
Photo-Fermentative Bacteria Used for Hydrogen Production

1
FibEnTech, Department of Chemistry, University of Beira Interior, 6201-001 Covilhã, Portugal
2
Departamento de Farmacología, Fisiología y Medicina Legal y Forense, Facultad de Veterinaria, Universidad de Zaragoza, Miguel Servet 177, 50013 Zaragoza, Spain
3
Departamento de Química Analítica, Instituto de Investigación en Ingeniería de Aragón (I3A), Escuela de Ingeniería y Arquitectura (EINA), Universidad de Zaragoza, María de Luna 3 (Edificio Torres Quevedo), 50018 Zaragoza, Spain
*
Author to whom correspondence should be addressed.
Appl. Sci. 2024, 14(3), 1191; https://doi.org/10.3390/app14031191
Submission received: 19 December 2023
/
Revised: 24 January 2024
/
Accepted: 26 January 2024
/
Published: 31 January 2024
(This article belongs to the Topic Advances in Renewable Energy Technologies and Systems Solutions)
Abstract
:Photo-fermentation is an efficient hydrogen production pathway in which purple non-sulfur bacteria (PNSB) play an active role and produce hydrogen as a part of their metabolism under optimal conditions. These bacteria work under the influence of light to advance their metabolism and use various substrates, such as simple sugars and volatile fatty acids, to produce hydrogen. This article presents a comparative review of several bacterial strains that have been efficiently used to produce hydrogen by photo-fermentation under different optimized conditions, including the substrate, its concentration, type and capacity of the bioreactor, light sources and intensities, and process conditions to achieve the maximum biohydrogen production rate. The analysis showed that the Rhodopseudomonas palustris is the main bacterium used for hydrogen production, with a maximum hydrogen production rate of 3.2 mM/h using 27.8 mM of glucose in a 165 mL serum bottle and 3.23 mM/h using 50 mM of glycerol at pH 7, followed by Rhodobacter sphaeroides, which gave a hydrogen production rate as high as 8.7 mM/h, using 40 mM of lactic acid, pH 7, and 30 °C temperature in a single-walled glass bioreactor. However, it is not preferred over R. palustris due to its versatile metabolism and ability to use an alternative mode if the conditions are not carefully adjusted, which can be a problem in hydrogen production.
1. Introduction
Hydrogen is a clean and alternative energy source to fossil fuels [1]. It is the most abundant element in the universe and can be produced from a variety of renewable resources, including water, biomass, and solar energy. Hydrogen is an attractive fuel for transport, and its environmental impact is determined in terms of greenhouse gases, acidification, eutrophication, toxicity potential, and eco-cost [2]. The energy content of hydrogen is 122 kJ/g, which is 2.72 times higher than that of petrol [3]. However, the current production methods are based on fossil fuels, which cause greenhouse gas emissions. Methods, such as the thermal cracking of water or water electrolysis, have been used to produce hydrogen, but they are energy-intensive and emit gases, such as carbon dioxide, that cause global warming [4]. Hydrogen gas is advantageous because it can be used to produce electricity in fuel cells with heat and water as end products [5], which helps to reduce air pollution and mitigate climate change. The development of fuels with net-zero emissions is a current research priority [6]. Hydrogen has a higher energy content per unit weight than fossil fuels and can, therefore, be used in fuel cells for electric vehicles, power generation, and in industrial processes. Hydrogen can be stored and transported, making it a flexible gas carrier. However, there are challenges associated with the production and use of hydrogen, including the high cost of production and storage, its corrosiveness to metals, the need for infrastructure development, and safety concerns related to handling and storage. In any case, the production and use of hydrogen is an important area of research and development as we strive to improve both economic and environmental sustainability in the near future [7].
A promising method of producing hydrogen from biomass using microorganisms has been favored because it is environmentally friendly [4]. Such a biological method contributes to the production of renewable and carbon-neutral hydrogen, referred to as “biohydrogen” [8].
Biological hydrogen production includes photo-fermentation and dark fermentation. Photo-fermentation, which is the main focus of this review, involves bacteria that can convert organic matter into hydrogen using solar energy. It has been widely used to produce clean, renewable energy sources [8]. Substrates, such as simple sugars, volatile fatty acids, and industrial and agricultural wastes under anaerobic conditions in the presence of light, can be used by purple non-sulfur bacteria in photo-fermentation [9]. Based on this, several photobioreactors (PBRs) have been developed to improve the production of hydrogen in continuous operation, including different configurations, such as flat panel, tubular, or torus reactors [10].
The operation of PBRs at ambient conditions reduces the need for additional energy input, thereby reducing the process costs compared to thermochemical and electrolytic methods [11]. Some of the microbes that have been studied to carry out photo-fermentation include anaerobic bacteria of the genera Rhodobacter, Rhodobium, Rhodopseudomonas, and Rhodospirillum, which help to convert organic acids into hydrogen and carbon dioxide. The amount of hydrogen produced in this process is comparable to biophotolysis and it is influenced by the type of microbe, the medium, and the design of the PBR [12].
Other hydrogen production methods include microbial electrohydrogenesis cells, which produce hydrogen simultaneously with wastewater treatment, biophotolysis, which dissociates the water molecule into hydrogen and oxygen and, finally, dark fermentation [7]. Dark fermentation involves strict anaerobes or facultative bacterial strains using organic matter, biomass, and wastewater as substrates. It is an efficient approach as it takes place at an ambient temperature and pressure, produces clean energy, helps in the disposal of organic waste, and is influenced by parameters, such as pH, organic loading rate, inoculum type, retention time, availability of nutrients, and the organic substrate type, which have a significant impact on the amount of hydrogen produced [13]. Some of the microbes involved in dark fermentation are Clostridium, Ethanoligenens, and Desulfovibrio species.
This review aims to analyze and determine various photo-fermentative bacteria under different environmental conditions, such as temperature, pH, reactor design and material, light sources required, and the substrates for optimal hydrogen production. The article discusses a comparative study between different bacterial strains based on different parameters as mentioned above, giving a detailed idea of the suitable microbes that can be considered for high hydrogen production using photo-fermentation under different environmental conditions. Figure 1 shows the possibilities for hydrogen production using different methods previously described in the literature.
2. Photo-Fermentative Bacteria for Hydrogen Production
The production of hydrogen by photo-fermentation was recognised in the year 1949 by Gest and Kamen as it produces hydrogen with high purity without generating oxygen [14]. In this process, the organic matter is decomposed by photosynthetic bacteria in the presence of light to produce hydrogen through a reaction catalyzed by nitrogenase.
C6H12O6 + 6 H2O → 6 CO2 + 12 H2 (photosynthetic bacteria)
Bacteria commonly used for photo-fermentation include purple non-sulfur bacteria (PNSB) because they have high metabolic flexibility and can grow as photoautotrophs and chemoheterotrophs. They do not require water, unlike plants, algae, and cyanobacteria, nor do they require much free energy for hydrogen production, so no oxygen is produced in the system. Inorganic ions, carbon compounds or hydrogen are the electron sources for carrying out metabolic activities, and the generation of energy is produced by photosynthesis. PNSB show a versatile ability to utilize a wide range of organic carbon compounds for metabolic activity including pyruvate, acetate, amino acids, alcohols, organic acids, and carbohydrates. In particular, certain species within this group can use C1 compounds, such as formate and methanol, as viable carbon sources. In addition, PNSB can also utilize aromatic organic compounds, such as benzoate, cinnamate, and chlorobenzoate, to meet their carbon requirements [15]. PNSB also demonstrate the ability to assimilate a range of organic acids, including acetic, butyric, propionic, malic, and lactic acids, which becomes relevant when using organic waste as a substrate for hydrogen production. During the acidogenic phase of anaerobic digestion, PNSB can use these organic acids as carbon sources for their conversion to hydrogen and carbon dioxide. Thus, the main significance and advantage of photo-fermentative bacteria is their ability to achieve high hydrogen yields. The theoretical yield achieved from 1 mol of glucose is 12 mol H2, as given in equation 1 above, but the yield can vary slightly depending on the type of microbe. As shown in Figure 2, the following PNSB species show the ability to produce hydrogen using photo-fermentation: Rhodobacter species [16], such as Rhodobacter sphaeroides [17] and Rhodobacter capsulatus [18]; Rhodopseudomonas species [19], such as Rhodopseudomonas palustris [20], Rhodopseudomonas capsulata; Rhodospirillum rubrum [21]; Rhodovulum species, including Rhodovulum sulfidophilum [22]. These microbes follow the pathways supported by ATP-dependent nitrogenase, as the metabolic pathway is similar in all these bacteria; among them, Rhodospirillum rubrum was reported as the first PNSB to produce hydrogen.
2.1. Metabolic Pathway of Photo-Fermentative Bacteria
Hydrogen metabolism is of great importance in maintaining the stability and performance of various microbial biotopes at the ecosystem level. Hydrogen has been used as an electron donor for reductive dehalogenation by several microbes and for the presence of hydrogenase enzymes [23]. There are certain key factors associated with the metabolic pathways of these microbes that enhance hydrogen production. These include light absorption, as these bacteria possess photosynthetic pigments that allow light energy to be absorbed from the environment, followed by the incorporation of organic substrates, such as sugars and volatile fatty acids, which are then catabolized through metabolic pathways.
This light energy enables the reduction of protons to produce molecular hydrogen while carbon fixation takes place for bacterial growth. R. palustris, which has been extensively studied for hydrogen production, assimilates carbon dioxide into organic compounds via the Calvin–Benson–Bassham cycle, which contributes to bacterial growth and its biomass production, although this cycle is not the primary route of hydrogen production in this species [24]. The photosystem of PNSB is not very efficient at splitting water, so no oxygen is involved, making it suitable for hydrogen production. Protons are pumped into the periplasmic space, as electrons move through the electron transport chain to ferredoxin, which delivers these electrons to the nitrogenase enzyme to reduce molecular nitrogen to ammonia, creating a proton gradient or proton motive force. This is followed by the hydrogenase enzyme present in these bacteria, which catalyzes the reversible reaction of proton reduction to form molecular hydrogen, which accumulates in the periplasmic space. In the absence of nitrogen, nitrogenase functions similarly to hydrogenase, catalyzing the reduction of protons with the electrons derived from ferredoxin.
The processes of hydrogen production and efflux generate ATP as a form of energy through a process called chemiosmotic ATP synthesis, which is used by the cells for various metabolic processes [25]. A general outline of the metabolic pathway followed by these photo-fermentative bacteria is presented in Figure 3, and a detailed description of the complete process is given in Figure 4.
2.2. Rhodopseudomonas
Rhodopseudomonas bacteria contain a reaction center with bacteriochlorophyll b [27], which was discovered in 1963 and has a wide range of metabolic processes. It is found in a variety of marine environments as well as in soil [28]. Some strains of these microbes are used in photo-fermentation and will be discussed here.
2.2.1. Rhodopseudomonas palustris
This is a Gram-negative bacterium that uses light energy for its metabolic pathway, which leads to the conversion of organic matter into hydrogen. These microbes can alleviate heavy metal toxicity, improve plant growth, and control various plant pathogens [28].
Several studies have been carried out using these bacteria to analyze hydrogen production during photo-fermentation. In a study conducted by Wang et al., the biochemical kinetics of photo-fermentative hydrogen production were investigated experimentally and numerically in order to optimize the photo-fermentative hydrogen production process. R. palustris showed a maximum specific growth rate of 0.26 h−1 at an irradiance of 47.4 W/m2, 30 °C culture temperature, at 12 h and a pH of 7.0. An initial glucose concentration of 9.9 g/L was used as a substrate in a batch mode study, and gas chromatography was performed to analyze the hydrogen content with the flow rate of argon as carrier gas adjusted to 25 mL/min; a maximum specific hydrogen production rate of 1.63 × 10−3 h−1 was observed. An increase in the specific hydrogen production rate was obtained with an increment in temperature to 30 °C, but this decreased to 1.11 × 10−3 h−1 when the temperature was further increased to 40 °C, suggesting a reversible inactivation of cellular enzymes and inhibition of hydrogen formation [29].
In a further analysis, mutant strains of R. palustris were synthesized by the manipulation of nitrogen regulatory genes, as it proved to be an effective way of increasing the ammonia tolerance in photosynthetic bacteria to improve hydrogen production. Acetate (20 mM) and butyrate (20 mM) were used as carbon sources for cell growth and hydrogen production at 30 °C in Sistrom’s medium with an irradiance of 39.5 W/m2. The mutant strain, named nifA draT2, was 25 times more tolerant to ammonium than the wild-type strain, demonstrating that manipulation of nitrogen regulatory genes is a reliable way to increase the ammonium tolerance of these bacteria and improve their hydrogen-producing capacity. The hydrogen produced was collected in syringes and analyzed by gas chromatography. The mutant strain nifA draT2 yielded 2744 ± 66 mL of hydrogen/L, which was higher than the wild-type strain, indicating that the hydrogen production was enhanced by the nifA mutation [30].
The review by Sagir and Alipour discussed the immobilization of photosynthetic bacteria, which helped to increase hydrogen production, and different materials, such as agar, glass beads, alginate, and porous glass have been used for the attachment and immobilization of R. palustris in different studies [31]. A strategy to increase the productivity of hydrogen was recently analyzed in R. palustris using lignocellulose substrate by supplementing it with iron and molybdenum, which acts as a biocatalyst to facilitate electron transfer and ultimately an increase in hydrogen production with a maximum of 2.15 mM/h [32]. A list of hydrogen-producing R. palustris strains in biofilm PBRs is given in Table 1.
The results of several studies showed that the optimum process conditions for maximizing hydrogen yields from R. palustris strains were close to pH 7 and at a temperature range of 30–40 °C in different bioreactors and varying light intensities from different sources. Therefore, this microbe has the potential to use different organic sources to produce hydrogen, offering a glimpse into the near future where sustainable and renewable energy sources will play an important role.
2.2.2. Rhodopseudomonas capsulata
R. capsulata can produce hydrogen by converting light into chemical energy. This microbe is mainly found in aquatic environments and is known for its ability to perform anoxygenic photosynthesis, as it does not produce oxygen as a by-product. Bacteriochlorophyll and carotenoids are the essential pigments for photosynthesis in these bacteria.
In a study conducted by Eroglu et al., the maximum hydrogen productivity was achieved at a temperature of 30 °C and an irradiance of 31.6 W/m2 [49]. In addition, a study carried out in 1971 to elucidate the photophosphorylation system of this microbe found that sonication of chromatophores in the presence of EDTA produced non-photo-phosphorylating particles and protein factors that helped to restore photophosphorylation [50].
It was observed that the metabolism of R. capsulata was affected by intermittent illumination, suggesting that the light conversion and biosynthesis are closely linked in these bacteria. Thus, the analysis of R. capsulata could help to gain an insight into the phototrophy of the bacteria and its ability to produce hydrogen, and to identify the relationship between the energy conversion and biosynthesis. R. capsulata has been identified for its ability to degrade toxic contaminants along with hydrogen. Benzoate is one of the identified inhibitory compounds that can prevent hydrogen production during anaerobic metabolism in the absence of light and electron acceptors, but under phototrophic conditions, R. capsulata can extract hydrogen and use benzoate as a carbon source [51]. Table 2 shows a list of substrates used by these bacteria hydrogen production.
The studies showed that a mixture of acetate, propionate, and butyrate is an optimal carbon source for R. capsulata strains to produce hydrogen in the PBRs. However, the microbe has not been analyzed in many studies but has shown a great potential for photo-biological hydrogen production, paving the way for the production of renewable energy sources in the future.
2.3. Rhodobacter
Rhodobacter is a Gram-negative bacterium belonging to the family Rhodobacteraceae, which has a 16S rRNA gene-based phylogeny and is known for its anoxygenic photosynthetic ability [52]. They are found in freshwater and marine habitats. This genus comprises a heterogeneous group of members, and the analysis of its pan-genome revealed that 1239 core genes are shared by 12 Rhodobacter [55].
2.3.1. Rhodobacter capsulatus
R. capsulatus is a member of the PNSB. Like other microbes in the genus Rhodobacter, these bacteria can also grow and function by using energy from light without releasing oxygen as a by-product [56]. These bacteria are facultative anaerobes that can switch metabolic pathways accordingly and can use a variety of organic compounds as carbon and energy sources. In these bacteria, bacteriochlorophyll absorb light energy to initiate photosynthesis, electrons are transferred from water to the photosynthetic reaction centers within a cell, and in return, protons are pumped into the periplasmic space, thus creating a proton gradient. The hydrogenase enzyme catalyzes the proton reduction reaction to produce hydrogen gas as the end product [57].
Ma et al. reduced the bacteriochlorophyll content of R. capsulatus and used genetic engineering and transposon mutagenesis to create a mutant strain that produces 50.5% more hydrogen. The reduction in pigment allows more light to reach the bacteria, resulting in increased phototrophic growth and hydrogen production [58].
R. capsulatus strains have been widely used for hydrogen production. These microorganisms can utilize different carbon sources, such as glucose, acetate, or sucrose, under different process conditions, as shown in Table 3. However, challenges, such as low yield and sensitivity to environmental conditions, have become a challenge. Despite these limitations, the organism still has a great potential to be used as a natural hydrogen producer.
2.3.2. Rhodobacter sphaeroides
R. sphaeroides is a photosynthetic bacterium found in freshwater and marine ecosystems with a metabolism that produces hydrogen under illumination in the presence of an inert, anaerobic atmosphere. The conditions for hydrogen production must be carefully adjusted, but sometimes it is unavoidable and an alternative mode prevails, and the microbe can modify its metabolic pathways due to its versatile metabolism [68].
These bacteria, like other PNSBs, have bacteriochlorophyll, which absorbs photons and converts them into chemical energy. NiFe hydrogenases, which catalyze the formation of molecular hydrogen, are commonly found in these bacteria. A list of certain conditions for the use of this microbe to produce hydrogen, which have been studied in various research papers, is given in Table 4.
R. sphaeroides has been analyzed in several studies with different carbon sources and has shown great potential as a renewable and environmentally friendly source with the ability to convert organic compounds into hydrogen gas. Further optimization of the environmental conditions for its growth and genetic engineering of its strains helps to enhance its hydrogen production capabilities, making this microbe an efficient candidate for the development of green energy.
2.4. Rhodospirillum rubrum
R. rubrum is a spiral, purple photosynthetic bacterium of the genus Rhodospirillum in the class Alphaproteobacteria. The bacteria are anoxygenic phototrophs that produce extracellular elemental sulfur instead of oxygen while harvesting light with their single complex LH1 rubrum and are found in stagnant freshwater ecosystems [79]. A carotenoid spirilloxanthin is bound to the LH1 complex, which has 16 subunits observed by electron microscopy [78]. These bacteria use similar carbon sources to other purple photosynthetic bacteria, including acetate, malate, glucose, fructose, and sucrose, while producing hydrogen [79].
Different strains of these bacteria were used in the PBRs to produce hydrogen and to determine exergy, which is the useful work potential or the available energy. In a study using R. rubrum (ATCC 10801), acetate was used as the carbon source in a 2 L Biostat (Sartorius, Goettingen, Germany) PBR at 30 °C, 1 atm, with 0–0.85 mL/min flow rates of liquid media, stirring at 150–500 rpm with a total of 540 h of continuous hydrogen production. Tungsten lamps with an average irradiance of 11.85 W/m2 were used as the light source, and gas samples were analyzed by gas chromatography. The process exergy efficiency was in the range of 14.71–22.90% under optimal conditions and 14.71–22.84% using conventional and eco-exergy concepts [80]. Another study demonstrating the efficiency of R. rubrum in the batch hydrogen production process was analyzed using different carbon sources, including formate, acetate, malate, glucose, fructose, and sucrose, to support microbial growth. The process used both conventional exergy and eco-exergy concepts to evaluate the exergy efficiency, simultaneously produced molecular hydrogen and identified acetate as the optimal substrate for hydrogen production. A pH of 7.5, 60 W tungsten lamps with a uniform irradiance of 7.9 W/m2, a pressure of 1 atm, and a constant speed of 200 rpm were used to produce hydrogen. Using acetate as a carbon source, a minimum of 189.67 and 181.40 kJ/kJ of H2 exergy destruction was obtained, proving that the exergy analysis can be used to determine and compare the renewability of hydrogen production [21].
2.5. Rhodovulum sulfidophilum
R. sulfidophilum is a marine acid-tolerant photo-fermentative bacterium. They are Gram-negative, rod-shaped bacteria and can accumulate poly(3-hydroxybutyrate) (PHB) to over 50% of cell dry weight when acetate is used as a substrate. Hydrogen production in the presence of PHB by R. sulfidophilum shows that the loss of reducing equivalents to produce hydrogen can be recovered by the degradation of PHB [81].
In most cases, a neutral initial pH is preferred by the photosynthetic bacteria for optimal hydrogen production, but screening for acid-tolerant strains is important to increase the production of hydrogen in an acidic environment and at higher temperatures. Thus, a Tn7-based transposon was inserted into the genomic DNA of R. sulfidophilum P5 and the hydrogen yield and average hydrogen production rate of 2.16 ± 0.10 mol/mol acetate and 10.06 ± 0.47 mL/L h were observed, respectively, which were approximately 17.32- and 15.37-fold higher than those of the wild-type strain, respectively [82]. Another strain, R. sulfidophilum TH-102, was studied, and it was analyzed individually and in co-culture with other microbes of dark fermentation; the results showed that the addition of strain TH-102 can stabilize pH, decrease oxygen reduction potential, and prolong hydrogen production. The analyses provided data showing that hydrogen production during dark and photo-fermentation alone was sustained for 72 and 168 h, but when using a co-culture it went from 168 to 216 h. The co-culture with the ratio of dark/photo-fermentation bacteria 1:10 produced the highest hydrogen yield of 1694 ± 21 mL/L [83].
Based on the studies with different photo-fermentative microbes, single-stage and two-stage fermentation have been carried out to produce hydrogen. Single-stage photo-fermentation is more cost-effective than two-stage because it utilizes a wide range of substrates, thus yielding more hydrogen as compared to dark fermentation [84]. Two-stage photo-fermentation involves either coupling dark fermentation with photo-fermentation in sequence or combining bacteria used for dark and photo-fermentation to increase hydrogen yield [85]. By dividing the hydrogen production process into two stages, each stage could be optimized based on specific requirements and could lead to an improved hydrogen production efficiency compared to single-stage photo-fermentation.
3. Discussion
This review discusses the microbes used in the photo-fermentative hydrogen production and the ideal conditions for increasing the efficiency and rates of hydrogen production are discussed. Initially, we considered the discussion based on exergy calculations. This is a very realistic approach in wastewater treatment plants where operating costs, energy losses, or fees per volume of water treated are critical. However, most of the studies reviewed do not provide essential data for such calculations, e.g., volatile fatty acid formation, amounts of CO2 produced, heat losses, heating, agitation, or other energy costs. We have, therefore, chosen to report all results in a homogeneous format, expressed as mmol of hydrogen per liter of substrate per hour (mM/h), a parameter that is also of great interest at an industrial level.
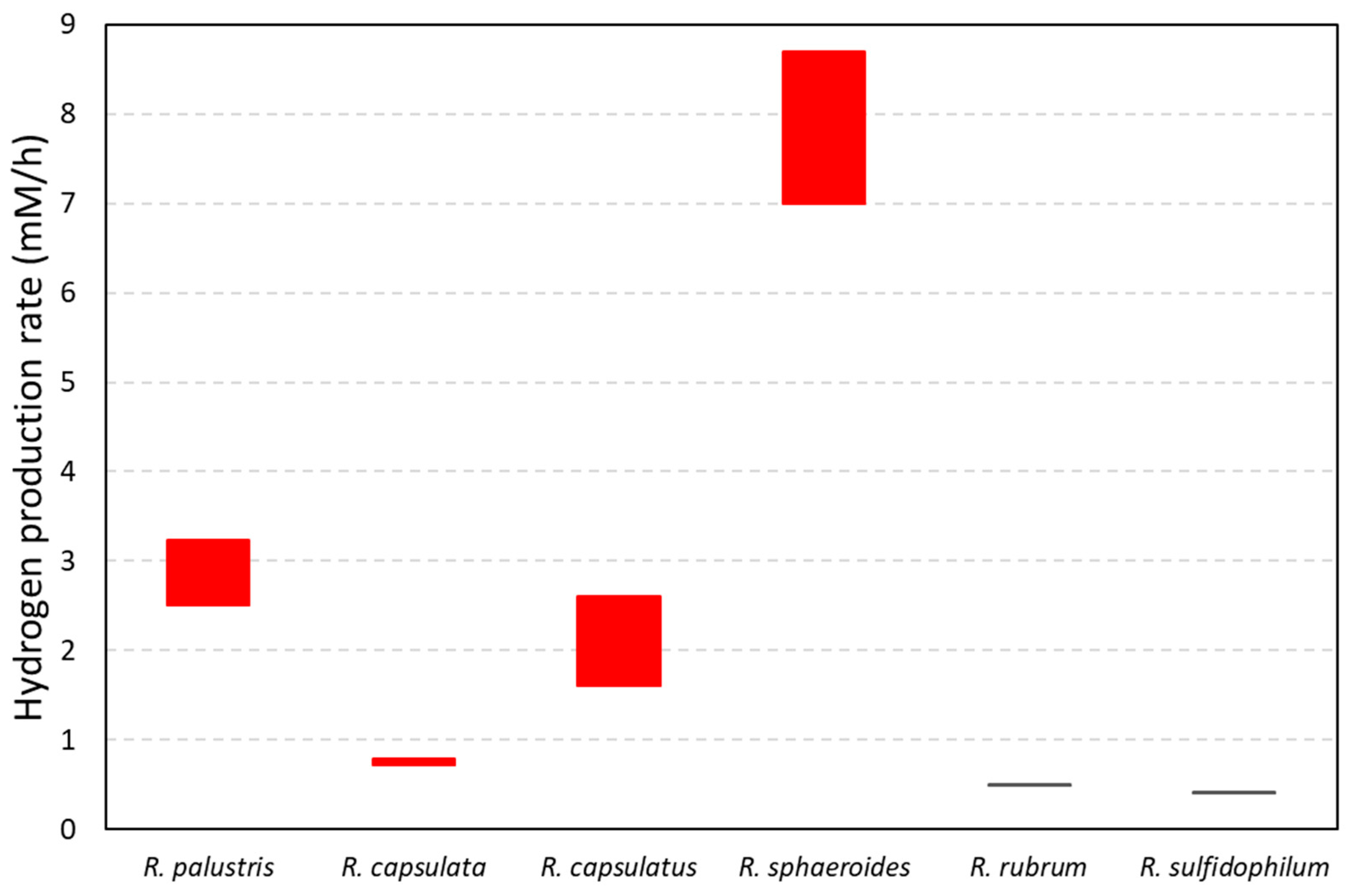
Table 5 gives a summary of the strains of different bacterial species with their maximum hydrogen production rate, providing a useful guide for comparison purposes. The ranges are shown in Figure 5.
A comparative analysis of hydrogen-producing bacteria by photo-fermentation showed that R. palustris is the preferred species for hydrogen production under certain environmental conditions due to its hydrogen production rate. A hydrogen production rate of 3.2 mM/h was observed by the CQK01 strain of this bacterium using glucose as a substrate in a serum bottle at pH 7 and a temperature of 30 °C, followed by its NCIMB 11774 strain using 50 mM glycerol as a substrate in a 1 L fluidized bed PBR, giving a hydrogen production rate of 3.23 mM/h. The results obtained for this, and for all the bacteria analyzed in this review, show that most of these microbes work well around a pH of 7, but the temperature and the type of bioreactor used in the research studies play a major role in the yield of hydrogen production, as can be observed in Table 1 and also in the microbial analysis as well. The same strain CQK01 of R. palustris with 56 mM of glucose as the substrate, at pH 7, and with a tungsten lamp of 31.6 W/m2 irradiance provided a hydrogen production rate of 1.75 mM/h at 30 °C in a PBR made of PMMA [33], while a 2.72 mM/h hydrogen production rate between the range 20–40 °C, when a cylindrical PMMA PBR with chitosan medium-coated optical fiber was used [36], demonstrating the importance of the bioreactor design, its material, and the temperature at which the reactor is operated for hydrogen production. On the other hand, R. capsulata, another bacterium of the same genus already discussed in the review, has mostly been used with substrates, such as acetate, propionate, butyrate, or their mixtures in PBRs or glass vials, but the studies shown in Table 2 indicate that the hydrogen production rates of these microbes are low, as the light conversion is closely linked to their biosynthesis. A maximum hydrogen production rate of 0.79 mM/h with acetate, 0.71 mM/h with propionate, and 0.65 mM/h with a mixture of acetate, propionate, and butyrate was observed at pH 6.8 and a temperature of about 32 °C [53]. By increasing the temperature to 35 °C, under similar conditions, the rate was even lower at 0.56 mM/h, demonstrating the dependence of hydrogen production on temperature and bioreactor design and material [52].
R. capsulatus, a Gram-negative Rhodobacter extensively considered in this review, is very sensitive to environmental conditions, yet these microbes have been used for hydrogen production. R. capsulatus strain JP91 gave a maximum hydrogen production rate under similar environmental conditions as other photo-fermentative bacteria, with pH 7, 30 °C, tungsten lamps as a light source, and glucose concentration of 56 and 35 mM as the substrates. The rate of hydrogen production as observed in R. palustris is dependent on the temperature, bioreactor design, volume, and material, with hydrogen produced at a rate of 1.66–2.3 mM/h using a 350 mL continuous photobioreactor and 2.6 mM/h hydrogen in a 100 mL serum bottle in a batch process [59,60]. On the other hand, acetate used as a substrate does not work well with these species, giving a hydrogen production rate as low as 0.15 mM/h, at pH 8 and temperature < 30 °C [66], indicating the specificity required for the optimal metabolic activity of this microbe. R. sphaeroides, a bacterium of the same genus, has shown a better potential than R. capsulatus and even R. palustris, with strain HY01 having a hydrogen production rate as high as 7 mM/h at a pH range of 7.0–8.2 and 30 °C temperature in a 30 mL syringe using a mixture of 25 mM acetate and 34 mM butyrate and halogen lamps for light irradiance [76], and strain DSM158 with a hydrogen production rate 8.7 mM/L with 40 mM of lactic acid as the substrate and tungsten lamps as light sources in a single-walled glass vessel at pH 7 and 30 °C [74]. These results make this microbe the most efficient of all for hydrogen production, but because its metabolic pathway is very diverse, the environmental conditions under which this bacterium can produce hydrogen are very specific. On the other hand, R. rubrum has only been studied for the exergy analysis in hydrogen production. The substrates used by the bacteria are similar to other PNSBs, including acetate, malate, and glucose, using light sources, such as tungsten lamps, which help to determine and compare the renewability of hydrogen production. Finally, R. sulfidophilum was studied and compared with the other PNSBs in the review. These acid-tolerant bacteria work well in acidic environments and at high temperatures. A strain P5 with acetate as a substrate has given a hydrogen production rate of 0.50 mM/h, and a strain TH-102 with glucose has given a hydrogen rate of 0.42 mM/h [83], making it an efficient hydrogen-producing bacterium. Substrate, pH, temperature, bioreactor design and material, and light sources were identified for each bacterium, and it was found that most of them worked well in a pH range of 6.8–8.0, as a more acidic pH significantly reduces the hydrogen yield due to the inhibition of metabolism by the formation of volatile fatty acids. The effect of the temperature varied among bacteria. R. palustris gave a maximum hydrogen yield at 28 °C with glycerol as a substrate after 192 h and at 30 °C with glucose after just 20 h. R. capsulata gave a maximum hydrogen yield at 32 °C with acetate as a substrate, while R. capsulatus reached the optimum with glucose at 30 °C, pH 6.8; R. sphaeroides’ optimum was at 30 °C using lactic acid, and R. sulfidophilum produced a maximum of 0.42 mM/h at 72 and 168 h, but using a co-culture it went from 168–216 h. In general, the simpler the substrate, the better the yield and the shorter the time required. For example, volatile fatty acids and monosaccharides (glucose) performed better than disaccharides, and these performed better than more complex molecules, such as starch. The analysis carried out in the review is efficient in determining the most compatible bacteria for photo-fermentative hydrogen production under the specific conditions available in the particular laboratory or industry at that particular time of the year as the temperature plays a major role in hydrogen production efficiency, along with other conditions, such as bioreactor design and material, type of bioprocess, light intensity, and the substrate used.
4. Conclusions
Hydrogen is a promising way to produce clean and sustainable energy resources. Biological processes can be used to produce renewable and carbon neutral hydrogen. Photo-fermentation using PNSBs has been extensively studied as these bacteria can utilize substrates, such as simple sugars, volatile fatty acids, and even industrial and agricultural wastes, making it clear that these bacteria have unique abilities to utilize light energy and fermentation pathways to produce hydrogen. However, their relatively low hydrogen production rate, potential toxicity to non-sulfur photosynthetic bacteria due to their high substrate concentrations, and the need for light to facilitate the process have made it difficult to scale it up for large-scale application.
However, photo-fermentation also has advantages over other processes in terms of both chemical oxygen demand (COD) removal efficiency and hydrogen content. Optimal conditions, such as the substrate provided, pH, temperature, agitation speed, bioreactor volume, and light source and intensity play an important role in enhancing hydrogen production by different bacteria. In addition, the incorporation of immobilization techniques, genetic engineering, and other biotechnological techniques can also help to enhance the production of hydrogen by these bacteria. This review covers this diverse range of photo-fermentative microbes with their specific strains having different enzyme systems and metabolic pathways for hydrogen production. Each strain discussed has been compared with other strains to analyze the most suitable bacteria with the specific environmental and process conditions that have been preferred in several studies over the years and that are available during individual research. This can help to make future research more reliable and easier by tracking the availability of data that still needs to be optimized or worked on and can also help in environmental remediation.
Author Contributions
Conceptualization, S.G., A.F., A.L., L.G. and J.S.; methodology, S.G. and L.G.; software, S.G. and L.G.; validation, A.F., A.L. and L.G.; formal analysis, L.G. and J.S.; investigation, S.G. and L.G.; resources, L.G. and J.S.; data curation, A.F., A.L. and L.G.; writing—original draft preparation, S.G., A.F., L.G. and J.S.; writing—review and editing, S.G., A.F., A.L., L.G. and J.S.; visualization, L.G. and J.S.; project administration, L.G. and J.S.; funding acquisition, A.F., L.G. and J.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Fundação para a Ciência e a Tecnologia, FCT, project UIDB/00195/2020, PhD grant PRT/BD/154415/2023 awarded to Soumya Gupta, and research contract CEECINST/00016/2021/CP2828/CT0006 awarded to Annabel Fernandes under the scope of the CEEC Institutional 2021. This work was carried out as part of the project PR-H2CVAL4-C1-2022-0049 “Valorización de aguas residuales industriales para la generación de hidrógeno biológico (Hi2biO)” from IDAE, financed by the EU Next Generation Funds.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Oh, W.-C.; Park, J.-Y. Hydrogen gas generation of metal-activated carbon for fuel cells. J. Ind. Eng. Chem. 2007, 13, 578–584. [Google Scholar]
- Hren, R.; Vujanović, A.; Van Fan, Y.; Klemeš, J.J.; Krajnc, D.; Čuček, L. Hydrogen production, storage and transport for renewable energy and chemicals: An environmental footprint assessment. Renew. Sust. Energ. Rev. 2023, 173, 113113. [Google Scholar] [CrossRef]
- Nortez, K.B.; Movillon, J.L.; Alfafara, C.G.; Sevilla-Nastor, J.B.; Ventura, R.L.G.; Ventura, J.R.S. Optimization of photo-fermentative biohydrogen production in a mixed volatile fatty acid medium by Rhodobacter sp. MAY2: A response surface methodology (RSM) approach. Int. J. Hydrogen Energy 2024, 56, 844–852. [Google Scholar] [CrossRef]
- Jeong, T.Y.; Cha, G.C.; Yeom, S.H.; Choi, S.S. Comparison of hydrogen production by four representative hydrogen-producing bacteria. J. Ind. Eng. Chem. 2008, 14, 333–337. [Google Scholar] [CrossRef]
- Jain, I.P.; Lal, C.; Jain, A. Hydrogen storage in Mg: A most promising material. Int. J. Hydrogen Energy 2010, 35, 5133–5144. [Google Scholar] [CrossRef]
- Ivanenko, A.A.; Laikova, A.A.; Zhuravleva, E.A.; Shekhurdina, S.V.; Vishnyakova, A.V.; Kovalev, A.A.; Kovalev, D.A.; Trchounian, K.A.; Litti, Y.V. Biological production of hydrogen: From basic principles to the latest advances in process improvement. Int. J. Hydrogen Energy 2024, 55, 740–755. [Google Scholar] [CrossRef]
- Vargas, S.R.; Macêdo, W.V.; Trindade, L.F.; Zaiat, M. Influence of organic carbon source on hydrogen production and nutrient removal by microbial consortium in anaerobic photobioreactors. Int. J. Hydrogen Energy 2024, 54, 1160–1168. [Google Scholar] [CrossRef]
- Ren, N.Q.; Liu, B.F.; Ding, J.; Guo, W.Q.; Cao, G.L.; Xie, G.J. The effect of butyrate concentration on photo-hydrogen production from acetate by Rhodopseudomonas faecalis RLD-53. Int. J. Hydrogen Energy 2008, 33, 5981–5985. [Google Scholar] [CrossRef]
- Reungsang, A.; Zhong, N.; Yang, Y.; Sittijunda, S.; Xie, A.; Liao, Q. Hydrogen from Photo Fermentation. In Bioreactors for Microbial Biomass and Energy Conversion; Liao, Q., Chang, J., Herrmann, C., Xia, A., Eds.; Springer Nature: Singapore, 2018; pp. 221–317. [Google Scholar] [CrossRef]
- Xie, G.J.; Liu, B.F.; Ding, J.; Ren, H.Y.; Xing, D.F.; Ren, N.Q. Hydrogen production by photo-fermentative bacteria immobilized on fluidized bio-carrier. Int. J. Hydrogen Energy 2011, 36, 13991–13996. [Google Scholar] [CrossRef]
- Genç, Ş.; Koku, H. A preliminary techno-economic analysis of photo-fermentative hydrogen production. Int. J. Hydrogen Energy 2024, 52, 212–222. [Google Scholar] [CrossRef]
- Tiang, M.F.; Hanipa, M.A.; Abdul, P.M.; Jahim, J.M.; Mahmod, S.S.; Takriff, M.S.; Lay, C.; Resunsang, A.; Wu, S.Y. Recent advanced biotechnological strategies to enhance photo-fermentative biohydrogen production by purple non-sulfur bacteria: An overview. Int. J. Hydrogen Energy 2020, 45, 13211–13230. [Google Scholar] [CrossRef]
- Monir, M.U.; Aziz, A.A.; Ahmed, M.T.; Hasan, M.Y. Hydrogen energy–potential in developing countries. In Renewable Energy and Sustainability; Khan, I., Ed.; Elsevier: Amsterdam, The Netherlands, 2022; pp. 299–325. [Google Scholar] [CrossRef]
- Gest, H.; Kamen, M.D. Photoproduction of molecular hydrogen by Rhodospirillum rubrum. Science 1949, 109, 558–559. [Google Scholar] [CrossRef] [PubMed]
- Harwood, C.S. Degradation of aromatic compounds by purple nonsulfur bacteria. In The Purple Phototrophic Bacteria; Hunter, C.N., Daldal, F., Thurnauer, M.C., Beatty, J.T., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 577–594. [Google Scholar] [CrossRef]
- Assawamongkholsiri, T.; Reungsang, A. Photo-fermentational hydrogen production of Rhodobacter sp. KKU-PS1 isolated from an UASB reactor. Electron. J. Biotechnol. 2015, 18, 221–230. [Google Scholar] [CrossRef]
- Tao, Y.; He, Y.; Wu, Y.; Liu, F.; Li, X.; Zong, W.; Zhou, Z. Characteristics of a new photosynthetic bacterial strain for hydrogen production and its application in wastewater treatment. Int. J. Hydrogen Energy 2008, 33, 963–973. [Google Scholar] [CrossRef]
- Obeid, J.; Flaus, J.M.; Adrot, O.; Magnin, J.P.; Willison, J.C. State estimation of a batch hydrogen production process using the photosynthetic bacteria Rhodobacter capsulatus. Int. J. Hydrogen Energy 2010, 35, 10719–10724. [Google Scholar] [CrossRef]
- Liu, B.F.; Jin, Y.R.; Cui, Q.F.; Xie, G.J.; Wu, Y.N.; Ren, N.Q. Photo-fermentation hydrogen production by Rhodopseudomonas sp. nov. strain A7 isolated from the sludge in a bioreactor. Int. J. Hydrogen Energy 2015, 40, 8661–8668. [Google Scholar] [CrossRef]
- Bianchi, L.; Mannelli, F.; Viti, C.; Adessi, A.; De, P.R. Hydrogen-producing purple non-sulfur bacteria isolated from the trophic lake Averno (Naples, Italy). Int. J. Hydrogen Energy 2010, 35, 12216–12223. [Google Scholar] [CrossRef]
- Hosseini, S.S.; Aghbashlo, M.; Tabatabaei, M.; Younesi, H.; Najafpour, G. Exergy analysis of biohydrogen production from various carbon sources via anaerobic photosynthetic bacteria (Rhodospirillum rubrum). Energy 2015, 93, 730–739. [Google Scholar] [CrossRef]
- Cai, J.; Wang, G. Hydrogen production by a marine photosynthetic bacterium, Rhodovulum sulfidophilum P5, isolated from a shrimp pond. Int. J. Hydrogen Energy 2012, 37, 15070–15080. [Google Scholar] [CrossRef]
- Vignais, P.M.; Billoud, B.; Meyer, J. Classification and phylogeny of hydrogenases. FEMS Microbiol. Rev. 2001, 25, 455–501. [Google Scholar] [CrossRef]
- Larimer, F.W.; Chain, P.; Hauser, L.; Lamerdin, J.; Malfatti, S.; Do, L.; Land, M.L.; Pelletier, D.A.; Beatty, J.T.; Lang, A.S.; et al. Complete genome sequence of the metabolically versatile photosynthetic bacterium Rhodopseudomonas palustris. Nat. Biotechnol. 2004, 22, 55–61. [Google Scholar] [CrossRef]
- Liu, X.; Huang, L.; Rensing, C.; Ye, J.; Nealson, K.H.; Zhou, S. Syntrophic interspecies electron transfer drives carbon fixation and growth by Rhodopseudomonas palustris under dark, anoxic conditions. Sci. Adv. 2023, 7, 1852. [Google Scholar] [CrossRef]
- Androga, D.D.; Özgür, E.; Eroglu, I.; Gündüz, U.; Yücel, M. Photo-fermentative hydrogen production in outdoor conditions. In Hydrogen Energy—Challenges and Perspectives; IntechOpen Limited: London, UK, 2012; pp. 77–120. [Google Scholar] [CrossRef]
- Imhoff, J.F. Anoxygenic phototrophic bacteria from extreme environments. In Modern Topics in the Phototrophic Prokaryotes: Environmental and Applied Aspects; Hallenbeck, P.C., Ed.; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; pp. 427–480. [Google Scholar] [CrossRef]
- Surachat, K.; Kantachote, D.; Deachamag, P.; Wonglapsuwan, M. In silico genomic analysis of Rhodopseudomonas palustris strains revealed potential biocontrol agents and crop yield enhancers. Biol. Control 2022, 176, 105085. [Google Scholar] [CrossRef]
- Wang, Y.Z.; Liao, Q.; Zhu, X.; Li, J.; Lee, D.J. Effect of culture conditions on the kinetics of hydrogen production by photosynthetic bacteria in batch culture. Int. J. Hydrogen Energy 2011, 36, 14004–14013. [Google Scholar] [CrossRef]
- Wu, X.M.; Zhu, L.Y.; Zhu, L.Y.; Wu, L. Improved ammonium tolerance and hydrogen production in nifA mutant strains of Rhodopseudomonas palustris. Int. J. Hydrogen Energy 2016, 41, 22824–22830. [Google Scholar] [CrossRef]
- Sagir, E.; Alipour, S. Photo-fermentative hydrogen production by immobilized photosynthetic bacteria: Current perspectives and challenges. Renew. Sust. Energ. Rev. 2021, 141, 110796. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, L.; Ma, Y.; Huang, H.; Wang, G.; Ma, S.; Li, Z.; Han, M. Improving photo-fermentative hydrogen productivity of photosynthetic bacteria using a formulated Fe and Mo metal supplemented lignocellulosic substrate. Int. J. Hydrogen Energy 2024, 49, 516–531. [Google Scholar] [CrossRef]
- Guo, C.L.; Zhu, X.; Liao, Q.; Wang, Y.Z.; Chen, R.; Lee, D.J. Enhancement of photo-hydrogen production in a biofilm photobioreactor using optical fiber with additional rough surface. Bioresour. Technol. 2011, 102, 8507–8513. [Google Scholar] [CrossRef] [PubMed]
- Fu, Q.; Li, Y.; Zhong, N.; Liao, Q.; Huang, Y.; Xia, A.; Zhu, X.; Hou, Y. A novel biofilm photobioreactor using light guide plate enhances the hydrogen production, Int. J. Hydrogen Energy 2017, 42, 27523–27531. [Google Scholar] [CrossRef]
- Li, Y.; Zhong, N.; Liao, Q.; Fu, Q.; Huang, Y.; Zhu, X.; Li, Q. A biomaterial doped with LaB6 nanoparticles as photothermal media for enhancing biofilm growth and hydrogen production in photosynthetic bacteria. Int. J. Hydrogen Energy 2017, 42, 5793–5803. [Google Scholar] [CrossRef]
- Liao, Q.; Zhong, N.; Zhu, X.; Huang, Y.; Chen, R. Enhancement of hydrogen production by optimization of biofilm growth in a photobioreactor. Int. J. Hydrogen Energy 2015, 40, 4741–4751. [Google Scholar] [CrossRef]
- Wen, H.; Cao, G.; Xie, G.; Xing, D.; Yin, T.; Ren, N.; Liu, B. Improved photo-fermentative hydrogen production by biofilm reactor with optimizing carriers and acetate concentration. Int. J. Hydrogen Energy 2019, 44, 25151–25159. [Google Scholar] [CrossRef]
- Basak, N.; Jana, A.K.; Das, D. CFD modeling of hydrodynamics and optimization of photo-fermentative hydrogen production by Rhodopseudomonas palustris DSM 123 in annular photobioreactor. Int. J. Hydrogen Energy 2016, 41, 7301–7317. [Google Scholar] [CrossRef]
- Mabutyana, L.; Pott, R.W. Photo-fermentative hydrogen production by Rhodopseudomonas palustris CGA009 in the presence of inhibitory compounds. Int. J. Hydrogen Energy 2021, 46, 29088–29099. [Google Scholar] [CrossRef]
- Ross, B.S.; Pott, R.W.M. Hydrogen production by immobilized Rhodopseudomonas palustris in packed or fluidized bed photobioreactor systems. Int. J. Hydrogen Energy 2021, 46, 1715–1727. [Google Scholar] [CrossRef]
- Zhang, D.; Xiao, N.; Mahbubani, K.T.; del Rio-Chanona, E.A.; Slater, N.K.H.; Vassiliadis, V.S. Bioprocess modelling of biohydrogen production by Rhodopseudomonas palustris: Model development and effects of operating conditions on hydrogen yield and glycerol conversion efficiency. Chem. Eng. Sci. 2015, 130, 68–78. [Google Scholar] [CrossRef]
- Zhu, X.; Xie, X.; Liao, Q.; Wang, Y.; Lee, D. Enhanced hydrogen production by Rhodopseudomonas palustris CQK 01 with ultra-sonication pretreatment in batch culture. Bioresour. Technol. 2011, 102, 8696–8699. [Google Scholar] [CrossRef]
- Wang, Y.Z.; Xie, X.W.; Zhu, X.; Liao, Q.; Chen, R.; Zhao, X.; Lee, D.J. Hydrogen production by Rhodopseudomonas palustris CQK 01 in a continuous photobioreactor with ultrasonic treatment. Int. J. Hydrogen Energy 2012, 37, 15450–15457. [Google Scholar] [CrossRef]
- Azbar, N.; Cetinkaya, D.F.T. The effect of dilution and l-malic acid addition on bio-hydrogen production with Rhodopseudomonas palustris from effluent of an acidogenic anaerobic reactor. Int. J. Hydrogen Energy 2010, 35, 5028–5033. [Google Scholar] [CrossRef]
- Jamil, Z.; Annuar, M.S.M.; Ibrahim, S.; Vikineswary, S. Optimization of phototrophic hydrogen production by Rhodopseudomonas palustris PBUM001 via statistical experimental design. Int. J. Hydrogen Energy 2009, 34, 7502–7512. [Google Scholar] [CrossRef]
- Wang, Y.Z.; Liao, Q.; Zhu, X.; Chen, R.; Guo, C.L.; Zhou, J. Bioconversion characteristics of Rhodopseudomonas palustris CQK 01 entrapped in a photobioreactor for hydrogen production. Bioresour. Technol. 2013, 135, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Y.; Lee, C.M.; Chang, J.S. Hydrogen production by indigenous photosynthetic bacterium Rhodopseudomonas palustris WP3-5 using optical fiber-illuminating photobioreactors. Biochem. Eng. J. 2006, 32, 33–42. [Google Scholar] [CrossRef]
- Oh, Y.K.; Seol, E.H.; Lee, E.Y.; Park, S. Fermentative hydrogen production by a new chemoheterotrophic bacterium Rhodopseudomonas palustris P4. Int. J. Hydrogen Energy 2002, 27, 1373–1379. [Google Scholar] [CrossRef]
- Eroglu, I.; Sevinç, P.; Gündüz, U.; Grube, T.; Stolten, D.; Yücel, M. The Effect of Temperature and Light Intensity on Hydrogen Production by Rhodobacter capsulatus. In Proceedings of the 18th World Hydrogen Energy Conference 2010: WHEC 2010 Proceedings Forschungszentrum Jülich, Essen, Germany, 16–21 May 2010. [Google Scholar]
- Baccarini-Melandri, A.; Melandri, B.A. Partial resolution of the photophosphorylating system of Rhodopseudomonas capsulata. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1971; pp. 556–561. [Google Scholar] [CrossRef]
- Shi, X.Y.; Li, W.W.; Yu, H.Q. Key parameters governing biological hydrogen production from benzoate by Rhodopseudomonas capsulata. Appl. Energy 2014, 133, 121–126. [Google Scholar] [CrossRef]
- Shi, X.Y.; Yu, H.Q. Continuous production of hydrogen from mixed volatile fatty acids with Rhodopseudomonas capsulata. Int. J. Hydrogen Energy 2006, 31, 1641–1647. [Google Scholar] [CrossRef]
- Shi, X.Y.; Yu, H.Q. Conversion of individual and mixed volatile fatty acids to hydrogen by Rhodopseudomonas capsulata. Int. Biodeterior. Biodegrad. 2006, 58, 82–88. [Google Scholar] [CrossRef]
- Shi, X.Y.; Yu, H.Q. Response surface analysis on the effect of cell concentration and light intensity on hydrogen production by Rhodopseudomonas capsulata. Process Biochem. 2005, 40, 2475–2481. [Google Scholar] [CrossRef]
- Suresh, G.; Sasikala, C.; Ramana, C.V. Reclassification of Gemmobacter changlensis to a new genus as Cereibacter changlensis gen. nov., comb. nov. Int. J. Syst. Evol. Microbiol. 2015, 65, 794–798. [Google Scholar] [CrossRef]
- Suresh, G.; Lodha, T.D.; Indu, B.; Sasikala, C.; Ramana, C.V. Taxogenomics resolves conflict in the genus Rhodobacter: A two and half decades pending thought to reclassify the genus rhodobacter. Front. Microbiol. 2019, 10, 2480. [Google Scholar] [CrossRef]
- Basak, N.; Das, D. The prospect of purple non-sulfur (PNS) photosynthetic bacteria for hydrogen production: The present state of the art. World J. Microbiol. Biotechnol. 2007, 23, 31–42. [Google Scholar] [CrossRef]
- Ma, C.; Wang, X.; Guo, L.; Wu, X.; Yang, H. Enhanced photo-fermentative hydrogen production by Rhodobacter capsulatus with pigment content manipulation. Bioresour. Technol. 2012, 118, 490–495. [Google Scholar] [CrossRef]
- Abo-Hashesh, M.; Desaunay, N.; Hallenbeck, P.C. High yield single stage conversion of glucose to hydrogen by photo-fermentation with continuous cultures of Rhodobacter capsulatus JP91. Bioresour. Technol. 2013, 128, 513–517. [Google Scholar] [CrossRef]
- Ghosh, D.; Sobro, I.F.; Hallenbeck, P.C. Optimization of the hydrogen yield from single-stage photo-fermentation of glucose by Rhodobacter capsulatus JP91 using response surface methodology. Bioresour. Technol. 2012, 123, 199–206. [Google Scholar] [CrossRef]
- Elkahlout, K.; Sagir, E.; Alipour, S.; Koku, H.; Gunduz, U.; Eroglu, I.; Yucel, M. Long-term stable hydrogen production from acetate using immobilized Rhodobacter capsulatus in a panel photobioreactor. Int. J. Hydrogen Energy 2019, 44, 18801–18810. [Google Scholar] [CrossRef]
- Boran, E.; Özgür, E.; Van Der Burg, J.; Yücel, M.; Gündüz, U.; Eroglu, I. Biological hydrogen production by Rhodobacter capsulatus in solar tubular photo bioreactor. J. Clean. Prod. 2010, 18, S29–S35. [Google Scholar] [CrossRef]
- Androga, D.D.; Sevinç, P.; Koku, H.; Yücel, M.; Gündüz, U.; Eroglu, I. Optimization of temperature and light intensity for improved photo-fermentative hydrogen production using Rhodobacter capsulatus DSM 1710. Int. J. Hydrogen Energy 2014, 39, 2472–2480. [Google Scholar] [CrossRef]
- Sagir, E.; Alipour, S.; Elkahlout, K.; Koku, H.; Gunduz, U.; Eroglu, I.; Yucel, M. Scale-up studies for stable, long-term indoor and outdoor production of hydrogen by immobilized Rhodobacter capsulatus. Int. J. Hydrogen Energy 2017, 42, 22743–22755. [Google Scholar] [CrossRef]
- Magnin, J.P.; Deseure, J. Hydrogen generation in a pressurized photobioreactor: Unexpected enhancement of biohydrogen production by the phototrophic bacterium Rhodobacter capsulatus. Appl. Energy 2019, 239, 635–643. [Google Scholar] [CrossRef]
- Boran, E.; Özgür, E.; Yücel, M.; Gündüz, U.; Eroglu, I. Biohydrogen production by Rhodobacter capsulatus in solar tubular photobioreactor on thick juice dark fermenter effluent. J. Clean. Prod. 2012, 31, 150–157. [Google Scholar] [CrossRef]
- Boran, E.; Özgür, E.; Yücel, M.; Gündüz, U.; Eroglu, I. Biohydrogen production by Rhodobacter capsulatus Hup− mutant in pilot solar tubular photobioreactor. Int. J. Hydrogen Energy 2012, 37, 16437–16445. [Google Scholar] [CrossRef]
- Koku, H.; Eroğlu, I.; Gündüz, U.; Yücel, M.; Türker, L. Aspects of the metabolism of hydrogen production by Rhodobacter sphaeroides. Int. J. Hydrogen Energy 2002, 27, 1315–1329. [Google Scholar] [CrossRef]
- Zagrodnik, R.; Łaniecki, M. Hydrogen production from starch by co-culture of Clostridium acetobutylicum and Rhodobacter sphaeroides in one step hybrid dark- and photo-fermentation in repeated fed-batch reactor. Bioresour. Technol. 2017, 224, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Pattanamanee, W.; Choorit, W.; Kantachote, D.; Chisti, Y. Repeated-batch production of hydrogen using Rhodobacter sphaeroides S10. Int. J. Hydrogen Energy 2012, 37, 15855–15866. [Google Scholar] [CrossRef]
- Al-Mohammedawi, H.H.; Znad, H. Impact of metal ions and EDTA on photo-fermentative hydrogen production by Rhodobacter sphaeroides using a mixture of pre-treated brewery and restaurant effluents. Biomass Bioenergy 2020, 134, 105482. [Google Scholar] [CrossRef]
- Hu, J.; Yang, H.; Wang, X.; Cao, W.; Guo, L. Strong pH dependence of hydrogen production from glucose by Rhodobacter sphaeroides. Int. J. Hydrogen Energy 2020, 45, 9451–9458. [Google Scholar] [CrossRef]
- Akroum-Amrouche, D.; Abdi, N.; Lounici, H.; Mameri, N. Effect of physico-chemical parameters on biohydrogen production and growth characteristics by batch culture of Rhodobacter sphaeroides CIP 60.6. Appl. Energy 2011, 88, 2130–2135. [Google Scholar] [CrossRef]
- Krujatz, F.; Härtel, P.; Helbig, K.; Haufe, N.; Thierfelder, S.; Bley, T.; Weber, J. Hydrogen production by Rhodobacter sphaeroides DSM 158 under intense irradiation. Bioresour. Technol. 2015, 175, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Basak, N.; Das, D. Photo-fermentative hydrogen production using purple non-sulfur bacteria Rhodobacter sphaeroides O.U.001 in an annular photobioreactor: A case study. Biomass Bioenergy 2009, 33, 911–919. [Google Scholar] [CrossRef]
- Ma, H.; Zheng, X.; Yang, H. Enhancement on hydrogen production performance of Rhodobacter sphaeroides HY01 by overexpressing fdxN. Int. J. Hydrogen Energy 2018, 43, 17082–17090. [Google Scholar] [CrossRef]
- Al-Mohammedawi, H.H.; Znad, H.; Eroglu, E. Synergistic effects and optimization of photo-fermentative hydrogen production of Rhodobacter sphaeroides DSM 158. Int. J. Hydrogen Energy 2018, 43, 15823–15834. [Google Scholar] [CrossRef]
- Seifert, K.; Waligorska, M.; Laniecki, M. Brewery wastewaters in photobiological hydrogen generation in presence of Rhodobacter sphaeroides O.U. 001. Int. J. Hydrogen Energy 2010, 35, 4085–4091. [Google Scholar] [CrossRef]
- Jaime-Pérez, N.; Kaftan, D.; Bína, D.; Bokhari, S.N.H.; Shreedhar, S.; Küpper, H. Mechanisms of sublethal copper toxicity damage to the photosynthetic apparatus of Rhodospirillum rubrum. Biochim. Biophys. Acta Bioenerg. 2019, 1860, 640–650. [Google Scholar] [CrossRef]
- Dadak, A.; Aghbashlo, M.; Tabatabaei, M.; Younesi, H.; Najafpour, G. Exergy-based sustainability assessment of continuous photobiological hydrogen production using anaerobic bacterium Rhodospirillum rubrum. J. Clean. Prod. 2016, 139, 157–166. [Google Scholar] [CrossRef]
- Maeda, I.; Miyasaka, H.; Umeda, F.; Kawase, M.; Yagi, K. Maximization of hydrogen production ability in high-density suspension of Rhodovulum sulfidophilum cells using intracellular poly(3-hydroxybutyrate) as sole substrate. Biotechnol. Bioeng. 2003, 81, 474–481. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Wang, G. Photo-biological hydrogen production by an acid-tolerant mutant of Rhodovulum sulfidophilum P5 generated by transposon mutagenesis. Bioresour. Technol. 2014, 154, 254–259. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Guan, Y.; Jia, T.; Yang, J.; Hu, Y.; Li, P.; Duan, Y.; Zhang, L.; Yu, P. Hydrogen production from high slat medium by co-culture of Rhodovulum sulfidophilum and dark fermentative microflora. Int. J. Hydrogen Energy 2018, 43, 10959–10966. [Google Scholar] [CrossRef]
- Keskin, T.; Hallenbeck, P.C. Hydrogen production from sugar industry wastes using single-stage photo-fermentation. Bioresour. Technol. 2012, 112, 131–136. [Google Scholar] [CrossRef]
- Hallenbeck, P.C. Chapter 7—Photo-fermentative Biohydrogen Production. In Biohydrogen; Pandey, A., Chang, J.S., Hallenbecka, P.C., Larroche, C., Eds.; Elsevier: Amsterdam, The Netherlands, 2013; pp. 145–159. [Google Scholar] [CrossRef]
Figure 1.
Schematic illustration representing hydrogen production by various pathways (adapted from Ref. [12]).
Figure 1.
Schematic illustration representing hydrogen production by various pathways (adapted from Ref. [12]).
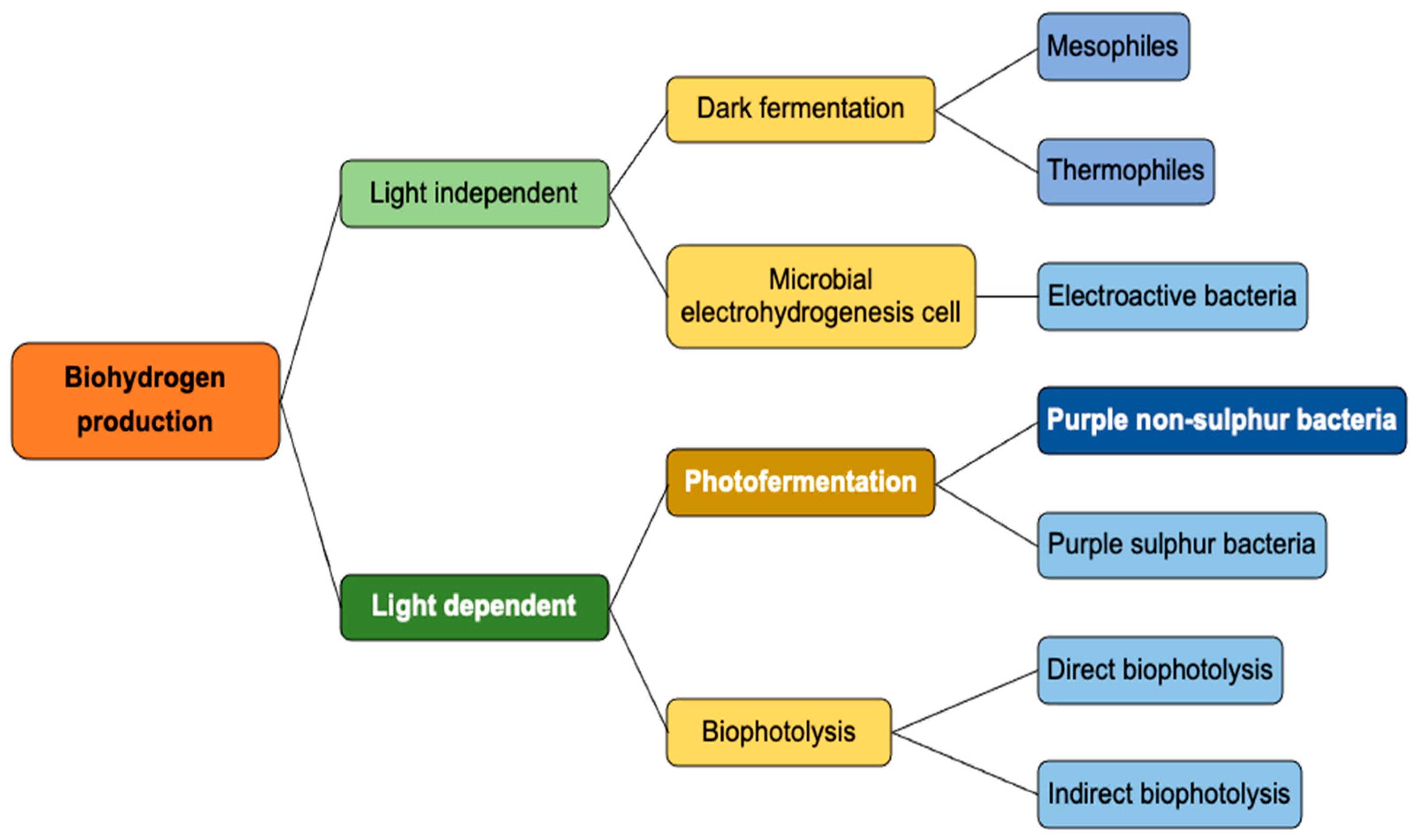
Figure 2.
Bacteria mostly studied for photo-fermentative hydrogen production.

Figure 3.
A general outline of hydrogen production by photo-fermentative bacteria.

Figure 4.
Photo-fermentative hydrogen production in PNSBs. Adapted with permission from Ref. [26]. 2012, Androga, Özgür, Eroglu, Gündüz, Yücel—IntechOpen Limited.
Figure 4.
Photo-fermentative hydrogen production in PNSBs. Adapted with permission from Ref. [26]. 2012, Androga, Özgür, Eroglu, Gündüz, Yücel—IntechOpen Limited.

Figure 5.
Ranges of maximum hydrogen production rates from different bacterial strains.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Hydrogen-producing Rhodopseudomonas palustris strains in PBRs.
| Ref. | Strain | Substrate (Concentration, mM) | Conditions | Bioreactor | Light Source | Irradiance (W/m2) | Hydrogen Production Rate (mM/h) |
|---|---|---|---|---|---|---|---|
| [33] | CQK 01 | Glucose (56) | pH 7.0, 30 °C | PBR made of polymethyl methacrylate (PMMA), continuous | Tungsten lamp | 31.6 | 1.75 |
| [34] | CQK 01 | Glucose (56) | pH 7.0, 25 °C | PBR 8 × 200 × 200 mm3 of PMMA, continuous | Light guided plate | 10.1 | 1.1 |
| SiO2 chitosan-medium-LGP | 7.8 | 1.4 | |||||
| [35] | CQK 01 | Glucose (56) | pH 7.0, 30 °C | Nanobiofilm of LaB6 powder | LED | 145.7 | 0.12 |
| [36] | CQK 01 | Glucose (55) | pH 7.0, 26.5–31.5 °C | Cylindrical PBR of PMMA with chitosan-medium coated optical fiber | Tungsten lamp | 31.6 | 2.72 |
| [37] | A7 | Acetate (68) | pH 7.0, 35 °C | Anaerobic PBR | Tungsten lamp | 1.05 | |
| [38] | DSM 123 | Malate (20) | pH 6.8, 33 °C | Triple-jacketed vertical annular PBR (1 L) | Tungsten lamp | 15 ± 1.1 | 0.31 |
| [39] | CGA009 | Glycerol (10) | pH 7.0, 35 °C | 250 mL glass bottles (200 mL working volume) | Tungsten lamp | 200 ± 5 | 0.25 |
| [40] | NCIMB 11774 | Glycerol (50) | pH 7.0, 28 ± 1 °C | Fluidized bed PBR (1 L) | Tungsten lamp | 100 | 3.23 |
| [41] | NCIB 11774 | Glycerol (10) | 25 ± 2 °C | Piecewise droop model, batch | Tungsten lamp | 174 | 1.54 |
| [42] | CQK 01 | Glucose (50) | pH 7.0, 30 °C | PBR of PMMA 100 × 50 × 100 mm3 | Monochromatic LED | 47.4 | 0.54 |
| [43] | CQK 01 | Glucose (50) | pH 7.0, 30 °C | PBR 120 × 60 × 190 mm3 | Monochromatic LED | 47.4 | 0.82 |
| [44] | DSM 127 | L-malic acid (30), 50% L-malic acid and 50% raw effluent | pH 6.9, 31 °C | 250 mL serum bottles (200 mL working volume), batch | Halogen lamp | 47.4 | 0.35 |
| [45] | PBUM001 | Palm oil mill effluent (POME) | pH 7.0, 30 °C | 121 mL serum bottles (50 mL working volume) | Tungsten lamp | 31.6 | 0.44 |
| [46] | CQK 01 | Glucose (44.7) | pH 7.0, 30 °C | Flat panel PBR 100 × 40 × 200 mm3, continuous | LEDs | 47.4 | 2.5 |
| [47] | WP3–5 | Acetate (14.8) | pH 7.1, 32 °C | Sealed glass vessel (500 mL) | Tungsten/halogen lamp | 95 | 0.76 |
| [48] | P4 | Glucose (27.8) | pH 7.0, 30 °C | 165 mL serum bottle (50 mL working volume), batch | Not described | Not described | 3.2 |
Table 2.
Hydrogen-producing R. capsulata strains in PBRs.
| Ref. | Substrate (Concentration, mM) | Conditions | PBR | Light Source | Irradiance (W/m2) | Hydrogen Production Rate (mM/h) |
|---|---|---|---|---|---|---|
| [51] | Benzoate (5) | pH 7.0, 30 °C | 150 mL glass PBR | Tungsten lamp | 31.6 | 0.36 |
| [52] | Acetate (30) + propionate (3) + butyrate (12) | pH 6.8, 35 °C | 1500 mL, continuous | Tungsten lamp | 39.5 | 0.56 |
| [53] | Individually and mixture: acetate (32) + propionate (5) + butyrate (10) | pH 6.8, 32 °C | Glass vials (300 mL working volume) | Not described | 31.6 | 0.79 (acetate), 0,71 (propionate), 0.26 (butyrate), 0.65 (mixture) |
| [54] | Acetate (30) + propionate (3) + butyrate (11) | pH 7.0, 31 °C | 300 mL glass PBR | Tungsten lamp | 39.5 | 0.31 |
Table 3.
Hydrogen-producing R. capsulatus strains used in PBRs.
| Ref. | Strain | Substrate (Concentration, mM) | Conditions | PBR | Light Source | Irradiance (W/m2) | Hydrogen Production Rate (mM/h) |
|---|---|---|---|---|---|---|---|
| [59] | JP91 | Glucose (56) | pH 6.8, 30 °C | 350 mL, continuous | Tungsten lamp | Not described | 1.6–2.3 |
| [60] | JP91 | Glucose (35) | pH 6.8, 30 °C | 100 mL serum bottles, batch | Tungsten lamp | 175 | 2.6 |
| [61] | DSM 1710, YO3 | Acetate (60) | pH 6.7, 30–32 °C | 1.4 L PMMA reactor, batch | Tungsten lamp | 200 | 0.75–1.3 |
| [62] | DSM 1710 | Acetic acid (40) | pH < 8, 10–35 °C | 80 L tubular PBR fed-batch | Artificial light source | >90 | 0.52 |
| [63] | DSM 1710 | Lactic acid (7.5) | pH 6.4, 27.5 °C | 55 mL glass bottle | Tungsten lamp | 287 | 0.566 |
| [64] | YO3 | Sucrose (5 indoors, 10 outdoors) | pH 7.5, 30 °C | 3.64 L PBR of two PMMA with PVC frame, batch | Tungsten lamps (indoors), sunlight (outdoors) | 200 | 0.73 (indoors), 0.87 (outdoors) |
| [65] | Wild type B10 | Lactate (35) | pH 6.9, 30 °C | 350 mL cylindrical glass vessel | Na lamps, LEDs | 812, 479 | 0.26–0.46 |
| [66] | DSM 1710 | Acetate (20) | pH < 8, <35 °C | Tubular PBR (90 L), fed-batch | Halogen lamp | 200 | 0.15 |
| [67] | MT1131 | Acetate (20) | pH < 8, <30 °C | Tubular PBR (90 L), fed-batch | Halogen lamp | 200 | 0.20 |
Table 4.
Hydrogen-producing R. sphaeroides strains used in PBRs.
| Ref. | Strain | Substrate (Concentration, mM) | Conditions | PBR | Light Source | Irradiance (W/m2) | Hydrogen Production Rate (mM/h) |
|---|---|---|---|---|---|---|---|
| [69] | O.U.001 | Acetic (4) + butyric acid (6) | pH 7.0, 32 °C | Batch mode and fed-batch | Ultra-Vitalux lamps (Osram, Premstaetten, Austria) | 192 | 0.89 |
| [70] | S10 | Oil palm + mixture of glucose (36.13), xylose (102.56), acetic acid (48.81) | pH 7.0, 35 °C | 500 mL serum cylinder reactors, batch | Tungsten lamps | 79 | 1.4 |
| [71] | DSM 158 | Brewery effluent (30%) | pH 7.2, 30 °C | 120 mL clear glass | Halogen lamp | 126 | 0.74 |
| Restaurant effluent (70%) | pH 7.6, 30 °C | ||||||
| [72] | HY01 | Glucose (40) | pH 6.9, 35 °C | 350 mL | Halogen lamp | 31.6 | 5.6 |
| [73] | CIP 60.6 | Lactate (50) | pH 7.5, 30 °C | Column reactor | Tungsten lamp | 35.6 | 1.8 |
| [74] | DSM 158 | Lactic acid (40) | pH 7.0, 30 °C | Single-walled glass vessel, batch | Halogen lamp | 2250 | 8.7 |
| 20 L feed tank, continuous | 7.4 | ||||||
| [75] | O.U.001 | DL malic acid (15) | pH 6.8, 33 °C, | Triple-jacketed vertical glass PBR (1 L) | Lumine tubular light (Xiamen, China) | 15 | 0.29 |
| [76] | HY01 | Acetate (25) + butyrate (34) | pH 7.0–8.2, 30 °C | 30 mL syringes, batch | Tungsten lamp | 31.6 | 7.0 |
| [77] | DSM 158 | DL malic acid (7.5) | pH 6.5–8.0, 30 °C | 120 mL glass PBR, batch | Halogen lamp | 35–185 | 1.9 |
| [78] | O.U.001 | L-malic acid (15) | pH 7.0–7.2, 28 °C | 25 mL sodium glass vials | Mercury-tungsten lamp | 116 | 1.2 |
| Biebl and Pfenning media + 20% wastewater | 1.1 |
Table 5.
Comparison of different bacterial strains with their maximum hydrogen production rate.
| Ref. | Bacteria | Strain | Substrate (mM) | Maximum Hydrogen Production Rate (mM/h) |
|---|---|---|---|---|
| [36] | R. palustris | CQK01 | Glucose (55) | 2.72 |
| [40] | NCIMB 11774 | Glycerol (50) | 3.23 | |
| [46] | CQK01 | Glucose (44.7) | 2.5 | |
| [48] | P4 | Glucose (27.8) | 3.2 | |
| [53] | R. capsulata | Acetate (32) | 0.79 | |
| Propionate (5) | 0.71 | |||
| [59] | R. capsulatus | JP91 | Glucose (56) | 1.6–2.3 |
| [60] | JP91 | Glucose (35) | 2.6 | |
| [74] | R. sphaeroides | DSM 158 | Lactic acid (40) | 8.7 |
| [76] | HY01 | Acetate (25) + butyrate (34) | 7.0 | |
| [21] | R. rubrum | Acetate | 0.50 | |
| [83] | R. sulfidophilum | TH-102 | 0.42 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gupta, S.; Fernandes, A.; Lopes, A.; Grasa, L.; Salafranca, J. Photo-Fermentative Bacteria Used for Hydrogen Production. Appl. Sci. 2024, 14, 1191. https://doi.org/10.3390/app14031191
AMA Style
Gupta S, Fernandes A, Lopes A, Grasa L, Salafranca J. Photo-Fermentative Bacteria Used for Hydrogen Production. Applied Sciences. 2024; 14(3):1191. https://doi.org/10.3390/app14031191
Chicago/Turabian StyleGupta, Soumya, Annabel Fernandes, Ana Lopes, Laura Grasa, and Jesús Salafranca. 2024. "Photo-Fermentative Bacteria Used for Hydrogen Production" Applied Sciences 14, no. 3: 1191. https://doi.org/10.3390/app14031191
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.