Neuroimage 2014, 85 Pt 1, 127–135. [Google Scholar] [CrossRef] [Green Version]
- Shader, M.J.; Luke, R.; Gouailhardou, N.; McKay, C.M. The use of broad vs restricted regions of interest in functional near-infrared spectroscopy for measuring cortical activation to auditory-only and visual-only speech. Hear. Res. 2021, 406, 108256. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subject ID | Gender | Etiology | Age at Identification (Years;Months) | Age at Implantation (Years;Months) | Age at Testing (Years;Months) | Implant | Vocation |
|---|---|---|---|---|---|---|---|
| CI1 | F | Genetic—connexin 26 | 0;01 | 1;00 (R) 6;06 (L) | 20;04 | Cochlear Nucleus (Both—Nucleus 6) | Student |
| CI2 | F | Genetic—connexin 26 | 0;01 | 1;01 (L) 6;08 (r) | 20;01 | Cochlear Nucleus (R—Freedom, L—Nucleus 5) | Unemployed |
| CI3 | F | Unknown | 0;08 | 2;00 (R) 10;00 (L) | 23;06 | Cochlear Nucleus (Both Nucleus 7) | Student |
| CI4 | M | CMV | 0;03 | 2;00 (L) 12;01 (R) | 22;05 | Cochlear nucleus (R—Nucleus 24, L—Freedom) | student |
| CI5 | F | CMV at pregnancy | 1;00 | 2;05 (R) 18;00 (L) | 26;01 | Cochlear Nucleus (R—Nucleus 5, L—Nucleus 6) | National service |
| CI6 | M | Genetic—connexin 26 | 0;10 | 3;00 (L) 25;04 (R) | 28;10 | Cochlear Nucleus (R—Nucleus 7, L—Nucleus 6) | Social worker |
| CI7 | F | Unknown | 4;06 | 14;00 (R) 16;00(L) | 24 | Both: AB-260 | Student |
| CI8 | F | Genetic | 0;08 | 25;11 (R) 30;06 (L) | 35;05 | Med-El (R—Concerto, L—Synchroni | Teacher |
| CI9 | F | Waardenburg syndrom | 0;01 | 3;00 (L) | 31;01 | Cochlear Nucleus—Espirit 3G | Teacher |
| CI10 | F | Genetic—connexin 26 | 1;00 | 4;06 (L) | 18;04 | Med-El Opus | Student |
| CI11 | M | Rubella in pregnancy | 0;03 | 5;00 (L) | 30;10 | Cochlear—Nucleus 6 | Technician |
| CI12 | F | Unknown | 0:06 | 24;00 (L) | 20;10 | Med-El Combi40+ | National service |
| CI13 | F | Genetic | 3;00 | 31;01 (L) | 33;04 | Cochlear-Kenso | Assistant to kindergarten teacher |
| CI4 | F | Ear infections | 5;06 | 34;00 (R) | 40 | AB Phonac | Kindergarten teacher |
| Channel | SD Pair | Pair on Map | MNI Coordinates x,y,z | SD Distance (mm) | Brodmann Number | Region | Specificity % | ||
|---|---|---|---|---|---|---|---|---|---|
| 1 | S1-D1 | AF7-F5 | −59.589 | 60.168 | −4.362 | 36 | 45 | Pars triangularis Broca’s | 48.79 |
| 46 | Dorsolateral prefrontal cortex | 43.20 | |||||||
| 3 | S2-D1 | AF3-F5 | −52.105 | 65.735 | 13.046 | 41.9 | 45 | Pars triangularis Broca’s | 43.88 |
| 46 | Dorsolateral prefrontal cortex | 32.12 | |||||||
| 4 | S2-D3 | AF3-Afz | −18.354 | 83.517 | 24.189 | 39.1 | 10 | Frontopolar area | 72.47 |
| 5 | S3-D1 | F3-F5 | −58.802 | 54.143 | 23.513 | 30.6 | 45 | Pars triangularis Broca’s | 70.67 |
| 6 | S3-D2 | F3-F1 | −41.387 | 58.42 | 48.855 | 30.5 | 9 | Dorsolateral prefrontal cortex | 66.61 |
| 8 | S4-D2 | Fz-F1 | −15.837 | 63.087 | 62.131 | 30.4 | 9 | Dorsolateral prefrontal cortex | 63.16 |
| 8 | Frontal eye fields | 34.73 | |||||||
| 9 | S4-D3 | Fz-Afz | 0.193 | 75.25 | 49.43 | 39.8 | 9 | Dorsolateral prefrontal cortex | 61.77 |
| 10 | S4-D4 | Fz-F2 | 14.824 | 63.339 | 62.24 | 30.2 | 9 | Dorsolateral prefrontal cortex | 68.93 |
| 11 | S5-D3 | AF4-Afz | 18.291 | 83.473 | 24.403 | 38.7 | 10 | Frontopolar area | 72.47 |
| 12 | S5-D5 | AF4-F6 | 51.371 | 66.117 | 14.439 | 41.2 | 46 | Dorsolateral prefrontal cortex | 49.34 |
| 45 | Pars triangularis Broca’s | 32.12 | |||||||
| 13 | S6-D4 | F4-F2 | 41.232 | 59.179 | 48.129 | 30.3 | 9 | Dorsolateral prefrontal cortex | 68.37 |
| 14 | S6-D5 | F4-F6 | 58.579 | 53.906 | 24.739 | 30.4 | 45 | Pars triangularis Broca’s | 70.67 |
| 16 | S7-D5 | AF8-F6 | 59.018 | 60.776 | −3.202 | 35.5 | 45 | Pars triangularis Broca’s | 43.88 |
| 46 | Dorsolateral prefrontal cortex | 43.18 | |||||||
| 18 | S8-D6 | T7-FT7 | −82.152 | −0.704 | −17.684 | 30.6 | 21 | Middle temporal gyrus | 67.39 |
| 19 | S8-D7 | T7-TP7 | −87.447 | −30.648 | −14.486 | 30.4 | 21 | Middle temporal gyrus | 49.28 |
| 20 | Inferior temporal gyrus | 47.32 | |||||||
| 20 | S8-D8 | T7-C5 | −85.287 | −14.179 | 3.539 | 41.1 | 22 | Superior temporal gyrus | 42.20 |
| 21 | Middle temporal gyrus | 37.02 | |||||||
| 22 | S9-D7 | CP5-TP7 | −83.896 | −46.505 | 6.45 | 40.8 | 22 | Superior temporal gyrus | 35.29 |
| 21 | Middle temporal gyrus | 34.85 | |||||||
| 23 | S9-D8 | CP5-C5 | −85.028 | −29.926 | 25.727 | 33.6 | 48 | Restosubicular area | 36.01 |
| 24 | S9-D15 | CP5-P5 | −77.374 | −64.104 | 26.858 | 33.6 | 39 | Angular gyrus part of Wernicke’s area | 32.67 |
| 25 | S10-D9 | T8-FT8 | 81.902 | 0.854 | −15.968 | 30 | 21 | Middle temporal gyrus | 84.30 |
| 26 | S10-D10 | T8-C6 | 85.078 | −12.698 | 4.825 | 41.1 | 22 | Superior temporal gyrus | 47.33 |
| 21 | Middle temporal gyrus | 38.02 | |||||||
| 27 | S10-D11 | T8-TP8 | 86.471 | −30.117 | −13.132 | 31 | 21 | Middle temporal gyrus | 54.96 |
| 20 | Inferior temporal gyrus | 41.52 | |||||||
| 29 | S11-D10 | CP6-C6 | 84.951 | −28.595 | 27.436 | 34.3 | 22 | Superior temporal gyrus | 31.60 |
| 30 | S11-D11 | CP6-TP8 | 83.928 | −46.19 | 8.435 | 40.9 | 21 | Superior temporal gyrus | 40.43 |
| 22 | Middle temporal gyrus | 35.40 | |||||||
| 31 | S11-D13 | CP6-P6 | 78.121 | −62.051 | 27.562 | 33.9 | 39 | Angular gyrus part of Wernicke’s area | 34.98 |
| 32 | S12-D9 | FT10-FT8 | 80.743 | 13.185 | −38.235 | 41.9 | 21 | Middle temporal gyrus | 69.07 |
| 33 | S12-D12 | FT10-F10 | 78.136 | 27.381 | −59.246 | 29.1 | 38 | Superior temporal gyrus | 38.91 |
| 20 | Inferior temporal gyrus | 32.41 | |||||||
| 35 | S13-D11 | P8-TP8 | 79.743 | −60.24 | −9.105 | 31.7 | 37 | Fusiform gyrus | 68.11 |
| 36 | S13-D13 | P8-P6 | 73.274 | −76.332 | 9.546 | 33.6 | 37 | Fusiform gyrus | 68.82 |
| 37 | S14-D6 | FT9-FT7 | −80.817 | 12.621 | −38.559 | 41.7 | 21 | Middle temporal gyrus | 69.10 |
| 38 | S14-D14 | FT9-F9 | −78.71 | 25.365 | −59.946 | 29.7 | 38 | Temporopolar area | 42.31 |
| 20 | Inferior temporal gyrus | 31.83 | |||||||
| 40 | S15-D7 | P7-TP7 | −79.372 | −61.256 | −10.554 | 31.4 | 37 | Fusiform gyrus | 71.20 |
| 41 | S15-D15 | P7-P5 | −72.834 | −77.39 | 8.481 | 33.5 | 37 | Fusiform gyrus | 68.55 |
| Group/Channel | NH | NHV | CI |
|---|---|---|---|
| 1 | 0 | 0 | 1 |
| 3 | 0 | 0 | 1 |
| 4 | 0 | 0 | 0 |
| 5 | 0 | 0 | 1 |
| 6 | 0 | 0 | 1 |
| 8 | 0 | 0 | 1 |
| 9 | 2 | 0 | 3 |
| 10 | 0 | 0 | 0 |
| 11 | 1 | 0 | 0 |
| 12 | 0 | 0 | 0 |
| 13 | 0 | 1 | 1 |
| 14 | 2 | 0 | 1 |
| 16 | 0 | 0 | 0 |
| 18 | 0 | 0 | 0 |
| 19 | 0 | 0 | 0 |
| 20 | 0 | 0 | 1 |
| 22 | 0 | 0 | 0 |
| 23 | 0 | 0 | 0 |
| 24 | 1 | 0 | 2 |
| 25 | 0 | 0 | 2 |
| 26 | 1 | 0 | 2 |
| 27 | 1 | 0 | 5 |
| 29 | 0 | 0 | 0 |
| 30 | 0 | 0 | 3 |
| 31 | 0 | 0 | 0 |
| 32 | 2 | 0 | 0 |
| 33 | 0 | 1 | 0 |
| 35 | 1 | 0 | 4 |
| 36 | 0 | 0 | 1 |
| 37 | 0 | 1 | 2 |
| 38 | 0 | 0 | 2 |
| 40 | 0 | 0 | 5 |
| 41 | 0 | 0 | 4 |
| CI | NHV | NH | |||
|---|---|---|---|---|---|
| Serial Number | SRTn | Serial Number | SRTn | Serial Number | SRTn |
| CI1 | −4 | NHV1 | −5.18 | NH1 | −9.62 |
| CI2 | −4 | NHV2 | −0.41 | NH2 | −6.30 |
| CI3 | −3.96 | NHV3 | −3.69 | NH3 | −8.75 |
| CI4 | 0.22 | NHV4 | −1.33 | NH4 | −10.69 |
| CI5 | CE | NHV5 | −0.41 | NH5 | −11.32 |
| CI6 | −3.46 | NHV6 | −2.26 | NH6 | −8.14 |
| CI7 | 3.53 | NHV7 | −3.54 | NH7 | −9.37 |
| CI8 | 0.49 | NHV8 | −0.41 | NH8 | −10.00 |
| CI9 | 2.40 | NHV9 | −3.23 | NH9 | −10.27 |
| CI10 | −5.52 | NHV10 | −1.42 | NH10 | −9.62 |
| CI11 | 6.75 | NHV11 | 0.74 | NH11 | −7.02 |
| CI12 | −3.36 | NHV12 | −2.55 | NH12 | −8.14 |
| CI13 | CE | NHV13 | −3.48 | NH13 | −9.62 |
| CI14 | CE | NHV14 | −1.42 | NH14 | −9.62 |
| NHV15 | −3.56 | NH15 | −7.64 | ||
| Mean (SD) | 0.34 (5.95) | −2.14 (1.64) | −9.03 (1.35) | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Levin, M.; Balberg, M.; Zaltz, Y. Cortical Activation in Response to Speech Differs between Prelingually Deafened Cochlear Implant Users with Good or Poor Speech-in-Noise Understanding: An fNIRS Study. Appl. Sci. 2022, 12, 12063. https://doi.org/10.3390/app122312063
Levin M, Balberg M, Zaltz Y. Cortical Activation in Response to Speech Differs between Prelingually Deafened Cochlear Implant Users with Good or Poor Speech-in-Noise Understanding: An fNIRS Study. Applied Sciences. 2022; 12(23):12063. https://doi.org/10.3390/app122312063
Chicago/Turabian StyleLevin, Michal, Michal Balberg, and Yael Zaltz. 2022. "Cortical Activation in Response to Speech Differs between Prelingually Deafened Cochlear Implant Users with Good or Poor Speech-in-Noise Understanding: An fNIRS Study" Applied Sciences 12, no. 23: 12063. https://doi.org/10.3390/app122312063