
Indole-3-Acetic Acid Action in Outdoor and Indoor Cultures of Spirulina in Open Raceway Reactors
by
 ,
,
Jéssica Teixeira da Silveira
1 ,
,
Ana Priscila Centeno da Rosa
1,
Michele Greque de Morais
2 and
Jorge Alberto Vieira Costa
1,* 1
Laboratory of Biochemical Engineering, College of Chemistry and Food Engineering, Federal University of Rio Grande, P.O. Box 474, Rio Grande 96201-900, RS, Brazil
2
Laboratory of Microbiology and Biochemistry, College of Chemistry and Food Engineering, Federal University of Rio Grande, P.O. Box 474, Rio Grande 96201-900, RS, Brazil
*
Author to whom correspondence should be addressed.
Appl. Sci. 2024, 14(9), 3715; https://doi.org/10.3390/app14093715
Submission received: 1 April 2024
/
Revised: 24 April 2024
/
Accepted: 24 April 2024
/
Published: 26 April 2024
(This article belongs to the Special Issue New Insights into Microalgae Cultivation and Downstream Processes, 3rd Edition)
Abstract
:A significant research gap exists in investigating large-scale microalgae cultures exposed to outdoor conditions, with the addition of phytohormones using non-sterile growth media. Implementing these conditions is crucial for verifying the industrial viability of this strategy. This study aimed to evaluate the effect of indole-3-acetic acid (IAA) supplementation on Spirulina sp. LEB 18 cultures conducted indoors and outdoors in raceway bioreactors. The outdoor experiments were performed under uncontrolled environmental conditions. The indoor cultures were maintained within a thermostat-controlled chamber at a consistent temperature and lighting intensity. The outdoor experiments supplemented with IAA achieved a biomass concentration of 5.43 g L−1 and productivity of 173.9 mg L−1 d−1. These values increased 122.5% and 130.9% in biomass concentration and productivity, respectively, compared to the indoor experiments with the same supplementation. Moreover, roughly half of the biomass generated from outdoor cultivation with IAA consisted of carbohydrates (45%). Compared to indoor cultivation, this approach reduced production costs for biomass (55%) and lowered production costs for carbohydrates, proteins, and lipids by 86%, 44%, and 50%, respectively. The successful application of phytohormones in microalgae cultures, particularly under larger scale, nonsterile, and outdoor conditions, represents a significant advancement toward industrial implementation.
1. Introduction
The pursuit of renewable sources of food and biofuels has emerged as a top priority in response to global challenges, such as population growth and the scarcity of natural resources [1]. Within this context, the microalga Spirulina (Arthrospira) holds promising potential to address these demands [2] Spirulina biomass contains iron, essential unsaturated fatty acids (such as γ-linoleic acid), vitamin B12, and a variety of B vitamins and natural carotenes, making them valuable for food applications and health foods. Additionally, Spirulina is seen as a sustainable feedstock for bioenergy and biofuels, offering potential as a carbon-neutral source for biodiesel, bioethanol, and biogas production [3]. Spirulina is one of the most extensively researched and cultivated microalgal genera worldwide [4] due to its adaptability to various cultivation conditions, diverse environments, culture media, and reactor types [3].
Among the factors impacting microalgal growth and productivity, light, temperature, and nutrients are the most influential [5]. Light is pivotal in photosynthesis, furnishing the energy needed for carbon fixation and microalgae growth. The intensity, spectral quality, and photoperiod represent critical factors influencing photosynthetic activity and, consequently, culture productivity [6]. The microalgae Spirulina boasts a wide range of growth temperatures, demonstrating its ability to adapt to tropical and subtropical conditions [7].
Furthermore, light availability and exposure to varying temperatures can regulate the synthesis of specific biomolecules, including photosynthetic pigments [8], proteins, and carbohydrates [9]. These molecules hold significance for applications in the food [10], cosmetics [11], pharmaceutical [12], biofuel [13], and environmental solution [14] sectors.
Specifically, concerning the microalga Spirulina, recent studies have unveiled the presence of its phytohormones [9] and their influence on cultivation modulation [15,16]. Among phytohormones, the best known are auxins, with indole-3-acetic acid (IAA) standing out for showing more significant activity in plants and algae [17] and stimulating Spirulina biomass and biomolecules production [14,15]. Phytohormones encompass natural chemical compounds that play a fundamental role in the development and metabolism of plants, algae, and microalgae. Phytohormones are effective in enhancing the microalgae growth and production of biomolecules by promoting the expression of genes involved in DNA replication and repair and energy metabolism (glycolysis, TCA cycle, and oxidative phosphorylation), as well as signaling and protective synthesis pathways [18]. In addition, phytohormones have advantages such as being efficient at minimal concentrations, natural, environmentally safe, and degrading quickly, leaving no environmental remnants [19]. However, the mechanism needs to be clarified. Therefore, further investigations should be conducted to identify the genes that control the effects of phytohormones.
Despite the extensive literature documenting the impact of phytohormone stimulation on microalgae biomass and biomolecule productivity [15,16,18,20,21,22,23,24], no research to date has explored the role of phytohormones within outdoor microalgae cultivation systems employing nonsterile media and operating on a larger scale [25].
In this study, we investigate the application of phytohormones in outdoor microalgae cultivation using raceway-type reactors and a nonsterile medium, focusing on the supplementation of indole-3-acetic acid (IAA). Our outdoor experimentation reveals promising enhancements compared to indoor studies. These findings highlight the potential of phytohormone supplementation as a cost-effective and environmentally friendly strategy for large-scale microalgae production, addressing a crucial research gap in the field.
In addition, the comparison between indoor and outdoor microalgae cultivation is crucial, as outdoor cultivation in raceway-type reactors offers the advantage of preventing the necessity for artificial lighting and aeration. This strategic decision aligns with sustainability principles and decreases energy consumption, making microalgae cultivation economically feasible and environmentally sustainable [3]. Reactors like the open raceway reactors utilized in this study, designed for outdoor operation, are extensively employed in commercial microalgae production due to their cost-effectiveness in construction and operation compared to enclosed systems [26]. In light of this context, a critical investigation into the influence of phytohormones in cultures conducted under uncontrolled environmental conditions becomes imperative. Such research could offer valuable insights to optimize cultivation parameters and maximize targeted biomolecule production.
Additionally, this cultivation approach considers energy efficiency, sustainability, and economic feasibility and provides pivotal guidance for implementing large-scale production strategies. Consequently, this study aims to assess the cost-effectiveness and productivity of Spirulina sp. LEB 18 supplemented with indole-3-acetic acid (IAA) when the cultivation is conducted within raceway bioreactors under indoor and outdoor conditions.
2. Materials and Methods
2.1. Microalga and Phytohormone
Spirulina LEB 18 was used for the experiments. This microorganism is sourced from the strain bank of the Laboratory of Biochemical Engineering at the Federal University of Rio Grande (FURG)—Brazil. The microalga was isolated at Lagoa Mangueira, at latitude 33°31′08″ S and longitude 53°22′05″ W [27].
For supplementation of the microalgal cultures, an auxin-type phytohormone was employed. The commercial indole-3-acetic acid (IAA) (98%, Sigma-Aldrich, Cotia, SP, Brazil) was introduced into Spirulina cultures from a 1000 ppm stock solution [15].
2.2. Culture Conditions
Both indoor and outdoor experiments were conducted in duplicate for 30 days, utilizing open reactors of the 6 L raceway type (with a usable volume of 5 L). Continuous agitation was achieved by paddle wheels rotating at 20 rpm. The initial biomass concentration of the cultures was set at 0.2 g L−1 [28]. The microalgae were cultivated in Zarrouk medium [29] employing a batch-fed system, with IAA supplementation introduced at a concentration of 0.1 mg L−1 on the 15th day of cultivation, according to the results obtained in previous research [15,16]. In addition, control experiments were performed. These were conducted in the same conditions as the supplemented experiments but without IAA treatment.
Indoor cultures were situated within a thermostat-controlled chamber, maintaining a consistent temperature of 30 °C, a 12 h light/dark photoperiod, and an illuminance of 80 μmol photons m−2 s−1 provided by fluorescent lamps. The outdoor experiments were carried out under unregulated environmental conditions during the winter period (August 2022) in the city of Rio Grande, RS, Brazil (latitude 32°4′23.028″ S, longitude 52°10′2.079″ W). The outdoor bioreactors were situated within a greenhouse covered by a transparent film, offering protection against UV rays [30]. Monitoring occurred daily in the early afternoon (2 p.m.) to record the cultivation and ambient temperatures. A digital thermometer (SH 102, J. Prolab, São Paulo, SP, Brazil) was employed, and solar radiation levels were measured with a universal light meter (ULM- 500, Walz, Effeltrich, BY, Germany).
The indoor experiments were designated C-I and IAA-I for the control and supplemented groups, respectively. The designations C-O and IAA-O were assigned to the control and supplemented experiments, respectively, conducted in the outdoor environment.
2.3. Evaluation of Cell Growth
At the initiation and end of the experiments, assessments were conducted to determine the biomass concentration, pH levels, fluorescence, and cell morphology. Biomass concentration was gauged through optical density measurements at 670 nm, utilizing a spectrophotometer (Shimadzu UV/VIS UVmini-1240 spectrophotometer, Kyoto City, Japan), aided by a calibration curve correlating optical density with biomass dry mass [29].
The optical density data enabled the determination of the final biomass concentration and subsequently facilitated calculating the final biomass productivity. pH measurements were taken using a digital pH meter (Mettler Toledo FiveGoTM, Greifensee, Switzerland). Fluorescence measurements were taken with a fluorimeter (AquaPen AP 100, Photon System-Instrument, Drásov, Czech Republic) on day 0 of cultivations (Fv/Fm initial) and the last day of experiments (Fv/Fm final). Samples of 3 mL were dark adapted for 15 min in the cuvettes before the measurement. A wavelength of 620 nm excitation was used. Cell morphology was assessed using an electron microscope (AxioCan ERc 5s Microscope camera, Zeiss, Germany).
2.4. Biomass Characterization
After 30 days of cultivation, in the stationary growth phase, the cultures were harvested and centrifugated (Hitachi, Himac CR-GIII, Tokyo, Japan) at 2000× g for 20 min to recover the biomass. Subsequently, a washing step was carried out to eliminate salts; following which, the biomass was frozen at –80 °C, subjected to lyophilization for 48 h, and then stored at –20 °C until characterization. The lyophilized biomass was utilized to analyze lipids, carbohydrates, proteins, phycocyanin, and moisture content.
Total lipids were directly extracted from the biomass using the organic solvent chloroform/methanol (1:2) and quantified through colorimetry [31], with readings correlated to a tripalmitin standard curve. Before carbohydrate and protein analyses, the cell walls of the biomasses were disrupted via an ultrasonic probe (Cole Parmer, CPX 130, Merrillville, IN, USA). Extracts were prepared by combining 5 mg of lyophilized biomass with 10 mL of distilled water and subjecting them to 10 cycles of sonication (59 s on/off). Quantification of total carbohydrates was conducted using the phenol-sulfuric method [32], referencing a standard glucose curve. Protein content was determined utilizing the colorimetric technique described by Lowry et al. [33], employing bovine serum albumin as a standard and including a preliminary step of protein solubilization with NaOH. Phycocyanin was extracted from the microalgal biomass [34], and the total pigment content was measured following the protocol of Bennett and Bogorad [35]. Furthermore, the purity of the phycocyanin extract was ascertained by calculating the ratio between the absorbance reading of the extract at 620 nm (indicative of pigment concentration) and the absorbance at 280 nm (reflecting protein concentration in the solution) [36]. The moisture content was assessed following the methodology outlined in the Official Methods of Analysis [37].
After these analyses, lipid, carbohydrate, protein, and phycocyanin productivities were calculated. This calculation involved multiplying the results of biomass productivity by the fractions of each biomolecule found in the biomass of Spirulina sp. LEB 18.
2.5. Cost Analysis
The price of the IAA phytohormone (98%) and other reagents (ACS-grade reagents) constituting the culture medium utilized in this study was obtained from the Sigma-Aldrich website on March 2023 (Supplementary Table S1). The expense associated with producing microalgal biomass and biomolecules was determined by employing the equation delineated by Park et al. [38] as follows:
where A signifies the quantity of IAA added per liter of culture medium (mg L−1), B stands for the price of IAA (USD mg−1), C denotes the price of the culture medium (USD L−1), and D represents the final concentration of biomass or biomolecule achieved at the culmination of the cultivation (g L−1). This analysis considered only the cost of the culture medium with and without the phytohormone.
Cost (USD g−1 = (A × B) + C/D
2.6. Statistical Analysis
Two biological replicates and three technical replicates were utilized to obtain the responses. All responses were subjected to biological duplicates and triplicate technical samples analysis using analysis of variance (ANOVA) followed by Tukey’s test for comparison of means, with a significance threshold set at the 95% confidence level (p ≤ 0.05).
3. Results
3.1. Biomass Growth and Productivity Evaluation
When evaluating the final biomass concentration (Xfinal) and productivity (Pfinal), both cultivation conditions supplemented with indole-3-acetic acid (IAA), specifically indoor cultivation supplemented with IAA (IAA-I) and outdoor cultivation supplemented with IAA (IAA-O), demonstrated superior performance compared to experiments conducted without supplementation, represented by indoor control culture (C-I) and outdoor control culture (C-O) (Table 1). A 13.9% increase in biomass concentration was observed when comparing the indoor conditions C-I with IAA-I, and an 18.0% increase was observed between the outdoor conditions control (C-O) and supplemented (IAA-O). The productivity was enhanced by 24% when comparing C-I with IAA-I and improved by 18.5% from C-O to IAA-O.
During the outdoor experimental period, environmental conditions were monitored, revealing a wide natural range with a minimum temperature of 4.2 °C and a maximum of 39.7 °C. The minimum and maximum illuminance levels were 160 and 1200 μmol photons m−2 s−1, respectively. The average minimum and maximum temperature values were 11 °C and 32 °C, respectively, and the average illuminance hovered at approximately 700 μmol photons m−2 s−1. Under these conditions, phytohormone supplementation (IAA-O) resulted in an approximately twofold increase in biomass generation compared to indoor experiments with the addition of the phytohormone (IAA-I) (Table 1).
During day zero of the cultures, the maximum initial photochemical efficiency (Fv/Fm initial) consistently showed similar values across all conditions (Table 1). However, a reduction in the final maximum photochemical efficiency (Fv/Fm final) was observed after 30 days of cultivation under indoor conditions (C-I and IAA-I). In contrast, the outdoor conditions (C-O and IAA-O) did not exhibit a statistically significant difference in the final response compared to the initial measurement. At the end of the experimental period, higher pH values were observed in outdoor cultures (C-O and IAA-O) compared to indoor experiments (C-I and IAA-I) (Table 1). Notably, control and supplemented conditions within the same environment exhibited no significant difference in pH values.
3.2. Biomass Characterization
Experiments with phytohormone supplementation (IAA-I and IAA-O) demonstrated lower protein content than those with control conditions (C-I and C-O) in indoor and outdoor settings. However, since biomass productivity was higher in the conditions with the supplement (IAA-I and IAA-O) compared to controls (C-I and C-O), the protein productivity followed the same trend (Figure 1). In both scenarios, the addition of IAA to Spirulina cultures yielded an approximately 11% increase in protein productivity compared to control experiments under similar conditions.
Compared to control experiments in the same environment, the supplemented conditions exhibited a higher carbohydrate concentration and productivity within the biomass (Figure 2). The IAA-I condition showed 99.7% and 144.9% increases in carbohydrate concentration and productivity, respectively, compared to the C-I control. In an outdoor setting, the IAA-O-supplemented condition demonstrated a 72.6% concentration increase and a 114.6% boost in carbohydrate productivity compared to the results obtained from the un-supplemented condition (C-O). Furthermore, outdoor cultivation with phytohormone supplementation (IAA-O) exhibited approximately 225% higher biomolecule content and productivity than indoor conditions (IAA-I). The robust biomass productivity observed in the supplemented outdoor experiment (Table 1) coupled with the substantial carbohydrate content (Figure 2) led to a twofold increase in carbohydrate productivity compared to the outdoor control (C-O) and an eightfold increase over the indoor control (C-I).
The microalgal biomass showed no significant difference in the lipid concentration response under the conditions tested (Figure 3). However, when the microalga was cultivated in an outdoor environment (C-O and IAA-O), it showed higher productivity of the lipid biomolecule than when grown in the indoor conditions (C-I and IAA-I).
The biomass generated from outdoor-supplemented cultivation (IAA-O) exhibited higher phycocyanin productivity compared to the other conditions (Figure 4). However, concerning phycocyanin concentration, the outdoor-supplemented condition (IAA-O) showed no significant difference from the control experiment conducted in the same environment (C-O).
The purity achieved in phycocyanin extracts was higher in outdoor experiments (C-O = 2.4a,b ± 0.1 and IAA-O = 2.5a ± 0.1) compared to indoor experiments (C-I = 2.0b ± 0.2 and IAA-I = 1.2c ± 0.1). However, there was no statistically significant difference when comparing the results of the extracts obtained from supplemented cultures with those obtained from controls in the outdoor condition. On the other hand, in the indoor condition, the control generated an extract with a greater purity.
3.3. Biomass and Biomolecule Production Costs
While various factors, such as energy, harvesting, and labor significantly influence the production costs of biomass and biomolecules, this study focused solely on the impact of phytohormone supplementation. As such, other cultivation parameters were not considered when calculating production costs. Therefore, only the costs associated with the culture medium, with and without phytohormones, were considered. However, we emphasize that further research is needed to delve deeper into the study of costs to enhance the robustness of this cultivation technique in the future.
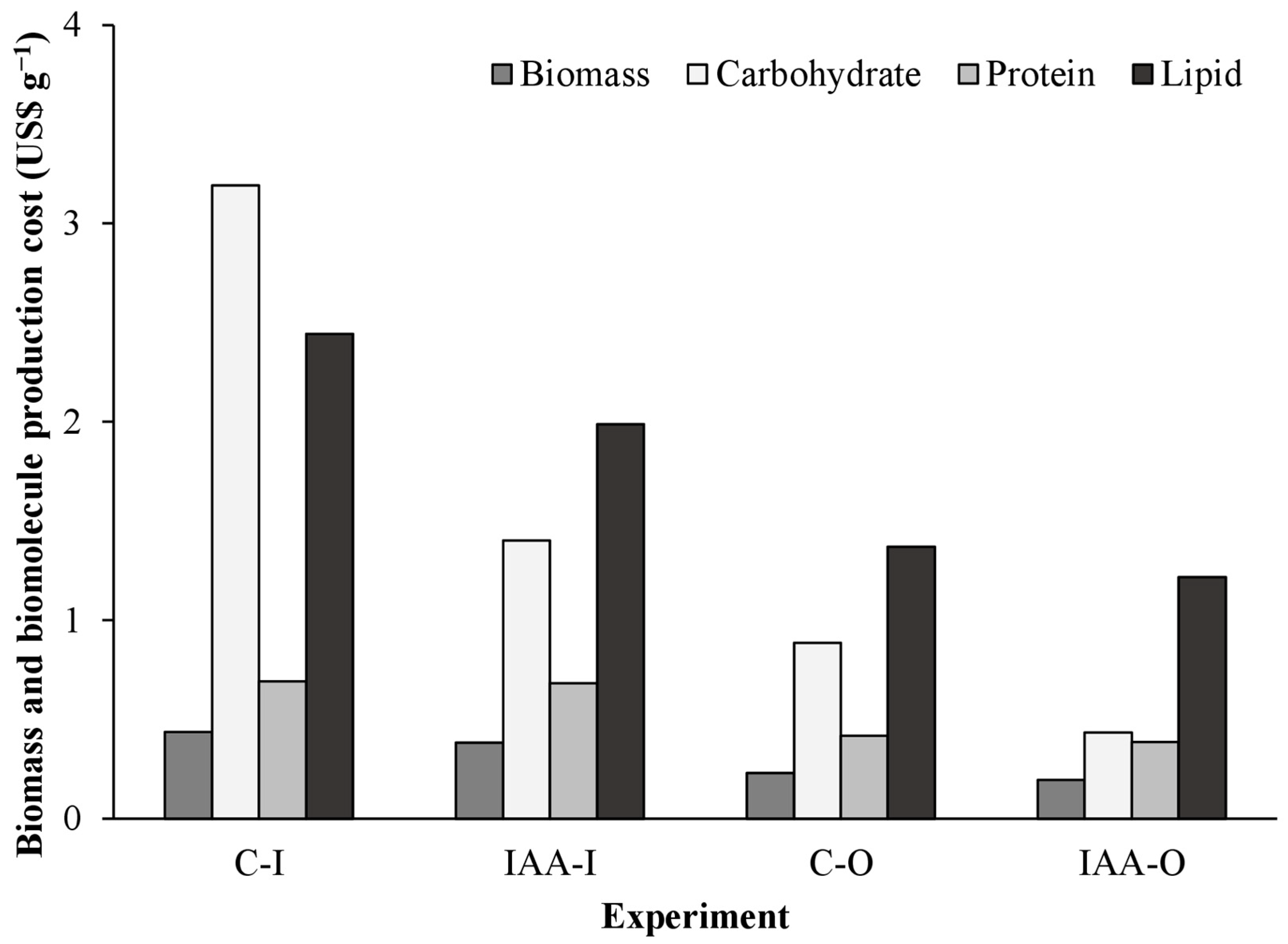
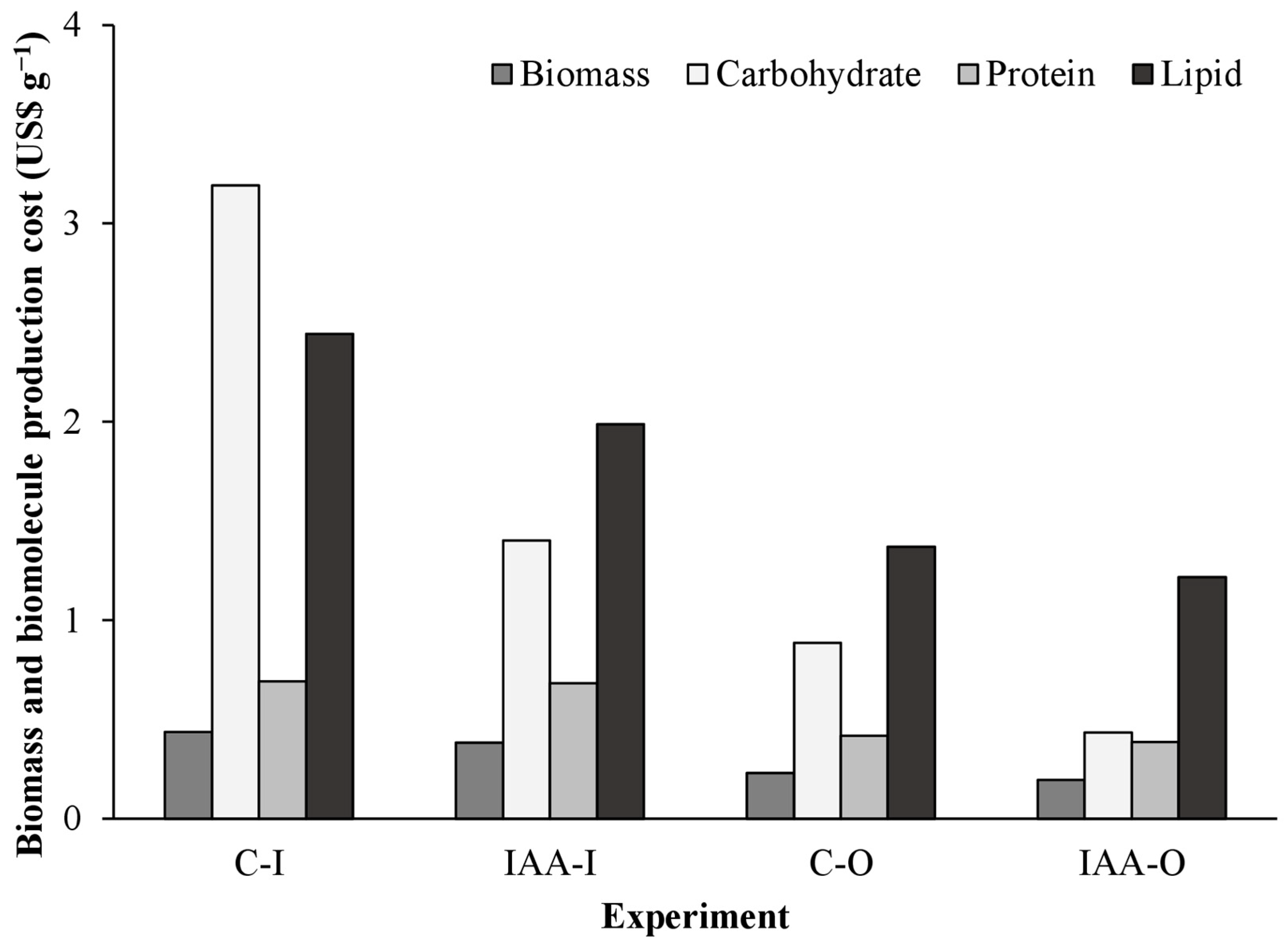
The implementation of IAA supplementation in outdoor cultivation (IAA-O) resulted in a 55%, 86%, 44%, and 50% reduction in the cost of biomass, carbohydrate, protein, and lipid production, respectively, in comparison with control experiments conducted indoors (C-I) (Figure 1). This cost comparison was based on the conditions with the highest and lowest production costs (C-I and IAA-O, respectively).
Outdoor experiments (C-O and IAA-O) yielded lower biomass and biomolecule production costs than indoor cultivation (C-I and IAA-I). Furthermore, supplemented conditions (IAA-I and IAA-O) demonstrated lower production costs compared to their respective controls (C-I and C-O) (Figure 5). Compared with C-I production costs, the IAA-I production cost of biomass, carbohydrates, and lipids was reduced by 12%, 56%, and 19%, respectively. Additionally, compared with C-O production costs, the IAA-O biomass production cost was 16% lower, the carbohydrate cost was 51% lower, and the lipid cost was 11% lower. However, the protein production cost was reduced by less than 10% compared to the control and supplemented conditions. Phycocyanin production costs were lower in the indoor control condition (USD 2.30).
4. Discussion
The findings from this study present significant implications for the scale-up of microalgae cultivation, particularly in outdoor settings and under uncontrolled conditions, with the supplementation of IAA showing promising potential. The observed outcomes pave the way for the industrial adoption of this strategy, given the substantial enhancements in productivity and reduction in biomass and biomolecule production costs achieved through the synergistic interaction of IAA and environmental factors.
The results regarding biomass growth and productivity are consistent with the previous literature indicating the positive impact of phytohormone supplementation on microalgae under adverse conditions [39,40,41,42,43]. Despite the extreme fluctuations in abiotic factors, outdoor cultivation supplemented with IAA exhibited a higher biomass concentration and productivity than controlled indoor conditions. Phytohormones are suggested to confer cellular stress resistance by inhibiting chloroplast structure degradation [19].
Similar to the synergistic effects observed between phytohormones and different light spectra to enhance biomass and biomolecule production [44,45], optimal light intensity is also pertinent for the efficacy of phytohormone performance, as evidenced by growth parameter evaluations. Notably, the outdoor experiments experienced higher light intensities, with a minimum of 160 μmol photons m−2 s−1, twice that of indoor cultivation (80 μmol photons m−2 s−1). Furthermore, the maximum illuminance in outdoor conditions was 15-fold higher (1200 μmol photons m−2 s−1) than indoors. The elevation in biomass concentration is attributed to the concurrent substantial augmentation in light intensity. Exposing Spirulina sp. LEB 18 to these intensified environmental conditions proved efficient in promoting its growth. Moreover, it amplified the phytohormone’s effectiveness on the microalgae, culminating in a synergistic growth effect.
The maximum quantum efficiency (Fv/Fm) indicates Photosystem II’s photosynthetic efficiency. A decline in the Fv/Fm value signifies a reduction in photosynthetic efficiency and can be attributed to the cellular stress experienced by microalgae [46]. Remarkably, the outdoor experiments displayed no disparity between initial and final values, signifying the consistent maintenance of maximum quantum efficiency throughout the cultivation period and an absence of noteworthy stress exposure. Conversely, the indoor experiments exhibited a decrease in this parameter after 30 days of cultivation. This phenomenon can be elucidated by microalgae’s diminished photosynthesis and energy generation when subjected to low-light indoor conditions [47].
Both indoor and outdoor experiments exhibited an elevation in pH levels. This phenomenon can be attributed to the bicarbonate assimilation mechanism inherent to microalgae. Initially, the Zarrouk medium contains sodium bicarbonate (NaHCO3) as the primary form of carbon, serving as the favored substrate for carbon assimilation by microalgae [48]. This process is reflected in the pH range of 6 to 10.5. Over time, the microalgae consume two bicarbonate ions: one is internalized as carbon dioxide (CO2), and the other is released as carbonate (CO3−2). This intricate interplay contributes to a pH elevation in the medium, resulting in values surpassing 10.5 [49].
Consequently, the outcomes derived from this study point toward a greater assimilation of carbon molecules in outdoor cultures. This assertion is supported by a pH level exceeding 11 at the culmination of the outdoor experiment, whereas indoor conditions maintained a pH of approximately 10. This can be compared to the findings of Deamici et al. [50], who observed similar trends when cultivating Spirulina sp. LEB 18 in raceways augmented with magnetic fields. Notably, their indoor tests yielded a maximum pH of 10.75, while the outdoor cultures recorded a pH peak of 12.77.
Microalgae can harness both CO2 and HCO3 as substrates for their CO2 metabolism. Through active transport of inorganic carbon, these microorganisms accumulate intracellular bicarbonate and activate internal carbonic anhydrase [51]. This biological process underpins the mechanism for the observed elevation in carbohydrate content resulting from IAA supplementation. Phytohormones likely contribute to carbon fixation during photosynthesis [19,52], stimulating a cascade of reactions within photosynthetic metabolism. Consequently, this stimulation culminates in an amplified synthesis of biomolecules, exemplified by heightened carbohydrate production.
Another significant determinant in fostering carbohydrate synthesis is the choice of reactor configuration. Raceways or analogous open systems facilitate enhanced cellular exposure to atmospheric CO2, consequently promoting biomolecule synthesis [53]. Moreover, outdoor cultivation settings exhibit an augmented carbohydrate yield. This phenomenon can be attributed to increased light exposure and elevated temperatures, which directly influence the synthesis of this macromolecule [54,55].
A contrasting trend was observed in protein content, with the elevation in carbohydrate concentration evident in the supplemented and outdoor experimental conditions compared to controls and indoor setups. This counterintuitive effect can be attributed to the phenomenon of nitrogen limitation because of cultivation duration, which prompts microalgae cells to shift their protein synthesis toward the production of carbohydrates or lipids [56]. The decline in protein synthesis and the concurrent rise in carbohydrate content align with findings reported by Xu and Xiong [18] and Salama et al. [57] in their investigations involving the supplementation of IAA in cultures of Chlorella pyrenoidosa FACHB-9 and Scenedesmus obliquus GU732418, respectively.
Another fact that may have influenced the increase in carbohydrate production and the decrease in protein synthesis in outdoor cultivation is the unfavorable conditions that stress the microalgae cells. This is in line with Zaparoli et al. [58], who described that an increase in light intensity is correlated with a reduction in protein content and an increase in carbohydrate content. The same was observed when the duration of light (photoperiod) increased. Higher carbohydrate content at the expense of lower protein content occurs consistently with higher daily irradiances. Firstly, higher light encourages photosynthesis processes that produce carbohydrates. Secondly, the higher irradiance also increases culture temperature, which induces cyanobacterial cell stress and reduces the enzymatic capacity for protein synthesis so that the first-order photosynthetic products of simple carbohydrates are accumulated in cells [59].
Moreover, analogous to the impact observed due to the type of reactor employed, elevated temperatures and increased illuminance further augmented carbohydrate production while concurrently exerting a deleterious effect on protein content within the biomass. Maintaining the cultivation temperature at approximately 30 °C and ensuring a pH range of 9.0–10.5 creates the optimal conditions for fostering heightened protein synthesis in Spirulina sp. [56,60,61]. Fluctuations in pH and temperature can disrupt the bioavailability of essential nutrients and influence the activity of microalgae cells [62]. Given that the outdoor conditions employed in this study encompassed a broad spectrum of temperatures and illuminance levels and that the resultant pH of the cultures surpassed the ideal range, the observed decline in protein synthesis can be understood.
Nevertheless, notwithstanding the diminished protein concentration observed in the IAA-supplemented conditions compared to the control conditions, the protein productivity exhibited a contrary outcome. This divergence is attributed to the augmented biomass productivity facilitated by the presence of the phytohormone in the culture, which offsets the lower protein content.
Similarly, in line with the investigations by Oliveira [54] and Jesus et al. [61], variations in the experimental conditions did not induce alterations in the lipid content of the microalgal biomass. These authors substantiate that while temperature and illuminance exert a substantial impact on protein and carbohydrate composition, they do not exert a discernible influence on lipid production in Spirulina biomass. Additionally, it is worth noting that supplementation with IAA at a concentration of 0.1 mg L−1 did not yield a significant increase in lipid content in Spirulina sp. LEB 18, as evidenced by the findings of Silveira et al. [16].
In contrast to findings reported in the prevailing literature, the introduction of phytohormone supplementation did not yield an observable enhancement in the production of the pigment phycocyanin [63,64,65,66,67]. Nevertheless, experiments conducted under outdoor conditions demonstrated higher phycocyanin yields than their indoor counterparts. Hence, the confluence of environmental factors may engender shifts in the microalgae’s biochemical composition, potentially augmenting or diminishing the proportion of phycocyanin [68]. Moreover, there is a suggestion of a total or partial correlation between maximum biomass concentration values and phycocyanin content in Spirulina cultures, as observed by Soni et al. [69], Deamici et al. [70], and Jesus et al. [61]. This further underscores the ability of cyanobacterial cells to modulate their photosynthetic efficacy by modulating pigment production levels [71].
Despite the augmented expenses incurred by the incorporation of phytohormones into the growth medium for microalgal cultures, extant research posits that judicious adjustments in supplementation and cultivation methodologies may yield cost savings while concurrently augmenting biomass and biomolecule output [15,16,24,38,72]. The accomplishment of a cost reduction exceeding fifty percent in the production of biomass and biomolecules constitutes a significant milestone. Notably, the available data indicate that numerous global companies are actively conducting experiments aimed at cost reduction while maintaining the photosynthetic efficiency in Spirulina [3]. Furthermore, cultivating microalgae in an outdoor environment utilizing raceway-type reactors confers the advantage of obviating the need for artificial lighting and aeration. This strategic choice aligns with sustainability principles and reduces energy consumption, making microalgal cultivation more economically viable and environmentally sustainable.
Although IAA is considered a high input, the increased production of biomass and biomolecules generated by supplementation makes its use advantageous. In this study, we used a small concentration to stimulate these effects. Thus, the cost of the cultivation medium was not increased significantly. It is essential to highlight that the microalgae, type of phytohormone used, supplementation concentration, and cultivation method, among other parameters, must be considered for the best cost–benefit of using the strategy.
An additional noteworthy aspect is that adopting phytohormones as a strategy does not necessitate structural overhauls within the industry. Reactors like those employed in this study, specifically raceway-type open reactors, are extensively utilized for commercial microalgae production due to their lower construction and operational expenses than enclosed systems [26]. This approach aligns well with industry practices and can be readily integrated without significant modifications.
The biomass produced in this study is rich in carbohydrates and presents high productivity and good cost–benefit, so it could be applied in various sectors. In energy terms, microalgae carbohydrates can produce different biofuels, including bioethanol, biobutanol, biomethane, and biohydrogen [73], just as this carbohydrate-rich biomass facilitates the production of pyrolytic gas [74]. Furthermore, the application of polysaccharides, a type of carbohydrate, from microalgae has evolved in the food, sustainable agriculture, and biomedical sectors [75,76]. However, more studies should be conducted on biomass produced with phytohormone supplementation in outdoor conditions to verify these potential applications.
Implementing phytohormones in nonsterile open reactors operating at larger scales and under varying environmental conditions represents a notable research gap in phytohormone’s application in microalgal cultures [25]. This study effectively addresses this void by showcasing successful experiments conducted in such settings, marking a significant contribution to the field.
5. Conclusions
Different from other studies involving phytohormones and microalgae, this is the first study involving microalgae within an outdoor setting, utilizing raceway-type reactors and a nonsterile medium and supplemented with indole-3-acetic acid phytohormone, which has yielded promising outcomes. When contrasted with data acquired from an indoor environment lacking phytohormone supplementation, the outdoor experiment with indole-3-acetic acid supplemented in Spirulina sp. LEB 18 cultivations demonstrated a notable increase of 122.5% and 130.9% in biomass concentration and productivity, respectively, resulting in a 55% reduced-cost product. Moreover, the carbohydrate concentration underwent a remarkable surge of 230%, a remarkable 697% boost in biomolecule productivity, and an 86% decrease in production costs. However, further studies are necessary to apply this technology on an industrial scale and usher in a new era of microalgae supplementation. Nonetheless, this study establishes that phytohormone supplementation represents a viable industrial strategy for reducing microalgae production costs in an environmentally friendly manner and enhancing large-scale microalgae production. Hence, a significant gap in the research regarding phytohormone supplementation in microalgae cultivation has been addressed.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/app14093715/s1, Table S1: Price of reagents used to compose Zarrouk medium obtained on March 2023, from the manufacturer’s website (Sigma-Aldrich Brazil).
Author Contributions
Conceptualization, J.T.d.S., A.P.C.d.R., M.G.d.M. and J.A.V.C.; methodology, J.T.d.S. and A.P.C.d.R.; formal analysis and investigation, J.T.d.S.; writing—original draft preparation, J.T.d.S.; writing—review and editing, J.T.d.S., A.P.C.d.R., M.G.d.M. and J.A.V.C.; funding acquisition, J.A.V.C.; resources, J.A.V.C. and M.G.d.M.; supervision, A.P.C.d.R. and J.A.V.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded in part by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior—Brasil (CAPES)—Finance Code 001 and also had the support of the MCTI (Ministry of Science, Technology, and Innovation).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in the study are included in the article, further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Esteves, A.F.; Pires, J.C.M.; Gonçalves, A.L. Current Utilization of Microalgae in the Food Industry beyond Direct Human Consumption. In Cultured Microalgae for the Food Industry; Lafarga, T., Acién, G., Eds.; Elsevier: London, UK, 2021; pp. 199–248. [Google Scholar]
- Fernández, F.G.A.; Reis, A.; Wijffels, R.H.; Barbosa, M.; Verdelho, V.; Llamas, B. The Role of Microalgae in the Bioeconomy. New Biotechnol. 2021, 61, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Costa, J.A.V.; Freitas, B.C.B.; Rosa, G.M.; Moraes, L.; Morais, M.G.; Mitchell, B.G. Operational and Economic Aspects of Spirulina-Based Biorefinery. Bioresour. Technol. 2019, 292, 121946. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Cardenas, J.A.; Manzano-Agugliaro, F.; Acien-Fernandez, F.G.; Molina-Grima, E. Microalgae Research Worldwide. Algal Res. 2018, 35, 50–60. [Google Scholar] [CrossRef]
- Borowitzka, M.A.; Vonshak, A. Scaling up Microalgal Cultures to Commercial Scale. Eur. J. Phycol. 2017, 52, 407–418. [Google Scholar] [CrossRef]
- Belay, A. Biology and Industrial Production of Arthrospira (Spirulina). In Handbook of Microalgal Culture: Applied Phycology and Biotechnology; Richmond, A., Hu, Q., Eds.; Blackwell Science: Oxford, UK, 2013; pp. 339–358. [Google Scholar]
- Morais, M.G.; Radmann, E.M.; Andrade, M.R.; Teixeira, G.G.; Brusch, L.R.F.; Costa, J.A.V. Pilot Scale Semicontinuous Production of Spirulina Biomass in Southern Brazil. Aquaculture 2009, 294, 60–64. [Google Scholar] [CrossRef]
- Jiang, L.; Yu, S.; Chen, H.; Pei, H. Enhanced Phycocyanin Production from Spirulina Subsalsa via Freshwater and Marine Cultivation with Optimized Light Source and Temperature. Bioresour. Technol. 2023, 378, 129009. [Google Scholar] [CrossRef] [PubMed]
- Cruz, C.G.; Rosa, A.P.C.; Costa, J.A.V. Identification of the Phytohormones Indole-3-acetic Acid and trans-Zeatin in Microalgae. J. Chem. Technol. Biotechnol. 2023, 98, 1048–1056. [Google Scholar] [CrossRef]
- Ahmad, A.M.R.; Intikhab, A.; Zafar, S.; Farooq, U.; Shah, H.B.U.; Akram, S.; Abid, J.; Parveen, Z.; Iqbal, S. Spirulina, an FDA-Approved Functional Food: Worth the Hype? Cell Mol. Biol. 2023, 69, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Ragusa, I.; Nardone, G.N.; Zanatta, S.; Bertin, W.; Amadio, E. Spirulina for Skin Care: A Bright Blue Future. Cosmetics 2021, 8, 7. [Google Scholar] [CrossRef]
- Shao, W.; Ebaid, R.; El-Sheekh, M.; Abomohra, A.; Eladel, H. Pharmaceutical Applications and Consequent Environmental Impacts of Spirulina (Arthrospira): An Overview. Grasas Aceites 2019, 70, 292. [Google Scholar] [CrossRef]
- Sivaramakrishnan, R.; Suresh, S.; Kanwal, S.; Ramadoss, G.; Ramprakash, B.; Incharoensakdi, A. Microalgal Biorefinery Concepts’ Developments for Biofuel and Bioproducts: Current Perspective and Bottlenecks. Int. J. Mol. Sci. 2022, 23, 2623. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Cheah, W.Y.; Ng, I.-S.; Chang, J.-S.; Zhao, M.; Show, P.L. Microalgae-Based Biotechnological Sequestration of Carbon Dioxide for Net Zero Emissions. Trends Biotechnol. 2022, 40, 1439–1453. [Google Scholar] [CrossRef] [PubMed]
- Silveira, J.T.; Rosa, A.P.C.; Costa, J.A.V. Modulating Phytohormone Supplementation Can Efficiently Increase Biomass and Lipid Production in Spirulina (Arthrospira). Bioenergy Res. 2022, 15, 112–120. [Google Scholar] [CrossRef]
- Silveira, J.T.; Rosa, A.P.C.; Morais, M.G.; Costa, J.A.V. Cost Reduction in the Production of Spirulina Biomass and Biomolecules from Indole-3-Acetic Acid Supplementation in Different Growth Phases. Appl. Biochem. Biotechnol. 2023, 195, 2882–2892. [Google Scholar] [CrossRef] [PubMed]
- Cooke, T.J.; Poli, D.; Sztein, A.E.; Cohen, J.D. Evolutionary Patterns in Auxin Action. Plant Mol. Biol. 2002, 49, 319–338. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.-M.; Xiong, J.-Q. Boosting the Yields of Microalgal Biomass and High-Value Added Products by Phytohormones: A Mechanistic Insight Using Transcriptomics. J. Clean. Prod. 2023, 393, 136175. [Google Scholar] [CrossRef]
- Chernyad’ev, I.I. The Protective Action of Cytokinins on the Photosynthetic Machinery and Productivity of Plants under Stress (Review). Appl. Biochem. Microbiol. 2009, 45, 351–362. [Google Scholar] [CrossRef]
- Lu, Y.; Xu, J. Phytohormones in Microalgae: A New Opportunity for Microalgal Biotechnology? Trends Plant Sci. 2015, 20, 273–282. [Google Scholar] [CrossRef]
- Chokshi, K.; Pancha, I.; Ghosh, A.; Mishra, S. Nitrogen Starvation-Induced Cellular Crosstalk of ROS-Scavenging Antioxidants and Phytohormone Enhanced the Biofuel Potential of Green Microalga Acutodesmus dimorphus. Biotechnol. Biofuels 2017, 10, 60. [Google Scholar] [CrossRef]
- Kozlova, T.A.; Hardy, B.P.; Krishna, P.; Levin, D.B. Effect of Phytohormones on Growth and Accumulation of Pigments and Fatty Acids in the Microalgae Scenedesmus quadricauda. Algal Res. 2017, 27, 325–334. [Google Scholar] [CrossRef]
- Chen, J.; Wei, D.; Lim, P.-E. Enhanced Coproduction of Astaxanthin and Lipids by the Green Microalga Chromochloris zofingiensis: Selected Phytohormones as Positive Stimulators. Bioresour. Technol. 2020, 295, 122242. [Google Scholar] [CrossRef] [PubMed]
- Vijay, A.K.; Salim, S.A.M.; Prabha, S.; George, B. Exogenous Carbon Source and Phytohormone Supplementation Enhanced Growth Rate and Metabolite Production in Freshwater Microalgae Scenedesmus Obtusus Meyen. Bioresour. Technol. Rep. 2021, 14, 100669. [Google Scholar] [CrossRef]
- Stirk, W.A.; van Staden, J. Potential of Phytohormones as a Strategy to Improve Microalgae Productivity for Biotechnological Applications. Biotechnol. Adv. 2020, 44, 107612. [Google Scholar] [CrossRef] [PubMed]
- Slade, R.; Bauen, A. Micro-Algae Cultivation for Biofuels: Cost, Energy Balance, Environmental Impacts and Future Prospects. Biomass Bioenergy 2013, 53, 29–38. [Google Scholar] [CrossRef]
- Morais, M.G.; da Reichert, C.; Dalcanton, F.; Durante, A.J.; Marins, L.F.; Costa, J.A.V. Isolation and Characterization of a New Arthrospira Strain. Z. Naturforschung C 2008, 63, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Costa, J.A.V.; Colla, M.; Duarte Filho, P.; Kabke, K.; Weber, A. Modelling of Spirulina platensis Growth in Fresh Water Using Response Surface Methodology. World J. Microbiol. Biotechnol. 2002, 18, 603–607. [Google Scholar] [CrossRef]
- Zarrouk, C. Contributionà l Étuded Une Cyanophycée: Influence de Divers Facteurs Physiques et Chimiques Sur La Croissance et Photosynthese de Spirulina Maxima Geitler. Ph.D. Thesis, Faculté des Sciences de L’université de Paris, Paris, France, 1966. [Google Scholar]
- Andrade, M.R.; Costa, J.A.V. Outdoor and Indoor Cultivation of Spirulina platensis in the Extreme South of Brazil. Z. Naturforschung C 2008, 63, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Marsh, J.B.; Weinstein, D.B. Simple Charring Method for Determination of Lipids. J. Lipid Res. 1966, 7, 574–576. [Google Scholar] [CrossRef] [PubMed]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein Measurement with the Folin Phenol Reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Prates, D.F.; Radmann, E.M.; Duarte, J.H.; Morais, M.G.; Costa, J.A.V. Spirulina Cultivated under Different Light Emitting Diodes: Enhanced Cell Growth and Phycocyanin Production. Bioresour. Technol. 2018, 256, 38–43. [Google Scholar] [CrossRef]
- Bennett, A.; Bogorad, L. Complementary Chromatic Adaptation in a Filamentous Blue-Green Alga. J. Cell Biol. 1973, 58, 419–435. [Google Scholar] [CrossRef] [PubMed]
- Abalde, J.; Betancourt, L.; Torres, E.; Cid, A.; Barwell, C. Purification and Characterization of Phycocyanin from the Marine Cyanobacterium Synechococcus sp. IO9201. Plant Sci. 1998, 136, 109–120. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of AOAC International, 16th ed.; AOAC International: Arlington, VA, USA, 1995. [Google Scholar]
- Park, W.-K.; Yoo, G.; Moon, M.; Kim, C.W.; Choi, Y.-E.; Yang, J.-W. Phytohormone Supplementation Significantly Increases Growth of Chlamydomonas reinhardtii Cultivated for Biodiesel Production. Appl. Biochem. Biotechnol. 2013, 171, 1128–1142. [Google Scholar] [CrossRef]
- Babu, A.G.; Wu, X.; Kabra, A.N.; Kim, D.-P. Cultivation of an Indigenous Chlorella sorokiniana with Phytohormones for Biomass and Lipid Production under N-Limitation. Algal Res. 2017, 23, 178–185. [Google Scholar] [CrossRef]
- Yu, Z.; Pei, H.; Jiang, L.; Hou, Q.; Nie, C.; Zhang, L. Phytohormone Addition Coupled with Nitrogen Depletion Almost Tripled the Lipid Productivities in Two Algae. Bioresour. Technol. 2018, 247, 904–914. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Pei, H.; Li, Y.; Yang, Z.; Xie, Z.; Hou, Q.; Nie, C. Inclined Algal Biofilm Photobioreactor (IABPBR) for Cost-Effective Cultivation of Lipid-Rich Microalgae and Treatment of Seawater-Diluted Anaerobically Digested Effluent from Kitchen Waste with the Aid of Phytohormones. Bioresour. Technol. 2020, 315, 123761. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, H.-P.; Han, B.; Yu, X. Coupling of Abiotic Stresses and Phytohormones for the Production of Lipids and High-Value by-Products by Microalgae: A Review. Bioresour. Technol. 2019, 274, 549–556. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, X.; Han, B.; Zhao, Y.; Li, T.; Zhao, P.; Yu, X. Improvement in Lipid Production in Monoraphidium sp. QLY-1 by Combining Fulvic Acid Treatment and Salinity Stress. Bioresour. Technol. 2019, 294, 122179. [Google Scholar] [CrossRef]
- Chang, W.; Li, Y.; Qu, Y.; Liu, Y.; Zhang, G.; Zhao, Y.; Liu, S. Mixotrophic Cultivation of Microalgae to Enhance the Biomass and Lipid Production with Synergistic Effect of Red Light and Phytohormone IAA. Renew. Energy 2022, 187, 819–828. [Google Scholar] [CrossRef]
- Fierli, D.; Barone, M.E.; Donnell, A.M.; Conlon, T.; Touzet, N. Combined Application of Exogenous Phytohormones and Blue Light Illumination to the Marine Diatom Phaeodactylum tricornutum. Algal Res. 2023, 71, 103052. [Google Scholar] [CrossRef]
- Sánchez, J.F.; Fernández, J.M.; Acién, F.G.; Rueda, A.; Pérez-Parra, J.; Molina, E. Influence of Culture Conditions on the Productivity and Lutein Content of the New Strain Scenedesmus almeriensis. Process Biochem. 2008, 43, 398–405. [Google Scholar] [CrossRef]
- Ferreira, V.S.; Pinto, R.F.; Sant’Anna, C. Low Light Intensity and Nitrogen Starvation Modulate the Chlorophyll Content of Scenedesmus dimorphus. J. Appl. Microbiol. 2016, 120, 661–670. [Google Scholar] [CrossRef]
- Ota, M.; Kato, Y.; Watanabe, H.; Watanabe, M.; Sato, Y.; Smith, R.L.; Inomata, H. Fatty Acid Production from a Highly CO2 Tolerant Alga, Chlorocuccum littorale, in the Presence of Inorganic Carbon and Nitrate. Bioresour. Technol. 2009, 100, 5237–5242. [Google Scholar] [CrossRef] [PubMed]
- Shiraiwa, Y.; Goyal, A.; Tolbert, N.E. Alkalization of the Medium by Unicellular Green Algae during Uptake Dissolved Inorganic Carbon. Plant Cell Physiol. 1993, 34, 649–657. [Google Scholar] [CrossRef]
- Deamici, K.M.; Santos, L.O.; Costa, J.A.V. Magnetic Field Action on Outdoor and Indoor Cultures of Spirulina: Evaluation of Growth, Medium Consumption and Protein Profile. Bioresour. Technol. 2018, 249, 168–174. [Google Scholar] [CrossRef]
- Ho, S.-H.; Chen, C.-Y.; Lee, D.-J.; Chang, J.-S. Perspectives on Microalgal CO2-Emission Mitigation Systems—A Review. Biotechnol. Adv. 2011, 29, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Renuka, N.; Guldhe, A.; Singh, P.; Ansari, F.A.; Rawat, I.; Bux, F. Evaluating the Potential of Cytokinins for Biomass and Lipid Enhancement in Microalga Acutodesmus obliquus under Nitrogen Stress. Energy Convers. Manag. 2017, 140, 14–23. [Google Scholar] [CrossRef]
- Cardoso, L.G.; Duarte, J.H.; Andrade, B.B.; Lemos, P.V.F.; Costa, J.A.V.; Druzian, J.I.; Chinalia, F.A. Spirulina sp. LEB 18 Cultivation in Outdoor Pilot Scale Using Aquaculture Wastewater: High Biomass, Carotenoid, Lipid and Carbohydrate Production. Aquaculture 2020, 525, 735272. [Google Scholar] [CrossRef]
- Oliveira, M.A.C.L.; Monteiro, M.P.C.; Robbs, P.G.; Leite, S.G.F. Growth and Chemical Composition of Spirulina maxima and Spirulina platensis Biomass at Different Temperatures. Aquac. Int. 1999, 7, 261–275. [Google Scholar] [CrossRef]
- Mata, S.N.; Santos, T.S.; Cardoso, L.G.; Andrade, B.B.; Duarte, J.H.; Costa, J.A.V.; Souza, C.O.; Druzian, J.I. Spirulina sp. LEB 18 Cultivation in a Raceway-Type Bioreactor Using Wastewater from Desalination Process: Production of Carbohydrate-Rich Biomass. Bioresour. Technol. 2020, 311, 123495. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, L.G.; Lombardi, A.T.; Silva, J.S.J.; Lemos, P.V.F.; Costa, J.A.V.; Souza, C.O.; Druzian, J.I.; Chinalia, F.A. Scaling-up Production of Spirulina sp. LEB 18 Grown in Aquaculture Wastewater. Aquaculture 2021, 544, 737045. [Google Scholar] [CrossRef]
- Salama, E.-S.; Kabra, A.N.; Ji, M.-K.; Kim, J.R.; Min, B.; Jeon, B.-H. Enhancement of Microalgae Growth and Fatty Acid Content under the Influence of Phytohormones. Bioresour. Technol. 2014, 172, 97–103. [Google Scholar] [CrossRef]
- Zaparoli, M.; Ziemniczak, F.G.; Mantovani, L.; Costa, J.A.V.; Colla, L.M. Cellular Stress Conditions as a Strategy to Increase Carbohydrate Productivity in Spirulina platensis. Bioenergy Res. 2020, 13, 1221–1234. [Google Scholar] [CrossRef]
- Chrismadha, T.; Satya, A.; Satya, I.A.; Rosidah, R.; Satya, A.D.M.; Pangestuti, R.; Harimawan, A.; Setiadi, T.; Chew, K.W.; Show, P.L. Outdoor Inclined Plastic Column Photobioreactor: Growth, and Biochemicals Response of Arthrospira Platensis Culture on Daily Solar Irradiance in a Tropical Place. Metabolites 2022, 12, 1199. [Google Scholar] [CrossRef]
- Ogbonda, K.H.; Aminigo, R.E.; Abu, G.O. Influence of Temperature and PH on Biomass Production and Protein Biosynthesis in a Putative Spirulina sp. Bioresour. Technol. 2007, 98, 2207–2211. [Google Scholar] [CrossRef] [PubMed]
- Jesus, C.S.; Uebel, L.S.; Costa, S.S.; Miranda, A.L.; Morais, E.G.; Morais, M.G.; Costa, J.A.V.; Nunes, I.L.; Ferreira, E.S.; Druzian, J.I. Outdoor Pilot-Scale Cultivation of Spirulina sp. LEB-18 in Different Geographic Locations for Evaluating Its Growth and Chemical Composition. Bioresour. Technol. 2018, 256, 86–94. [Google Scholar] [CrossRef]
- Andersen, R.A. Algal Culturing Techniques, 1st ed.; Academic Press: Burlington, NJ, USA, 2004. [Google Scholar]
- Pan, X.; Chang, F.; Kang, L.; Liu, Y.; Li, G.; Li, D. Effects of Gibberellin A3 on Growth and Microcystin Production in Microcystis aeruginosa (Cyanophyta). J. Plant Physiol. 2008, 165, 1691–1697. [Google Scholar] [CrossRef]
- Mansouri, H.; Talebizadeh, R. Effects of Indole-3-Butyric Acid on Growth, Pigments and UV-Screening Compounds in Nostoc linckia. Phycol. Res. 2017, 65, 212–216. [Google Scholar] [CrossRef]
- Mc Gee, D.; Archer, L.; Fleming, G.T.A.; Gillespie, E.; Touzet, N. The Effect of Nutrient and Phytohormone Supplementation on the Growth, Pigment Yields and Biochemical Composition of Newly Isolated Microalgae. Process Biochem. 2020, 92, 61–68. [Google Scholar] [CrossRef]
- Tiwari, S.; Patel, A.; Prasad, S.M. Phytohormone Up-Regulates the Biochemical Constituent, Exopolysaccharide and Nitrogen Metabolism in Paddy-Field Cyanobacteria Exposed to Chromium Stress. BMC Microbiol. 2020, 20, 206. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, S.; Patel, A.; Prasad, S.M. Auxin and Cytokinin Alleviate Chromium-Induced Oxidative Stress in Nostoc muscorum and Anabaena sp. by Modulating Ascorbate–Glutathione Cycle. J. Plant Growth Regul. 2022, 41, 2743–2758. [Google Scholar] [CrossRef]
- Walter, A.; Carvalho, J.C.; Soccol, V.T.; Faria, A.B.B.; Ghiggi, V.; Soccol, C.R. Study of Phycocyanin Production from Spirulina platensis under Different Light Spectra. Braz. Arch. Biol. Technol. 2011, 54, 675–682. [Google Scholar] [CrossRef]
- Soni, B.; Kalavadia, B.; Trivedi, U.; Madamwar, D. Extraction, Purification and Characterization of Phycocyanin from Oscillatoria quadripunctulata—Isolated from the Rocky Shores of Bet-Dwarka, Gujarat, India. Process Biochem. 2006, 41, 2017–2023. [Google Scholar] [CrossRef]
- Deamici, K.M.; Costa, J.A.V.; Santos, L.O. Magnetic Fields as Triggers of Microalga Growth: Evaluation of Its Effect on Spirulina sp. Bioresour. Technol. 2016, 220, 62–67. [Google Scholar] [CrossRef]
- Olaizola, M.; Duerr, E.O. Effects of Light Intensity and Quality on the Growth Rate and Photosynthetic Pigment Content of Spirulina platensis. J. Appl. Phycol. 1990, 2, 97–104. [Google Scholar] [CrossRef]
- Salama, E.-S.; Jeon, B.-H.; Chang, S.W.; Lee, S.; Roh, H.-S.; Yang, I.-S.; Kurade, M.B.; El-Dalatony, M.M.; Kim, D.-H.; Kim, K.-H.; et al. Interactive Effect of Indole-3-Acetic Acid and Diethyl Aminoethyl Hexanoate on the Growth and Fatty Acid Content of Some Microalgae for Biodiesel Production. J. Clean. Prod. 2017, 168, 1017–1024. [Google Scholar] [CrossRef]
- Silvello, M.A.C.; Gonçalves, I.S.; Azambuja, S.P.H.; Costa, S.S.; Silva, P.G.P.; Santos, L.O.; Goldbeck, R. Microalgae-Based Carbohydrates: A Green Innovative Source of Bioenergy. Bioresour. Technol. 2022, 344, 126304. [Google Scholar] [CrossRef]
- Morais, E.G.; Silveira, J.T.; Schüler, L.M.; Freitas, B.C.B.; Costa, J.A.V.; Morais, M.G.; Ferrer, I.; Barreira, L. Biomass Valorization via Pyrolysis in Microalgae-Based Wastewater Treatment: Challenges and Opportunities for a Circular Bioeconomy. J. Appl. Phycol. 2023, 35, 2689–2708. [Google Scholar] [CrossRef]
- Moreira, J.B.; Vaz, B.S.; Cardias, B.B.; Cruz, C.G.; Almeida, A.C.A.; Costa, J.A.V.; Morais, M.G. Microalgae Polysaccharides: An Alternative Source for Food Production and Sustainable Agriculture. Polysaccharides 2022, 3, 441–457. [Google Scholar] [CrossRef]
- Moreira, J.B.; Santos, T.D.; Cruz, C.G.; Silveira, J.T.; Carvalho, L.F.; Morais, M.G.; Costa, J.A.V. Algal Polysaccharides-Based Nanomaterials: General Aspects and Potential Applications in Food and Biomedical Fields. Polysaccharides 2023, 4, 371–389. [Google Scholar] [CrossRef]
Figure 1.
Concentration and productivity of protein (dry basis) of indoor and outdoor Spirulina sp. LEB 18 biomass supplemented with IAA and control experiments. C-I = indoor control culture; IAA-I = indoor cultivation supplemented with IAA; C-O = outdoor control culture; IAA-O = outdoor cultivation supplemented with IAA. Lowercase letters that are identical within columns signify no significant difference in the means of protein content at the 95% confidence level (p > 0.05). Similarly, uppercase letters that are identical on the line indicate that there is no significant difference in the means of protein productivity at the 95% confidence level (p > 0.05). Data are represented as the means ± standard deviation (n = 6).
Figure 1.
Concentration and productivity of protein (dry basis) of indoor and outdoor Spirulina sp. LEB 18 biomass supplemented with IAA and control experiments. C-I = indoor control culture; IAA-I = indoor cultivation supplemented with IAA; C-O = outdoor control culture; IAA-O = outdoor cultivation supplemented with IAA. Lowercase letters that are identical within columns signify no significant difference in the means of protein content at the 95% confidence level (p > 0.05). Similarly, uppercase letters that are identical on the line indicate that there is no significant difference in the means of protein productivity at the 95% confidence level (p > 0.05). Data are represented as the means ± standard deviation (n = 6).
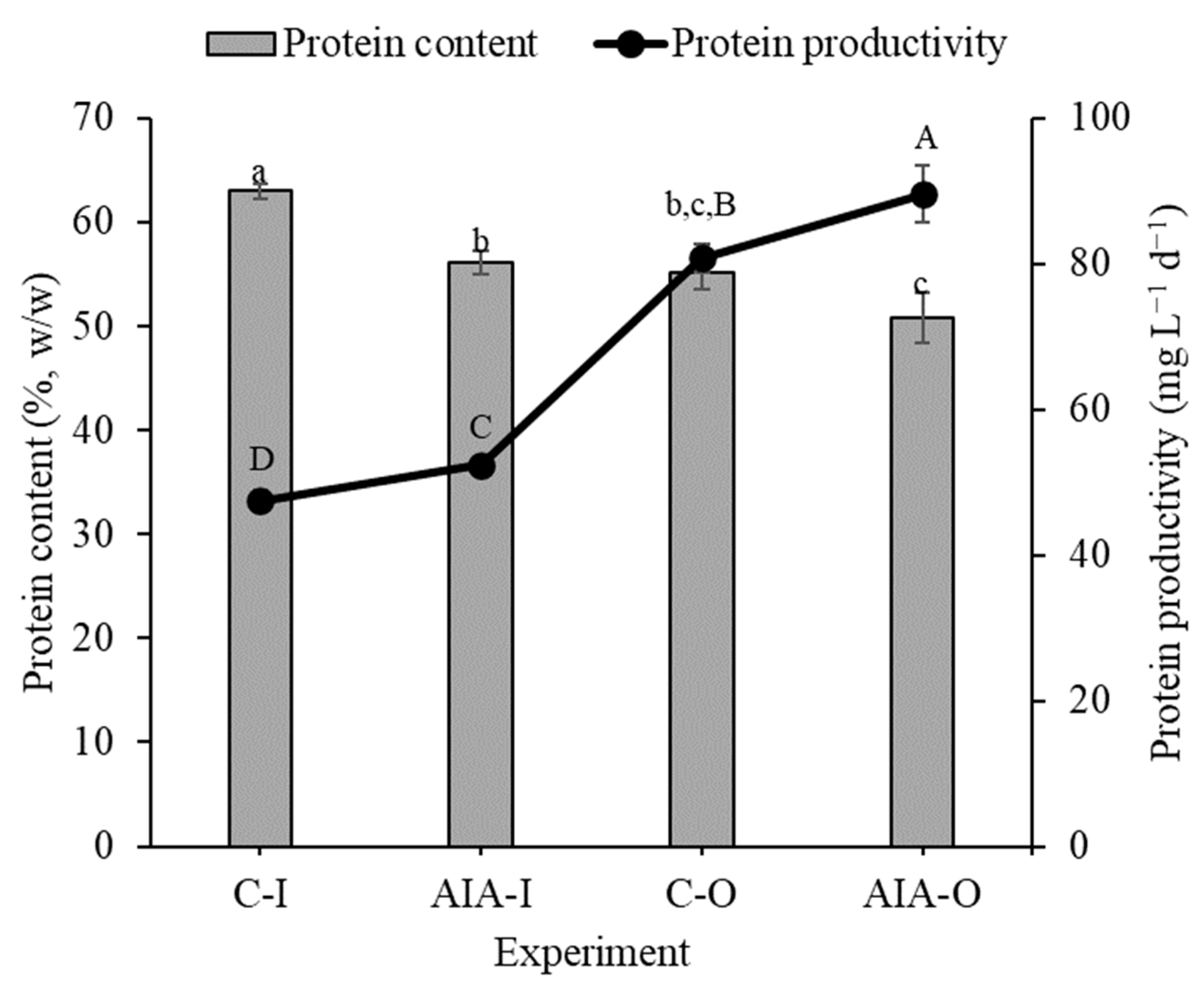
Figure 2.
Concentration and productivity of carbohydrates (dry basis) of indoor and outdoor Spirulina sp. LEB 18 biomass supplemented with IAA and control experiments. C-I = indoor control culture; IAA-I = indoor cultivation supplemented with IAA; C-O = outdoor control culture; IAA-O = outdoor cultivation supplemented with IAA. Lowercase letters that are identical within columns signify no significant difference in the means of carbohydrate content at the 95% confidence level (p > 0.05). Similarly, uppercase letters that are identical on the line indicate that there is no significant difference in the means of carbohydrate productivity at the 95% confidence level (p > 0.05). Data are represented as the means ± standard deviation (n = 6).
Figure 2.
Concentration and productivity of carbohydrates (dry basis) of indoor and outdoor Spirulina sp. LEB 18 biomass supplemented with IAA and control experiments. C-I = indoor control culture; IAA-I = indoor cultivation supplemented with IAA; C-O = outdoor control culture; IAA-O = outdoor cultivation supplemented with IAA. Lowercase letters that are identical within columns signify no significant difference in the means of carbohydrate content at the 95% confidence level (p > 0.05). Similarly, uppercase letters that are identical on the line indicate that there is no significant difference in the means of carbohydrate productivity at the 95% confidence level (p > 0.05). Data are represented as the means ± standard deviation (n = 6).
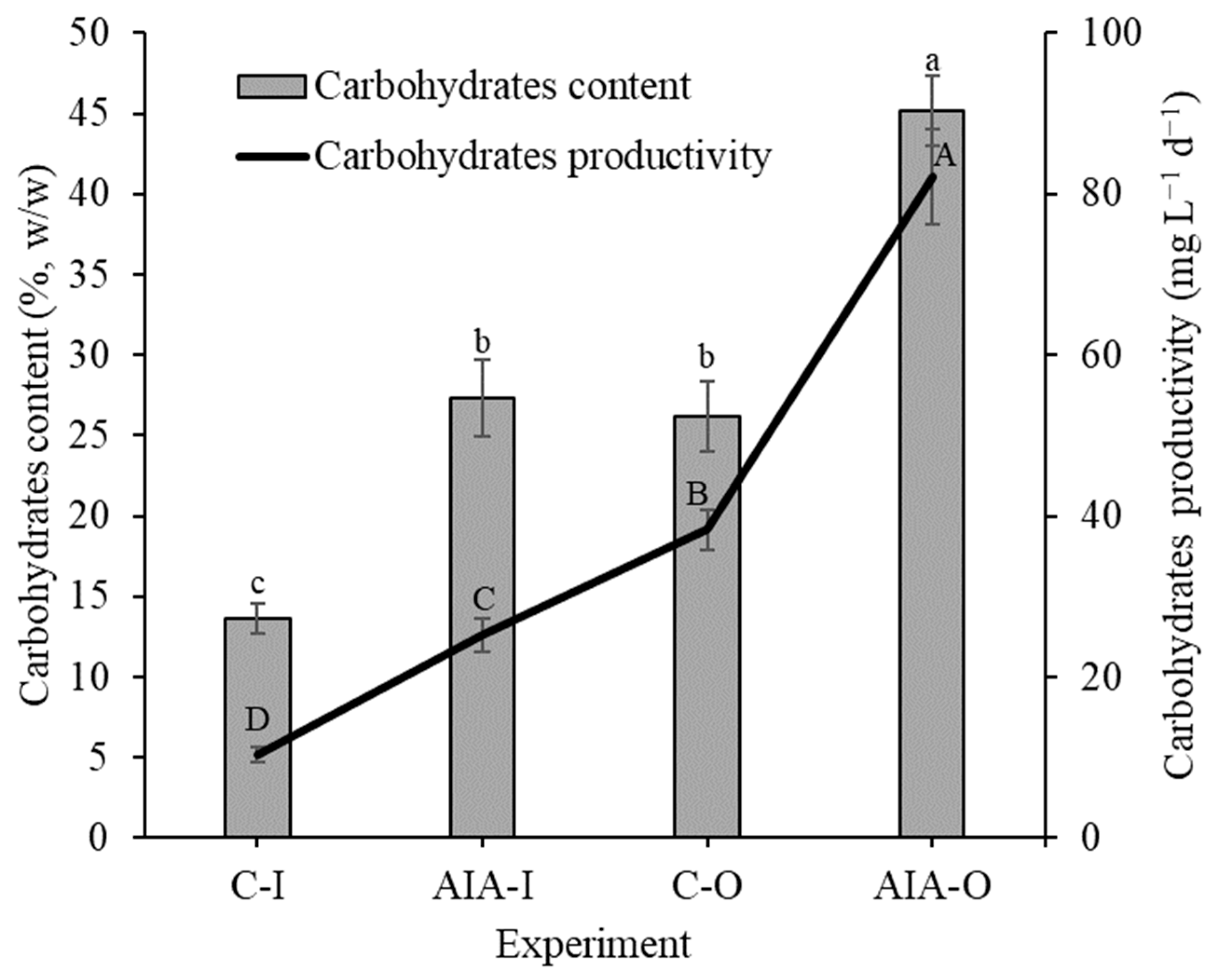
Figure 3.
Concentration and productivity of lipids (dry basis) of indoor and outdoor Spirulina sp. LEB 18 biomass supplemented with IAA and control experiments. C-I = indoor control culture; IAA-I = indoor cultivation supplemented with IAA; C-O = outdoor control culture; IAA-O = outdoor cultivation supplemented with IAA. Lowercase letters that are identical within columns signify no significant difference in the means of lipids content at the 95% confidence level (p > 0.05). Similarly, uppercase letters that are identical on the line indicate that there is no significant difference in the means of lipid productivity at the 95% confidence level (p > 0.05). Data are represented as the means ± standard deviation (n = 6).
Figure 3.
Concentration and productivity of lipids (dry basis) of indoor and outdoor Spirulina sp. LEB 18 biomass supplemented with IAA and control experiments. C-I = indoor control culture; IAA-I = indoor cultivation supplemented with IAA; C-O = outdoor control culture; IAA-O = outdoor cultivation supplemented with IAA. Lowercase letters that are identical within columns signify no significant difference in the means of lipids content at the 95% confidence level (p > 0.05). Similarly, uppercase letters that are identical on the line indicate that there is no significant difference in the means of lipid productivity at the 95% confidence level (p > 0.05). Data are represented as the means ± standard deviation (n = 6).
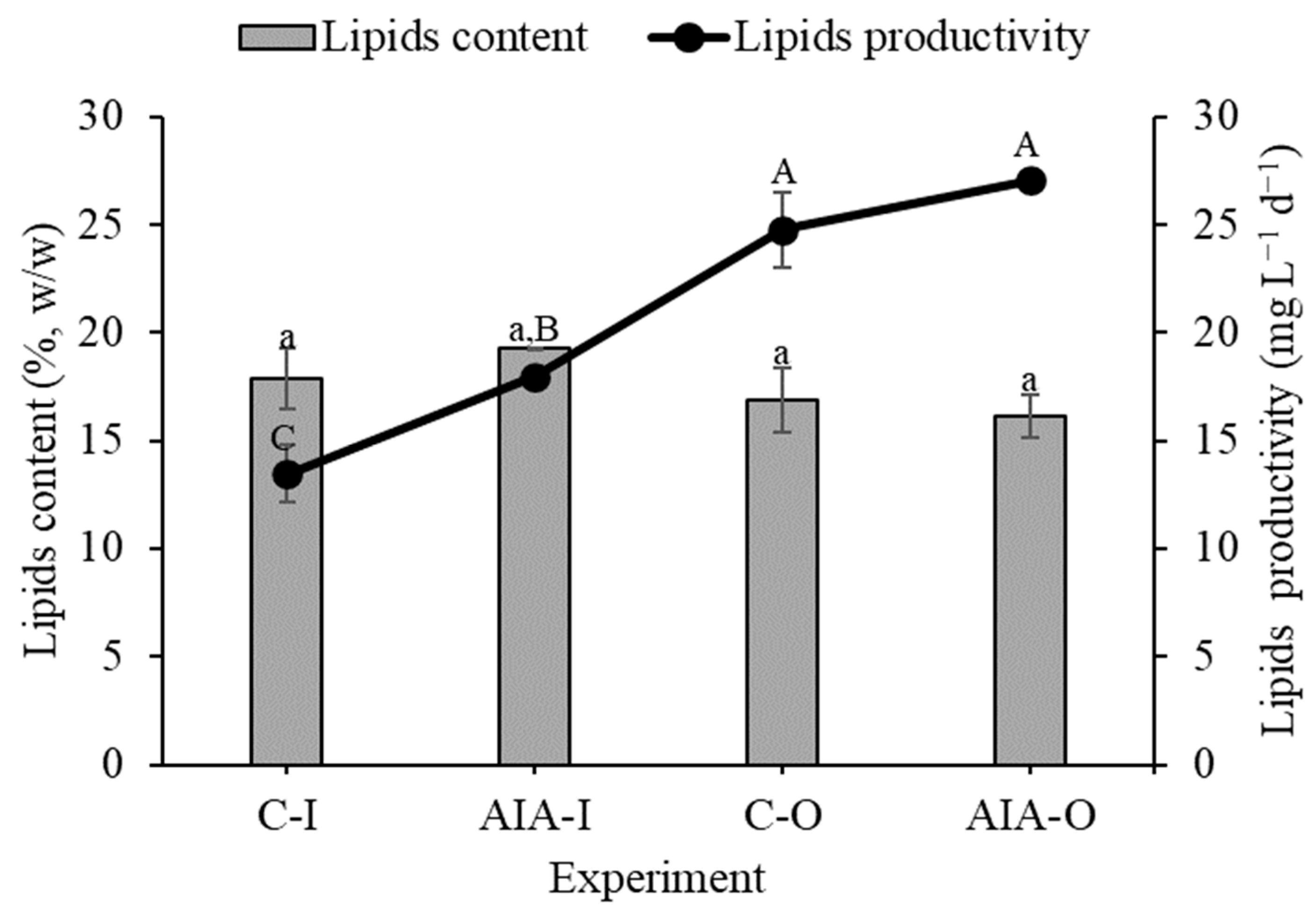
Figure 4.
Yield and productivity of phycocyanin (dry basis) of indoor and outdoor Spirulina sp. LEB 18 biomass supplemented with IAA and control experiments. C-I = indoor control culture; IAA-I = indoor cultivation supplemented with IAA; C-O = outdoor control culture; IAA-O = outdoor cultivation supplemented with IAA. Lowercase letters that are identical within columns signify no significant difference in the means of phycocyanin yield at the 95% confidence level (p > 0.05). Similarly, uppercase letters that are identical on the line indicate that there is no significant difference in the means of phycocyanin productivity at the 95% confidence level (p > 0.05). Data are represented as the means ± standard deviation (n = 6).
Figure 4.
Yield and productivity of phycocyanin (dry basis) of indoor and outdoor Spirulina sp. LEB 18 biomass supplemented with IAA and control experiments. C-I = indoor control culture; IAA-I = indoor cultivation supplemented with IAA; C-O = outdoor control culture; IAA-O = outdoor cultivation supplemented with IAA. Lowercase letters that are identical within columns signify no significant difference in the means of phycocyanin yield at the 95% confidence level (p > 0.05). Similarly, uppercase letters that are identical on the line indicate that there is no significant difference in the means of phycocyanin productivity at the 95% confidence level (p > 0.05). Data are represented as the means ± standard deviation (n = 6).
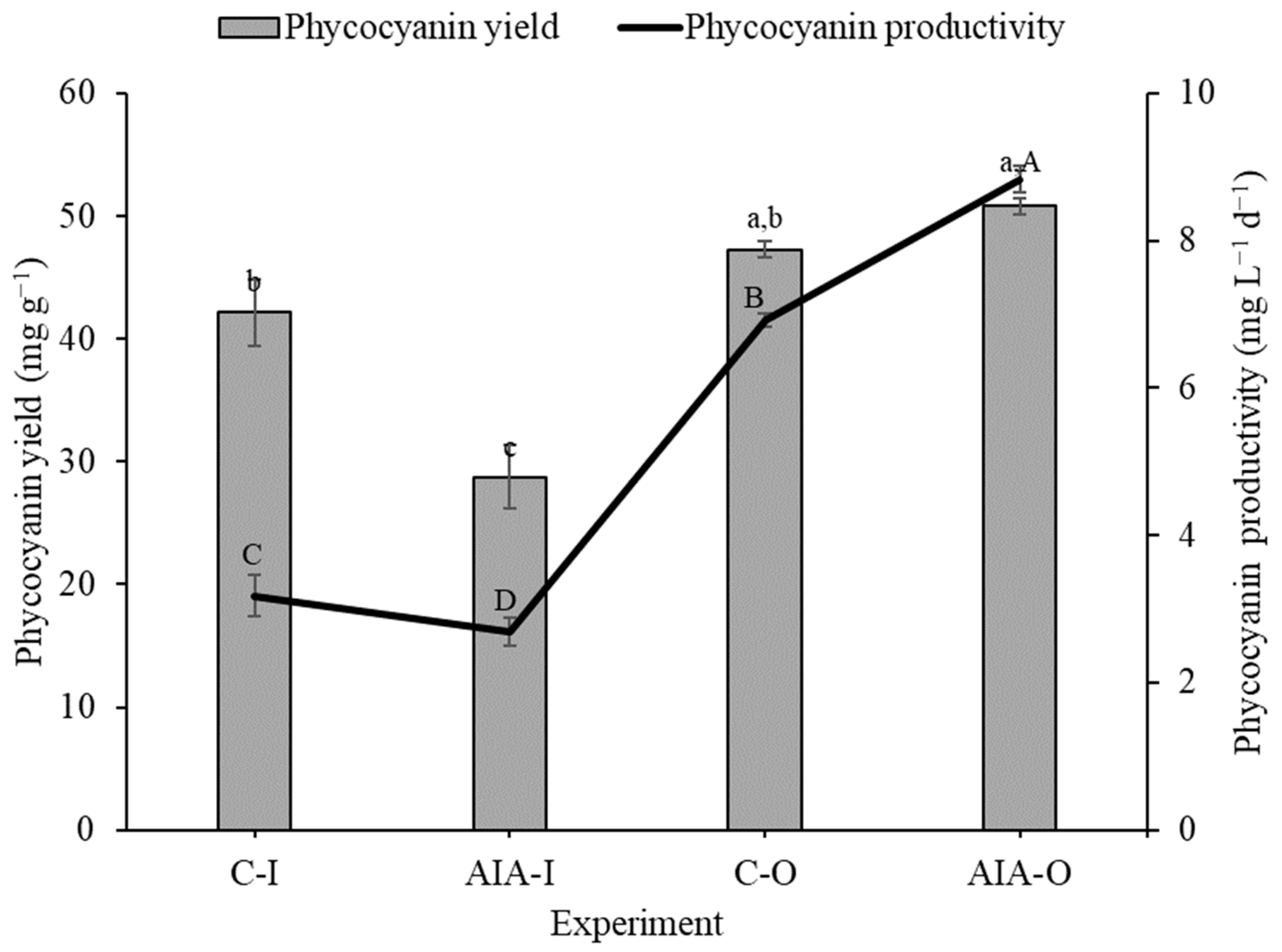
Figure 5.
Cost of biomass production and biomolecules produced in indoor and outdoor cultures of Spirulina sp. LEB 18 supplemented with IAA. C-I = indoor control culture; IAA-I = indoor culture supplemented with IAA; C-O = outdoor control culture; IAA-O = outdoor cultivation supplemented with IAA.
Figure 5.
Cost of biomass production and biomolecules produced in indoor and outdoor cultures of Spirulina sp. LEB 18 supplemented with IAA. C-I = indoor control culture; IAA-I = indoor culture supplemented with IAA; C-O = outdoor control culture; IAA-O = outdoor cultivation supplemented with IAA.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Final biomass concentration (Xfinal) and productivity (Pfinal), and maximum photochemical efficiency (Fv/Fm) and pH of indoor and outdoor Spirulina sp. LEB 18 supplemented with IAA and control experiments.
Table 1.
Final biomass concentration (Xfinal) and productivity (Pfinal), and maximum photochemical efficiency (Fv/Fm) and pH of indoor and outdoor Spirulina sp. LEB 18 supplemented with IAA and control experiments.
| Parameters | Indoor | Outdoor | ||
|---|---|---|---|---|
| C-I | IAA-I | C-O | IAA-O | |
| Xfinal (g L−1) | 2.44 c ± 0.09 | 2.78 c ± 0.29 | 4.60 b ± 0.01 | 5.43 a ± 0.07 |
| Pfinal (mg L−1 d−1) | 75.3 d ± 2.9 | 93.4 c ± 1.2 | 146.5 b ± 0.4 | 173.9 a ± 2.1 |
| Fv/Fm initial | 0.435 a ± 0.01 | 0.440 a ± 0.01 | 0.445 a ± 0.01 | 0.430 a ± 0.01 |
| Fv/Fm final | 0.385 b ± 0.01 | 0.385 b ± 0.01 | 0.450 a ± 0.01 | 0.440 a ± 0.01 |
| pH initial | 9.13 a ± 0.01 | 9.12 a ± 0.01 | 9.09 a ± 0.03 | 9.12 a ± 0.01 |
| pH final | 10.16 b ± 0.01 | 10.18 b ± 0.01 | 11.13 a ± 0.01 | 11.14 a ± 0.05 |
C-I = indoor control culture; IAA-I = indoor cultivation supplemented with IAA; C-O = outdoor control culture; IAA-O = outdoor cultivation supplemented with IAA. Identical superscript letters in the same row indicate that the means do not differ significantly at the 95% confidence level (p > 0.05). Data are presented as the means ± standard deviation (n = 6).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Silveira, J.T.d.; Rosa, A.P.C.d.; Morais, M.G.d.; Costa, J.A.V. Indole-3-Acetic Acid Action in Outdoor and Indoor Cultures of Spirulina in Open Raceway Reactors. Appl. Sci. 2024, 14, 3715. https://doi.org/10.3390/app14093715
AMA Style
Silveira JTd, Rosa APCd, Morais MGd, Costa JAV. Indole-3-Acetic Acid Action in Outdoor and Indoor Cultures of Spirulina in Open Raceway Reactors. Applied Sciences. 2024; 14(9):3715. https://doi.org/10.3390/app14093715
Chicago/Turabian StyleSilveira, Jéssica Teixeira da, Ana Priscila Centeno da Rosa, Michele Greque de Morais, and Jorge Alberto Vieira Costa. 2024. "Indole-3-Acetic Acid Action in Outdoor and Indoor Cultures of Spirulina in Open Raceway Reactors" Applied Sciences 14, no. 9: 3715. https://doi.org/10.3390/app14093715
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.