
Laminar Ulva Species: A Multi-Tool for Humankind?




Marine Resources, Conservation and Technology, Marine Algae Lab, CFE—Centre for Functional Ecology: Science for People & Planet, Department of Life Sciences, University of Coimbra, 3000-456 Coimbra, Portugal
*
Authors to whom correspondence should be addressed.
Appl. Sci. 2024, 14(8), 3448; https://doi.org/10.3390/app14083448
Submission received: 21 March 2024
/
Revised: 16 April 2024
/
Accepted: 17 April 2024
/
Published: 19 April 2024
(This article belongs to the Section Ecology Science and Engineering)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Featured Application
Firstly, it is an extensive review of one of the most talked about “hot topics” today. Subsequently, it could be a complete guide to the cultivation and use for aquaculture of one of the most cosmopolitan species ever, as well as a guide to controlling “green tides”.
Abstract
Green algae, phylum Chlorophyta, due to their green appearance as higher plants, are seen as one of the raw materials to be widely used by humanity for different purposes. How can these different purposes achieve ONU Sustainable Development Goals? The genus Ulva sp. is widely distributed through all continents, tolerating different ecosystems (freshwater and marine), different intensities of light, temperature, and salinity. The Ulva sp. life cycle is isomorphic and biphasic type, also affected by biotic factors such as thallus age, phytohormones, microbiome, sporulation inhibitors and metabolomic. Due to that, types of farming can be implemented depending on the cultivation method and it is final biomass exploitation. Thus, this critical review analyzes the laminar Ulva species from the ecology and demonstrates that the seaweed biomass application, may make significant contributions to marine ecosystems, humans, aquaculture, and biotechnological innovation, indicating its importance in both environmental and socioeconomic contexts based on experiments across the world, time and critical thinking. This means that explaining the actual road and future roads of laminar Ulva into a multi-tool development from humankind welfare. With right management of resources and human empowerment, Ulva sp. products can be produced facing climate change and support different industries. However, responsible management of Ulva populations and farming is essential to prevent overgrowth, green floods, and maintain environmental equilibrium.
Keywords:
Ulva sp.; life cycle; cultivation; reproduction; gametogenesis; abiotic factors; biotic factors; feedstock; future1. Introduction
According to UN data, the Homo sapiens species reached 8 billion inhabitants on 15 November 2022. Despite past progress, our planet suffers from degraded ecosystems and loss of biodiversity, and humanity faces a climate, financial, and food crisis with hunger and malnutrition. This increase calls for an urgent transformation in agri-food systems and a focus on sustainability. Therefore, to contribute to the 7 Sustainable Development Goals (SDG), we began to focus on aquaculture and algae [1].
Algae are aquatic organisms that (with few exceptions to the rule) develop photosynthesis and are oxygenated autotrophs that are typically smaller and less structurally complex than plants. They are characterized by their cell wall nature, pigment composition (such as chlorophyll a, b, c, e; phycobilin, xanthophyll, carotenoids, peridinin and fucoxanthin, phycoerythrin), and reserve substance types (paramylon, starch, chrysolaminarin, laminarin, and mannitol). The “body” of algae, known as “thallus” (“thalli” in plural), is regularly seen in diverse forms [2].
Green algae, often known as chlorophytes, have the same green hue as higher plants. This occurs because green algae have chlorophyll b and their principal storage product is starch, which accumulates in the chloroplast rather than the cytoplasm and is frequently coupled with a pyrenoid. Ulva sp. is one of the most prevalent green macroalgae species in the world [2,3].
An Old Latin name for “edge,” Ulva sp. is portrayed by a foliaceous thalli with polygonal cells grouped into two defined layers forming conspicuous flat sheets up to 1 m long, irregularly lobed, wedge-shaped, linear, lanceolate, oblanceolate, or deeply divided into linear laciness [2,4]. As will be discussed later, species of the genus Ulva show ease of adaptation to fluctuations in abiotic and biotic conditions and can be found in a greater proportion in marine and estuarine environments. This means it is ideal for growing in virtually any location, according to Ben-Ari et al. [5], to obtain biomass for commercial interests.
Furthermore, because of its rapid development rate, this species is facing ecological threats. If the aquatic estuary environment or sea becomes entropized [6], Ulva sp. can overrun another species and cause an environmental problem [7,8,9,10]. It is also important to note and refer to Abdel-Fattah and Edrees [11] who noted that the nutritional value might change depending on environmental conditions (such as temperature, salinity, light, and nutrients) and mineral availability. The chemical composition of lipids, fatty acids, polysaccharides, and pigments in this seaweed has been discovered to alter dramatically. Seaweed farming helps to reduce environmental strain in natural habitats, ensure food safety, and promote a blue-green economy [12]. Nonetheless, little progress has been made beyond the laboratory scale in the establishment of controlled commercial production systems. It is also worth noting that the nutritional value might change depending on environmental conditions (including temperature, salinity, light, and nutrients) and mineral availability. The chemical makeup of this seaweed (pigments, polysaccharides, fatty acids, lipids) has been discovered to fluctuate dramatically [11]. However, minimal effort has been undertaken beyond the laboratory scale to build regulated commercial farming systems.
The objective of this critical review was to understand fully the exploitation circuit and all the back-to-back understanding of the potential of laminar Ulva species to be a keystone for humankind’s welfare, from the ecological and environmental data to the seaweed-based products and tools for humans (Figure 1).
2. The Biology of Ulva sp.
Ulva is one of the few genera of the green lineage with freshwater and marine species, living on rocky coastal shores, connected to substrates through rhizoid branches, or occurring in floating masses. They end up being rapid colonizers of bare substrates, such as jetty walls and surfaces of maritime vessels, where they also facilitate colonization by other algae. Large growths can also occur in salt marshes [2].
As they present tolerance to changes in salinity, the occurrence of forms adapted to high and low temperatures, the ability to use bicarbonate as a source of inorganic carbon, very high rates of light-saturated photosynthesis, high reproductive rates, seasonality in growth with abundance in winter and induction of swarms in summer, and the ability to flagellate cells to adhere to substrates quickly, they are considered fast-growing opportunists that can dominate coastal regions influenced by nutrient-rich effluents. Ulva species are distributed across seven continents: Asia (56), followed by Australia (40), Europe (38), North America (34), Africa (31), South America (20), and Antarctica (12). Of these, 18 are endemic to Asia, 11 to Australia, 9 to Europe, 6 to Africa, 2 to North America, and 1 to South America [2,4].
Rotting masses of these algae produce harmful H2S (Hydrogen Sulfide), and there is some concern that the continued increase in ulvaleans biomass could increase the production of ozone-depleting halocarbons by seaweed [2].
Ulva is governed by a complex process, and the type of life cycle pattern has been deciphered in 14 different species to date. Two of the key factors are to understand the seaweed life cycle and the factors that can influence the species for better or worse. This is a critical step in the successful cultivation and maintenance of the seaweed profile.
2.1. Reproduction
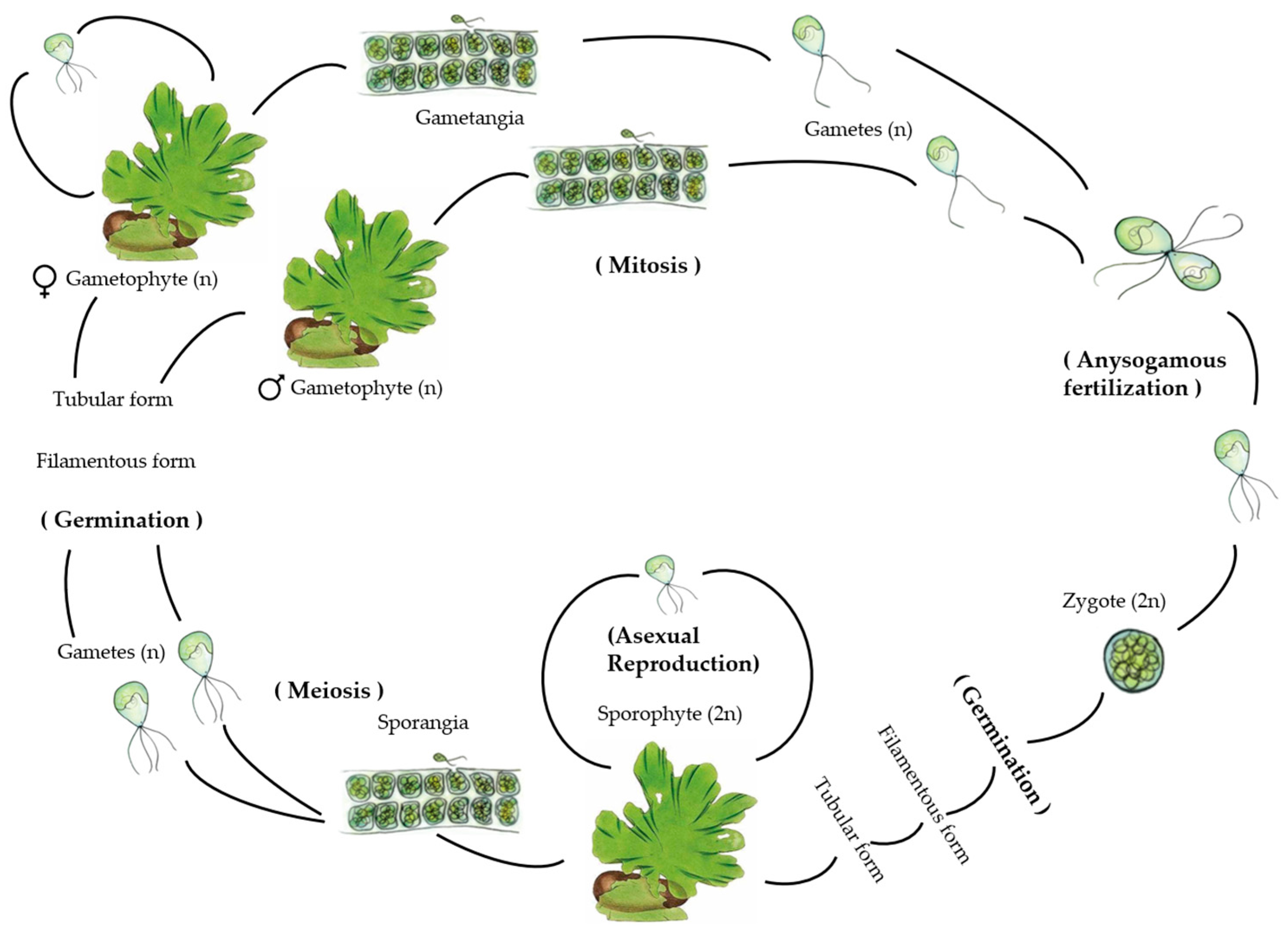
Reproduction can occur both sexually and asexually (Figure 2). The life cycle is isomorphic and biphasic, where the multicellular adult sporophyte and gametophytic phases are morphologically similar but genetically different, as sporophytes contain twice the level of nuclear DNA (2C) and number of chromosomes (2N) per cell as gametophytes (1C or 1N) [2,4].
Sexual reproduction involves the production of haploid biflagellate gametes that fuse during copulation, resulting in the formation of a diploid zygote. This zygote then develops into a diploid multicellular adult sporophyte thallus. As these typical vegetative cells mature, they transition into zoospore mother cells, which undergo meiosis to produce haploid quadriflagellate zoospores. These zoospores germinate and give rise to haploid male and female isomorphic adult gametophytic thalli through mitosis. At maturity, these thalli once again produce their respective gametes through mitosis. This reproductive process is termed a “haplodiplontic” life cycle (Figure 2). Both zoospores and gametes have no cell walls and are described as naked and are produced mainly in cells at the end of the thallus [2,4].
A difference in the duration of the division stages was reported; for example, the 4-cell and 8-cell states last longer in the gametophyte than in the sporophyte, which is evident from the greater DNA synthesis during this period in the gametophyte [13]. Gametophytes’ multicellularity allows them to create significant quantities of photosynthesis and so produce a bigger number of gametes (potential sporophytes) than unicellular haploids. Ulvaleans’ widespread ecological success is most likely aided by the alternation of multicellular generations [2].
Algae of the genus Ulva are opportunistic and have similar reproductive characteristics compared to “selected” species: many spores released by the reproductive cycle, small spores (approximately 10 μm), a wide range of dispersive potential (flagellates), and fast growth and short life cycles.
Liu et al., in 2022 [14], conducting research on gene expression during the various stages of gamete formation in Ulva sp., found that 8296 distinct genes (62.2% of annotated genes) were expressed during gametogenesis, with relatively low variation in the total number of expressed genes between time points, ranging from 7146 (0 h) to 7949 expressed genes (72 h). They also identified 6056 differentially expressed genes (DEGs) between any two of the five time periods (0 h, 6 h, 24 h, 48 h, 72 h) during gametogenesis, accounting for 45% of the annotated genome. They also obtained consistency with results from Stratmann et al. [15], where the analysis of DNA synthesis carried out during gametogenesis observed that DNA replication begins 25 h after induction.
In 2018, De Clerck et al. [16] documented a haploid genome of 98.5 Mbp (comprising 12,924 protein-coding genes) in Ulva compressa, previously known as U. mutabilis. Through the construction of a phylogenetic tree using a concatenated alignment of 58 nuclear protein-coding genes (totaling 42,401 amino acids), Ulva was determined to be closely related to Chlorophyta. The identification of specific features such as ecotypic genetic variation in bloom-forming organisms will aid in our comprehension of the molecular pathways involved in growth and reproduction in response to environmental conditions [13,16].
2.2. Influence of Factors
Ulva reproductive processes are affected by abiotic factors, such as light, temperature, and desiccation, but the mode of action for each factor is different and always works in combination and not alone. Furthermore, biological factors such as algal microbial interaction, thallus age, concentrations of growth regulators (hormonal second messenger), and sporulation, as well as the existence of inhibitors also play a critical role in maturation.
2.2.1. Biotic Factors
Biotic factors are the living components of an ecosystem, which include all live species and their interactions. These elements have a significant impact on ecosystem functioning and structure. Studying the interactions between these biotic elements is critical for understanding ecosystem dynamics, population ecology, and the general balance between organisms in various contexts. Ecosystems are structured by intricate interactions and interdependence between their living components, and a shift in one area of the system’s functioning can have far-reaching consequences for the whole ecosystem.
To utilize the laminar Ulva sp., we must first understand how the biotic factors affect their metabolism, health, and development.
Thallus Age
Anywhere within a stalk of Ulva sp., the formation of zoospores may occur; however, the gamete is formed close to the margin of the foliage by the repeated bipartition of the protoplast of a cell according to Lüning et al. [17].
The initial cleavage takes place along the surface of the stalk, followed by a second cleavage perpendicular to the first. Conical projections, referred to as papilla, form at the outer end. Subsequently, the microtubule apparatus is formed, and the nucleus divides into subsequent gamete nuclei. According to Katsaros et al. [18], cytokinesis gives rise to 16 oval-shaped gametes, which are ready to be released. However, divisions continue to occur, and the zoospore mother cells divide to form zoospores. Thallus cells thus enter sporogenesis or gametogenesis based on their maturity and the level of sporulation inhibitors. Stratmann et al. [15] registered that the completion of reproductive induction before the elimination of gametes or zoospores is marked by a change in thallus color from yellow green (fully vegetative state) to dark olive green. The white color is evidence of the release of reproductive cells [13].
Phytohormones
Hormones are very small signal molecules that could control several physiological processes, such as helping steer the growth, cell elongation, adventitious bud formation, and development of the plant. Hormones are usually produced in one part of the plant and transferred to another part of the plant to initiate physiological changes. The most important hormones are auxins, cytokinins, and gibberellins [19].
Auxins are small organic compounds constituting an aromatic ring and a side chain with a carboxylic acid group. They are also involved in multiple developmental processes in planta, such as key regulators of cell division and elongation, tissue differentiation, tropisms, apical dominance, senescence, abscission, and flowering. This is reported in bacteria, animal tissues, fungi, algae, and higher plants. Because of its versatile influence, auxin has been long recognized as “the plant growth hormone” [19].
Cytokinins have a variety of activities in algae, including cell division, differentiation, and morphogenesis; rhizoid and thallus growth; chloroplast formation; photosynthesis control; metabolite generation; and stress tolerance. Authors like Jones et al. [20], Woodward and Bartel [21], and Žižková et al. [22] reported that high levels of auxin, however, may inhibit cytokinin biosynthesis via negative feedback, especially in mature algal cells or tissues, in higher plants.
Gibberellins are involved in cell expansion, activation of cell division, increased metabolism, and the accumulation of pigments and lipids.
Abscisic acid (ABA) is often referred to as a stress hormone or negative regulator whose concentrations increase under various biotic and abiotic stresses, controlling embryogenesis, seed dormancy, and senescence and stimulating physiological responses for stress adaptation such as stomatal closure. In algae, this can occur as two types: endogenous as a response to unfavorable environments such as salinity, pH, oxidative, temperature, and nitrogen-deficient stresses and exogenous downregulating auxin signaling and repressing growth by stimulating the transition from a vegetative phase to a cyst or reproductive stage, as observed by Kobayashi et al. [23] and by Nimura and Mizuta [24] with Haematococcus lacustris (formerly H. pluvialis) (Chlorophyta) and Saccharina japonica (Phaeophyceae).
It has already been proven in literature such as Nagata [25], Hu [26], Ikeuchi [27], and Skoog [28] that a callus may be formed from a single differentiated cell, and many callus cells are totipotent, meaning they can regenerate the entire plant. In other cases, callus cells contribute to somatic embryogenesis, a procedure in which adult somatic cells are used to create embryos. This is frequently due to a high ratio of auxin to cytokinin or cytokinin to auxin, which drive root and shoot regeneration, respectively, whereas a transitory auxin to cytokinin ratio favors callus formation. Other hormones, including abscisic acid and brassinosteroids, can contribute to callus development and, in certain situations, replace auxin or cytokinin in the process. Auxin and cytokinin have been the most utilized and researched hormones for callus development, followed by organ regeneration [29].
As reported by Yokoya [30], Yokoya et al. [31], and Kumar [32], those auxins and cytokinins, such as Indole-3-acetic acid (IAA) and 6-Benzylaminopurine (BAP), can be used in combination to accelerate the callus growing from species of the Gracilariaceae family. Another hormone, Kinetin (KIN), was also observed to cause callus development in Gracilaria perplexa but had a detrimental impact on Gracilaria tenuistipitata (Rhodophyta). Yoon and Soh in 1998 [33] found that callus formation of the brown alga Silvetia siliquosa (formerly Pelvetia siliquosa) is induced by up to 90% of IAA supplement in solid PES medium; however, at higher doses, the rate of induction decreases. Sulistiani (2012) [34] noted a higher rate of Kappaphycus alvarezii (Rhodophyta) callus formation in PES media compared to CW media when supplemented with IAA. However, the optimal induction of callus in this seaweed occurred on solid PES medium containing 0.1–1 mg L−1 of 1-naphthaleneacetic acid (NAA) or 6-Benzylaminopurine (BAP) [34]. Furthermore, NAA, phenylacetic acid (PAA), or a mixture of NAA, PAA, and BAP produce an optimal callus growth of K. alvarezii [29].
To induce seaweed callus, according to Uji et al. [35] and Muhamad [36], α-lipoic acid (ALA), a growth promoter and retardant, can also be utilized. Tabuchi et al. [37] obtained the development of callus in all segments of Saccharina japonica (Phaeophyceae), at a concentration of 100 mg L−1 of ALA; nonetheless, at excessive concentrations, the growth of callus-like cells was suppressed. The two scientists referred to above also reported that supplementation with UNI, a triazole-type inhibitor, led to maximum callus induction among PGRs of the genus Sargassum (Phaeophyceae). Uji et al. (2016) [35] further demonstrated that this inhibitor could hinder cellulose production, leading to the formation of calluses. Apart from the aforementioned growth regulators, evidence suggests that picloram (PIC), indole-3-butyric acid (IBA), 2-isopentenyladenine (2iP), zeatin (ZEA), gibberellic acid (GA), and N-(2-chloro-4-pyridyl)-N’-phenylurea (CPPU) are also effective in regulating growth in macroalgae callus cultures [29,35].
Another study on the Ulva genome ended up providing more information about the hormonal role in development, with the existence of genes responsible for the biosynthesis of abscisic acid (ABA), ethylene, salicylic acid, and auxin (IAA). While culture studies have indicated the presence of Gibberellic acid (GA3) production in axenic Ulva sp., no precursor of GA3, such as ent-kaurene (CPP synthase, ent-kaurene synthase), has been identified in the Ulva sp. genome. Additionally, the Ulva genome lacks homologs of angiosperm hormone sensing and signaling genes, suggesting the existence of a distinct and independent hormonal signaling mechanism in Ulva sp., which may bear similarities to those observed in microalgae, such as diatoms (Bacillariophyceae) [4].
Sporulation Inhibitors
In the various works carried out by scientists on different species of Ulva, two sporulation inhibitors were identified (inhibitor-1a (SI-1a) and inhibitor-2 (SI-2)) and a compound as a swarming inhibitor (SWI), acting on the joint in the process of inducing reproduction and releasing gametes or zoospores, regardless of photoperiod control.
Stratmann et al. [15] outlined that vegetative thalli secrete a high molecular mass cell wall glycoprotein (SI-1) into the surrounding medium while simultaneously harboring a second low molecular weight inhibitor or non-protein molecule (SI-2) within the space between the two cellular layers of the thallus. The transition of a lamina cell into a gametangium only occurs when SI-1 levels diminish and the continually present SI-2 is no longer detected by the algae. SWI serves as a mechanism to coordinate gamete release during the swarming phase, with a decline in concentration observed on the third day post-induction, further amplified by an additional light stimulus, thereby enhancing the likelihood of mating [13,15,38]. Cells are no longer susceptible to the action of sporulation inhibitors after 23–26 h (when cells enter the determination phase) [13,39].
Unmated gametes can develop parthenogenetically into haploid clonal gametophytes, ideal for genetic manipulation and reproducible standardized experiments. It was reported according to Løvlie [40] and Stratmann et al. [15] that the generation time of Ulva compressa (as U. mutabilis) is short: only 3 to 5 weeks of growth are required between the potential inducibility of gametogenesis.
Stratmann et al. [15], utilizing Ulva compressa (formerly known as U. mutabilis), along with Wichard and Oertel [38] and Vesty et al. [41], who studied U. lactuca, U. linza, and U. rigida, observed that U. linza generates similar self-inhibitors (SI) as U. compressa during gametogenesis. Both types of SI from U. compressa (SIM1 and SIM2) demonstrated the ability to impede gamete production in U. linza and U. compressa, albeit with a somewhat lesser effect (particularly for medium-derived SIM1 and SIM2 between cell layers) in U. linza [15,38,41]. On the other hand, U. linza SI (SIL1 and SIL2) were able to inhibit gamete formation in both Ulva species tested. They concluded that these SI use analogous signals regulate the induction of reproduction in different species and that life cycle regulators are not specific to the species but rather to the clade in Ulva. Thus, to increase the potential of Ulva sp. as a model organism, gametogenesis can be artificially induced by removing both IS through cutting the thallus into single-layer fragments and subsequent washing, as exemplified in the studies above [4,38,41].
Algae Microbial Interaction
We learn in ecology that in each ecosystem there are different forms of interactions between the beings that constitute it and determine everything from obtaining food, shelter, and protection; reproduction; and survival. These interactions range from cooperative ones (mutualism, commensalism, tenants) to disharmonious ones (parasitism, competition, predation, etc.). In this way, marine bacteria have developed several strategies to survive. According to existing literature such as Dang and Lovell 2016 [42], Egan et al., 2013 [43], and Hardoim et al., 2012 [44], they can have a free-living planktonic lifestyle or can be organized in biofilms on rocks, particles, micro and macroalgae, sponges, and animals. Azam and Malfatti [45] state that bacterial interactions with primary producers can be mutualistic, commensal, or pathogenic and can alternate between different forms depending on the partner’s life stage [39].
Marshall et al. [46] identified roughly 38 distinct bacteria from U. linza and classified them based on morphogenetic activity during 28 days of incubation. Four types were determined based on the number of tubular extensions formed from a core callus. One category relates to axenic civilizations and reflects a morphotype that is extremely similar to the axenic morphotype found in this study. However, none of the other categories characterized the complete return of morphogenesis, and a combination of the isolated bacteria was not examined [41,47].
Burke et al. [48] found that the algal microbiome associated with Ulva australis undergoes seasonal fluctuations and varies between closely located sample sites. While they did not extensively confirm the mono-specificity of their Ulva samples, their findings led them to conclude that Ulva lacks a consistent core microbial community. Instead, they suggested that the composition of epi-bacteria on Ulva is influenced by random selection, akin to a “lottery”, rather than being controlled by specific mechanistic interactions, such as mutualistic relationships with the green algae [47,48].
Alsufyani et al., 2014 [49] conducted the first analysis of the full microbiome of U. bulbosa for AGMPF-producing bacteria and compared it to the microbiome of U. compressa, collected in the Mediterranean Sea (Ria Formosa, Portugal), where U. mutabilis was originally found by Føyn 1958 [50]. The microbiomes differed significantly from each other; while the Mediterranean species was associated with α-Proteobacteria and Bacteroidites, the microbiome of Ulva hookeriana (formerly U. bulbosa) was dominated by γ-Proteobacteria and Bacteroidites. The quantity of Operational Taxonomic Units (OTUs) identified in cultivated U. bulbosa (41) markedly contrasts with the higher count (251) found in uncultivated U. compressa. This discrepancy reflects a common trend observed in algal-bacteria interactions, as noted previously by Califano et al., 2020 [51] and van der Loos et al., 2021 [51,52]. It is important to highlight that the genera Maribacter and Sulfitobacter, essential producers of AGMPF, were associated with both species [53].
Briefly, in the relevant literature, around 1500 different bacterial strains associated with Ulva have been reported as α-Proteobacteria or γ-Proteobacteria, and several of them have been considered responsible for morphogenesis [54].
In Duan et al., 1995 [55], Nakanishi et al., 1999 [56], Matsuo et al., 2003 [57], and Marshal et al., 2006 [46], it was reported that morphogenesis in macrophytic green algae from the families Ulvaceae and Monostromaceae is controlled by bacteria belonging to the genera Cytophaga, Pseudomonas, Staphylococcus, Vibrio, Bacillus, and Flavobacterium [58].
Provasoli and Pintner 1980 [59], referenced by Fjeld 1972 [60], and Wichard et al., 2015 [47] discovered that the distinctive foliose morphology of Ulva was lost when grown under axenic conditions in a specified synthetic medium, but the introduction of bacteria restored differentiation. It should be emphasized that the propensity was observed prior to germ cell formation, alluding to the genetic regulation of cell differentiation in the natural bleb-like morphotype of Ulva compressa (as U. mutabilis). Wichard et al., 2015 [47] also noted that although bacteria cannot pass endogenously to the next generation, direct contact between bacteria and seaweed or a message through secreted chemicals is essential for growth, development, and differentiation. In Matsuo et al., 2003 [57], the discovery was reported that the substance that induces Thallus morphogenesis is secreted by Cytophaga sp. (YM2-23) of the Cytophaga—Flavobacterium—Bacteroide complex. Therefore, we can say that these organisms are also responsible for the release of spores, recruitment, and initial establishment on an appropriate substrate [13].
Wheeler et al., 2006 [61], when working with a specimen of Ulva intestinalis, noted that in addition to development and morphogenesis, bacteria-derived “acyl-homoserine lactones” also play a vital role in modulating the rate of settlement of zoological spores through a chemokinetic mechanism [13].
Singh et al. [58] carried out a study stating that Marinomonas sp. and Bacillus spp. induced wild-type morphology and growth in U. lactuca (formerly Ulva fasciata) (it was the first study to demonstrate involvement in this species). Nakanishi et al. (1996) [62] also reported the involvement of Bacillus spp. affecting morphology and growth in Ulva australis (formerly U. pertusa). However, the effect of bacteria on the growth rate of macroalgae has not been well quantified [58].
Spoerner et al. (2012) [63] conducted axenic cultivation of Ulva and observed that complete morphogenesis was achieved through the combination of two bacterial strains, Roseobacter sp. and Cytophaga sp., or via morphogenetic complexes obtained from the bacterial supernatant. These bacteria perform complementary functions: Roseobacter sp. triggers cell division akin to the plant hormone cytokinin, while Cytophaga sp. fosters the development of a viable undifferentiated basal cell and primary rhizoid cells, akin to the plant hormone auxin, in addition to promoting proper cell wall formation. Notably, the Roseobacter species displays a specific chemotactic attraction to the rhizoid cells of Ulva compressa (formerly U. mutabilis) and appears to engage in chemical communication with both the Cytophaga strain and the alga, forming a distinctive tripartite symbiotic community [47,54,63]. Notably, while other α-proteobacteria (such as Sulfitobacter sp.) and γ-proteobacteria (like Halomonas sp.) can substitute for Roseobacter, the presence of Cytophaga sp. seems to be indispensable. This implies that certain genes might play a crucial role in shaping the Ulva community and its accompanying bacteria [47]. Furthermore, laboratory studies have demonstrated that different bacterial communities with similar functional attributes can facilitate successful algal morphogenesis in Ulva compressa (as U. mutabilis) and U. intestinalis as reported in Ghaderiardakani et al., 2017 [64].
Kessler et al., 2017 [39] studied the metabolic profile of fragmentation-induced reproduction in U. prolifera, where they reported a dynamic change within 48 h. The metabolites, namely γ-aminobutyric acid (GABA), glutamic acid, gallic acid, and malic acid, were responsible for the formation and release of reproductive cells [39]. Surface metabolomic snapshots were employed to monitor the gametogenesis of Ulva compressa (formerly known as U. mutabilis), during which lamina cells undergo synchronous differentiation into gametangia, leading to the production of gametes. Molecular ion imaging of the metabolites unveiled dynamic alterations in surface-associated compounds. An extensive dataset comprising spatially and temporally resolved compounds, including DMSP and 55 specific molecular biomarkers, facilitated the distinction of four distinct time points in the gametogenesis process [13,39].
Kessler et al. [39] explored whether the algae-bacteria relationship arose due to the mutual exploitation of common environmental signals. They subjected a sample of Ulva to axenic conditions, leading to the development of callus-like structures resembling pincushion morphotypes. These structures were characterized by unusual cell wall formations, a lack of cell differentiation, and slow growth. Upon the introduction of two bacterial strains—Roseovarius sp. and Maribacter sp.—the morphogenesis of Ulva compressa (formerly U. mutabilis) was fully restored, resulting in the formation of a tripartite community, as previously described in studies by Grueneberg et al., 2016 [65] and Spoerner et al., 2012 [39,63,65]. DMSP was then identified as an attractant and tracked from algal production to bacterial uptake, i.e., Ulva releases significant amounts of DMSP into its chemosphere. Roseovarius sp. are attracted to DMSP and absorb this metabolite quickly. Although they do not directly stimulate bacterial growth, once a specific threshold is reached, bacteria uptake and store more DMSP than Ulva releases de novo. This leads to bacterial biofilm expansion as Ulva provides a boundary layer of glycerol, serving as a carbon source for Roseovarius sp. Additionally, chemical communication can occur bidirectionally; bacteria can attract Ulva zoospores using N-acyl homoserine lactones, as reported by Joint et al., 2007 [66]. Upon initial interactions, morphogenetic compounds trigger cell division in Ulva (Roseovarius factor) and rhizoid formation (Maribacter factor). These morphogenetic compounds overall promote biomass production and facilitate the formation of retention structures, allowing algae to integrate directly into the bacterial biofilm [39,66].
We can therefore say, taking into account the various examples described above, that bacteria are essential and play a crucial role in the physiology of Ulva sp. from development and growth to the induction of reproduction, sporulation, and the settlement of its zoospores through various mechanisms, such as production of growth-promoting substances, quorum sensing, signaling mediated by bioactive compounds, etc. [54]. To this end, Ulva sp. needs to cultivate bacteria that create protective biofilms around rhizoid cells and provide morphogens for their growth (Figure 2). Furthermore, they also release various organic compounds into their environment to harm other organisms and to maintain the balance of osmotic pressure in the surrounding seawater [39]. Because bacteria can selectively utilize nutrients from algae, they play a key role in biotransformation and nutrient recycling [54]. However, on the other hand, we can also find epiphytes. Epiphytic individuals can be other unwanted species of algae, viruses, bacteria, and fungi. They can attract herbivores such as crabs, lobsters, shrimps, crayfish, fish, and turtles, which have positive (enriched biodiversity throughout the food chain, ecosystem services, etc.) and negative (loss of yield, etc.) impacts on seaweed cultivation [67]. According to Goecke et. al. 2010 [68], epiphytic bacteria also provide a defensive function through their antibiotic activity or by forcing pathogenic bacteria to compete for space within the biofilm [47,54].
Mutualistic Symbiotic Relationships
Another example was the case of the investigation in Veerse Meer, a brackish lagoon located in the southwest of the Netherlands carried out by Kamermans et al., 2002 [69] on the role of isopod and amphipod grazing in Ulva spp. They identified the amphipod Gammarus locusta and the isopods Idotea chelipes and Sphaeroma hookeri as potential herbivores of Ulva spp. However, in the presence of epiphytic diatoms on Ulva spp. thalli, Gammarus and Sphaeroma species primarily grazed on the diatoms rather than on the Ulva tissue. Only Idotea continued to graze on Ulva spp. In a laboratory experiment, it was found that Gammarus had a positive effect on Ulva growth, likely due to its selective removal of epiphytic diatoms from the thalli. Herbivores that remove epiphytes, such as Gammarus, may stimulate Ulva development, thereby explaining the higher growth rates observed in the field. When assessing the potential role of herbivorous invertebrates in reducing macroalgal biomass accumulation, it is crucial to consider epiphyte abundance on macroalgae. Preferential removal of epiphytes may promote growth and consequently have an opposite effect [4,69].
Ingle et al., 2018 [67], when working on a proposed framework for the integrated management of marine pests in seaweed cultivation, based on several cases studied, noted that a low density of Ulva rigida and Cladophora sp. (minimum 11.5 g) under closed in-door plastic photobioreactor culture led to culture collapse due to proliferation of epiphytes (other marine algae, nematodes, copepods, and bacteria), but a high inoculum density (minimum 179–264 g) yielded a 15% daily growth rate. Furthermore, the culture of Ulva sp. in a cage with the inclusion of a double safety net resulted in a daily growth rate of 8.1% per day in open sea agriculture in Israel in a one-month growth cycle, which in the absence of the safety net dropped to −2.5% per day [67].
Finally, it should be noted that the composition of the microbiome associated with Ulva sp. changes depending on geographic location, seasonality, and abiotic factors such as temperature, salinity, depth, light, etc. Therefore, understanding this microbiome-Ulva complex is essential given its importance for the adaptation of Ulva spp. to their environment, which will vary between aquaculture systems [70].
Metabolomic Profile
Metabolites are compounds, generally organic, that participate in chemical reactions that occur at the cellular level. Classified as primary and secondary, they are differentiated by whether they are directly involved or not in the normal growth, development, and reproduction of an organism. Some authors like [71,72] consider that metabolites released into the environment “act as spoken language, transmitting signals from the genetic architecture and the environment.” Therefore, metabolomics can help provide a functional understanding of an organism’s physiological state or symbiotic interactions [73].
A study was carried out to investigate the metabolic profiles of U. prolifera during proliferation induced by fragmentation. Filaments and fragments were collected at intervals of 12, 24, 36, 48, 60, 72, 96, and 120 h and subjected to metabolic profile analysis using GC-MS. Within 48 h, the procedure detected a dynamic shift in the metabolic profile. A total of 156 metabolites were measured, with 63 showing substantial change. GABA, glutamic acid, gallic acid, and malic acid are the metabolites responsible for the creation and release of reproductive cells. Furthermore, n-hexanol, 2-methyl-3-phenylindole, and 3-indoleacetonitrile were responsible for stress reduction. Changes in the levels of metabolites including sugars, organic acids, and alcohol with photoperiod may be the strategy adopted by U. prolifera to deal with fragmentation in nature [74]. According to Kessler et al. [39], matrix-assisted laser desorption/ionization mass spectrometric imaging (MALDI-MSI) directly enabled the spatial distribution of chemical molecules during cell differentiation processes. Surface metabolomic snapshots were used to monitor Ulva compressa (as U. mutabilis) gametogenesis, which occurs when lamina cells develop into gametangia and produce gametes at the same time. Molecular ion imaging of metabolites revealed dynamic changes in surface-associated chemicals. A vast collection of geographically and temporally resolved chemicals (DMSP and 55 unique molecular biomarkers) helped to separate four discrete time periods in the gametogenesis process. Although the identification of effective biomarkers is the first step, the isolation and elucidation of the structure of these compounds still require further work [13,39].
In a separate investigation focusing on the cellular metabolic responses of a temperate species and a cold-adapted species, cultures of Ulva compressa (formerly known as U. mutabilis) from Ria Formosa, Portugal, and U. bulbosa from the polar region of Antarctica were exposed to a simultaneous temperature shift from 18 to 5 °C in the initial experiment. Following a 14-day incubation period, Ulva biomass was harvested, extracted, and analyzed using HILIC-HR-ESI-MS. The results revealed significant differences in 18 out of 30 metabolites. The researchers discovered that ectoine was the sole component not detected in axenic cultures of Ulva compressa (as U. mutabilis) at 18 °C or 5 °C, indicating that it is generated only by bacteria associated with the holobiont. There was an increase in the production of taurine (a sulfur-containing β-amino acid, potentially an important carbon source for heterotrophic bacterial growth) in U. bulbosa but not in Ulva compressa (as U. mutabilis), emphasizing the difference in the responses of these two species. Ulva to heat stress; cysteinolic acid (a member of the marine sulfonate class, it has physiological antioxidant activity and osmoregulatory effects) was elevated in Ulva compressa (as U. mutabilis) (9.5 times) after the temperature change to cold, but the concentration was below the limit of quantification. The proline concentration increased sharply by 24.9% after the temperature change, while the methionine concentration did not change. Similarly, Lee and Liu 1999 [75] noted that proline accumulated (to 6.48 mg g−1 DW, a 9.8-fold increase) in U. lactuca (as U. fasciata) exposed to high salinity. Dawson et al. (2020) [76] observed a significant increase in proline levels, reaching a four-fold elevation, in response to decreasing temperatures (−1 °C compared to 4 °C) in the polar diatom, Nitzschia lecointei (Bacillariophyceae) [76].
Similar to the 4.5-fold increase observed in proline levels in Fragilariopsis cylindrus (Bacillariophyceae), a prevalent psychrophilic diatom found encased in freshly formed sea ice and subjected to heightened external salinities and lower temperatures, this change reflects alterations seen under cold stress conditions. According to Gawryluk et al., 2019 [77] and Qiu et al., 2020 [78], the concentration of glutamine, a proxy for its precursor glutamate, increased by 20.6-fold in Ulva compressa (formerly U. mutabilis) under similar conditions. Glutamate serves as a precursor to various metabolites, including proline, arginine, and histidine, and plays a role in numerous biochemical reactions [77,78], including those to pathogen resistance and abiotic stress (such as cold, heat, and drought). Glycine betaine, which rose 13.4-fold following cold stress, is commonly regarded as a potent protectant against the harmful effects of stressful situations on plants. Finally, the level of production of dimethylsulfoniopropionate (DMSP) was higher in Ulva compressa (as U. mutabilis), while Van Alstyne and Puglisi 2007 [79] found a level lower than expected for species from cold regions. In summary, hot–cold temperature changes affect various morphological, cytological, physiological, and biochemical characteristics in plants. Plants acclimated to harsh ecosystems, such as polar regions, must maintain strategies to deal with severe stresses [53].
2.2.2. Abiotic Factors
Abiotic variables are inanimate components of ecosystems that have an impact on both creatures and the surrounding ecosystem. These elements have a significant impact on ecosystems’ functioning and structure. Changes in abiotic variables can have a significant impact on the abundance, distribution, quantity, and behavior of organisms in an ecological system. These characteristics are used to learn more about how ecosystems work and how they adapt to natural changes or human-caused disruptions, from an ecological and biotechnological point of view. The abiotic factors can be physical or chemical, and they can affect the ecosystem’s status quo as well as modify the organism’s metabolism to a survival mode or induce distress to the organisms.
Salinity
Salinity can influence spore release by affecting turgor pressure and pore diameter of sporangia [80]. According to Sousa et al., 2007 [81], for Ulva intestinalis species, spore biomass was strongly affected by salinity, having been limited to 5 psu and then favored at 20 PSU and highly increased at 35 PSU. They also demonstrated that spore biomass tends to increase with increasing salinity and increasing light intensity [81]. Han et al., in 2008 [82], also found a maximum spore release for Ulva australis (as U. pertusa) of 25 to 35 PSU.
In a study by Carl et al. (2014) [83], Ulva filaments were subjected to dechlorinated tap water for 10 min at reduced salinity. Following this, all filaments were dried using paper towels to eliminate excess water and then dehydrated by exposure to air for 45 min in darkness at 25 °C (dehydration treatments), or they were kept in a dark environment for 45 min as part of the salinity control [83]. Third, the effect of segmentation was tested by cutting a section of approximately 50–70 mm from each filament. The authors found that there was no effect of salinity shock on the release of gametes after two and three days [83].
Light Intensity
The release of reproductive cells in Ulva is reliant on photosynthesis, making light a crucial factor in its spawning process. There exists a positive correlation between the presence of light in the water column and spore biomass.
Mantri et al. (2020) [4] have reported a linear association between growth rate and light intensity, up to 30 μmol photons m−2 s−1. However, growth reaches saturation levels beyond 70 μmol photons m−2 s−1 [4]. Furthermore, it can be noted that ultraviolet (UV) radiation has a negative effect on growth. Mitigation of UV exposure is achieved by the accumulation of UV-absorbing compounds such as carotenoids, coumarins, phenolic compounds, and mycosporin-like amino acids (MAAs).
Sousa et al., 2007 [81] found that algal spore biomass increased with the increasing amount of available light, where the higher tested photon flux density (PFD) (90 μE m−2s−1) allowed significantly greater algae recruitment when compared to the other two PFDs tested (20 and 40 μE m−2s−1) [84].
Another point is the types of endogenous rhythms: ultradian (with periods of several hours or much less), circadian (with periods between 20 and 28 h), and infradian (with periods longer than 28 h).
There is evidence that the brief infradian clock controls physiological processes in animals; for example, bivalves, rats, ducks, horses, and humans all have infra-radian rhythms with durations ranging from 2 to 7 days. However, information on these infradian rhythms in marine algae is scarce, and most reports concern reproduction. For example, in Fucus vesiculosus (Phaeophyceae) there is a semilunar rhythmicity, and there are reports of a weekly or 5-day reproductive rhythm that occurs in U. lactuca, U. pseudocurvata, and U. fenestrate, and an interval of 4 to 5 days between the formation of gametangia occurs in Derbesia [4,85]. However, the question arises whether such rhythms found in algae from different geographic areas will be the same.
Kalita and Titlyanov 2013 [85] carried out a study comprising two experiments. The first was to analyze the effects of temperature and irradiation on the infradian rhythm of growth rate, where they placed 48 discs with U. lactuca stalk in different combinations of temperature (5, 10, 15, 20 °С) and irradiance (40 or 60 μmol m−2s−1 photons) in 12 h of light per day. The second experiment was the analysis of the effects of photoperiod on the infradian rhythm of growth rate, where they placed 81 discs containing male and female gametophytes and sporophytes cultivated in two groups of 8 h light:16 h dark (8:16 L:D) and 12 h light:12 h dark (12:12 L:D).
They found that the growth rate, in terms of rate of increase in biomass, exhibited a rhythmic pattern with a major peak every 2 or 3 days, both at a temperature of 5 °C and photons of 40 μmol m−2s−1 and at a temperature of 10 °C and 60 μmol photons m−2s−1 [85].
Temperatures of 5 and 10 °С caused an increase in the prevalence of 3-day cycles, keeping U. lactuca in the vegetative growth stage, while fertile tissue appeared mainly at 10 °C and 60 μmol photons m−2s−1. Considering references that indicate that higher minimum light (10 µmol photons m−2s−1) is required for reproduction versus vegetative growth (5 µmol photons m−2s−1) for Ulva australis (as U. pertusa) [85].
Exposure to 15 and 20 °С caused a predominance of 2-day cycles and induced reproduction after 5 days of cultivation, representing 56–90% of all cycles detected. And finally, lengthening the photoperiod from 8 to 12 h altered the amplitude of the growth rate, so that growth accelerated as the light phase increased [85].
Temperature
Temperature affects photosynthesis resulting in altered productivity and therefore supplies carbohydrates to the tissue. Temperature response is exhibited through phenological attributes mediated by complex metabolic pathways, such as significantly affecting respiration rates via adenylate control, i.e., ADP/ATP ratios; increasing the content of nitrate reductase, amylase, invertase and peroxidase; and reducing the growth rate as the temperature increases [54].
Mantri et al. (2020) [4] identified the optimal effective temperature range for inducing reproduction in various Ulva spp. to be between 15–20 °C. However, they noted that a difference of 5 °C is crucial for effectively inducing gametes or spores. Additionally, a temperature difference of 10 °C is essential for the growth of tropical species [4]. There is the possibility of relating it to the seasons. For example, U. lactuca from Groton, USA, only reproduces in the warmest months at 21 °C and is inhibited at 25 °C, while populations of Ulva pseudocurvata from the North Sea have approximate weekly reproductive peaks during the summer and peak biweekly during the colder seasons [54].
Another example of the influence of temperature is due to the experiment by Kalita and Titlyanov 2011 [85], where they collected a stalk of Ulva fenestrata in Japan (temperature 6 °C and irradiation at 60 UE/(m2s)) and placed it in the laboratory (water temperature 15 °C and irradiation at 40 EU/(m2s)). They found that the reproductive period decreased from 30 to 5 days when the temperature increased from 10 to 20 °C and ceased at 5 °C [54].
With this, researchers investigated the relationship between rapid changes in temperature and the induction of gamete release over shorter periods of days and hours.
Niesenbaum (1988) [86] employed a temperature shock with 2 °C saltwater to induce sporulation in U. lactuca when no reproduction occurred between samples after 30 days. Plants were withdrawn from 22 °C containers and cleaned completely in filtered saltwater at 2 °C before being reintroduced to clean growth chambers with fresh media at 22 °C. This study involved two reproductive therapies along with a control group. The experiment was limited to these conditions because, within 18 h of washing at 2 °C, all samples had transitioned to a reproductive state, rendering no vegetative algae available. The author observed that the conversion of vegetative tissue to a reproductive condition occurred almost instantly after washing the algae in water at 2 °C, and the production of gametes over 12 h was rapid and consistent. Carl et al. [83], working with a tropical filamentous species of Ulva spp., subjected the stalks to a temperature shock (4 °C) for 10 min, followed by immersion in autoclaved, filtered seawater under a 12-h photoperiod at 25 °C. Gamete release reached its peak 2 days after the onset of treatment, resulting in an approximately 10% increase in sporulation [83]. In 2017, Gao et al. [87] demonstrated that a 20-min temperature shock at 4 °C induced an average reproductive response of 94.7% in the tissue of the lamina of Ulva rigida on the fifth day with a two-step process, the first being from high temperature to low temperature and the second the return to high temperature. The results demonstrated that none of the steps is dispensable, as continuous induction of low temperature for 3 weeks, without returning to high temperature, did not trigger reproduction [54].
Some authors have suggested that a minimum of 2 days is required for the transition from the vegetative to the reproductive state and that enhanced reproduction under abiotic temperature shock may be a survival strategy under unfavorable conditions such as occurs in certain animals.
Nutrients
The nutrients required for seaweed growth can be found in seawater; however, they are often concentrated in plant tissue and can be divided into three categories: micronutrients or trace elements (e.g., Fe, Zn, Cu, Mn, etc.); vitamins (B12, thiamine, and biotin); and macronutrients (e.g., N, P, C, etc.). Seaweed concentrates nitrogen and phosphorus about 100,000 times [54]. Nitrogen is a key cofactor in cellular anabolic activities that produce amino acids, including nicotinamide adenine dinucleotide phosphate (NADP+). Phosphorus has a similar role in nucleic acid production and promotes photosynthesis, nutrition transport, and energy transmission via energy-rich adenosine triphosphate (ATP) linkages [54].
The optimal ratio refers to the amount of nutrients necessary for maximum plant development. The typical ratio for seaweed is 30N:1P, ranging from 10:1 to 80:1. When an excessive amount of a nutrient is delivered, the nutrient becomes a “limiting factor.” Chemical variables, such as nutrient content and the type of the inorganic nutrient (e.g., non-ionized ammonium nitrate NO3− or ionized ammonia NH4+), can influence absorption rates [54].
Some marine algae can absorb NO3− and NH4+ simultaneously and in equal proportions, but most prefer to assimilate NH4+, as is the case with U. lactuca. When nitrate is ingested, it can be retained intracellularly in the vacuole and cytoplasm or converted to nitrite by the enzyme nitrate reductase. Nitrite is transferred from the cytoplasm to the chloroplasts and converted to ammonia by the enzyme urease. Ammonium is taken up or transformed into amino acids by glutamine synthetase in chloroplasts [54].
Pedersen [88] observed three phases of NH4+ uptake (first observed by Conway et al. [89]) in U. lactuca. These phases are epidemics that are managed both within and outside. When the absorption rate was several times higher than the growth rate, they referred to it as a “surge”. This increased absorption rate enables the cell to “catch up” on its earlier nutritional shortage by absorbing the limiting material much quicker than required for development. For these reasons, it is critical to acclimate wild samples in order to homogenize any differences in nutrient absorption or needs. It should be noted that some nutrient-limited algae exhibit a decline in nutrient absorption over time after being introduced to saltwater. As a result, while performing research, it is critical to document the duration of nutrient measurements. It has been shown for Ulva spp. that NH4+ absorption rates decrease by 50% in the dark compared to light [54].
Sousa et al. [81] intended to evaluate the influence of light and phosphorus (PO4–P) on the germination and growth of Ulva intestinalis spores; three light intensity levels (20, 40, and 90 μE m−2s−1) and three PO4–P levels (6.4, 2, and 0.8 μmol L−1) were combined and three levels of NH4–N (102.4, 32, and 12.8 μmol L−1). They noted that spore biomass increased with light and PO4–P concentration and the interaction between these factors was slightly statistically significant; that is, at a PO4–P concentration of 6.4 μmol L−1, spore biomasses increased when subjected to 40 and 90 μEm−2s−1 [81]. Regarding NH4–N, the spore biomass was inversely proportional to the concentration: the biomass of algal spores tended to decrease with increasing NH4–N concentrations. The highest salinity tested (35 PSU) together with the lowest concentration of NH4–N allowed a greater spore biomass and, consequently, a higher growth rate [81].
Gao et al. in 2017 [87], when working with eight thalli (five wild type and three sterile mutants) of U. rigida collected in the intertidal zone of Cullercoats Bay, United Kingdom, noticed that the growth rate of the wild thalli decreased with time of cultivation and it was close to zero on day 12, suggesting that biomass would not increase after day 12. This growth was possibly due, in addition to sterility, to the absorption of nutrients, where they observed that the absorption of nitrate and phosphorus by sterile stalks was 40% and 30.9% greater than the wild ones, respectively.
Temperature plays a critical role in regulating various aspects of seaweed physiology, including enzyme activity, the pace of chemical reactions, and the efficiency of nutrient transport across boundary layers. Temperature can influence these processes by impacting the activity of membrane transporters, particularly for nutrients absorbed through active transport, although its effect on passive diffusion absorption may be relatively smaller [54].
Elevated temperatures have been observed to accelerate growth and reproduction in seaweeds by enhancing enzyme activity. Additionally, higher nitrogen availability can expedite the synthesis of nucleotides and proteins, facilitating rapid growth and reproduction [90].
Thallus Fragmentation
Thallus fragmentation is thought to play an essential role in the fast induction of this change and reproduction. The method of generating fragmentation was once thought to be the cutting of the damaged cell walls and the additional structure of the cell matrix, allowing the algal segments to filter out the inhibitors and thereby reducing the regulatory hurdles to reproduction. Grinding the thallus into monolayer pieces and washing away sporulation inhibitors artificially promote gametogenesis in mature laminae [74].
According to authors like Hiraoka and Enomoto in 1998 [91], fragmentation significantly increases the sporulation rate of Ulva compressa (as U. mutabilis) from 15.8% to 80.0%. Additionally, within 2–3 days after fragmentation, the reproduction of Ulva australis (as U. pertusa) was induced [74]. Gao et al. (2010) [92] discovered that nearly all fragments of U. australis with a diameter of 0.5 mm were transformed into sporangia, while larger fragments only formed sporangia from marginal and submarginal cells [74]. He et al. (2019) [74] observed that nearly 90% of reproductive cells appeared within 48 h after cutting the U. prolifera thallus into segments of approximately 0.5 mm. Moreover, almost all cells (96.29%) in the physiological state transformed into the reproductive state within 120 h in the control group. This suggests that fragmentation induction was not the primary determinant of U. prolifera sporangia formation but rather acted as a promoter [74,91,92].
3. Ulva Cultivation Methods: Towards an Innovative Future?
Algae can be cultivated depending on the species, location, and cultivation facilities. Initially we choose our seaweed cultivation method based on the life cycle. They can be divided into different groups according to different criteria: based on manipulating or cultivation methods, cultivation systems, and several cultures [93].
Seaweeds can be categorized into two groups based on the manipulation of their life cycles: (1) those initiated from microscopic spores, where the entire seaweed life cycle is fully controlled during cultivation, and (2) those initiated from macroscopic seaweed fragments, where only a portion of the life cycle is controlled during cultivation [93]. Ulva sp. exhibits various reproductive modes, such as asexual reproduction, sexual reproduction, parthenogenesis, and vegetative propagation, which will be explored further [93,94].
Cultivation systems can be carried out offshore (the open sea method, which is defined as a marine product farm located at a certain minimum distance from the coast); onshore (the land method, where the cultivator can continuously monitor growth indicators; it is devoid of environmental threats such as storms, food, and other natural disasters); or nearshore (which is the best-known and most widely used seaweed aquaculture technique, which can be developed in estuarine and near-coastal locations). Photobioreactors are devices designed for the massive cultivation of microalgae; IMTA (integrated multitrophic aquaculture) is characterized by creating species of different trophic levels close to each other; saline aquaculture is terrestrial aquaculture that uses saline groundwater; and recirculating aquaculture systems are designed to control all environmental aspects of production by continuously filtering, treating, and reusing water, thus increasing operational efficiency and reducing risks from plant protection products and climate change [6,95,96,97].
Finally, based on the number of cultivated species, the cultivation methods can be di-vided into monoculture and polyculture (or integrated culture). Open-water cultivation and land-based farming can be distinguished by cultivation locations. In general, both entire life cycle and partial life cycle approaches, as well as monoculture and polyculture, can be used in open-water and land-based production [93].
3.1. Vegetative Propagation Method
Asexual reproduction is classified as parthenogenetic duplication and vegetative re-production. Parthenogenesis is the process by which biflagellate gametes evolve into adult algae by the formation of spores, and it is one of the primary causes of the fast reproduction and large biomass of green tides [94]. Liu et al. (2015) [98] discovered that the gametophytes of U. prolifera exhibit two reproductive outcomes: they can generate tetraflagellate spores or persist in producing biflagellate gametes. Additionally, Zhang et al. (2011) [99] noted that gametes can endure for more than 6 weeks and demonstrate positive phototaxis, aiding their aggregation on the sea surface and attachment to algae [98,99].
Vegetative proliferation is the most prevalent way of culture. This is a type of clonal propagation that is also used in plants that do not require life cycle control. According to Góes and Reis [100], the asexual structure or portion of seaweed thallus is used as initial “seed material” or “parent”, and new fronds will be genetically identical to the initial “parent”, which is useful for consistency in production [13,101]. Zhang et al. [102] investigated the asexual reproduction of U. prolifera and identified four vegetative reproduction modes, which include algal fragment regeneration, algal tissue regeneration, single-cell regeneration, and protoplast regeneration [94,102]. This method increases reproductive diversity enormously and enables it to colonize new territory more quickly and efficiently [103].
3.2. Life Cycle Method
The life cycle method is characterized by the gametophyte phase and must be induced (Figure 3). Gametogenesis can be artificially induced by removing sporulation inhibitors by cutting the thallus into single-layer fragments and subsequently washing (exemplified by Stratmann et al., 1996 [15]; Wichard and Oertel 2010 [38]; Vesty et al., 2015 [41] for Ulva compressa (as U. mutabilis), U. lactuca, U. linza, and U. rigida [47]).
The process of induction of gametogenesis can be divided into three phases (Figure 4): the determination phase (~0–26 h; the cells enter the next S-phase and become irreversibly committed to gametangium differentiation [14,38]; i.e., some cells in the segments gradually enlarge in size and became sphere-shaped, making the surface of the segments irregular [103]), the differentiation phase (~26–70 h, characterized by a reorientation of the chloroplast, followed by four consecutive cell divisions forming sixteen pro-gametes per cell and their maturation [14,38]; i.e., brown spots of gametangium were formed [103]), and the swarming phase (~72 h onwards—the mature gametes are eventually released if the SWI declines in concentration on the third day after induction, following a further light stimulus) [14,38].
There are two types of gametes: “male” and “female” gametes, both with red eye spots and an elliptical shape; the female ones are slightly larger than the male ones [103]. Unmated gametes can grow parthenogenetically into haploid clonal gametophytes, making them excellent for genetic manipulation and replication in standardized research. Løvlie 1964 [40] and Stratmann et al., 1996 [15] noted that only 3 to 5 weeks of growth are necessary between the potential inducibility of synchronous gametogenesis in Ulva compressa (as U. mutabilis) [47].
Ulva’s early development comprises five stages that a seedling must undergo to progress from a swarmer (either gamete or zoid) to a juvenile thallus:
- (I)
- Connected, germinating cell
- (II)
- Germinating cell with a discernible germination tube
- (III)
- Initiation of distal cell division
- (IV)
- Onset of lateral cell division
- (V)
- Juvenile thallus displaying evident expansion resulting from distal and lateral cell divisions [105]
3.3. In Vitro
In vitro is the expression used to study all biological process that happens in all organisms in a controlled and closed environment (inside of “a glass”). According to Morales-Rubio [106], two of the most frequent ways for plant tissue culture are by applying organogenesis, which refers to the creation of plant organs (roots or shoots) and can be performed in vitro, an can be done in seaweeds. Plants can be propagated either directly from meristems or indirectly from dedifferentiated cells known as callus. These cultures can be utilized for mass plant production through techniques like micropropagation or for the development of specific organs, such as roots in hairy root culture. Callogenesis results in an amorphous mass of cells when explants are exposed to various growth regulators [107].
All plant tissue culture methods involve a sequence of steps:
- (1)
- The plant of interest must be chosen; this is normally determined by the purpose of the study, although disease- and insect-free plants are desired; if the plant demands it, pre-treatments can be used.
- (2)
- The start of the in vitro culture necessitates the removal of tiny plant parts (explants) or the use of seeds with their surfaces sterilized.
- (3)
- The explants are then put in appropriate culture conditions and cultured for a short amount of time, with infected explants destroyed. The stages that follow differ based on the intended culture.
- (4)
- Organogenesis is the propagation phase in which explants are cultivated on suitable culture conditions for shoot or root multiplication, while in callogenesis, the callus is multiplied.
- (5)
- Callus and root cultures are grown in bioreactors, while propagated shoots are moved to root-promoting culture medium in the case of micropropagation.
- (6)
- Micropropagated plants are toughened in order to produce individual photosynthesis-capable plants. Hardening is done gradually, allowing the plants to adjust to ex vitro environments [107].
Micropropagation is a possible cultivation technique for clonal multiplication, and it has been successfully used for tissue culture of Ulva sp. [13].
The second methodology is through axenic cultures. These are an essential tool, for example, (i) to study the chemical ecology of macroalgal-bacteria interactions, (ii) to identify (allelopathic) compounds released by the macroalgae, (iii) to research the effects of bacterial morphogenetic compounds happening in the algal growth, and (iv) to have a controlled feedstock for algal aquacultures [108]. Vesty et al., 2015 [41] prepared the axenic cultures in U. linza by phototactic migration of freshly released gametes down a thin horizontal capillary. Under strictly sterile conditions within a laminar flow hood, movement towards a light source was induced. Sterile Pasteur pipettes equipped with 15 cm capillaries were assembled. Gametes exhibited upward movement within the pipettes, congregating at the top, where they were collected. Subsequently, they were transferred to another Pasteur pipette for further purification in the subsequent run [41]. Wichard 2015 includes a table with studies with Ulva sp., including the methodology to prepare cultures [109].
The third methodology is using a photobioreactor, which is the most recent method and is not yet fully developed. It may be limited to the manufacture of high-value compounds for pharmaceutical or nutraceutical purposes since it necessitates specialized infrastructure and expertise, which are costly. A study conducted by Chemodanov et al., 2017 [110] studied Ulva compressa and Ulva rigida, which were grown for 6 months in a macroalgae photobioreactor system incorporated inside a building. U. compressa grew at an average rate of 2.89–3.65% each day, but U. rigida did not grow well [13].
3.4. Cultivation Control or Monitoring Techniques
As previously said, any seaweed nursery or culture system requires strategies to be effective. Controlling seawater media (seawater and nutrients), temperature, and light is crucial, but so is monitoring the stage of growth.
Regarding light intensity, in the laboratory, fluorescent lamps can provide energy. Daytime lamps have the highest colour temperature, 5000 K or higher, and are designed to mimic natural sunlight; cool white bulbs emit light primarily in the blue spectrum, typically with a colour temperature of around 4000 K. On the other hand, warm white bulbs emit light in the red portion of the spectrum, with a lower colour temperature of 3000 K or less. Quantum light meters detect the quantity of light accessible for algae growth and the photosynthetic photon flux density, which is expressed in micromoles of photons per square meter per second (μmol m2 s1) [101].
Water quality requires a refractometer (a portable instrument for measuring salinity) and a pH meter. These must be calibrated using buffer solutions before each use, washed before and after each use in deionized water, and kept in an electrode storage solution [101]. To facilitate the work by having the possibility of in situ analysis and obtaining data in real-time, both in the field and the laboratory, multiparametric probes can be used. These have different types of sensors that measure temperature, pH, turbidity, dissolved oxygen in the water, electrical conductivity (EC), total solids dissolved (TDS), and salinity.
Regarding observation, a microscope is essential to determine successful spore release, spore density, and development of microscopic stages [101]. A magnifying glass can also be used to monitor the growth of inoculum. In more advanced locations, a high-powered dissecting microscope is also very useful for monitoring larger plants throughout the culture period or for isolating spores [101].
A spectrophotometer is a spectrophotometric instrument that measures the intensity of light in relation to wavelength. It is used to detect the concentration of substances; elucidate the structure of organic compounds; identify impurities, the characteristics of a protein, classes of compounds, and dissolved oxygen content and for functional group detection and determination of the molecular weight of a specific compound. There are several types depending on the type of light used: visible light, UV/Visible, near infrared, nuclear magnetic resonance (NMR), atomic absorption, mercury, and fluorometers [111,112].
3.5. Cultivation Security and Safety Measures
Macroalgae have a long history of usage in the creation of thickening and gelling agents used as additives in foods, cosmetics, and pharmaceutical items; therefore, in the EU, every product that is sold must adhere to particular regulations. In the food sector, there are three areas that are linked to algae, from its production through direct to indirect consumption. Regarding their production, organic macroalgae create a specific regulatory category, and Regulations EC No. 710/2009 and EU No. 2018/848 established detailed rules on the biological production of macroalgae including harvesting from natural reserves (i.e., the collection does not substantially affect the strength of the natural ecosystem). For algae and extracts to be commercialized, three Novel Food Regulations must be followed: EC No. 258/97 (which establishes that species that have not been used as food to a significant degree in any of the EU member countries before 15 May 1997 need to undergo an authorization procedure); EC No. 2015/2283 (where an additional notification system is provided for species that have demonstrated a history of safe food use for at least 25 years in a country outside the EU), and EU No. 2017/2470 (or Catalog of Novel Foods, which contains a list of foods including European and imported seaweed intended for human consumption) [113].
As for indirect consumption (such as food additives and supplements), it is controlled by EC regulations No. 1333/2008 (which contains a list of authorized food additives and comprises eight additives derived from macroalgae under codes E401-E407a); EU No. 231/2012 (which contains details of the origin, composition, and use of accepted additives); EU No. 1379/2013 (which is necessary to indicate the origin and method of production or harvest, establishing the common organization of fishing and aquaculture markets); EC No. 1924/2006 (where EFSA assesses the scientific evidence verifying nutritional allegations, e.g., “rich in fiber,” “source of vitamin/mineral x,” or “contains vitamin/mineral x”); and Annex XIII of EU No. 1169/2011 (nutritional information regulation for vitamins and minerals) [113].
Because algae have the propensity to collect toxins, we can propose the following regulations: CE No. 1881/2006 (establishes maximum levels for certain toxic substances, such as heavy metals, for certain foods); CE No. 396/2005 (relating to maximum limits for pesticide residues in food and animal feed, being 0.01 mg/kg of mercury); CE No. 1881/2006 (constitutes a market barrier for most food products derived from macroalgae); and EU Recommendation No. 2018/464 (for monitoring the levels of cadmium, lead, mercury, arsenic, and iodine in foods for human and animal consumption). However, there are still gaps in the regulations in terms of maximum levels and whether the levels are for dry or fresh weight.
In pharmacology, medications are authorized through one of four procedures: the centralized method, wherein the European Medicines Agency evaluates evidence and the EU Commission issues the license; the national procedure; the mutual recognition procedure; or the decentralized procedure. In addition to the “normal” approval procedure for synthetic medicines, Directive 2011/83/EC establishes distinct procedures for (i) herbal medicines and (ii) traditional herbal medicines. Algae ingredients and preparations are considered herbal remedies [113].
There are international standards that can be met not only in Europe but also around the world. The ISO 22000 standard (Food Safety Management System) is based on the internationally recognized Codex Alimentarius HACCP principles. It proposes to the market and consumers that there is a management system capable of providing safe products from suppliers to the market, including establishments, cleaning, waste, and pest control [114,115]. Another standard worth mentioning is ISO 14001 (Environmental Management System) as it helps organizations manage their environmental impact through a commitment to pollution prevention and mitigation, legal compliance both locally and internationally, waste management, and continuous improvement [116,117].
4. Ulva Compounds of Interest
4.1. Ulvan
Ulvans are a distinct polydisperse sulfated heteropolysaccharide made up of variable quantities of uronic acids, including glucuronic acid (2.6–52.0%) and iduronic acid (0.6–15.3%mol), interspersed with sugar moieties. Neutral examples are rhamnose (5.0–92.2%mol), xylose (0.0–38.0%mol), and glucose, connected by α and β-1 → 4 bonds [118], representing according to the literature between 8 and 29% of the dry weight of biomass [119].
Ulvans are the principal polysaccharide compound of the cell wall of Ulva sp. [120], and they present gelling properties [120]; water solubility [119]; and antiviral, antioxidant, anticoagulant, antihyperlipidemic, and anticancer activity, adding to immunostimulant effects (for more information read Kraan 2013 [121], Lahaye and Robic 2007 [122], Alves et al., 2013 [123]) and the ability to accumulate heavy metals (for more information read Webster and Gadd 1996 [124], Bocanegra et al., 2009 [125], Schijf and Ebling 2010 [126], Garcia-Poza et al. [118]).
The literature has demonstrated the significance of rhamnose owing to its impact on the biosynthetic pathways of the dermis and plant immunity. Uronic acids, including glucuronic and iduronic acids, along with their sulfate esters, are vital components of mammalian glycosaminoglycans (GAGs) such as heparin, heparan sulfate, and dermatan sulfate [127].
Cellulose is a linear polysaccharide composed of repeating glucose units linked together by β-1,4 glycosidic bonds; it is the most abundant polysaccharide found in nature and serves as a structural component in the cell walls of plants and algae, providing strength and rigidity, thus contributing to their overall structural integrity or mechanical support. The molecules are organized into microfibrils, long fibers, and insoluble forms, which form a network [128] that is also known for its high resistance to enzymatic degradation [127].
The composition and structure of polysaccharides in Ulva sp. can be influenced by several factors, including environmental conditions, growth stage, and extraction methods [128]. As techniques used to analyze the composition and structural characteristics of these polysaccharides, we have Fourier transform infrared spectroscopy (FTIR) and nuclear magnetic resonance (NMR) spectroscopy [128].
4.2. Pigments
4.2.1. Chlorophylls
As per Aronoff (1957) [129], chlorophylls are green photosynthetic pigments employed by photoautotrophic organisms like plants and algae. They absorb light and, in conjunction with carbon dioxide fixation, produce the carbohydrates necessary for the growth of plants and algae [129,130,131].
Structurally, they are composed of a reduced porphyrin ring, with a central magnesium atom, and a long hydrophobic tail (phytol), which confers them low solubility in water. However, due to the presence of ester and carbonyl groups, polar functions are lipid-soluble [130]. The most common types of chlorophylls present in green algae are the a and b, considering that the concentrations exhibit variability contingent upon species, geographical location, and environmental conditions [132].
Chlorophylls are used in the food industry as natural colorants in foods and beverages; however, other important features are being reported, namely their antioxidant, antitumoral, and antimicrobial activities (according to Lordan et al., 2011 [133], Lanfer-Marquez et al., 2005 [134], Luo et al., 2015 [135] and Sudakin 2003 [136], respectively). A recent study conducted by Diogo et al., 2018 [137] proved the possibility of chlorophylls to be used as precursors of photosensitizers for photodynamic therapy, namely for cancer treatment and inactivation of microorganisms [130].
4.2.2. Carotenoids
Carotenoids are natural pigments originating from five-carbon isoprene units, which enzymatically polymerize to create regular, extensively conjugated 40-carbon structures containing up to 15 double conjugated bonds [130,131]. They are the most prevalent pigments found in nature, existing in all algae, higher plants, and numerous photosynthetic bacteria [130,131]. They represent photosynthetic pigments in red, orange, and yellow wavelengths. Green seaweed species are characterized as containing β-carotene, lutein, violaxanthin, neoxanthin, and zeaxanthin [131]. Ulva spp. have been reported to contain various carotenes, including 9-cis-β-carotene, all-trans-β-carotene, α-carotene, and other carotene isomers. Additionally, reported xanthophylls include lutein, violaxanthin, antheraxanthin, zeaxanthin, neoxanthin, and β-cryptoxanthin [128].
In terms of commercial production, carotenoids are widely recognized for their antioxidative properties, which play a crucial role in counteracting the detrimental effects of free radicals, thereby mitigating oxidative stress and safeguarding cellular and tissue integrity [128]. For that, they have an important role as food additives, animal feed, colorants, medications, and nutraceuticals. For example, according to Pham et al., 2021 [138], beta-carotene is a precursor to vitamin A and is pivotal in preserving optimal vision and reinforcing the immune system. Madhavan et al., 2018 [139] noted that Lutein and zeaxanthin have a crucial role in sustaining optimal eye health by accumulating within the retina and offering protection against age-related macular degeneration and various other vision-related disorders [128].
4.3. Phenolic Compounds
Phenolic compounds are secondary metabolites found in plants and algae [128] and are defined as molecules with hydroxylated aromatic rings [140,141]. These phytochemicals show a wide variety of chemical structures, from simple moieties to high molecular polymers. The biogenetically primary synthetic pathways that produce these phytochemicals are the shikimate or the acetate pathways [140,141].
Phlorotannins, bromophenols, flavonoids, phenolic terpenoids, and mycosporine-like amino acids (MAAs) are the polyphenolic compounds found in seaweeds [140,141]. Phlorotannins are oligomers of phloroglucinol and serve as primary and secondary metabolites according to Santos et al., 2019 [142]. Bromophenols are characterized by the presence of phenolic groups with varying degrees of bromination with ecological functions, such as chemical defence and deterrence, with studies according to Liu et al., 2011 [143] and Stengel et al., 2011 [144] revealing a wide variety of beneficial ecological activities. Flavonoids, as evidenced by Bilal Hussain et al., 2019 [145] and Mukherjee 2019 [146], are characterized as having heterocyclic oxygen attached to two aromatic rings, which might vary depending on the degree of hydrogenation. The different types are rutin, quercetin, hesperidin, kaempferol, catechin, daidzein, or genistein [140,141]. The origin of phenolic terpenoids (also known as terpenes or isoprenoids) form the largest group of natural products with more than 30,000 different structures and are products of the isoprenoid biosynthetic pathway [147]. Lastly, MAAs are a group of UV-absorbing compounds [141] whose molecular structure is based on a cyclohexanone or cyclohexenine ring with amino acid substituents [140].
Bromophenols and flavonoids of green seaweeds have antioxidant activities. They have already been tested and proven in different species; for example, Tang et al., 2004 [148] and Khanavi et al., 2012 [149] referred to the phenolic fraction of Ulva clathrata and Ulva flexuosa, which has an antitumoral and antibacterial effect; also, research on Ulva prolifera showed antidiabetic properties in Yan et al., 2019 [150] and Lin et al., 2018 [151]. Other examples were proved in other species like Ulva compressa (as Enteromorpha compressa), Ulva intestinalis, Ulva linza, Ulva australis (as Ulva pertusa), Capsosiphon fulvescens, and Chaetomorpha moniligera [141].
Farvin and Jacobsen (2012) [152], who worked with water and ethanolic extracts of 16 species of seaweeds collected along the Danish coasts, observed that U. lactuca had the following acids: gallic, protocatechuic, and gentisic (only by ethanol), hydroxybenzoic, chlorogenic, vanillic, syringic, caffeic, salicylic, and coumaric (by water as well). Another study developed by Aslan et al. (2019) [153], which collected four macroalgae from the European and Asian shores of Istanbul Strait to find the most suitable solvent system to extract polyphenolic compounds, noted that ascorbic, gallic, coumaric, caffeic, and ferulic acids, catechin, rutin, and quercetin were found at the highest concentration on U. lactuca.
U. lactuca has additional flavonoids, kaempferol, and quercetin. They are known for their antioxidant and anti-inflammatory effects. Kaempferol has been extensively investigated for its possible health advantages, especially its involvement in lowering the likelihood of chronic illnesses such as cancer, cardiovascular problems, and neurological ailments [141].
4.4. Fatty Accids
Fatty acids constitute vital components of lipids and hold significant roles in human health and metabolism. The composition and concentrations of fatty acids can vary depending on factors such as species, geographic location, and environmental conditions [128].
Gao et al., 2017 [87] obtained for wild stalks lipids 5.4 ± 0.4%, while for sterile stalks, lipid content was 13.8%. The lipid content for sterile thalli was higher compared to other Ulva species: 2.5–3.5% in U. clathrata and 7.9% in U. lactuca. There was also a high proportion of monounsaturated fatty acids (MUFA), polyunsaturated fatty acids (PUFA), omega-3 fatty acids (n3-FA), and Omega-6 fatty acids (n6-FA) such as linoleic acid (LA) and gamma-linolenic acid (GLA) [87,128].
Omega-6 fatty acids are crucial for preserving healthy cell membranes, moderating inflammation, and promoting overall well-being. In a study by Valério Filho et al. (2023) [154], a thorough examination of the fatty acid composition in U. lactuca unveiled notable quantities of various fatty acids. Palmitic acid (16:0) emerged as the primary constituent, accounting for 21.51%, while oleic acid (18:1n9c) contributed 10.60% as a monounsaturated fatty acid. Some literature also shows that green algae also contain large amounts of palmitolinolenic (16:3ω3) and palmitidonic (16:4ω-3) acid [118,128,155,156].
Also, notable components are linolelaidic acid (LA; C18:2ω-6), which is found in most of the species, and α-linoleic acid (ALA, C18:3ω-3), which is characteristic of Ulvales. Furthermore, the omega-3 fatty acid eicosapentaenoic acid is 1.67%. The presence of these fatty acids in U. lactuca suggests its potential as a dietary source of essential fatty acids [128].
4.5. Proteins
Plant proteins are frequently called inadequate protein sources because they lack one or more necessary amino acids [157]. Ulva sp., according to several authors, contains substantial quantities of proteins (7–29% dry weight, DW) [70,132]. However, some study has indicated that it can reach up to roughly 40% in IMTA systems, where large quantities of nitrogen are incorporated into proteins with a N to P conversion factor of 5 [32].
Kazir et. al. in 2019 used five different ways to extract proteins and obtained between 20% and 62% of the DW of protein on a DW basis, whereas the carbohydrate percentage ranged from 5 to 37%. The remaining non-protein and non-carbohydrate fraction in all extracts consisted of phenolic compounds, minerals, and other phytochemicals such as pigments [132].
Ulva protein contains large levels of essential amino acids, making it a potentially valuable dietary protein source. Indeed, there is remarkable resemblance in the overall amino acid content of the alga and that of egg ovalbumin, such as histidine [96].
Among with amino acids, Shuuluka et al., 2013 [158] and Fleurence et al., 1995 [159] noted the most prevalent amino acids are aspartic and glutamic acids, which can account for up to 26 and 32% of the total amino acids in U. rigida and U. lactuca (as U. rotundata), respectively [70]. On the contrary, Gao et al., 2017 showed that glutamic acid and alanine existed as the highest amino acids in wild-type and sterile thalli of Ulva sp. [87].
Other essential amino acids such as valine, leucine, lysine, and threonine make up 42% of the total amino acid composition in U. lactuca, according to Yaich et al., 2011 [160].
5. Commercial Potential
According to FAO, worldwide algal output (including cultivation and wild collection) rose by more than 60 times between 1950 and 2019, from 0.56 million (wet) tons to 35.82 million tons. In 2019, 54 countries/territories contributed 35.8 million tons of global algal output (including seaweeds and microalgae), with cultivation accounting for 97% of the total. Green algae contributed with 32,926 tons; the highest production countries were the Republic of Korea through cultivation and India through wild collection [161].
Ulva species have been farmed in many regions of the world in experimental commercial systems due to their potential utility as food, animal feed, biofuel, and bioremediation, as detailed more below. The initial trials of commercial Ulva culture, according to Fannin 1983 [156], documented the inaugural commercial Ulva culture trials in America, coinciding with the initiation of the US Ocean Food and Energy Farm project in 1973, where Ulva was employed as biomass for biomethane conversion. However, due to the challenges associated with offshore development methods, Ulva production for biofuels was subsequently discontinued [156]. In the meantime, Dan et al., 2002 [162] and Hiraoka and Oka, 2008 [163] showed there is a folklore in Japan to eat Ulva as food, and if we look to the statistics [93], it is sold as “aonori” and “aosa,” reaching around 1500 tons of dry weight per year. In 2019, according to FAO, 2021, Ulva sp. contributed 2155 tons to different industries [161].
In addition, Ulva has also been cultivated for other sectors, for instance as a biofilter in integrated multi-trophic systems where Ulva culture is combined with aquaculture of marine animals reported by Neori et al., 2000 [164] in Israel and Al-Hafedh et al., 2014 [165] in Saudi Arabia. Robertson-Andersson et al., 2008 [166] reported that it has been intensively studied in South Africa as animal feed; also, Ulva was cultivated in outdoor tanks to investigate its biomass production potential for bioenergy in Denmark according to Bruhn et al., 2011 [167] and Mata et al., 2016 [168] referred to the cultivation in Australia to explore its potential in nutraceuticals and cosmeceuticals [93].
5.1. Human Food
Algae vary in composition based on the species, geographic location, water conditions, and harvest season. However, the distinguishing feature of all seaweeds is their high-water content of more than 70%, which implies that they should be consumed soon after harvesting or dried [169], and between 20 and 50% of its dry weight contains essential minerals for human consumption.
In terms of nutritional value, green algae such as Ulva, as previously mentioned, may be a source of important amino acids, with some of them, such as histidine, present at amounts equivalent to vegetables and eggs [96]. It should also be noted that the overall quantity of an element in seaweed does not properly predict the absorbable amount in the gastrointestinal (GI) tract. Cherry et al. (2019) [170] reported that to enable saccharolytic fermentation in the human colon, the gut microbiota must produce functional carbohydrate-active enzymes (CAZymes) capable of breaking down seaweed glycans as carbon sources. Meanwhile, Ripol et al. (2017) [171] investigated the bioaccessibility of bioactive compounds from green algae (Chaetomorpha linum, Ulva intestinalis, U. lactuca, and Ulva prolifera) cultivated in multitrophic aquaculture environments. Their findings suggested that the absence of bioaccessible phenolic compounds in human digestion indicates the inability to eliminate these compounds, thus suggesting the likely necessity for extraction [170,171].
Furthermore, this alga can be a rich source of minerals (especially calcium, magnesium, iron, and zinc, as well as high levels of potassium; calcium, sodium, and iron levels were higher than those recorded for land plants), lipids, and phytochemicals. Extraction of proteins from two marine macroalgae, Ulva sp. (Chlorophyta) and Gracilaria sp. (Rhodophyta), for food application, evaluated digestibility, amino acid content, and antioxidant characteristics of the protein concentrates [96,132].
MacArtain et al. [172], when analyzing the nutritional composition of ten species of algae most used in food, observed that for U. lactuca the total fiber corresponds to 3.8 g/100 g, and, compared to food, it is below that of lentils and above that of rice, carrots, apples, bananas, cabbage, and plums; while carbohydrates are smaller than the foods mentioned above with 4.1 g/100 g of dry weight. The mineral composition (Ca, Po, Mg, Na, Cu, Fe, I, Zn) is equivalent to 1.4 g/100 g. Algae contain many forms of antioxidants, including vitamins (hydro- and fat-soluble) and pigments. In this experiment, vitamin C and B3 were in the highest proportion (10 and 8 mg/8 g dry weight, respectively) for a total of 18.1 mg/8 g [172].
Around 20% of the Asian diet consists of seaweed, and Ulva is commonly consumed as a meal or as an extract, for example, in health supplements; in Western nations, seaweed is mostly utilized as food additives or extracts (as natural colorants in foods and beverages) [70,173]. Another approach utilized by the food business is to improve the visual attractiveness of items [132].
5.2. Agriculture and Livestock Animals
Historically, seaweed has been used not only for human food but also for animal food since the 19th and early 20th centuries in several European countries, such as Norway, Scotland, and France, and in America. These feeds were used for cattle, horses, and poultry [70].
Abudabos et al., 2013 [175] found that including U. lactuca (up to 3%) in maize diets for 21 days showed no detrimental impacts on assessed chicken production parameters but enhanced carcass features in terms of yield and breast yield [70,157].
References in Ventura and Castañón 1998 [176] and Cabrita et al., 2017 [177] showed that nutritional assessment of U. lactuca for goats and sheep helps its presence as medium-quality forage [4]. Holdt and Kraan 2011 [178] and Dhargalkar and Pereira 2005 [179] state that the use of macroalgae in feed led to an increase in milk output in cattle and the growth rate in lambs and an improvement in the color of the yolk in eggs [180].
In terms of marine animals, past research in fish aquaculture has shown that integrating green algae in their diet improves fish development, with Ulva biomass being mostly used as feed for abalone, shrimp, and fish aquaculture. For example, Hashim and Saat 1992 [181] showed that incorporation of 5% Ulva spp. in Snakehead Murrel feeding increased growth rate, feed efficiency, and feed consumption [70]. Mustafa et al., 1995 [182] showed that the body weight improvement and feed effectiveness of red sea bream Pagurus major was boosted by the adding of 5% algae meal. Furthermore, Gabrielsen and Austreng 1998 [183] found that Atlantic salmon (Salmo salar) fed Ascophyllum nodosum had significantly higher levels of lysozyme (3.28 ± 0.49 g mL−1) compared to controls (2.99 ± 0.65 g mL−1). This suggests that alginate has an immunostimulant effect, improving stress response and disease resistance in marine animals. According to Naidoo et al. (2006) [184], abalone fed a mixed diet of U. lactuca, Gracilaria gracilis, and kelp exhibited a growth rate of 0.066 mm per day in shell length and 0.074 g per day in body weight. However, abalone fed solely dried kelp showed a lower growth rate, with only 0.029 mm per day in shell length and 0.021 g per day in body weight [93,184].
The alga A. nodosum is also used in agriculture, commonly found on the northwestern coast of Europe and in the northwest of North America. Its extensive use is mainly due to its abundance, ease of collection, and growth in areas close to processing infrastructures [180]. It has already been demonstrated in Sharma et al., 2014 [185] and Van Oosten et al., 2017 [186] that commercial hydrolyzed extracts of A. nodosum (ANE) exhibit growth-stimulating activities in treated plants, when repeatedly applied at very low doses, and are called “biostimulants”. Basak 2008 [187], Chouliaras et al., 2009 [188], Abdel-Mawgoud et al., 2010 [189], and Frioni et al., 2018 [190] mention that foliar spraying improved the quality of fruits of watermelons, apples, olives, and grapes [191].
Another use is as a natural biopesticide, where it can control pests and diseases. There are several bibliographies from which we can take the following examples: in Hankins and Hockey 1990 [192], a reduction of the population of two-spotted red spider mites on treated plants on strawberry crops occurred; in Subramanian et al., 2011 [193], it is demonstrated that it decreases the advancement of diseases, which is connected with the expression of jasmonic-acid-related gene transcript in Pseudomonas syringae and Sclerotinia sclerotiorum, on Arabidopsis crops; and in Panjehkeh and Abkhoo 2016 [194], tomato crops stimulate the appearance of defense-related genes or proteins off Phytophthora capsici [191].
5.3. Pharmaceuticals
There are several studies throughout the literature demonstrating that various phytochemicals extracted from the algae in general exhibit different activities such as
- Anticancer—Aronoff 1957 [129] showed that chlorophylls may be employed as precursors of photosensitizers for photodynamic therapy, namely for cancer treatment and microorganism inactivation [130,195]. Another study carried out by Lee et al. (2004) [196] tested the water-soluble component of U. lactuca extract at concentrations up to 140 µg/mL on human leukemia cells (U937). Surprisingly, 50% growth inhibition was reported following therapy. At a dosage of 100 µg/mL, splenocytes showed increased proliferation. Furthermore, the macrophage cell line (RAW 264.7) increased nitric oxide production, which is thought to regulate cytokine action [173].
- Antioxidant—Antioxidants found in chlorophylls, xanthophylls, and carotenoids can function as free radical scavengers, thus helping to safeguard cellular and tissue integrity against oxidative damage and helping to preserve cellular well-being and mitigate susceptibility to disease chronic illnesses such as cancer and improve immune system response [130,180].
- Antihypercholesterolemic—The high yield of soluble dietary fiber is shown in species like Kappaphycopsis cottonii (formerly Eucheuma cottonii) (Rhodophyta), Caulerpa lentillifera (Chlorophyta), Sargassum polycystum (Phaeophyceae), Gymnogongrus durvillei (formerly Ahnfeltiopsis concinna) (Rhodophyta), Gayralia oxysperma (Chlorophyta), Chondrus ocellatus (Rhodophyta), and U. lactuca (as U. fasciata) (Chlorophyta), which can help reduce cholesterol [130,180]. Another study carried out by Hassan et al. (2011) [197] subjected albino rats as an in vivo model to being fed precipitated polysaccharides for 21 days. The control group was given a reference medication named Lapitor (Atorvastatin Ca). Ulvan-treated rats had decreased levels of blood total lipids and triglycerides, total cholesterol, LDL (low-density lipoprotein), and VLDL. Improved levels of atherogenic index, HDL (high-density lipoprotein) (180%), and enzyme activity creatine kinase and lactate dehydrogenase were also seen. Furthermore, hepatic enzyme activity (glutathione peroxidase, superoxide dismutase, and catalase) were enhanced. On the other hand, a decrease of total glutathione and thiol was also detected, indicating that ulvan is a strong drug against induced hypercholesterolemic situations [173].
- Anti-inflammatory—Awad (2000) [198] extracted a physiologically active steroid (3-O-β-D-glucopyranosyl clerosterol) from U. lactuca and tested it on mice ear edema. At dosages of 1000 and 1500 mg/year, edema decreased significantly (p < 0.05) (62.2 and 72.2%, respectively). The green macroalgae, U. lactuca, has been studied in a rat model to prevent edema by lowering inflammation [173].
- Antimicrobial—It was identified in Abbassy et al., 2014 [199] that U. lactuca has insecticidal properties. The acetone extract was considered the best against inhibiting the mosquito population, killing the larvae of Culex pipiens (a hematophagous mosquito belonging to the Culicidae family, a vector of diseases such as Japanese encephalitis, meningitis, and urticaria). Spavieri et al., 2010 [200] reported that Ulva extract indicated only moderate antiprotozoal (specifically trypanocidal) activity (IC50 = 34.9 µg/mL) against Trypanosoma cruzi but good leishmanicidal activity (IC50 = 12 µg/mL) [173].
Antiviral—Sathivel et al., 2008 [201] worked on hepatitis, one of the most common liver diseases, which can be caused by certain germs, viruses, or chemical toxins. U. lactuca polysaccharide extract was evaluated against chemo-induced hepatitis in rats, using Galactosamine at 500 mg/kg body weight. Rats pre-treated with ulvan for 3 weeks (30 mg/kg body weight) indicated a substantial value in retardation of any form of abnormality and gave the liver protection, presumably by demonstrating free-radical quenching activity [173].
- Anticoagulants—These are remarkably exploited in medicines to avoid blood clotting. Mao et al., 2006 [202] identified the anticoagulant properties of pure polysaccharides from Ulva conglobata using a hot water and ethanol extraction procedure, followed by ion exchange and HPLC purification. Sulfated polysaccharides were thought to limit thrombin activity both directly and via increasing the potency of heparin cofactor II (HCII) [173].
- Antiobesity and antidiabetic effects—BelHadj et al. (2013) [200] found that extracts from U. lactuca may offer benefits in managing obesity and diabetes. Components of U. lactuca exhibit anti-obesity effects by inhibiting adipogenesis and enhancing lipid metabolism. Additionally, they have been shown to enhance glucose metabolism and insulin sensitivity, suggesting potential antidiabetic properties [128,203].
5.4. Cosmetics
Literature like Biba 2014 [204] and Cardozo et al., 2007 [205] mention that bioactive compounds with antioxidant activity in the cosmetics industry help to delay skin aging, skin inflammation, and skin cancer and provide protection against ultraviolet rays (sunscreens). In Kim et al., 2008 [206], Chondrus crispus can be cited as an example, which is full of polysaccharides and minerals including manganese, zinc, calcium, and magnesium and has a moisturizing, conditioning, calming, and healing action. Brown algae, which in general have many vitamins, minerals, and essential fatty acids, including omega 3 and 6, are known to help with skin regeneration and health. Leal et al., 2016 [207] refers to Asparagopsis spp. (Rhodophyta), which is already cultivated to produce extracts for skin treatment in France, where approximately 5 tons (fresh weight) of algae are used to meet industry demand [180].
5.5. Biofuel
Fossil energy has been a key driver of the technical, social, economic, and developmental progress that has trailed since the Industrial Revolution between 1760 and 1820/1840. Fossil fuels (coal, oil, gas) continue to play a dominant role in global energy systems. It is estimated that around 136,018 TWh were consumed in 2021 [209]. However, when burned, they emit carbon dioxide (CO2), which is the leading cause of global climate change and air pollution. One option is the usage of biofuels.
Biofuels are a form of energy derived from biomass. Currently, bioethanol from sugar cane or maize, as well as biodiesel from oilseed plants, are generated on a wide scale thanks to environmentally friendly oil extraction and well-established farming processes [210]. However, recent developments in the generation of biofuels from food crops have highlighted numerous ethical problems due to the food vs. fuel debate, permitting openness and interest in algae.
According to Osman et al., 2020 [210], the production of biodiesel and bioethanol from 22 most promising seaweed species, harvested from Abu Qir Bay, Egypt, was assessed using three distinct methods. The first approach included producing biodiesel by extracting lipids and then trans-esterifying them. The second technique included directly fermenting all biomasses to make bioethanol without first extracting the lipids. The third approach involved producing biodiesel from the extracted lipids, followed by fermenting the remaining lipid-free biomass to make bioethanol. They found that Ulva intestinalis presented the highest value of relative increase in energy compounds; the third route increased the efficiency of hydrolysis and the availability of sugar, which resulted in a 14.1% higher bioethanol yield; and the sequential recovery of energy presented gross energy production 170.9% and 82.0% higher than the individual production of biodiesel or bioethanol, respectively. Korzen et al., 2015 [211] developed a simultaneous saccharification and fermentation process based on sonication to produce bioethanol from Ulva rigida and generated a maximum of 6.2 wt% ethanol pursuant to sonication in 3 h (65.5%) and only 4.9% by weight of ethanol under incubation even after 48 h (67.9%).
However, according to the research, bioethanol produced from sugar cane or other agricultural crops is significantly less expensive than the Ulva biorefinery, independent of the technique. As a result, efforts to develop a competitive supply of bioethanol from green algae will not be financially feasible in the future.
5.6. Bioplastic
Plastic pollution is a major problem across the world, with approximately 20% of plastic in the oceans coming from marine sources and 80% coming from terrestrial sources. Plastic is a unique substance because it is inexpensive, adaptable, light, and resistant. It is utilized in various applications such as medicinal equipment, packaging, 3D printing, home appliances, autos, electronics, and so on, making people dependent [212].
Annual plastic production has increased from 2 million metric tons in 1950 to 460 million metric tons in 2019 [212]. However, these plastics will take thousands of years to degrade and will eventually accumulate in landfills or the natural environment.
There are many methods and materials for making bio-based plastics, and the essential ingredients are polysaccharides, proteins, and lipids. Several substances utilized today include corn, potatoes, vegetable oils, wood, food waste, beetroot, sugar cane, and cereals, as the major forms of bioplastics nowadays are starch-based, followed by polylactic acid, (PLA—a highly versatile and biodegradable aliphatic polyester), poly-3 hydroxybutyrate (PHB—a polymer that belongs to the polyester class), polyamide 11 (PA 11—polymer from the nylon family), and polyethylene (PE—organic polymer) [213].
Seaweed is promising to produce bioplastics as it can form films. Ulva spp. can also be used to generate polylactic acid (PLA) film because of their starch content, which acts as biomass for fermentation with Lactobacillus plantarum. Some green algae taxa are underused for the manufacture of bioplastics, whereas others are solely employed to make polymers, such as Caulerpa sp., Codium sp., and Cladophora sp. (Chlorophyta) [213].
Bioplastic properties and characterization details tend to be categorized into the following categories: physical (thickness, solubility, water vapor permeability, conveyance of water vapor rate, and moisture content); optical (transparency, opacity, and light transmittance value); mechanical (thickness); structural, thermal, antioxidant, antimicrobial, and biodegradable [213]. However, generic methods are not sustainable to produce bioplastics due to yield, time, and costs; hence it is advised to do more research with different species.
5.7. Feed Aquaculture and Bioremediation
Fed aquaculture (animal feed) is a growing industry, but according to scientists like Chopin [214,215] and others [216,217,218], it dumps heavy loads of nutrients into coastal waters, potentially leading to the deterioration of local marine environments. A possible solution to this problem is to integrate seaweed into fish farming, that is, to combine fed aquaculture with extractive aquaculture (algaculture). There are already studies that combine seaweed culture with terrestrial fish tanks or open sea fish cages [219], as well as studies on Ulva species used mainly from the point of view of treating lagoon effluents/land tanks [220].
According to Neori et al., 1996 [218], seaweed removed up to 90% of the nutrients discarded from intensive fish farming [219]. Al-Hafedh et al., 2011 [165] selected U. lactuca to investigate its relevance in integrated aquaculture on the Red Sea (Saudi Arabia). The results confirm that tank-grown Ulva was greatly effective in treating fish farming effluents. They also determined that the algae stalks require greater surface area than the overall size of the culture tank to absorb enough light for photosynthesis, and it functioned well with marine tilapia (O. spilurus), as well as with effluent from sea bream culture, Sparus aurata in Israel. The literature also states that it is suitable for integration with shrimp systems (Kuruma, Penaeus japonicus larvae) resulting in greater survival and better growth of shrimp larvae and lower bacterial density due to high tolerance and affinity for ammonia absorption [165].
Natify et al., in 2015 [221], discovered that Nile tilapia given 5% seaweed outperformed those fed the control (and 10% seaweed). This study concluded that U. lactuca bran may be used as a supplemental element in the diet of Nile tilapia without affecting their zootechnical performance, feed consumption, or body structure.
Gao et al., 2016 [222], working with U. rigida stems, discovered that higher temperatures boosted nitrate and phosphate absorption when they were vegetative. However, the greater temperature resulted in negative nutrient intake during reproductive events, indicating that the thalli were releasing rather than absorbing nutrients from the ocean. Possibly due to the release of spores, nutrients are also discharged, and increasing temperature may increase the rate of decomposition, thereby increasing the rate of nutrient release. As a result, it was proposed that Ulva species may actually be a source of nutrients when they reproduce, which has consequences for biofilter effectiveness, as sustaining a long-term vegetative state seemed to be crucial to improving Ulva species’ biofiltration effectiveness [220].
We can thus say that the cultivation of algae along the coast can function as an effective biofilter to alleviate the problem of eutrophication throughout the world and provide benefits in fish growth; certain species can be used as bioindicators of nutrients in the water column, thus being able to improve aquaculture systems and restore ecosystems.
6. Ulva sp. as Environmental Danger
Ulva spp. is widely recognized for generating uncontrolled, fast, and massive mass production known as “green tides.” The free-floating shards can operate as a nucleus to produce green floods in new sites, offering enormous ecological and economic risks [4]. There are several examples in the literature: in Belfast (1911), Laguna of Venice (1930), Galicia (1999), New Zealand (1930s, 1991–1993, 1998, and 2003–2007), Maine through Long Island Sound in the USA (reports in 1958, 1973, 2000, 2004, 2005, 2009, and 2013), Bahía de la Paz in California (2013 to 2015), Tokyo Bay (1995, 1999, 2002, 2004), Brittany and Florida coasts (2008), Australia (2012), Yellow Sea in China (from 2007 to 2019), and Muttukadu Lagoon, India (2023) [223,224,225,226,227].
During the bloom event, the fast-growing seaweed begin vying for sunlight and nutrients, causing ecosystem harm through hypoxia and acidity and interfering with the variety and richness of the habitat [228].
Algal blooms are natural in the environment but have increased significantly due to pollution. According to Also and Park [229], during El Niño circumstances (e.g., 2007), when nutrient-rich deep waters upwell offshore and reach the harbor, Ulva blooms are especially severe [223]. In summary, increased urbanization has led to changes in the global carbon cycle [230], increased coastal eutrophication [231,232], altered meteorological patterns [233,234], and overfishing [235], which act together in intricated paths to affect nearshore habitats [236].
This also shows that algal blooms will increase in different parts of the world, but, like the old saying, “In every difficulty there is always an opportunity.” One of these opportunities is to create more sustainable measures for overfishing. According to Eriksson et al. [237], there are depleted sections of top predator populations, indicating increases in smaller fish and significant covering of ephemeral algae. This allows consideration of the need for broader ecosystem processes and trophic interactions [223].
However, if we compare with other seaweed such as red and brown algae, Ulva sp. cannot be cultivated in deep waters due to the light intensity need. At the commercial level, the most interesting seaweed belongs to the genera Porphyra, Fucus, Chondrus, Laminaria, Gracilaria, Sargassum, Palmaria, Kappaphycus, Gelidium, Spirulina, and others, i.e., red and brown seaweeds. This means that Ulva sp. is not very well explored scientifically and economically.
7. Future Roads
CO2 emissions account for 82% of global warming. The remainder is mainly associated with methane and nitrous oxide gases, with a much greater potential for temperature rise. In 2018, a UN Environment publication revealed that global greenhouse gas emissions in 2030 need to be 25% to 55% lower than in 2017 to put the world on a lower-cost path to limit warming global between 2 °C and 1.5 °C warmer [238].
Algal communities serve a crucial function in marine ecosystems by absorbing CO2 due to their autotrophic nature and higher organic matter production through photosynthesis compared to consumption through ecosystem respiration. According to Duarte et al., seaweed and its agricultural activities consume approximately 2.48 million tons of CO2 per year. In 2014, global seaweed production reached 27.3 million tons, capturing 100% of carbon, equivalent to 24.8% of dry weight carbon production [96].
The rate of carbon capture is determined by the growth rate and the carbon concentration. Some researchers observed that algae and marine plants have been estimated to be accountable for over 70% of the world’s carbon storage. They were able to demonstrate that Ulva species had the greatest carbon capture capacity compared to algae belonging to the Chlorophyta, Phaeophyceae, and Rhodophyta [220]. Ulva spp.s’ quick development is related to its strong photosynthetic rates and high absorption capacity of carbon and nitrogen/nitrogen nutrients required for growth [96,220].
Studies that Gao and others carried out found that high temperatures can raise the increase of Ulva biomass, as happened with U. lactuca (as U. fasciata) doubling its growth rate with an increase of 5 °C (from 20 to 25 °C) with a salinity of 25, according to Mantri et al. [239]. The findings show that cooler temperatures in higher latitudes inhibit the development of U. rigida even throughout the summer, and so Ulva species may benefit from future oceanic rising temperatures. Higher temperatures, pCO2, and nitrate triggered more reproductive events, leading to a slower growth rate in the near term as well as higher carbon content, culminating in a higher carbon capture efficiency [220].
In the areas of medicine and pharmaceuticals, from Ulva sp. can be developed a food supplement with antioxidant and nutraceutical properties accordingly to age and physical conditioning, for example, for athletes and younger and older people. The world is also going to the nutraceutical niche; people are focusing on fighting diseases not only with medicine/pills but with food as well. There are several gastroenterologists and biochemistries revealing that the intestine is the human body’s second brain, and “We are what we eat.” I imagine in the supermarkets, food based on different seaweeds will support the medication to fight cancer and other diseases. Moreover, comprehending the metabolic patterns of seaweed opens up the potential for developing novel cultivation systems tailored to produce seaweeds rich in specific compounds. This in the future can present an important field, and seaweeds can be a key to obtaining high-quality natural drugs.
In addition to the areas described above, from what we know with the help of science, there will be the possibility that algae biomass could be used for construction, as is the case with Sargablock (Omar Sánchez) who invented a brick partially made out of sargassum that spoils Riviera Maya shores between April and October, inspired by the memory of his family’s little adobe house, and Ms. Larsen, who built an installation from seaweed [240,241,242]. Seaweed also offers great insulation of floors, walls, and ceilings, as well as having extraordinarily good acoustic qualities and contributing to the regulation of indoor air humidity parameters [242,243].
From textile production, AlgiKnit created an algae-based yarn fiber, and Violaine Buet employs weaving, coloring, printing, embroidery, embossing, engraving, stitching, pressing, and braiding methods on seaweed to make personalized items using the biodegradable algae [244]. From Ulva sp. can be extracted pigments to be used for drink industries and especially the textile industry for coloring and printing methods in clothes.
The human population grows every day, and the pressure on nature is also increasing exponentially. Resources are limited, and creative ways to find new resources while facing climate change are needed. Seaweed is one of the options. Seaweeds are considered one of the primary forms of life on Earth that do not compete with arable land to be cultivated. As basic organisms, they can produce a variety of chemicals, depending on location and abiotic conditions. In the end, seaweed offers several chances to tackle key global issues such as food security, climate change, and environmental degradation, as well promote sustainable development, thus providing an attractive resource for the foreseeable future of humanity.
Ulva sp. is one of the most cosmopolitan algae, adapted to high and low temperatures, and as an “r selected” species is also opportunistic, creating “green waves” in some parts of the world. Light, temperature, salinity, nutrients, oxygen, and microbiomes have a big influence on their reproduction, growth, and density in the ecosystems. Different types of cultivation and methodology can be applied as the objective to cultivate seaweed. Based on the composition, Ulva sp. can be used for different industries based on the composition, such as food (human and animal)—itself, pigments, chlorophylls, and proteins; pharmaceuticals—phenolic compounds, fatty acids; cosmetics—polysaccharides, extracts; and biofuels—fatty acids, bioplastics, and bioremediation.
8. Conclusions
Thus, this review shows Ulva sp. can make important contributions to the marine ecosystems, humankind, aquaculture, and biotechnological innovation, demonstrating its value throughout both environmental and socioeconomic contexts. On the other hand, it is critical to manage and control Ulva populations and farm it responsibly in order to avoid overgrowth and green tides and preserve environmental equilibrium, although there are several pathways and industries that can benefit from Ulva sp. biomass to promote the circular bioeconomy. Research still needs to develop the Ulva sp. bioactive chemicals segment, which might be used in a variety of biotechnological industries, including medical, cosmetical, and pharmaceutical. Ulva extracts have been researched for their antioxidant, antibacterial, and anti-inflammatory behaviour, indicating their high prospective potential for application in the development of novel products and therapies.
As the seaweed world is still complex and unknown, we must be creative. As George Scialabba said, “Maybe imagination is just intelligence having fun,” and in a world where everything changes, perhaps creativity is our greatest ally in using so much biomass.
Author Contributions
Conceptualization, S.P.C. and J.C.; investigation, S.P.C.; writing—original draft preparation, S.P.C. and J.C.; writing—review and editing, S.P.C., J.C., and L.P.; supervision, L.P. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by FCT—Fundação para a Ciência e Tecnologia, I.P. by project reference UIDB/04004/2020 and DOI identifier 10.54499/UIDB/04004/2020 (https://doi.org/10.54499/UIDB/04004/2020). This work was supported by FCT—Fundação para a Ciência e Tecnologia, I.P. by project reference UIDP/04004/2020 and DOI identifier 10.54499/UIDP/04004/2020 (https://doi.org/10.54499/UIDP/04004/2020).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts.
References
- FAO; IFAD; UNICEF; WFP; WHO. The State of Food Security and Nutrition in the World 2021; FAO: Rome, Italy, 2021; ISBN 9789251343258. [Google Scholar]
- Graham, L.E.; Wilcox, L.W. Algae; Prentice-Hall: Hoboken, NJ, USA, 2000. [Google Scholar]
- Gallardo, T. Marine Algae: General Aspects (Biology, Systematics, Field and Laboratory Techniques). In Marine Algae; CRC Press: Boca Raton, FL, USA, 2014; pp. 1–67. [Google Scholar]
- Mantri, V.A.; Kazi, M.A.; Balar, N.B.; Gupta, V.; Gajaria, T. Concise Review of Green Algal Genus Ulva Linnaeus. J. Appl. Phycol. 2020, 32, 2725–2741. [Google Scholar] [CrossRef]
- Ben-Ari, T.; Neori, A.; Ben-Ezra, D.; Shauli, L.; Odintsov, V.; Shpigel, M. Management of Ulva lactuca as a Biofilter of Mariculture Effluents in IMTA System. Aquaculture 2014, 434, 493–498. [Google Scholar] [CrossRef]
- García-Poza, S.; Morais, T.; Leandro, A.; Cotas, J.; Marques, J.C.; Pereira, L.; Gonçalves, A.M.M. A Comparative Study of the Fatty Acids and Monosaccharides of Wild and Cultivated Ulva sp. J. Mar. Sci. Eng. 2022, 10, 233. [Google Scholar] [CrossRef]
- Zhang, J.; Huo, Y.; Wu, H.; Yu, K.; Kim, J.K.; Yarish, C.; Qin, Y.; Liu, C.; Xu, R.; He, P. The Origin of the Ulva Macroalgal Blooms in the Yellow Sea in 2013. Mar. Pollut. Bull. 2014, 89, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Xia, J.; Zhuang, M.; Zhang, J.; Yu, K.; Zhao, S.; Sun, Y.; Tong, Y.; Xia, L.; Qin, Y.; et al. Controlling the Source of Green Tides in the Yellow Sea: NaClO Treatment of Ulva Attached on Pyropia Aquaculture Rafts. Aquaculture 2021, 535, 736378. [Google Scholar] [CrossRef]
- Liu, J.; Zhuang, M.; Zhao, L.; Liu, Y.; Wen, Q.; Fu, M.; Yu, K.; Zhang, J.; He, P. Taxonomy and Genetic Diversity of Amphipods Living on Ulva lactuca L. from Gouqi Coast, China. Pac. Sci. 2020, 74, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Shi, J.; Gao, S.; Huo, Y.; Cui, J.; Shen, H.; Liu, G.; He, P. Annual Patterns of Macroalgal Blooms in the Yellow Sea during 2007–2017. PLoS ONE 2019, 14, e0210460. [Google Scholar] [CrossRef]
- Abdel-Fattah, A.F.; Edrees, M. Seasonal Changes in the Constituents of Ulva lactuca. Phytochemistry 1973, 12, 481–485. [Google Scholar] [CrossRef]
- Leandro, A.; Pacheco, D.; Cotas, J.; Marques, J.C.; Pereira, L.; Gonçalves, A.M.M. Seaweed’s Bioactive Candidate Compounds to Food Industry and Global Food Security. Life 2020, 10, 140. [Google Scholar] [CrossRef]
- Balar, N.B.; Mantri, V.A. Insights into Life Cycle Patterns, Spore Formation, Induction of Reproduction, Biochemical and Molecular Aspects of Sporulation in Green Algal Genus Ulva: Implications for Commercial Cultivation. J. Appl. Phycol. 2020, 32, 473–484. [Google Scholar] [CrossRef]
- Liu, X.; Blomme, J.; Bogaert, K.A.; D’hondt, S.; Wichard, T.; Deforce, D.; Van Nieuwerburgh, F.; De Clerck, O. Transcriptional Dynamics of Gametogenesis in the Green Seaweed Ulva mutabilis Identifies an RWP-RK Transcription Factor Linked to Reproduction. BMC Plant Biol. 2022, 22, 19. [Google Scholar] [CrossRef]
- Stratmann, J.; Paputsoglu, G.; Oertel, W. Differentiation of Ulva mutabilis (Chlorophyta) Gametangia and Gamete Release Are Controlled by Extracellular Inhibitors 1. J. Phycol. 1996, 32, 1009–1021. [Google Scholar] [CrossRef]
- De Clerck, O.; Kao, S.-M.; Bogaert, K.A.; Blomme, J.; Foflonker, F.; Kwantes, M.; Vancaester, E.; Vanderstraeten, L.; Aydogdu, E.; Boesger, J.; et al. Insights into the Evolution of Multicellularity from the Sea Lettuce Genome. Curr. Biol. 2018, 28, 2921–2933.e5. [Google Scholar] [CrossRef]
- Lüning, K.; Kadel, P.; Pang, S. Control Of Reproduction Rhythmicity by Environmental and Endogenous Signals in Ulva pseudocurvata (Chlorophyta). J. Phycol. 2008, 44, 866–873. [Google Scholar] [CrossRef]
- Katsaros, C.; Weiss, A.; Llangos, I.; Theodorou, I.; Wichard, T. Cell Structure and Microtubule Organisation during Gametogenesis of Ulva mutabilis Føyn (Chlorophyta). Bot. Mar. 2017, 60, 123–135. [Google Scholar] [CrossRef]
- Tan, C.-Y.; Dodd, I.C.; Chen, J.E.; Phang, S.-M.; Chin, C.F.; Yow, Y.-Y.; Ratnayeke, S. Regulation of Algal and Cyanobacterial Auxin Production, Physiology, and Application in Agriculture: An Overview. J. Appl. Phycol. 2021, 33, 2995–3023. [Google Scholar] [CrossRef]
- Jones, B.; Gunnerås, S.A.; Petersson, S.V.; Tarkowski, P.; Graham, N.; May, S.; Dolezal, K.; Sandberg, G.; Ljung, K. Cytokinin Regulation of Auxin Synthesis in Arabidopsis Involves a Homeostatic Feedback Loop Regulated via Auxin and Cytokinin Signal Transduction. Plant Cell 2010, 22, 2956–2969. [Google Scholar] [CrossRef]
- Woodward, A.W.; Bartel, B. Auxin: Regulation, Action, and Interaction. Ann. Bot. 2005, 95, 707–735. [Google Scholar] [CrossRef]
- Žižková, E.; Kubeš, M.; Dobrev, P.I.; Přibyl, P.; Šimura, J.; Zahajská, L.; Záveská Drábková, L.; Novák, O.; Motyka, V. Control of Cytokinin and Auxin Homeostasis in Cyanobacteria and Algae. Ann. Bot. 2017, 119, 151–166. [Google Scholar] [CrossRef]
- Kobayashi, M.; Hirai, N.; Kurimura, Y.; Ohigashi, H.; Tsuji, Y. Abscisic Acid-Dependent Algal Morphogenesis in the Unicellular Green Alga Haematococcus pluvialis; Kluwer Academic Publishers: Alphen aan den Rijn, The Netherlands, 1997; Volume 22. [Google Scholar]
- Nimura, K.; Mizuta, H. Inducible Effects of Abscisic Acid on Sporophyte Discs from Laminaria japonica Areschoug (Laminariales, Phaeophyceae). J. Appl. Phycol. 2002, 14, 159–163. [Google Scholar] [CrossRef]
- Nagata, T.; Takebe, I. Plating of Isolated Tobacco Mesophyll Protoplasts on Agar Medium. Planta 1971, 99, 12–20. [Google Scholar] [CrossRef]
- Hu, Y.; Bao, F.; Li, J. Promotive Effect of Brassinosteroids on Cell Division Involves a Distinct CycD3-induction Pathway in Arabidopsis. Plant J. 2000, 24, 693–701. [Google Scholar] [CrossRef]
- Ikeuchi, M.; Sugimoto, K.; Iwase, A. Plant Callus: Mechanisms of Induction and Repression. Plant Cell 2013, 25, 3159–3173. [Google Scholar] [CrossRef]
- Skoog, F.; Miller, C.O. Chemical Regulation of Growth and Organ Formation in Plant Tissues Cultured in Vitro. Symp. Soc. Exp. Biol. 1957, 11, 118–130. [Google Scholar]
- Chandimali, N.; Park, E.H.; Bak, S.-G.; Lim, H.-J.; Won, Y.-S.; Lee, S.-J. Seaweed Callus Culture: A Comprehensive Review of Current Circumstances and Future Perspectives. Algal Res. 2024, 77, 103376. [Google Scholar] [CrossRef]
- Yokoya, N.S. Apical Callus Formation and Plant Regeneration Controlled by Plant Growth Regulators on Axenic Culture of the Red Alga Gracilariopsis tenuifrons (Gracilariales, Rhodophyta). Phycol. Res. 2000, 48, 133–142. [Google Scholar] [CrossRef]
- Yokoya, N.S.; West, J.A.; Luchi, A.E. Effects of Plant Growth Regulators on Callus Formation, Growth and Regeneration in Axenic Tissue Cultures of Gracilaria tenuistipitata and Gracilaria perplexa (Gracilariales, Rhodophyta). Phycol. Res. 2004, 52, 244–254. [Google Scholar] [CrossRef]
- Kumar, N.; Reddy, M.P. In Vitro Plant Propagation: A Review. J. For. Environ. Sci. 2011, 27, 61–72. [Google Scholar]
- Yoon, J.T.; Soh, W.Y. Developmental Morphology on the Regeneration of Pelvetia siliquosa. Algae 1998, 13, 261–270. [Google Scholar]
- Sulistiani, E.; Soelistyowati, T.; Yani, A. Callus Induction and Filaments Regeneration from Callus of Cottonii Seaweed (Doty) Collected from Natuna Islands, Riau Islands Province Kappaphycus alvarezii. Biotropia 2012, 19, 103–114. [Google Scholar]
- Uji, T.; Nanaumi, D.; Kawagoe, C.; Saga, N.; Miyashita, K. Factors Influencing the Induction of Adventitious Bud and Callus in the Brown Alga Sargassum horneri (Turner) C. Agardh. J. Appl. Phycol. 2016, 28, 2435–2443. [Google Scholar] [CrossRef]
- Muhamad, S.N.S.; Ling, A.P.-K.; Wong, C.-L. Effect of Plant Growth Regulators on Direct Regeneration and Callus Induction from Sargassum polycystum C. Agardh. J. Appl. Phycol. 2018, 30, 3299–3310. [Google Scholar] [CrossRef]
- Tabuchi, K.; Mizuta, H.; Yasui, H. Promotion of Callus Propagation by 5-Aminolevulinic Acid in a Laminaria japonica Sporophyte. Aquac. Res. 2009, 41, 1–10. [Google Scholar] [CrossRef]
- Wichard, T.; Oertel, W. Gametogenesis and Gamete Release of Ulva mutabilis and Ulva lactuca (Chlorophyta): Regulatory Effects and Chemical Characterization of the “Swarming Inhibitor”. J. Phycol. 2010, 46, 248–259. [Google Scholar] [CrossRef]
- Kessler, R.W.; Weiss, A.; Kuegler, S.; Hermes, C.; Wichard, T. Macroalgal–Bacterial Interactions: Role of Dimethylsulfoniopropionate in Microbial Gardening by Ulva (Chlorophyta). Mol. Ecol. 2018, 27, 1808–1819. [Google Scholar] [CrossRef] [PubMed]
- Løvlie, A. Genetic Control of Division Rate and Morphogenesis in Ulva mutabilis Foeyn. Comptes-Rendus Trav. Lab. Carlsberg 1964, 34, 77–168. [Google Scholar]
- Vesty, E.F.; Kessler, R.W.; Wichard, T.; Coates, J.C. Regulation of Gametogenesis and Zoosporogenesis in Ulva linza (Chlorophyta): Comparison with Ulva mutabilis and Potential for Laboratory Culture. Front. Plant Sci. 2015, 6, 126163. [Google Scholar] [CrossRef]
- Dang, H.; Lovell, C.R. Microbial Surface Colonization and Biofilm Development in Marine Environments. Microbiol. Mol. Biol. Rev. 2016, 80, 91–138. [Google Scholar] [CrossRef] [PubMed]
- Egan, S.; Harder, T.; Burke, C.; Steinberg, P.; Kjelleberg, S.; Thomas, T. The Seaweed Holobiont: Understanding Seaweed–Bacteria Interactions. FEMS Microbiol. Rev. 2013, 37, 462–476. [Google Scholar] [CrossRef]
- Hardoim, C.C.P.; Esteves, A.I.S.; Pires, F.R.; Gonçalves, J.M.S.; Cox, C.J.; Xavier, J.R.; Costa, R. Phylogenetically and Spatially Close Marine Sponges Harbour Divergent Bacterial Communities. PLoS ONE 2012, 7, e53029. [Google Scholar] [CrossRef]
- Azam, F.; Malfatti, F. Microbial Structuring of Marine Ecosystems. Nat. Rev. Microbiol. 2007, 5, 782–791. [Google Scholar] [CrossRef] [PubMed]
- Marshall, K.; Joint, I.; Callow, M.E.; Callow, J.A. Effect of Marine Bacterial Isolates on the Growth and Morphology of Axenic Plantlets of the Green Alga Ulva linza. Microb. Ecol. 2006, 52, 302–310. [Google Scholar] [CrossRef] [PubMed]
- Wichard, T.; Charrier, B.; Mineur, F.; Bothwell, J.H.; De Clerck, O.; Coates, J.C. The Green Seaweed Ulva: A Model System to Study Morphogenesis. Front. Plant Sci. 2015, 6, 123849. [Google Scholar] [CrossRef] [PubMed]
- Burke, C.; Steinberg, P.; Rusch, D.; Kjelleberg, S.; Thomas, T. Bacterial Community Assembly Based on Functional Genes Rather than Species. Proc. Natl. Acad. Sci. USA 2011, 108, 14288–14293. [Google Scholar] [CrossRef] [PubMed]
- Alsufyani, T.; Engelen, A.H.; Diekmann, O.E.; Kuegler, S.; Wichard, T. Prevalence and Mechanism of Polyunsaturated Aldehydes Production in the Green Tide Forming Macroalgal Genus Ulva (Ulvales, Chlorophyta). Chem. Phys. Lipids 2014, 183, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Føyn, B. Über Die Sexualität Und Den Generationswechsel von Ulva mutabilis. Arch. Protistenkd. 1958, 102, 473–480. [Google Scholar]
- Califano, G.; Kwantes, M.; Abreu, M.H.; Costa, R.; Wichard, T. Cultivating the Macroalgal Holobiont: Effects of Integrated Multi-Trophic Aquaculture on the Microbiome of Ulva rigida (Chlorophyta). Front. Mar. Sci. 2020, 7, 52. [Google Scholar] [CrossRef]
- van der Loos, L.M.; D’hondt, S.; Willems, A.; De Clerck, O. Characterizing Algal Microbiomes Using Long-Read Nanopore Sequencing. Algal Res. 2021, 59, 102456. [Google Scholar] [CrossRef]
- Ghaderiardakani, F.; Langhans, L.; Kurbel, V.B.; Fenizia, S.; Wichard, T. Metabolite Profiling Reveals Insights into the Species-Dependent Cold Stress Response of the Green Seaweed Holobiont Ulva (Chlorophyta). Environ. Exp. Bot. 2022, 200, 104913. [Google Scholar] [CrossRef]
- Mcgovern, M. Domestication of the Green Seaweed Ulva Ohnoi Aka “sea Lettuce” for Biofloc Effluent Bioremediation. Master’s Thesis, Universidade do Algarve, Faro, Portugal, 2020. [Google Scholar]
- Duan, D.; Xu, L.; Fei, X.; Xu, H. Marine Organisms Attached to Seaweed Surfaces in Jiaozhou Bay, China. World J. Microbiol. Biotechnol. 1995, 11, 351–352. [Google Scholar] [CrossRef]
- Nakanishi, K.; Nishijima, M.; Nomoto, A.M.; Yamazaki, A.; Saga, N. Requisite Morphologic Interaction for Attachment between Ulva pertusa (Chlorophyta) and Symbiotic Bacteria. Mar. Biotechnol. 1999, 1, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, Y.; Suzuki, M.; Kasai, H.; Shizuri, Y.; Harayama, S. Isolation and Phylogenetic Characterization of Bacteria Capable of Inducing Differentiation in the Green Alga Monostroma oxyspermum. Environ. Microbiol. 2003, 5, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.P.; Mantri, V.A.; Reddy, C.R.K.; Jha, B. Isolation of Seaweed-Associated Bacteria and Their Morphogenesis-Inducing Capability in Axenic Cultures of the Green Alga Ulva fasciata. Aquat. Biol. 2011, 12, 13–21. [Google Scholar] [CrossRef]
- Provasoli, L.; Pintner, I.J. Bacteria Induced Polymorphism in an Axenic Laboratory Strain of Ulva lactuca (Chlorophyceae). J. Phycol. 1980, 16, 196–201. [Google Scholar] [CrossRef]
- Fjeld, A. Genetic Control of Cellular Differentiation in Ulva mutabilis. Gene Effects in Early Development. Dev. Biol. 1972, 28, 326–343. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, G.L.; Tait, K.; Taylor, A.; Brownlee, C.; Joint, I. Acyl-homoserine Lactones Modulate the Settlement Rate of Zoospores of the Marine Alga Ulva intestinalis via a Novel Chemokinetic Mechanism. Plant Cell Environ. 2006, 29, 608–618. [Google Scholar] [CrossRef] [PubMed]
- Nakanishi, K.; Nishijima, M.; Nishimura, M.; Kuwano, K.; Saga, N. Bacteria That Induce Morphogenesis in Ulva pertusa (Chlorophyta) Grown Under Axenic Conditions. J. Phycol. 1996, 32, 479–482. [Google Scholar] [CrossRef]
- Spoerner, M.; Wichard, T.; Bachhuber, T.; Stratmann, J.; Oertel, W. Growth and Thallus Morphogenesis of Ulva mutabilis (Chlorophyta) Depends on A Combination of Two Bacterial Species Excreting Regulatory Factors. J. Phycol. 2012, 48, 1433–1447. [Google Scholar] [CrossRef] [PubMed]
- Ghaderiardakani, F.; Coates, J.C.; Wichard, T. Bacteria-Induced Morphogenesis of Ulva Intestinalis and Ulva mutabilis (Chlorophyta): A Contribution to the Lottery Theory. FEMS Microbiol. Ecol. 2017, 93, fix094. [Google Scholar] [CrossRef]
- Grueneberg, J.; Engelen, A.H.; Costa, R.; Wichard, T. Macroalgal Morphogenesis Induced by Waterborne Compounds and Bacteria in Coastal Seawater. PLoS ONE 2016, 11, e0146307. [Google Scholar] [CrossRef]
- Joint, I.; Tait, K.; Wheeler, G. Cross-Kingdom Signalling: Exploitation of Bacterial Quorum Sensing Molecules by the Green Seaweed Ulva. Philos. Trans. R. Soc. B Biol. Sci. 2007, 362, 1223–1233. [Google Scholar] [CrossRef] [PubMed]
- Ingle, K.N.; Polikovsky, M.; Chemodanov, A.; Golberg, A. Marine Integrated Pest Management (MIPM) Approach for Sustainable Seagriculture. Algal Res. 2018, 29, 223–232. [Google Scholar] [CrossRef]
- Goecke, F.; Labes, A.; Wiese, J.; Imhoff, J. Chemical Interactions between Marine Macroalgae and Bacteria. Mar. Ecol. Prog. Ser. 2010, 409, 267–299. [Google Scholar] [CrossRef]
- Kamermans, P.; Malta, E.J.; Verschuure, J.M.; Schrijvers, L.; Lentz, L.F.; Lien, A.T.A. Effect of Grazing by Isopods and Amphipods on Growth of Ulva spp. (Chlorophyta). Aquat. Ecol. 2002, 36, 425–433. [Google Scholar] [CrossRef]
- Simon, C.; McHale, M.; Sulpice, R. Applications of Ulva Biomass and Strategies to Improve Its Yield and Composition: A Perspective for Ulva Aquaculture. Biology 2022, 11, 1593. [Google Scholar] [CrossRef] [PubMed]
- Jewett, M.C.; Hofmann, G.; Nielsen, J. Fungal Metabolite Analysis in Genomics and Phenomics. Curr. Opin. Biotechnol. 2006, 17, 191–197. [Google Scholar] [CrossRef]
- Roessner, U.; Bowne, J. What Is Metabolomics All About? Biotechniques 2009, 46, 363–365. [Google Scholar] [CrossRef]
- Alsufyani, T.; Weiss, A.; Wichard, T. Time Course Exo-Metabolomic Profiling in the Green Marine Macroalga Ulva (Chlorophyta) for Identification of Growth Phase-Dependent Biomarkers. Mar. Drugs 2017, 15, 14. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Wang, Y.; Hu, C.; Sun, X.; Li, Y.; Xu, N. Dynamic Metabolic Profiles of the Marine Macroalga Ulva prolifera during Fragmentation-Induced Proliferation. PLoS ONE 2019, 14, e0214491. [Google Scholar] [CrossRef]
- Lee, T.-M.; Liu, C.-H. Correlation of Decreased Calcium Contents with Proline Accumulation in the Marine Green Macroalga Ulva fasciata Exposed to Elevated NaCl Contents in Seawater. J. Exp. Bot. 1999, 50, 1855–1862. [Google Scholar] [CrossRef]
- Dawson, H.M.; Heal, K.R.; Boysen, A.K.; Carlson, L.T.; Ingalls, A.E.; Young, J.N. Potential of Temperature- and Salinity-Driven Shifts in Diatom Compatible Solute Concentrations to Impact Biogeochemical Cycling within Sea Ice. Elem. Sci. Anthr. 2020, 8, 25. [Google Scholar] [CrossRef]
- Gawryluk, R.M.R.; Tikhonenkov, D.V.; Hehenberger, E.; Husnik, F.; Mylnikov, A.P.; Keeling, P.J. Non-Photosynthetic Predators Are Sister to Red Algae. Nature 2019, 572, 240–243. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.-M.; Sun, Y.-Y.; Ye, X.-Y.; Li, Z.-G. Signaling Role of Glutamate in Plants. Front. Plant Sci. 2020, 10, 1743. [Google Scholar] [CrossRef] [PubMed]
- Van Alstyne, K.L.; Puglisi, M.P. DMSP in Marine Macroalgae and Macroinvertebrates: Distribution, Function, and Ecological Impacts. Aquat. Sci. 2007, 69, 394–402. [Google Scholar] [CrossRef]
- Chávez-Sánchez, T.; Piñón-Gimate, A.; Serviere-Zaragoza, E.; López-Bautista, J.M.; Casas-Valdez, M. Ulva Blooms in the Southwestern Gulf of California: Reproduction and Biomass. Estuar. Coast. Shelf Sci. 2018, 200, 202–211. [Google Scholar] [CrossRef]
- Sousa, A.I.; Martins, I.; Lillebø, A.I.; Flindt, M.R.; Pardal, M.A. Influence of Salinity, Nutrients and Light on the Germination and Growth of Enteromorpha sp. Spores. J. Exp. Mar. Biol. Ecol. 2007, 341, 142–150. [Google Scholar] [CrossRef]
- Han, T.; Kang, S.-H.; Park, J.-S.; Lee, H.-K.; Brown, M.T. Physiological Responses of Ulva pertusa and U. armoricana to Copper Exposure. Aquat. Toxicol. 2008, 86, 176–184. [Google Scholar] [CrossRef] [PubMed]
- Carl, C.; de Nys, R.; Lawton, R.J.; Paul, N.A. Methods for the Induction of Reproduction in a Tropical Species of Filamentous Ulva. PLoS ONE 2014, 9, e97396. [Google Scholar] [CrossRef] [PubMed]
- Pettett, P. Preliminary Investigation into the Induction of Reproduction in Ulva spp. in Southeast Queens-land for Mass Cultivation Purposes. Master’s Thesis, University of the Sunshine Coast, Sippy Downs, QLD, Australia, 2009; pp. 2–71. [Google Scholar]
- Kalita, T.L.; Titlyanov, E.A. Influence of Temperature on the Infradian Growth Rhythm in Ulva lactuca (Chlorophyta). Eur. J. Phycol. 2013, 48, 210–220. [Google Scholar] [CrossRef]
- Niesenbaum, R.A. The Ecology of Sporulation by the Macroalga Ulva lactuca L. (Chlorophyceae). Aquat. Bot. 1988, 32, 155–166. [Google Scholar] [CrossRef]
- Gao, G.; Clare, A.S.; Rose, C.; Caldwell, G.S. Reproductive Sterility Increases the Capacity to Exploit the Green Seaweed Ulva rigida for Commercial Applications. Algal Res. 2017, 24, 64–71. [Google Scholar] [CrossRef]
- Pedersen, M.F. Transient Ammonium Uptake in the Macroalga Ulva lactuca (Chlorophyta): Nature, Regulation, And The Consequences for Choice of Measuring Technique. J. Phycol. 1994, 30, 980–986. [Google Scholar] [CrossRef]
- Conway, H.L.; Harrison, P.J. Marine Diatoms Grown in Chemostats under Silicate or Ammonium Limitation. IV. Transient Response of Chaetoceros debilis, Skeletonema costatum, and Thalassiosira gravida to a Single Addition of the Limiting Nutrient. Mar. Biol. 1977, 43, 33–43. [Google Scholar] [CrossRef]
- Kalita, T.L.; Titlyanov, E.A. The Effect of Temperature on Infradian Rhythms of Reproduction in Ulva fenestrata Postels et Ruprecht, 1840 (Chlorophyta: Ulvales). Russ. J. Mar. Biol. 2011, 37, 52–61. [Google Scholar] [CrossRef]
- Hiraoka, M.; Enomoto, S. The Induction of Reproductive Cell Formation of Ulva pertusa Kjellman (Ulvales, Ulvophyceae). Phycol. Res. 1998, 46, 199–203. [Google Scholar] [CrossRef]
- Gao, S.; Chen, X.; Yi, Q.; Wang, G.; Pan, G.; Lin, A.; Peng, G. A Strategy for the Proliferation of Ulva prolifera, Main Causative Species of Green Tides, with Formation of Sporangia by Fragmentation. PLoS ONE 2010, 5, e8571. [Google Scholar] [CrossRef]
- Gao, G. Developing Systems for the Commercial Culture of Ulva Species in the UK. Ph.D. Thesis, Newcastle University, Newcastle, UK, 2016. [Google Scholar]
- Zhao, S.; Xia, Z.; Liu, J.; Sun, J.; Zhang, J.; He, P. Morphology, Growth, and Photosynthesis of Ulva prolifera O.F. Müller (Chlorophyta, Ulvophyceae) Gametophytes, the Dominant Green Tide Species in the Southern Yellow Sea. J. Sea Res. 2023, 193, 102375. [Google Scholar] [CrossRef]
- Naylor, R.L.; Hardy, R.W.; Buschmann, A.H.; Bush, S.R.; Cao, L.; Klinger, D.H.; Little, D.C.; Lubchenco, J.; Shumway, S.E.; Troell, M. A 20-Year Retrospective Review of Global Aquaculture. Nature 2021, 591, 551–563. [Google Scholar] [CrossRef] [PubMed]
- Dinesh Kumar, S.; Satish, L.; Dhanya, N.; Malar Vizhi, J.; Nadukkattu Nayagi, N.; Gopala Krishnan, S.; Ganesan, M. Tank Cultivation of Edible Seaweeds: An Overview of the Indian Perspective for Opportunities and Challenges. Biomass Convers. Biorefin 2023. [Google Scholar] [CrossRef]
- Mullikin, R.K.; Rorrer, G.L. A Tubular Recycle Photobioreactor for Macroalgal Suspension Cultures. In BioHydrogen; Springer: Boston, MA, USA, 1998; pp. 403–414. [Google Scholar]
- Liu, Q.; Yu, R.-C.; Yan, T.; Zhang, Q.-C.; Zhou, M.-J. Laboratory Study on the Life History of Bloom-Forming Ulva prolifera in the Yellow Sea. Estuar. Coast. Shelf Sci. 2015, 163, 82–88. [Google Scholar] [CrossRef]
- Zhang, X.; Xu, D.; Mao, Y.; Li, Y.; Xue, S.; Zou, J.; Lian, W.; Liang, C.; Zhuang, Z.; Wang, Q.; et al. Settlement of Vegetative Fragments of Ulva prolifera Confirmed as an Important Seed Source for Succession of a Large-scale Green Tide Bloom. Limnol. Oceanogr. 2011, 56, 233–242. [Google Scholar] [CrossRef]
- de Góes, H.G.; Reis, R.P. An Initial Comparison of Tubular Netting versus Tie–Tie Methods of Cultivation for Kappaphycus alvarezii (Rhodophyta, Solieriaceae) on the South Coast of Rio de Janeiro State, Brazil. J. Appl. Phycol. 2011, 23, 607–613. [Google Scholar] [CrossRef]
- Redmond, S.; Green, L.; Yarish, C.; Kim, J.; Neefus, C. New England Seaweed Culture Handbook; University of Connecticut: Groton, CT, USA, 2014. [Google Scholar]
- Zhang, J.; Kim, J.K.; Yarish, C.; He, P. The Expansion of Ulva prolifera O.F. Müller Macroalgal Blooms in the Yellow Sea, PR China, through Asexual Reproduction. Mar. Pollut. Bull. 2016, 104, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Lin, A.; Shen, S.; Wang, J.; Yan, B. Reproduction Diversity of Enteromorpha prolifera. J. Integr. Plant Biol. 2008, 50, 622–629. [Google Scholar] [CrossRef] [PubMed]
- Gao, G.; Clare, A.S.; Rose, C.; Caldwell, G.S. Intrinsic and Extrinsic Control of Reproduction in the Green Tide-Forming Alga, Ulva rigida. Environ. Exp. Bot. 2017, 139, 14–22. [Google Scholar] [CrossRef]
- Steinhagen, S.; Larsson, K.; Olsson, J.; Albers, E.; Undeland, I.; Pavia, H.; Toth, G.B. Closed Life-Cycle Aquaculture of Sea Lettuce (Ulva fenestrata): Performance and Biochemical Profile Differ in Early Developmental Stages. Front. Mar. Sci. 2022, 9, 942679. [Google Scholar] [CrossRef]
- Morales-Rubio, M.E.; Espinosa-Leal, C.; Garza-Padrón, R.A. Cultivo de Tejidos Vegetales y Su Aplicación En Productos Naturales. In Investigación en Plantas de Importancia Médica; OmniaScience: Barcelona, Spain, 2016; pp. 351–410. [Google Scholar]
- Espinosa-Leal, C.A.; Puente-Garza, C.A.; García-Lara, S. In Vitro Plant Tissue Culture: Means for Production of Biological Active Compounds. Planta 2018, 248, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Califano, G.; Wichard, T. Preparation of Axenic Cultures in Ulva (Chlorophyta). In Protocols for Macroalgae Research; CRC: Boca Raton, FL, USA, 2018; pp. 159–171. [Google Scholar]
- Wichard, T. Exploring Bacteria-Induced Growth and Morphogenesis in the Green Macroalga Order Ulvales (Chlorophyta). Front. Plant Sci. 2015, 6, 123850. [Google Scholar] [CrossRef] [PubMed]
- Chemodanov, A.; Robin, A.; Golberg, A. Design of Marine Macroalgae Photobioreactor Integrated into Building to Support Seagriculture for Biorefinery and Bioeconomy. Bioresour. Technol. 2017, 241, 1084–1093. [Google Scholar] [CrossRef]
- Gore, M. (Ed.) Spectrophotometry and Spectrofluorimetry; Oxford University Press: Oxford, UK, 2000; ISBN 9780199638130. [Google Scholar]
- Vo, K. 2.1.5: Spectrophotometry. Available online: https://chem.libretexts.org/Bookshelves/Physical_and_Theoretical_Chemistry_Textbook_Maps/Supplemental_Modules_(Physical_and_Theoretical_Chemistry)/Kinetics/02%3A_Reaction_Rates/2.01%3A_Experimental_Determination_of_Kinetics/2.1.05%3A_Spectrophotometry (accessed on 15 March 2024).
- Lähteenmäki-Uutela, A.; Rahikainen, M.; Camarena-Gómez, M.T.; Piiparinen, J.; Spilling, K.; Yang, B. European Union Legislation on Macroalgae Products. Aquac. Int. 2021, 29, 487–509. [Google Scholar] [CrossRef]
- FSSC. Food Safety System Certification 22000 Guidance Document: ISO 22000 Interpretation; FSSC: Gorinchem, The Netherlands, 2019. [Google Scholar]
- SGS. Entendendo a Norma de Certificação de Sistemas de Segurança de Alimentos FSSC 22000 Guia Técnico Sobre Os. Desafios, Impactos e Oportunidades Da FSSC 22000; SGS: Geneva, Switzerland, 2014. [Google Scholar]
- ABNT. Sistemas de Gestão Ambiental-Requisitos Com. Orientações Para. Uso Environmental Management Systems-Requirements with Guidance for Use; ABNT: Rio de Janeiro and São Paulo, Brazil, 2015. [Google Scholar]
- Ociepa-Kubicka, A.; Deska, I.; Ociepa, E. Organizations towards the Evaluation of Environmental Management Tools ISO 14001 and EMAS. Energies 2021, 14, 4870. [Google Scholar] [CrossRef]
- García-Poza, S.; Leandro, A.; Cotas, C.; Cotas, J.; Marques, J.C.; Pereira, L.; Gonçalves, A.M.M. The Evolution Road of Seaweed Aquaculture: Cultivation Technologies and the Industry 4.0. Int. J. Environ. Res. Public Health 2020, 17, 6528. [Google Scholar] [CrossRef] [PubMed]
- Pereira, L. Algal Polysaccharides. In Functional Ingredients from Algae for Foods and Nutraceuticals; Elsevier: Amsterdam, The Netherlands, 2023; pp. 151–212. [Google Scholar]
- Li, C.; Tang, T.; Du, Y.; Jiang, L.; Yao, Z.; Ning, L.; Zhu, B. Ulvan and Ulva Oligosaccharides: A Systematic Review of Structure, Preparation, Biological Activities and Applications. Bioresour. Bioprocess. 2023, 10, 66. [Google Scholar] [CrossRef]
- Kraan, S. Mass-Cultivation of Carbohydrate Rich Macroalgae, a Possible Solution for Sustainable Biofuel Production. Mitig. Adapt. Strateg. Glob. Change 2013, 18, 27–46. [Google Scholar] [CrossRef]
- Lahaye, M.; Robic, A. Structure and Functional Properties of Ulvan, a Polysaccharide from Green Seaweeds. Biomacromolecules 2007, 8, 1765–1774. [Google Scholar] [CrossRef] [PubMed]
- Alves, A.; Sousa, R.A.; Reis, R.L. A Practical Perspective on Ulvan Extracted from Green Algae. J. Appl. Phycol. 2013, 25, 407–424. [Google Scholar] [CrossRef]
- Webster, E.A.; Gadd, G.M. Cadmium Replaces Calcium in the Cell Wall of Ulva lactuca. BioMetals 1996, 9, 241–244. [Google Scholar] [CrossRef]
- Bocanegra, A.; Bastida, S.; Benedí, J.; Ródenas, S.; Sánchez-Muniz, F.J. Characteristics and Nutritional and Cardiovascular-Health Properties of Seaweeds. J. Med. Food 2009, 12, 236–258. [Google Scholar] [CrossRef] [PubMed]
- Schijf, J.; Ebling, A.M. Investigation of the Ionic Strength Dependence of Ulva lactuca Acid Functional Group pKas by Manual Alkalimetric Titrations. Environ. Sci. Technol. 2010, 44, 1644–1649. [Google Scholar] [CrossRef]
- Kidgell, J.T.; Magnusson, M.; de Nys, R.; Glasson, C.R.K. Ulvan: A Systematic Review of Extraction, Composition and Function. Algal Res. 2019, 39, 101422. [Google Scholar] [CrossRef]
- Putra, N.R.; Fajriah, S.; Qomariyah, L.; Dewi, A.S.; Rizkiyah, D.N.; Irianto, I.; Rusmin, D.; Melati, M.; Trisnawati, N.W.; Darwati, I.; et al. Exploring the Potential of Ulva lactuca: Emerging Extraction Methods, Bioactive Compounds, and Health Applications—A Perspective Review. S. Afr. J. Chem. Eng. 2024, 47, 233–245. [Google Scholar] [CrossRef]
- Aronoff, S. Photosynthesis. Bot. Rev. 1957, 23, 65–107. [Google Scholar] [CrossRef]
- Martins, M.; Oliveira, R.; Coutinho, J.A.P.; Faustino, M.A.F.; Neves, M.G.P.M.S.; Pinto, D.C.G.A.; Ventura, S.P.M. Recovery of Pigments from Ulva rigida. Sep. Purif. Technol. 2021, 255, 117723. [Google Scholar] [CrossRef]
- Pereira, L.; Kraan, S. Pigments and Minor Compounds in Algae. In Functional Ingredients from Algae for Foods and Nutraceuticals; Elsevier: Amsterdam, The Netherlands, 2023; pp. 397–461. [Google Scholar]
- Kazir, M.; Abuhassira, Y.; Robin, A.; Nahor, O.; Luo, J.; Israel, A.; Golberg, A.; Livney, Y.D. Extraction of Proteins from Two Marine Macroalgae, Ulva sp. and Gracilaria sp., for Food Application, and Evaluating Digestibility, Amino Acid Composition and Antioxidant Properties of the Protein Concentrates. Food Hydrocoll. 2019, 87, 194–203. [Google Scholar] [CrossRef]
- Lordan, S.; Ross, R.P.; Stanton, C. Marine Bioactives as Functional Food Ingredients: Potential to Reduce the Incidence of Chronic Diseases. Mar. Drugs 2011, 9, 1056–1100. [Google Scholar] [CrossRef] [PubMed]
- Lanfer-Marquez, U.M.; Barros, R.M.C.; Sinnecker, P. Antioxidant Activity of Chlorophylls and Their Derivatives. Food Res. Int. 2005, 38, 885–891. [Google Scholar] [CrossRef]
- Luo, L.; Lai, X.; Chen, B.; Lin, L.; Fang, L.; Tam, N.F.Y.; Luan, T. Chlorophyll Catalyse the Photo-Transformation of Carcinogenic Benzo[a]Pyrene in Water. Sci. Rep. 2015, 5, 12776. [Google Scholar] [CrossRef]
- Sudakin, D.L. Dietary Aflatoxin Exposure and Chemoprevention of Cancer: A Clinical Review. J. Toxicol. Clin. Toxicol. 2003, 41, 195–204. [Google Scholar] [CrossRef]
- Diogo, P.; Mota, M.; Fernandes, C.; Sequeira, D.; Palma, P.; Caramelo, F.; Neves, M.G.P.M.S.; Faustino, M.A.F.; Gonçalves, T.; Santos, J.M. Is the Chlorophyll Derivative Zn(II)e 6 Me a Good Photosensitizer to Be Used in Root Canal Disinfection? Photodiagn. Photodyn. Ther. 2018, 22, 205–211. [Google Scholar] [CrossRef]
- Pham, V.T.; Dold, S.; Rehman, A.; Bird, J.K.; Steinert, R.E. Vitamins, the Gut Microbiome and Gastrointestinal Health in Humans. Nutr. Res. 2021, 95, 35–53. [Google Scholar] [CrossRef]
- Madhavan, J.; Chandrasekharan, S.; Priya, M.; Godavarthi, A. Modulatory Effect of Carotenoid Supplement Constituting Lutein and Zeaxanthin (10:1) on Anti-Oxidant Enzymes and Macular Pigments Level in Rats. Pharmacogn. Mag. 2018, 14, 268. [Google Scholar] [CrossRef]
- Cotas, J. Marine Phenolics. In Functional Ingredients from Algae for Foods and Nutraceuticals; Elsevier: Amsterdam, The Netherlands, 2023; pp. 213–245. [Google Scholar]
- Cotas, J.; Leandro, A.; Monteiro, P.; Pacheco, D.; Figueirinha, A.; Gonçalves, A.M.M.; da Silva, G.J.; Pereira, L. Seaweed Phenolics: From Extraction to Applications. Mar. Drugs 2020, 18, 384. [Google Scholar] [CrossRef] [PubMed]
- Santos, S.A.O.; Félix, R.; Pais, A.C.S.; Rocha, S.M.; Silvestre, A.J.D. The Quest for Phenolic Compounds from Macroalgae: A Review of Extraction and Identification Methodologies. Biomolecules 2019, 9, 847. [Google Scholar] [CrossRef]
- Liu, M.; Hansen, P.E.; Lin, X. Bromophenols in Marine Algae and Their Bioactivities. Mar. Drugs 2011, 9, 1273–1292. [Google Scholar] [CrossRef] [PubMed]
- Stengel, D.B.; Connan, S.; Popper, Z.A. Algal Chemodiversity and Bioactivity: Sources of Natural Variability and Implications for Commercial Application. Biotechnol. Adv. 2011, 29, 483–501. [Google Scholar] [CrossRef] [PubMed]
- Bilal Hussain, M.; Hassan, S.; Waheed, M.; Javed, A.; Adil Farooq, M.; Tahir, A. Bioavailability and Metabolic Pathway of Phenolic Compounds. In Plant Physiological Aspects of Phenolic Compounds; IntechOpen: London, UK, 2019. [Google Scholar]
- Mukherjee, P.K. Bioactive Phytocomponents and Their Analysis. In Quality Control and Evaluation of Herbal Drugs; Elsevier: Amsterdam, The Netherlands, 2019; pp. 237–328. ISBN 9780128133743. [Google Scholar]
- Konuk, H.B.; Ergüden, B. Phenolic –OH Group Is Crucial for the Antifungal Activity of Terpenoids via Disruption of Cell Membrane Integrity. Folia Microbiol. 2020, 65, 775–783. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Inoue, M.; Uzawa, Y.; Kawamura, Y. Anti-tumorigenic Components of a Sea Weed, Eeteromorpha clathrata. BioFactors 2004, 22, 107–110. [Google Scholar] [CrossRef] [PubMed]
- Khanavi, M.; Gheidarloo, R.; Sadati, N.; Shams Ardekani, M.R.; Bagher Nabavi, S.M.; Tavajohi, S.; Ostad, S.N. Cytotoxicity of Fucosterol Containing Fraction of Marine Algae against Breast and Colon Carcinoma Cell Line. Pharmacogn. Mag. 2012, 8, 60–64. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Yang, C.; Lin, G.; Chen, Y.; Miao, S.; Liu, B.; Zhao, C. Antidiabetic Potential of Green Seaweed Enteromorpha prolifera Flavonoids Regulating Insulin Signaling Pathway and Gut Microbiota in Type 2 Diabetic Mice. J. Food Sci. 2019, 84, 165–173. [Google Scholar] [CrossRef]
- Lin, G.; Liu, X.; Yan, X.; Liu, D.; Yang, C.; Liu, B.; Huang, Y.; Zhao, C. Role of Green Macroalgae Enteromorpha prolifera Polyphenols in the Modulation of Gene Expression and Intestinal Microflora Profiles in Type 2 Diabetic Mice. Int. J. Mol. Sci. 2018, 20, 25. [Google Scholar] [CrossRef]
- Sabeena Farvin, K.H.; Jacobsen, C. Phenolic Compounds and Antioxidant Activities of Selected Species of Seaweeds from Danish Coast. Food Chem. 2013, 138, 1670–1681. [Google Scholar] [CrossRef] [PubMed]
- Aslan, E.; Aksu, A.; Korkmaz, N.E.; Taskin, O.S.; Caglar, N.B. Monitoring the Antioxidant Activities by Extracting the Polyphenolic Contents of Algae Collected from the Bosphorus. Mar. Pollut. Bull. 2019, 141, 313–317. [Google Scholar] [CrossRef] [PubMed]
- Valério Filho, A.; Santana, L.R.; Motta, N.G.; Passos, L.F.; lnes Wolke, S.; Mansilla, A.; Astorga-España, M.S.; Becker, E.M.; de Pereira, C.M.P.; Carreno, N.L.V. Extraction of Fatty Acids and Cellulose from the Biomass of Algae Durvillaea antarctica and Ulva lactuca: An Alternative for Biorefineries. Algal Res. 2023, 71, 103084. [Google Scholar] [CrossRef]
- Pereira, T.; Horta, A.; Barroso, S.; Mendes, S.; Gil, M.M. Study of the Seasonal Variations of the Fatty Acid Profiles of Selected Macroalgae. Molecules 2021, 26, 5807. [Google Scholar] [CrossRef] [PubMed]
- Fannin, K.F.; Srivastava, V.; Mensinger, J.; Chynoweth, D. Marine Biomass Program: Anaerobic Digestion Systems Development and Stability Study. Final Report; Gas Research Institute: Chicago, IL, USA, 1982. [Google Scholar]
- Bleakley, S.; Hayes, M. Algal Proteins: Extraction, Application, and Challenges Concerning Production. Foods 2017, 6, 33. [Google Scholar] [CrossRef] [PubMed]
- Shuuluka, D.; Bolton, J.J.; Anderson, R.J. Protein Content, Amino Acid Composition and Nitrogen-to-Protein Conversion Factors of Ulva rigida and Ulva capensis from Natural Populations and Ulva lactuca from an Aquaculture System, in South Africa. J. Appl. Phycol. 2013, 25, 677–685. [Google Scholar] [CrossRef]
- Fleurence, J.; Le Coeur, C.; Mabeau, S.; Maurice, M.; Landrein, A. Comparison of Different Extractive Procedures for Proteins from the Edible Seaweeds Ulva rigida and Ulva rotundata. J. Appl. Phycol. 1995, 7, 577–582. [Google Scholar] [CrossRef]
- Yaich, H.; Garna, H.; Besbes, S.; Paquot, M.; Blecker, C.; Attia, H. Chemical Composition and Functional Properties of Ulva lactuca Seaweed Collected in Tunisia. Food Chem. 2011, 128, 895–901. [Google Scholar] [CrossRef]
- FAO. Global Seaweeds and Microalgae Production, 1950–2019; Food and Agricultural Organization of the United Nations: Rome, Italy, 2021; 172p. [Google Scholar]
- Dan, A.; Hiraoka, M.; Ohno, M.; Critchley, A.T. Observations on the Effect of Salinity and Photon Fluence Rate on the Induction of Sporulation and Rhizoid Formation in the Green Alga Enteromorpha prolifera (Muller) J. Agardh (Chlorophyta, Ulvales). Fish. Sci. 2002, 68, 1182–1188. [Google Scholar] [CrossRef]
- Hiraoka, M.; Oka, N. Tank Cultivation of Ulva Prolifera in Deep Seawater Using a New “Germling Cluster” Method. J. Appl. Phycol. 2008, 20, 97–102. [Google Scholar] [CrossRef]
- Neori, A.; Msuya, F.E.; Shauli, L.; Schuenhoff, A.; Kopel, F.; Shpigel, M. A Novel Three-Stage Seaweed (Ulva lactuca) Biofilter Design for Integrated Mariculture. J. Appl. Phycol. 2003, 15, 543–553. [Google Scholar] [CrossRef]
- Al-Hafedh, Y.S.; Alam, A.; Buschmann, A.H. Bioremediation Potential, Growth and Biomass Yield of the Green Seaweed, Ulva lactuca in an Integrated Marine Aquaculture System at the Red Sea Coast of Saudi Arabia at Different Stocking Densities and Effluent Flow Rates. Rev. Aquac. 2015, 7, 161–171. [Google Scholar] [CrossRef]
- Robertson-Andersson, D.V.; Potgieter, M.; Hansen, J.; Bolton, J.J.; Troell, M.; Anderson, R.J.; Halling, C.; Probyn, T. Integrated Seaweed Cultivation on an Abalone Farm in South Africa. J. Appl. Phycol. 2008, 20, 579–595. [Google Scholar] [CrossRef]
- Bruhn, A.; Dahl, J.; Nielsen, H.B.; Nikolaisen, L.; Rasmussen, M.B.; Markager, S.; Olesen, B.; Arias, C.; Jensen, P.D. Bioenergy Potential of Ulva lactuca: Biomass Yield, Methane Production and Combustion. Bioresour. Technol. 2011, 102, 2595–2604. [Google Scholar] [CrossRef] [PubMed]
- Mata, L.; Magnusson, M.; Paul, N.A.; de Nys, R. The Intensive Land-Based Production of the Green Seaweeds Derbesia tenuissima and Ulva ohnoi: Biomass and Bioproducts. J. Appl. Phycol. 2016, 28, 365–375. [Google Scholar] [CrossRef]
- Uribe, E.; Vega-Gálvez, A.; García, V.; Pastén, A.; López, J.; Goñi, G. Effect of Different Drying Methods on Phytochemical Content and Amino Acid and Fatty Acid Profiles of the Green Seaweed, Ulva spp. J. Appl. Phycol. 2019, 31, 1967–1979. [Google Scholar] [CrossRef]
- Cherry, P.; O’Hara, C.; Magee, P.J.; McSorley, E.M.; Allsopp, P.J. Risks and Benefits of Consuming Edible Seaweeds. Nutr. Rev. 2019, 77, 307–329. [Google Scholar] [CrossRef] [PubMed]
- Ripol, A.; Cardoso, C.; Afonso, C.; Varela, J.; Quental-Ferreira, H.; Pousão-Ferreira, P.; Bandarra, N.M. Composition, Anti-Inflammatory Activity, and Bioaccessibility of Green Seaweeds from Fish Pond Aquaculture. Nat. Prod. Commun. 2018, 13, 603–608. [Google Scholar] [CrossRef]
- MacArtain, P.; Gill, C.I.R.; Brooks, M.; Campbell, R.; Rowland, I.R. Nutritional Value of Edible Seaweeds. Nutr. Rev. 2008, 65, 535–543. [Google Scholar] [CrossRef]
- Yu-Qing, T.; Mahmood, K.; Shehzadi, R.; Ashraf, M.F. Ulva lactuca and Its Polysaccharides: Food and Biomedical Aspects. J. Biol. 2016, 6, 140–151. [Google Scholar]
- Hall, A.C.; Fairclough, A.C.; Mahadevan, K.; Paxman, J.R. Ascophyllum Nodosum Enriched Bread Reduces Subsequent Energy Intake with No Effect on Post-Prandial Glucose and Cholesterol in Healthy, Overweight Males. A Pilot Study. Appetite 2012, 58, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Abudabos, A.M.; Okab, A.B.; Aljumaah, R.S.; Samara, E.M.; Abdoun, K.A.; Al-Haidary, A.A. Nutritional Value of Green Seaweed (Ulva lactuca) for Broiler Chickens. Ital. J. Anim. Sci. 2013, 12, e28. [Google Scholar] [CrossRef]
- Ventura, M.R.; Castañón, J.I.R. The Nutritive Value of Seaweed (Ulva lactuca) for Goats. Small Rumin. Res. 1998, 29, 325–327. [Google Scholar] [CrossRef]
- Cabrita, A.R.J.; Correia, A.; Rodrigues, A.R.; Cortez, P.P.; Vilanova, M.; Fonseca, A.J.M. Assessing In Vivo Digestibility and Effects on Immune System of Sheep Fed Alfalfa Hay Supplemented with a Fixed Amount of Ulva rigida and Gracilaria vermiculophylla. J. Appl. Phycol. 2017, 29, 1057–1067. [Google Scholar] [CrossRef]
- Holdt, S.L.; Kraan, S. Bioactive Compounds in Seaweed: Functional Food Applications and Legislation. J. Appl. Phycol. 2011, 23, 543–597. [Google Scholar] [CrossRef]
- Dhargalkar, V.K.; Pereira, N. Seaweed: Promising Plant of the Millennium. Sci. Cult. 2005, 71, 60–66. [Google Scholar]
- Figueira, T.A. Macroalgas e Suas Aplicações Biotecnológicas. In VIII Botânica no Inverno; Instituto de Biociências: São Paulo, Brazil, 2020. [Google Scholar]
- Hashim, R.; Saat, M.A.M. The Utilization of Seaweed Meals as Binding Agents in Pelleted Feeds for Snakehead (Channa striatus) Fry and Their Effects on Growth. Aquaculture 1992, 108, 299–308. [Google Scholar] [CrossRef]
- Mustafa, G.; Wakamatsu, S.; Takeda, T.; Umino, T.; Nakagawa, H. Effects of Algae Meal as Feed Additive on Growth, Feed Efficiency, and Body Composition in Red Sea Bream. Fish. Sci. 1995, 61, 25–28. [Google Scholar] [CrossRef]
- Gabrielsen, B.O.; Austreng, E. Growth, Product Quality and Immune Status of Atlantic Salmon, Salmo salar L., Fed Wet Feed with Alginate. Aquac. Res. 1998, 29, 397–401. [Google Scholar] [CrossRef]
- Naidoo, K.; Maneveldt, G.; Ruck, K.; Bolton, J.J. A Comparison of Various Seaweed-Based Diets and Formulated Feed on Growth Rate of Abalone in a Land-Based Aquaculture System. J. Appl. Phycol. 2006, 18, 437–443. [Google Scholar] [CrossRef]
- Sharma, H.S.S.; Fleming, C.; Selby, C.; Rao, J.R.; Martin, T. Plant Biostimulants: A Review on the Processing of Macroalgae and Use of Extracts for Crop Management to Reduce Abiotic and Biotic Stresses. J. Appl. Phycol. 2014, 26, 465–490. [Google Scholar] [CrossRef]
- Van Oosten, M.J.; Pepe, O.; De Pascale, S.; Silletti, S.; Maggio, A. The Role of Biostimulants and Bioeffectors as Alleviators of Abiotic Stress in Crop Plants. Chem. Biol. Technol. Agric. 2017, 4, 5. [Google Scholar] [CrossRef]
- Basak, A. Effect of Preharvest Treatment with Seaweed Products, Kelpak® and Goëmar BM 86®, on Fruit Quality in Apple. Int. J. Fruit Sci. 2008, 8, 1–14. [Google Scholar] [CrossRef]
- Chouliaras, V.; Tasioula, M.; Chatzissavvidis, C.; Therios, I.; Tsabolatidou, E. The Effects of a Seaweed Extract in Addition to Nitrogen and Boron Fertilization on Productivity, Fruit Maturation, Leaf Nutritional Status and Oil Quality of the Olive (Olea europaea L.) Cultivar Koroneiki. J. Sci. Food Agric. 2009, 89, 984–988. [Google Scholar] [CrossRef]
- Abdel-Mawgoud, A.M.R.; Tantaway, A.S.; Hafez, M.M.; Habib, H.A.M. Seaweed Extract Improves Growth, Yield and Quality of Different Watermelon Hybrids. Res. J. Agric. Biol. Sci. 2010, 6, 161–168. [Google Scholar]
- Frioni, T.; Sabbatini, P.; Tombesi, S.; Norrie, J.; Poni, S.; Gatti, M.; Palliotti, A. Effects of a Biostimulant Derived from the Brown Seaweed Ascophyllum nodosum on Ripening Dynamics and Fruit Quality of Grapevines. Sci. Hortic. 2018, 232, 97–106. [Google Scholar] [CrossRef]
- Shukla, P.S.; Mantin, E.G.; Adil, M.; Bajpai, S.; Critchley, A.T.; Prithiviraj, B. Ascophyllum nodosum-Based Biostimulants: Sustainable Applications in Agriculture for the Stimulation of Plant Growth, Stress Tolerance, and Disease Management. Front. Plant Sci. 2019, 10, 462648. [Google Scholar] [CrossRef] [PubMed]
- Hankins, S.D.; Hockey, H.P. The Effect of a Liquid Seaweed Extract from Ascophyllum nodosum (Fucales, Phaeophyta) on the Two-Spotted Red Spider Mite Tetranychus urticae. In Thirteenth International Seaweed Symposium; Springer: Dordrecht, The Netherlands, 1990; pp. 555–559. [Google Scholar]
- Subramanian, S.; Sangha, J.S.; Gray, B.A.; Singh, R.P.; Hiltz, D.; Critchley, A.T.; Prithiviraj, B. Extracts of the Marine Brown Macroalga, Ascophyllum nodosum, Induce Jasmonic Acid Dependent Systemic Resistance in Arabidopsis thaliana against Pseudomonas syringae Pv. Tomato DC3000 and Sclerotinia sclerotiorum. Eur. J. Plant Pathol. 2011, 131, 237–248. [Google Scholar] [CrossRef]
- Panjehkeh, N.; Abkhoo, J. Influence of Marine Brown Alga Extract (Dalgin) on Damping-off Tolerance of Tomato. J. Mater. Environ. Sci. 2016, 7, 2369–2374. [Google Scholar]
- Eismann, A.I.; Perpetuo Reis, R.; Ferreira da Silva, A.; Negrão Cavalcanti, D. Ulva spp. Carotenoids: Responses to Environmental Conditions. Algal Res. 2020, 48, 101916. [Google Scholar] [CrossRef]
- Lee, D.-G.; Hyun, J.-W.; Kang, K.-A.; Lee, J.-O.; Lee, S.-H.; Ha, B.-J.; Ha, J.-M.; Lee, E.Y.; Lee, J.-H. Ulva lactuca: A Potential Seaweed for Tumor Treatment and Immune Stimulation. Biotechnol. Bioprocess. Eng. 2004, 9, 236–238. [Google Scholar] [CrossRef]
- Hassan, S.; El-Twab, S.A.; Hetta, M.; Mahmoud, B. Improvement of Lipid Profile and Antioxidant of Hypercholesterolemic Albino Rats by Polysaccharides Extracted from the Green Alga Ulva lactuca Linnaeus. Saudi J. Biol. Sci. 2011, 18, 333–340. [Google Scholar] [CrossRef]
- Awad, N.E. Biologically Active Steroid from the Green Alga Ulva lactuca. Phytother. Res. 2000, 14, 641–643. [Google Scholar] [CrossRef] [PubMed]
- Moustafa, A.; Marzouk, M.A.; Rabea, E.I.; Abd-Elnabi, A.D. Insecticidal and Fungicidal Activity of Ulva lactuca Linnaeus (Chlorophyta) Extracts and Their Fractions. Annu. Res. Rev. Biol. 2014, 4, 2252–2262. [Google Scholar] [CrossRef]
- Spavieri, J.; Kaiser, M.; Casey, R.; Hingley-Wilson, S.; Lalvani, A.; Blunden, G.; Tasdemir, D. Antiprotozoal, Antimycobacterial and Cytotoxic Potential of Some British Green Algae. Phytother. Res. 2010, 24, 1095–1098. [Google Scholar] [CrossRef] [PubMed]
- Sathivel, A.; Raghavendran, H.R.B.; Srinivasan, P.; Devaki, T. Anti-Peroxidative and Anti-Hyperlipidemic Nature of Ulva lactuca Crude Polysaccharide on d-Galactosamine Induced Hepatitis in Rats. Food Chem. Toxicol. 2008, 46, 3262–3267. [Google Scholar] [CrossRef] [PubMed]
- Mao, W.; Zang, X.; Li, Y.; Zhang, H. Sulfated Polysaccharides from Marine Green Algae Ulva conglobata and Their Anticoagulant Activity. J. Appl. Phycol. 2006, 18, 9–14. [Google Scholar] [CrossRef]
- BelHadj, S.; Hentati, O.; Elfeki, A.; Hamden, K. Inhibitory Activities of Ulva lactuca Polysaccharides on Digestive Enzymes Related to Diabetes and Obesity. Arch. Physiol. Biochem. 2013, 119, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Biba, E. Protection: The Sunscreen Pill. Nature 2014, 515, S124–S125. [Google Scholar] [CrossRef]
- Cardozo, K.H.M.; Guaratini, T.; Barros, M.P.; Falcão, V.R.; Tonon, A.P.; Lopes, N.P.; Campos, S.; Torres, M.A.; Souza, A.O.; Colepicolo, P.; et al. Metabolites from Algae with Economical Impact. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2007, 146, 60–78. [Google Scholar] [CrossRef]
- Kim, S.-K.; Ravichandran, Y.D.; Khan, S.B.; Kim, Y.T. Prospective of the Cosmeceuticals Derived from Marine Organisms. Biotechnol. Bioprocess Eng. 2008, 13, 511–523. [Google Scholar] [CrossRef]
- Leal, M.C.; Rocha, R.J.M.; Rosa, R.; Calado, R. Aquaculture of Marine Non-food Organisms: What, Why and How? Rev. Aquac. 2018, 10, 400–423. [Google Scholar] [CrossRef]
- Sun, Y.; Chavan, M. Cosmetic Compositions Comprising Marine Plants. U.S. Patent No. US9603790B2, 28 March 2017. [Google Scholar]
- Ritchie, H.; Rosado, P. “Fossil Fuels”. 2017. Available online: https://ourworldindata.org/fossil-fuels (accessed on 1 March 2024).
- Osman, M.E.H.; Abo-Shady, A.M.; Elshobary, M.E.; Abd El-Ghafar, M.O.; Abomohra, A.E.-F. Screening of Seaweeds for Sustainable Biofuel Recovery through Sequential Biodiesel and Bioethanol Production. Environ. Sci. Pollut. Res. 2020, 27, 32481–32493. [Google Scholar] [CrossRef] [PubMed]
- Korzen, L.; Pulidindi, I.N.; Israel, A.; Abelson, A.; Gedanken, A. Single Step Production of Bioethanol from the Seaweed Ulva rigida Using Sonication. RSC Adv. 2015, 5, 16223–16229. [Google Scholar] [CrossRef]
- Ritchie, H.; Samborska, V.; Roser, M. Plastic Pollution. Our World in Data. 2023. Available online: https://ourworldindata.org/plastic-pollution (accessed on 1 March 2024).
- Lim, C.; Yusoff, S.; Ng, C.G.; Lim, P.E.; Ching, Y.C. Bioplastic Made from Seaweed Polysaccharides with Green Production Methods. J. Environ. Chem. Eng. 2021, 9, 105895. [Google Scholar] [CrossRef]
- Chopin, T.; Buschmann, A.H.; Halling, C.; Troell, M.; Kautsky, N.; Neori, A.; Kraemer, G.P.; Zertuche-González, J.A.; Yarish, C.; Neefus, C. Integrating Seaweeds into Marine Aquaculture Systems: A Key Toward Sustainability. J. Phycol. 2001, 37, 975–986. [Google Scholar] [CrossRef]
- Chopin, T.; Yarish, C.; Wilkes, R.; Belyea, E.; Lu, S.; Mathieson, A. Developing Porphyra/Salmon Integrated Aquaculture for Bioremediation and Diversification of the Aquaculture Industry. J. Appl. Phycol. 1999, 11, 463–472. [Google Scholar] [CrossRef]
- Troell, M.; Rönnbäck, P.; Halling, C.; Kautsky, N.; Buschmann, A. Ecological Engineering in Aquaculture: Use of Seaweeds for Removing Nutrients from Intensive Mariculture. In Sixteenth International Seaweed Symposium; Springer: Dordrecht, The Netherlands, 1999; pp. 603–611. [Google Scholar]
- Hernández, I.; Tovar, A.; Vergara, J.J. Biofiltering Efficiency in Removal of Dissolved Nutrients by Three Species of Estuarine Macroalgae Cultivated with Sea Bass (Dicentrarchus labrax) Waste Waters 2. Ammonium. J. Appl. Phycol. 2002, 14, 375–384. [Google Scholar] [CrossRef]
- Neori, A.; Krom, M.D.; Ellner, S.P.; Boyd, C.E.; Popper, D.; Rabinovitch, R.; Davison, P.J.; Dvir, O.; Zuber, D.; Ucko, M.; et al. Seaweed Biofilters as Regulators of Water Quality in Integrated Fish-Seaweed Culture Units. Aquaculture 1996, 141, 183–199. [Google Scholar] [CrossRef]
- Lüning, K.; Pang, S. Mass Cultivation of Seaweeds: Current Aspects and Approaches. J. Appl. Phycol. 2003, 15, 115–119. [Google Scholar] [CrossRef]
- Gao, G.; Clare, A.S.; Rose, C.; Caldwell, G.S. Ulva Rigida in the Future Ocean: Potential for Carbon Capture, Bioremediation and Biomethane Production. GCB Bioenergy 2018, 10, 39–51. [Google Scholar] [CrossRef]
- Kut Güroy, B.; Cirik, Ş.; Güroy, D.; Sanver, F.; Tekinay, A.A. Effects of Ulva rigida and Cystoseira barbata Meals as a Feed Additive on Growth Performance, Feed Utilization, and Body Composition of Nile Tilapia, Oreochromis niloticus. Turk. J. Vet. Anim. Sci. 2007, 31, 91–97. [Google Scholar]
- Gao, G.; Liu, Y.; Li, X.; Feng, Z.; Xu, J. An Ocean Acidification Acclimatised Green Tide Alga Is Robust to Changes of Seawater Carbon Chemistry but Vulnerable to Light Stress. PLoS ONE 2016, 11, e0169040. [Google Scholar] [CrossRef] [PubMed]
- Nelson, W.; Neill, K.; D’Archino, R. When Seaweeds Go Bad: An Overview of Outbreaks of Nuisance Quantities of Marine Macroalgae in New Zealand. N. Z. J. Mar. Freshw. Res. 2015, 49, 472–491. [Google Scholar] [CrossRef]
- Thornber, C.S.; Guidone, M.; Deacutis, C.; Green, L.; Ramsay, C.N.; Palmisciano, M. Spatial and Temporal Variability in Macroalgal Blooms in a Eutrophied Coastal Estuary. Harmful Algae 2017, 68, 82–96. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Cui, J.; Zhang, J.; Shi, J.; Kang, X.; Liu, J.; Wen, Q.; He, P. Reproductive Strategy of the Floating Alga Ulva prolifera in Blooms in the Yellow Sea Based on a Combination of Zoid and Chromosome Analysis. Mar. Pollut. Bull. 2019, 146, 584–590. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, H.; Loret, E.P. Ulva lactuca, A Source of Troubles and Potential Riches. Mar. Drugs 2019, 17, 357. [Google Scholar] [CrossRef]
- Yabe, T.; Ishii, Y.; Amano, Y.; Koga, T.; Hayashi, S.; Nohara, S.; Tatsumoto, H. Green Tide Formed by Free-Floating Ulva spp. at Yatsu Tidal Flat, Japan. Limnology 2009, 10, 239–245. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, D.; Jiao, N. Tapping Carbon Sequestration Potential of Blooming Macroalgae to Mitigate Climate Change. Ocean-Land-Atmos. Res. 2023, 2, 0033. [Google Scholar] [CrossRef]
- Park, S. Sea Lettuce and Nutrient Monitoring in Tauranga Harbour 1991–2010; Bay of Plenty Regional Council: Whakatane, New Zealand, 2011. [Google Scholar]
- Cox, P.M.; Betts, R.A.; Jones, C.D.; Spall, S.A.; Totterdell, I.J. Acceleration of Global Warming Due to Carbon-Cycle Feedbacks in a Coupled Climate Model. Nature 2000, 408, 184–187. [Google Scholar] [CrossRef]
- Rabalais, N.N.; Turner, R.E.; Díaz, R.J.; Justić, D. Global Change and Eutrophication of Coastal Waters. ICES J. Mar. Sci. 2009, 66, 1528–1537. [Google Scholar] [CrossRef]
- Teichberg, M.; Fox, S.E.; Olsen, Y.S.; Valiela, I.; Martinetto, P.; Iribarne, O.; Muto, E.Y.; Petti, M.A.V.; Corbisier, T.N.; Soto-Jiménez, M.; et al. Eutrophication and Macroalgal Blooms in Temperate and Tropical Coastal Waters: Nutrient Enrichment Experiments with Ulva spp. Glob. Change Biol. 2010, 16, 2624–2637. [Google Scholar] [CrossRef]
- Trenberth, K. Changes in Precipitation with Climate Change. Clim. Res. 2011, 47, 123–138. [Google Scholar] [CrossRef]
- Sydeman, W.J.; García-Reyes, M.; Schoeman, D.S.; Rykaczewski, R.R.; Thompson, S.A.; Black, B.A.; Bograd, S.J. Climate Change and Wind Intensification in Coastal Upwelling Ecosystems. Science 2014, 345, 77–80. [Google Scholar] [CrossRef] [PubMed]
- Jackson, J.B.C.; Kirby, M.X.; Berger, W.H.; Bjorndal, K.A.; Botsford, L.W.; Bourque, B.J.; Bradbury, R.H.; Cooke, R.; Erlandson, J.; Estes, J.A.; et al. Historical Overfishing and the Recent Collapse of Coastal Ecosystems. Science 2001, 293, 629–637. [Google Scholar] [CrossRef] [PubMed]
- Green-Gavrielidis, L.A.; Thornber, C.S. Will Climate Change Enhance Algal Blooms? The Individual and Interactive Effects of Temperature and Rain on the Macroalgae Ulva. Estuaries Coasts 2022, 45, 1688–1700. [Google Scholar] [CrossRef]
- Eriksson, B.K.; Ljunggren, L.; Sandström, A.; Johansson, G.; Mattila, J.; Rubach, A.; Råberg, S.; Snickars, M. Declines in Predatory Fish Promote Bloom-forming Macroalgae. Ecol. Appl. 2009, 19, 1975–1988. [Google Scholar] [CrossRef]
- Pereira, F. Agência Ambiental da ONU dá Exemplo em Neutralizar Emissões de Carbono. Available online: https://www.unep.org/pt-br/noticias-e-reportagens/story/agencia-ambiental-da-onu-da-exemplo-em-neutralizar-emissoes-de-carbono (accessed on 1 March 2024).
- Mantri, V.A.; Singh, R.P.; Bijo, A.J.; Kumari, P.; Reddy, C.R.K.; Jha, B. Differential Response of Varying Salinity and Temperature on Zoospore Induction, Regeneration and Daily Growth Rate in Ulva fasciata (Chlorophyta, Ulvales). J. Appl. Phycol. 2011, 23, 243–250. [Google Scholar] [CrossRef]
- MND. Staff ‘Mr. Sargassum’ Has Built 13 Houses with Blocks Made from the Smelly Seaweed. Mexico News Daily, 20 July 2022. [Google Scholar]
- Mkrtichian, E. Sargablock. Available online: https://fortomorrow.org/explore-solutions/sargablock (accessed on 1 March 2024).
- Buckley, S. Seaweed as Insulation? Available online: https://www.architectureanddesign.com.au/news/seaweed-as-insulation (accessed on 1 March 2024).
- Widera, B. Possible Application of Seaweed as Building Material in the Modern Seaweed House on Laesø. In Proceedings of the 30th International PLEA Conference: Sustainable Habitat for Developing Societies—Choosing the Way Forward, CEPT University, Ahmedabad, India, 16–18 December 2014. [Google Scholar]
- Textile Value Chain Fashion from the Sea. Available online: https://textilevaluechain.in/news-insights/fashion-from-the-sea-algae-seaweed/ (accessed on 1 March 2024).
Figure 1.
Flow-chart summarizing the discussed topics in this review.
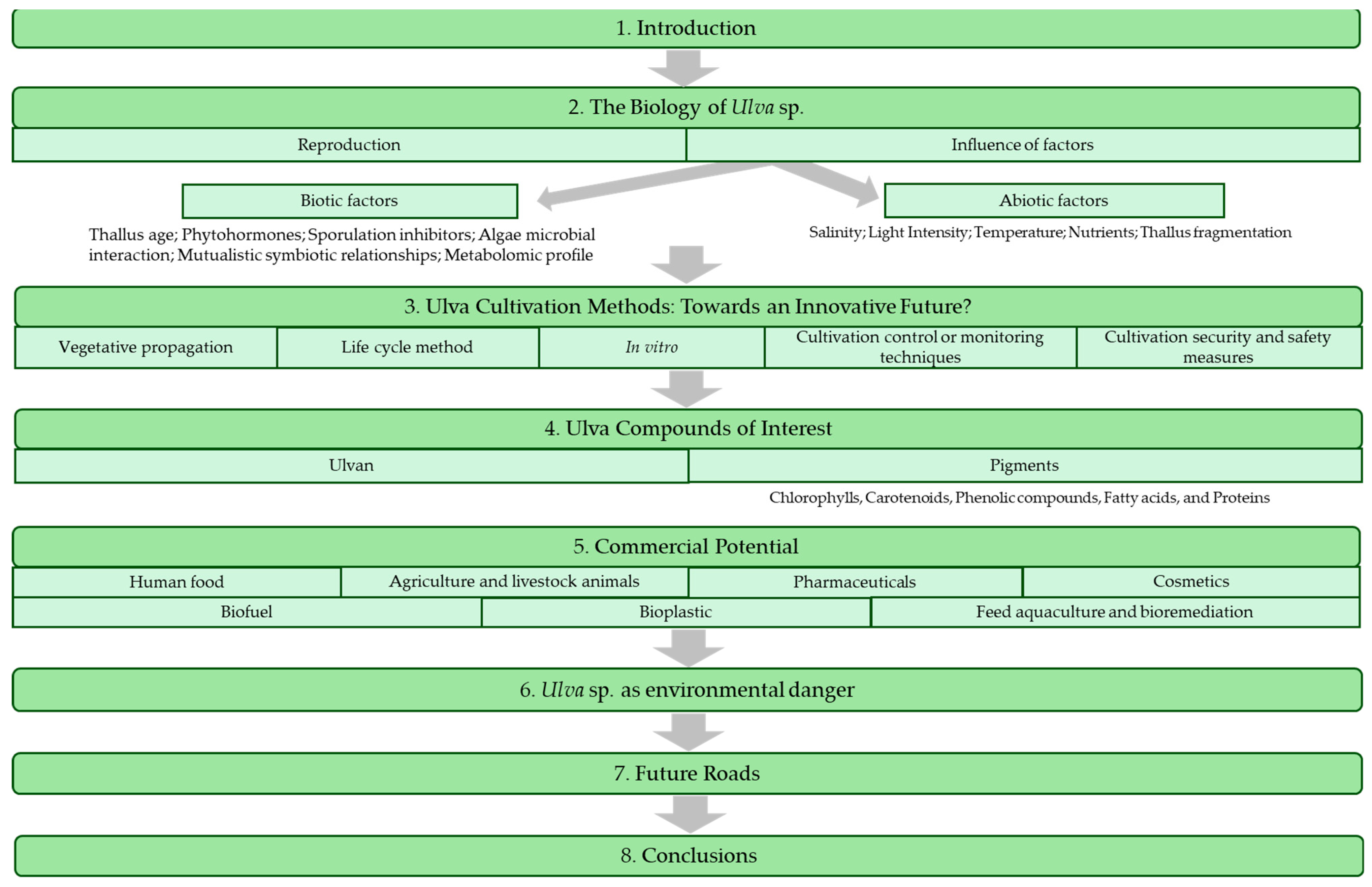
Figure 2.
Typical life cycle of Ulva sp.

Figure 3.
Natural bacterial community interaction with Ulva sp. juveniles. Scale bar created using Microscope VIS software.
Figure 3.
Natural bacterial community interaction with Ulva sp. juveniles. Scale bar created using Microscope VIS software.

Figure 4.
Time course of gametogenesis in Ulva sp. based on the description from [14,41]. (a) “determination phase”—gametogenesis induction after 24 h with chloroplast reorientation, (b) swarming phase—reproductive cells with fully formed gametes and gametes can be released upon removal of the SWI, (c) zoospores within a sporangium (sporophyte phase), (d) gametes fertilization, (e) zygotes, (f) individual on phase IV, and (g) juvenile (phase V) compared with [105]. Scale bars created using Microscope VIS software. Image sources from the authors.
Figure 4.
Time course of gametogenesis in Ulva sp. based on the description from [14,41]. (a) “determination phase”—gametogenesis induction after 24 h with chloroplast reorientation, (b) swarming phase—reproductive cells with fully formed gametes and gametes can be released upon removal of the SWI, (c) zoospores within a sporangium (sporophyte phase), (d) gametes fertilization, (e) zygotes, (f) individual on phase IV, and (g) juvenile (phase V) compared with [105]. Scale bars created using Microscope VIS software. Image sources from the authors.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Costa, S.P.; Cotas, J.; Pereira, L. Laminar Ulva Species: A Multi-Tool for Humankind? Appl. Sci. 2024, 14, 3448. https://doi.org/10.3390/app14083448
AMA Style
Costa SP, Cotas J, Pereira L. Laminar Ulva Species: A Multi-Tool for Humankind? Applied Sciences. 2024; 14(8):3448. https://doi.org/10.3390/app14083448
Chicago/Turabian StyleCosta, Sofia Pereira, João Cotas, and Leonel Pereira. 2024. "Laminar Ulva Species: A Multi-Tool for Humankind?" Applied Sciences 14, no. 8: 3448. https://doi.org/10.3390/app14083448
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.