1. Introduction
The production of food manufacturing wastes (FMWs) is potentially damaging to the environment; these materials, if not properly disposed of, can lead to the release of greenhouse gases or contaminate soil and water as they decay. Biotechnological approaches may be one solution to utilize these materials before they cause damage to the environment [
1,
2].
In Chile, three important waste materials generated from the food manufacturing industry are olive-mill solid waste (OMSW), tomato pomace (TP), and grape pomace (GP). OMSW is a sub-product of olive oil production by two-phase centrifugation. It is composed of a liquid fraction also known as “alpechin” and a semisolid residue known as “alperujo”. The chemical characterization of “alperujo” reveals a high moisture content, slightly acidic pH, and high content of organic matter, mainly consisting of lignin, hemicellulose, and cellulose. Furthermore, “alperujo” contains lipids and water-soluble polyphenols [
3,
4]. TP is a sub-product of tomato processing which mainly consists of peels and seeds and is composed of lignocellulosic material, proteins, minerals, and lipids [
5]. GP is a sub-product of grape processing, mainly composed of lignocellulosic residues, glucan, alcohol, phenols, and flavonoids [
6].
One of the simplest strategies for disposal of these FMWs is biodegradation through enzymatic attack. FMWs are mainly composed of lignocellulose, of which cellulose is the major component (20–50%
w/w) [
7]. In nature, a wide range of microorganisms can use cellulose as a carbon and energy source. To accomplish this, fungi and bacteria secrete extracellular enzymes able to degrade this complex polymer [
8]. Cellulose is hydrolyzed by three different classes of enzymes: endo-β-1,4-D-glucanases (EC 3.2.1.4), which hydrolyze internal β-1,4-glucosidic linkages, 1,4-β-D-glucan cellobiohydrolases (EC 3.2.1.91), which cleave off cellulose fragments from the cellulose chain ends, generating the cellobiose disaccharide and β-D-glucosidases (EC 3.2.1.21), which use cellobiose hydrolysis to generate glucose [
9].
Cellulases have been applied in many different industries, such as food, textiles, pulp, paper and wastewater treatment of residue effluents [
10,
11]. However, one of the most significant current applications is in bioethanol production from lignocellulosic materials. Bioethanol is considered a fuel alternative for reducing the dependence on crude oil [
7]. The majority of bioethanol produced globally is made from biomass containing starch (from corn) or sucrose (from molasses or cane juice) [
12]. However, bioethanol can also be made from non-edible lignocellulosic biomass like wood or agricultural residues [
13].
Generally, the production of bioethanol from lignocellulosic biomass requires several steps: pretreatment, enzymatic hydrolysis to monomers (also called saccharification), fermentation, distillation, and dehydration [
14]. The pretreatment phase can be performed using several different processes, such as concentrated acid hydrolysis, dilute acid hydrolysis, alkali treatment, sodium sulfite treatment, and steam explosion, among others [
15]. The objective of the pretreatment is to remove lignin and improve the access of the enzymes to the cell wall carbohydrates (mainly cellulose) to obtain fermentable sugars [
16]. The next step is saccharification, in which cellulases, typically from
Trichoderma spp., are applied to produce fermentable monomer sugars from polysaccharides, primarily cellulose [
14]. After saccharification, sugar monomers can be fermented to ethanol, typically with
Saccharomyces spp., resulting in concentrations of 40–50 g/L [
17].
Almost all commercially available lignocellulolytic enzymes needed for the hydrolysis step are produced by fermentation with the ascomycete
Trichoderma reesei [
18]. However, the purchase of this enzymes can make up 20–40% of the operating costs in a lignocellulosic biomass-based biorefinery. Accordingly, enzyme cost is a major determinant of the viability of commercial-scale production of bioethanol [
19,
20]. Onsite production of enzymes using low-value substrates could be an option to make a bioethanol facility more economical. In addition, using FMWs materials as a substrate for fungal cellulases production would make the biorefinery more environmentally sustainable.
In order to improve enzyme performance, extensive research has been carried out on the cellulolytic system of
T. reesei to elucidate the enzymes and gene regulation involved in the utilization of lignocellulosic materials. Despite this, enzyme production from
T. reesei still faces two major bottlenecks. The first one is because some of the produced enzymes have low catalytic activity under industrial conditions, due to the presence of high concentrations of inhibitory compounds generated from the degradation of carbohydrates and lignin during the pretreatment of lignocellulosic biomass [
21,
22]. The second one is because certain key enzymes are present in limited amounts [
18,
23]. Most research to date has focused on overcoming these bottlenecks by improving the performance of
T. reesei secretion systems and investigating other
Trichoderma spp. with higher secretion potential. Few studies have centered on evaluating enzyme production using less expensive lignocellulosic raw materials [
24]. Consequently, the main objective of this research was to study the use of FMWs for cellulases production using an isolated fungal strain from olive stones. Production of cellulases from low-cost residues such as FMWS, could support future on-site production of cellulase enzymes from locally available waste materials for bioethanol production.
2. Materials and Methods
2.1. Substrate Preparation
FMWs were dried at 60 °C on a hot plate for at least 24 hours, avoiding burning. Subsequently, they were ground in a crushing machine and sieved through a 300 μm mesh (VWR, Radnor, PA, USA). This material was used at 0.5% as the carbon source for liquid and solid medium experiments.
2.2. Chemicals
Glycine, sodium hydroxide (NaOH), sodium deoxycholate (DOC), 3,5-dinitro-2-hydroxybenzoic acid (DNS), trichloroacetic acid (TCA), trifluoroacetic acid, ammonium carbonate, iodoacetamide, acetonitrile, potassium ferricyanide, sodium acetate, sodium thiosulfate, Congo red, carboxymethylcellulose, agar, hexadecyltrimethylammonium bromide (CTAB), glucose, acetic acid, NaCl, (NH4)2SO4, KH2PO4, MgSO4, FeSO4 x 7H2O, isopropanol, chloroform, Milli-Q water, isoamyl alcohol, acetone, and lactophenol (Merck, Darmstadt, Germany). CaCl2, ZnSO4, CoCl2, and MnSO4 (Phytotechnology Laboratories, Lenexa, KS, USA). Wizard® SV Gel and PCR Clean-Up System, PureYield™ Plasmid Miniprep System, pGEMT®-T Vector System I, and urea (Promega, Madison, WI, USA). Neopeptone, Luria Bertani (LB) broth, and potato dextrose agar (PDA) (Becton, Dickinson and Company, Franklin Lakes, NJ, USA). 4-nitrophenol-β-D-glucopyranoside (pNPG), 4-nitrophenol, Folin–Ciocalteu reagent, dithiothreitol, trypsin, and RNAse (Sigma-Aldrich, Missouri, WI, USA). Agarose (Fermelo Biotec, Santiago, Chile).
2.3. Fungal Strain and Culture Conditions
The fungal strain was isolated from olive stones kept at room temperature. Later, it was laid onto PDA plates. The plates were incubated at 30 °C for 1 week, and plugs of PDA were extracted from the edge of the mycelium and placed on a new PDA plate. This procedure was repeated several times to obtain homogeneous colony formation. For the mycelium staining procedure, the fungal slide procedure was used. Briefly, a plug of a 0.4 mm
2 PDA was deposited on a glass slide. The mycelium was deposited on the plug and covered with a microscope coverslip. After growth of the mycelium on the PDA plug and the surface of the coverslip, this was carefully removed, and the mycelium was stained with a solution of lactophenol cotton blue and observed using an optical microscope (Optika, Lombardy, Italy). Liquid cultures were performed in Mandels medium as described previously [
25]. In this step, a plate containing the mycelium was scraped using an inoculation loop. The mycelium was dissolved in an Erlenmeyer flask containing a sterile 0.9% NaCl solution and glass beads to facilitate spore release. After shaking, this solution was filtered using sterile dressing into an empty sterile flask. Spores were counted in a Petroff–Hauser chamber, and 4 × 10
6 spores were inoculated in 100 mL of Mandel medium supplemented with 0.5% FMWs as the only carbon source. For solid-medium experiments, a 0.4 mm
2 area of a PDA plug containing the mycelium was extracted and placed on the surface of plates containing Mandel medium, 4% agar, and 0.5% FMWs. Mycelium radial growth was monitored each day by measurement of the colony diameter [
26]. For the analysis of the presence of fungal cellulases on the plates, the Congo red protocol was performed according to Zeng et al. [
27].
2.4. Strain Identification
The mycelium was grown in LB media at 30 °C for 5 days. Then, 1 mL of culture was centrifuged, resuspended in 800 μL of CTAB solution, and subsequently lysed in Precellys® 24 (Bertin, Provenza, France) using 0.5 mm beads. After lysis, the tubes were kept at 65 °C for 30 min, and a mixture of chloroform/isoamyl alcohol was added. The samples were centrifuged at 13,000 rpm for 11 min at 25 °C. The supernatants were transferred to clean tubes, and an equal volume of cold isopropanol was added. The tubes were kept at −20 °C for 2 h, then centrifuged at 13,000 rpm for 11 min at 4 °C. The supernatants were discarded, and the pellets were air-dried at room temperature and then resuspended in Milli-Q water. RNAse (10 mg/mL) was added to the tubes, which were incubated at 37 °C for 30 min. One volume of chloroform/isoamyl alcohol was added, and the samples were centrifuged at 13,000 rpm for 10 min at 4 °C. The supernatants were extracted, and 2.5 volumes of cold ethanol were added to the tubes, which were kept at −20 °C for 30 min. The samples were centrifuged at 13,000 rpm for 10 min, and the pellets were air-dried at room temperature. DNA was resuspended in Milli-Q water, quantified using Nanodrop 2000 (Thermo Scientific, Delaware, USA), and visualized using 1% agarose gel electrophoresis. Aliquots were used in PCR reactions using ITS-1 (5′-TCCGTAGGTGAACCTGCGG-3′) and ITS-4 (5′-TCCTCCGCTTATTGATATGC-3′) primers [
26]. The obtained PCR fragment corresponding to the ITS1-5.8-ITS2 ribosomal region was purified using Wizard® SV Gel and PCR Clean-Up System, cloned in pGEMT®-T Vector System I, and used to transform JM101 competent cells. Positive colonies containing the fragment were verified with colony PCR using T7 (5′-TAATACGACTCACTATAGGG-3′) and SP6 (5-ATTTAGGTGACACTATAG-3′) primers. A white colony was inoculated in LB medium for plasmid DNA purification using PureYield™ Plasmid Miniprep System. An aliquot of this sample was sequenced using T7 and SP6 primers (Macrogen, Seoul, Korea). The sequence was edited with Editseq software (DNASTAR, Madison, WI, USA) and compared with sequences from GenBank using BLAST algorithm (Megablast) (
http://blast.ncbi.nlm.nih.gov/Blast.cgi). The sequences were aligned with ClustalW and analyzed with the MEGA-7 software [
28]. Phylogenetic analyses were carried out using the neighbor-joining method with the Kimura-2-parameter model [
29]. There were 569 positions in the final dataset. Bootstrap analyses were performed on 10,000 replicates.
2.5. Enzyme Activity
Filter paper assays were done using 50 mg of Whatman grade 1 paper introduced in a glass tube in the presence of 1 mL of 50 mM citrate buffer pH 5.0 and 0.5 mL of enzyme. The sample was incubated at 50 °C for 1 h, and reducing sugars were quantified by the DNS method. Glucose in the range 0.2 μg/μL to 2 μg/μL was used as a standard. The samples were measured at 540 nm [
30]. β-glucosidase activity was determined using 1.5 mg/mL of 4-nitrophenol-β-D-glucopyranoside (pNPG) in the presence of 20 mM sodium acetate buffer pH 3.5. Then, 250 μL of enzyme was added to the mixture, and the samples were incubated at 50 °C for 10 min. Then the reaction was stopped by addition of 500 μL of 0.4 M glycine/NaOH buffer, pH 10.7, and the mixture was measured at 405 nm. The standard curve was measured using 4-nitrophenol in the range of 15 to 75 μM. All activities were expressed in international units (U), corresponding to the quantity of enzyme hydrolyzing 1 μmol of the substrate in 1 min. Triplicate assays were performed. Total protein content was determined using the Lowry method modified by Tan [
31], using bovine serum albumin in the range of 0.02 μg/μL to 0.2 μg/μL as a standard and measuring samples at 660 nm. Specific enzyme activity in both filter paper and β-glucosidase activity assays is expressed as U/mg total protein.
2.6. Sample Preparation and SDS-PAGE
For the preparation of the extracellular protein fractions, cultures from the fifth day were harvested by centrifugation at 4000 rpm for 30 min at 4 °C, and the supernatants were filtered through a 0.22 μm filter (Millipore, Burlington, MA, USA). A total of 10 μg of protein was precipitated by the DOC–TCA method and collected by centrifugation (13,000 rpm, 5 min, 4 °C). The resulting pellet was washed two times with cold acetone and air-dried. The protein pellet was resuspended in loading buffer, boiled, and loaded onto 12% SDS-PAGE (denaturant electrophoresis). The gel was run at a constant voltage of 200 V for 2 h. After running, the gel was stained with silver according to Schevchenko et al. [
32]. Gels from two different biological replicates were used for sequencing by means of mass spectrometry.
2.7. Peptide Preparation for Mass Spectrometry
For each lane of gel, 10 zones were selected for cutting, and each excised band was distained with a 1:1 mixture of 30 mM potassium ferricyanide and 100 mM sodium thiosulfate. After washing with Milli-Q water, a mixture of 1:1 ammonium carbonate and acetonitrile was added for 2 h. Then, acetonitrile was added until drying of the gel piece, which was hydrated in 0.1 M ammonium carbonate for 5 min at room temperature. An equal volume of acetonitrile was added, and the sample was incubated for 15 min at 37 °C; then, the liquid was removed, and the tube was dried in a speed vac. Gel pieces were reduced with 10 mM dithiothreitol in 0.1 M ammonium carbonate at 56 °C for 45 min. The liquid was discarded, and the gel was incubated in 0.1 M ammonium carbonate and 55 mM iodoacetamide for 30 min. The gel was washed two times with a 1:1 mixture of ammonium carbonate and acetonitrile for 15 min and dried in a speed vac. The dried samples were incubated with 10 ng of trypsin in ammonium carbonate solution overnight. After digestion, the samples were incubated with a mixture of 25 mM ammonium carbonate and acetonitrile for 50 min in a sonicator bath. Then, the samples were incubated in mixtures composed of 30% acetonitrile/0.1% trifluoroacetic acid and 50% acetonitrile/0.1% trifluoroacetic acid. Each supernatant was collected and concentrated in the speed vac for 30 min. Finally, 20 μL of trifluoroacetic acid was added, and the solutions were purified by Ziptip (Millipore, Burlington, MA, USA).
2.8. Mass Spectrometry
Dried samples were dissolved in 10 μL of 2% acetonitrile/0.1% trifluoroacetic acid and applied to an Ultimate 3000 Nano-HPLC (Dionex, Idstein, Germany). Each sample was first trapped on a 1 mm PepMap-trapping column (Dionex, Idstein, Germany) for 10 min at 30 μL/min of 2% acetonitrile/0.1% trifluoroacetic acid and subsequently subjected to a 75 μm ID, 5 cm PepMap C18-column (Dionex, Idstein, Germany). Peptide separation was performed by an acetonitrile gradient at 300 nL/min. The separation column outlet was coupled online to a nano-spray interface of an Esquire HCT ETDII-ion-trap mass spectrometer (Bruker Daltonics, Bremen, Germany). Mass spectra were acquired in positive MS-mode, tuned for tryptic peptides. MS/MS-precursor selection was performed in an optimized automatic regime, with preference for double- and triple-charged ions. Every selected precursor was fragmented by collision-induced dissociation (CID) and electron transfer dissociation (ETD), respectively. MS/MS spectra were processed by the Data Analysis and BioTools software from Bruker. Combined CID/ETD-derived fragment lists were analyzed by the MASCOT algorithm in in-house and Swissprot databases, generating a presence–absence table. The total number of proteins belonging to the fungi kingdom were counted for each treatment. Cellulases were selected to construct a final table.
3. Results and Discussion
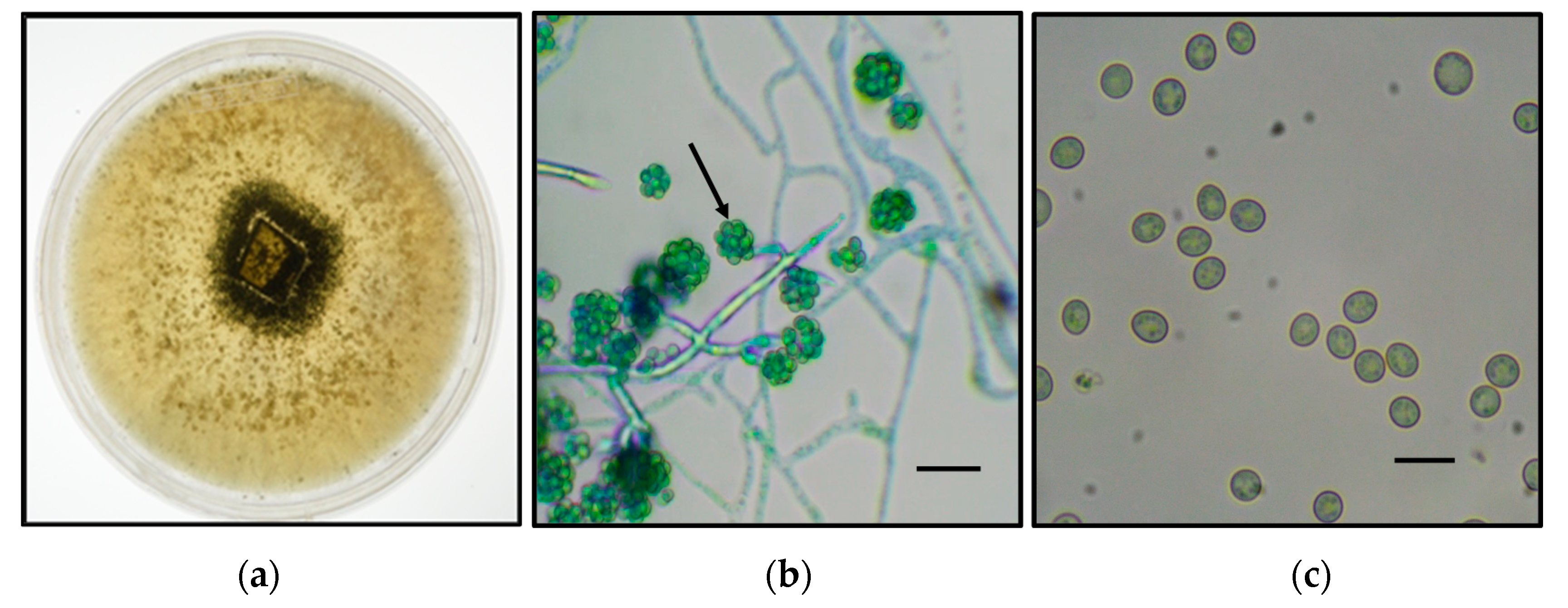
A fungal strain was isolated from olive stones kept at room temperature. After successive passages onto PDA plates, a homogenous fluffy-type mycelium of white color was observed which turned green after 5 days of cultivation (
Figure 1a). Examination of samples of the lactophenol-cotton-blue-stained mycelium under optical microscopy showed the presence of segmented hyphae, tree-like reproductive structure, and conidia showing round to ovoidal form. Similar features are found in
Trichoderma species (
Figure 1b,c) [
33].
PCR amplification using ITS-1 and ITS-4 primers and the mycelium DNA produced a fragment of around 570 bp (
Figure 2a). This fragment was cloned in pGEM-T vector and used for transformation of
Escherichia coli competent cells. After selection of colonies containing the recombinant plasmid, positive colonies were inoculated in LB medium for plasmid purification. Plasmids containing the PCR product were further sequenced using the T7 and SP6 vector primers. The amplified ITS1-5.8S-ITS2 sequence from strain FG displayed 100% homology to the same region of several strains of
Trichoderma asperellum associated with plants worldwide. Also, this region is 99% identical to that of
T. asperellum UC 1 (Genbank ID: MF774876) isolated in Malaysia, which is reported to be a high cellulase and xylanase producer [
34]. A phylogenetic tree was constructed using these data and related sequences extracted from the NCBI database, as shown in
Figure 2b. This figure shows that strain FG sequence was located in a clade together with other
Trichoderma species. These data suggest that strain FG belongs to
Trichoderma spp.
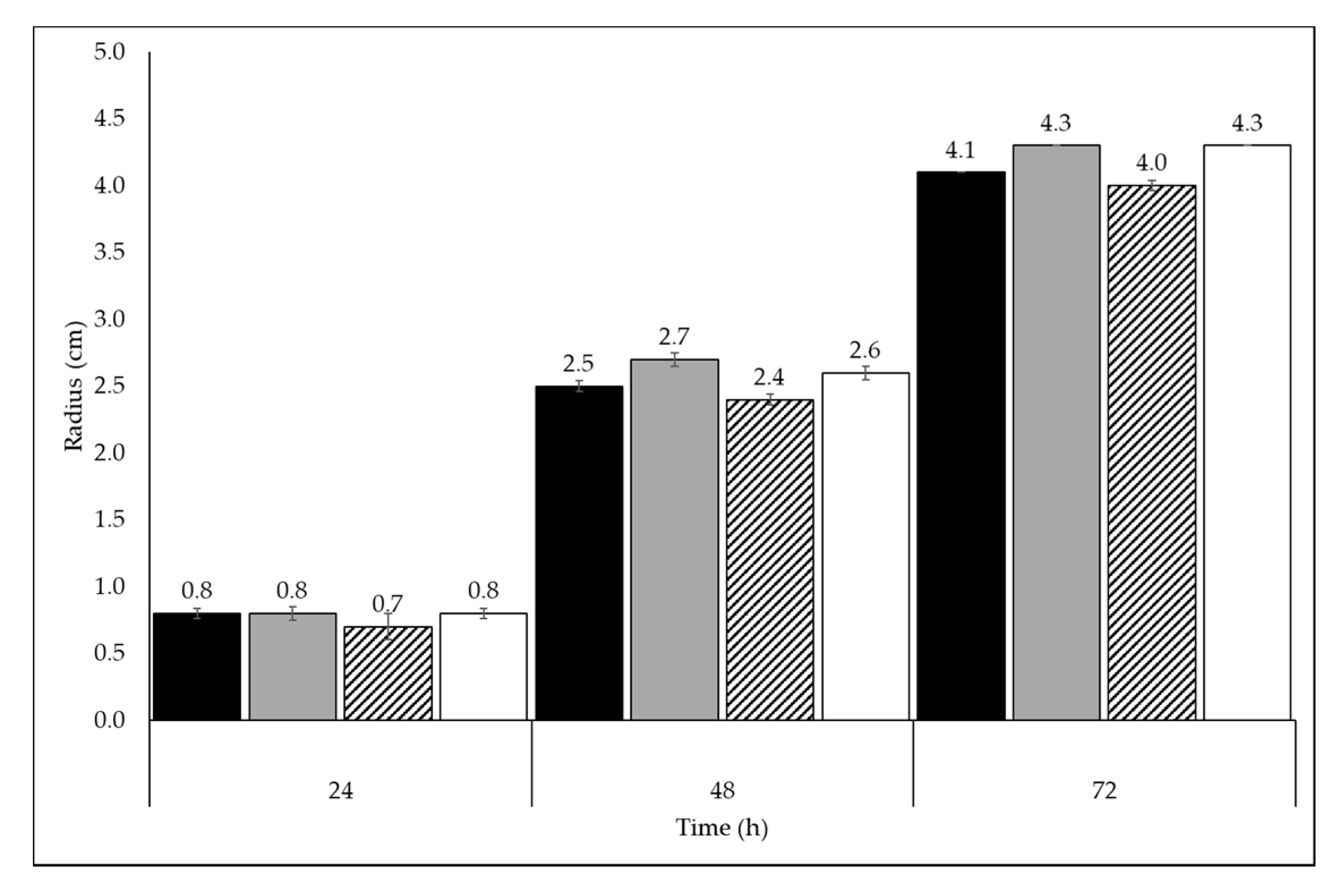
To evaluate the behavior of the
Trichoderma sp. on FMWs, the fungus was grown on both solid and liquid Mandel medium supplemented with 0.5% FMWs. For comparison purposes, glucose was added to a fourth set of liquid cultures and plates. On these plates,
Trichoderma sp. growth was observed on the three FMWs, however the highest radius value at 72 h was observed in OMSW and GP plates (4.3 cm) followed by glucose (4.1 cm) and TP (4.0 cm) (
Figure 3) plates.
The samples were incubated in 100 mL liquid cultures at 30 °C, and the total biomass (wet weight) obtained after 5 d of culture was 17.4 g for glucose, 11.0 g for OMSW, 9.8 g for G,P and 8.4 g for TP. In addition, the final protein concentrations in these samples were 0.6, 0.7, 0.3, and 0.4 mg/mL, respectively. The visual inspection of the FMW cultures on the fifth day revealed incomplete degradation; however, OMSW seemed to be the most preferable carbon source among the three FMWs.
Fungal growth was evidenced when the fungal strain was inoculated on plates containing LB medium with 1% carboxymethyl cellulose stained with Congo red solution. Initially, the plates were fully stained red. Once the cellulose was degraded after 5 d of incubation, the red color disappeared leaving a pale-colored halo, which was indicative of the presence of cellulases released into the culture medium (
Figure 4a). This preliminary evidence prompted us to search for cellulases when the fungi were grown in FMWs.
Culture supernatants prepared from FMWs liquid cultures after 5 d of fungal growth were evaluated for their total cellulase activity (i.e., endo-β-1,4-D-glucanase, 1,4-β-D-glucan cellobiohydrolase, and β-glucosidase) and β-glucosidase activities through filter paper and pNPG assays, respectively. Filter paper specific activity was clearly higher in the FMW culture supernatants in comparison to the glucose supernatant, with the GP culture supernatant having the highest value (
Figure 4b). The same trend was observed for β-glucosidase specific activities, although the TP supernatant had the highest value (
Figure 4c).
To confirm the presence of cellulases, proteins were extracted from the culture supernatants of the
Trichoderma sp. grown on glucose and FMWs on the fifth day. Proteins were separated by SDS-PAGE (denaturant electrophoresis) and analyzed by mass spectrometry. This qualitative analysis identified 336, 219, 244, and 210 proteins in the supernatant media from cells grown in glucose, OMSW, GP, and TP, respectively (results not shown). The identity of the proteins in each fraction is found in
Table 1, and a detailed description is given in
Supplementary Material (ST1–ST8). In total, 24 distinct cellulases were found in 4 four different residues. Endo-β-1,4-D-glucanase,1,4-β-D-glucan cellobiohydrolase and β-glucosidases were found in the culture supernatants of FMWs, whereas the glucose supernatants did not contain any exo- or endo-glucanases.
The ability of species belonging to genus
Trichoderma to grow in complex substrates has been previously reported. Using a synthetic medium, containing cellulose as the sole carbon source, Do Vale et al. identified few cellulases in the culture supernatant of
Trichoderma harzianum [
36]. In contrast, after growth of
T. harzianum using the same medium supplemented with sugarcane bagasse, Gomez-Mendoza et al. found that this complex substrate induced a higher secretion of cellulolytic and xylanolytic activities when compared with other substrates such as glucose [
37]. In a comparative study of the extracellular proteins produced by
T. asperellum S4F8 and
T. reesei Rut C30 after solid-state fermentation on the same substrate, Marx et al. found 14 and 12 cellulases, respectively [
38]. In a study using
Aspergillus niger and
T. reesei grown on sugarcane with two levels of cell wall complexity, culm in natura and pretreated, Borin et al. found that although
A. niger and
T. reesei both produced cellulases,
A. niger produced two times more enzymes than
T. reesei [
39].
In general, the activities from the culture supernatants agreed with the mass spectrometry data. Glucose was found repressive as expected; however, five β-glucosidases were still found in the culture supernatant of this simple substrate. These results are similar to those of Gómez-Mendoza et al. who found that
T. harzianum cultured with glucose presented two β-glucosidases [
37]. These enzymes might be responsible for the residual activities seen in
Figure 4c.
Only one endo-β-1,4-D-glucanase was found in our analysis. However, 14 1,4-β-D-glucan cellobiohydrolases were found in the culture supernatants from FMWs samples. The cellulolytic system of
T. reesei has been studied in detail. It is composed by seven endo-β-1,4-D-glucanases, two 1,4-β-D-glucan cellobiohydrolases, and seven β-D-glucosidases. The endo-β-1,4-D-glucanases belong to the GH5, GH7, GH12, and GH45 families, and the 1,4-β-D-glucan cellobiohydrolases belong to the GH6 and GH7 families. Finally, the β-glucosidases belongs to the GH3 family [
18,
23,
40]. In the cellulose degradation model by enzymes from
T. reesei, the enzyme endo-β-1,4-D-glucanases acts by cleaving off the amorphous regions of the chain, while the 1,4-β-D-glucan cellobiohydrolases cleave off at the nonreducing and reducing ends. The oligosaccharide products from this hydrolysis are further hydrolyzed into monosaccharides by
β-glucosidase into glucose. Other accessory enzymes such as lytic polysaccharide monooxygenases and swollenin, which expands cellulose chains to improve enzyme accessibility, were not found in our four culture conditions [
18,
23,
40].
From a biorefinery prospective, enzyme cost is one of the main drivers of the viability of commercial-scale production of bioethanol, representing around 20–40% of the operating costs in a lignocellulosic biomass-based biorefinery [
19,
22]. Using low-value substrates to produce enzymes onsite could be an option to make a bio-ethanol facility more sustainable and economical. Currently, most economic studies assume purchasing cellulases from an enzyme manufacturer [
19,
41,
42]. Although this approach avoids the capital cost of on-site enzyme production, it includes the transportation of the enzyme to the biorefinery facility, adding a non-trivial amount to the enzyme cost [
43]. Humbird et al. modeled on-site enzyme generation for ethanol production and found that the carbon source makes up 57% of the cost of enzyme production, followed by 21% capital cost, 13% of electricity, and 9% of other nutrients and fixed costs [
44]. Consequently, the utilization of less expensive carbon sources like agroindustrial residues such as OMSW, GP, and TP will allow reducing the cost of enzymes, thereby reducing ethanol production costs.
This investigation suggests that
Trichoderma sp. can grow on a wide range of carbon sources, including complex substrates such as FMWs. This enables a great variability in the production of cellulases and other enzymes (data not shown), since gene expression and enzyme secretion are directly dependent on the substrate being consumed. Accordingly, in FMWs, whose full chemical composition is still unknown, sugars released during fungal hydrolysis might induce or repress cellulase genes at different levels. Some cellulase inducers are cellulose, β-glucan, xylan, lactose, cellobiose, and sophorose; glucose is the main cellulase repressor [
40].

 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}