Effect of Glycerol Concentration and Light Intensity on Growth and Biochemical Composition of Arthrospira (Spirulina) Platensis: A Study in Semi-Continuous Mode with Non-Aseptic Conditions

 ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Cultivation Conditions
2.2. Experimental Set-Up
2.3. Analytical Methods
3. Results and Discussion
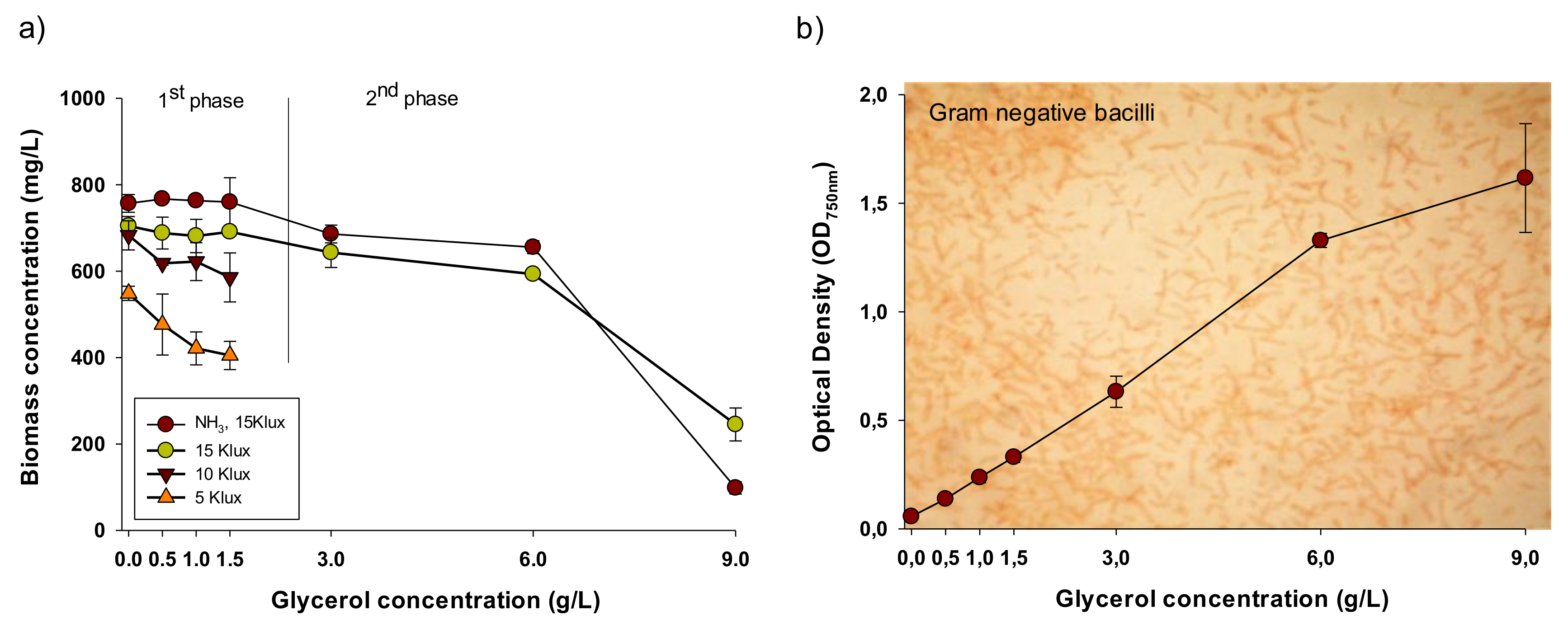
3.1. Effect of Light and Glycerol Concentration on the Growth Performance of A. platensis
3.2. Biochemical Composition
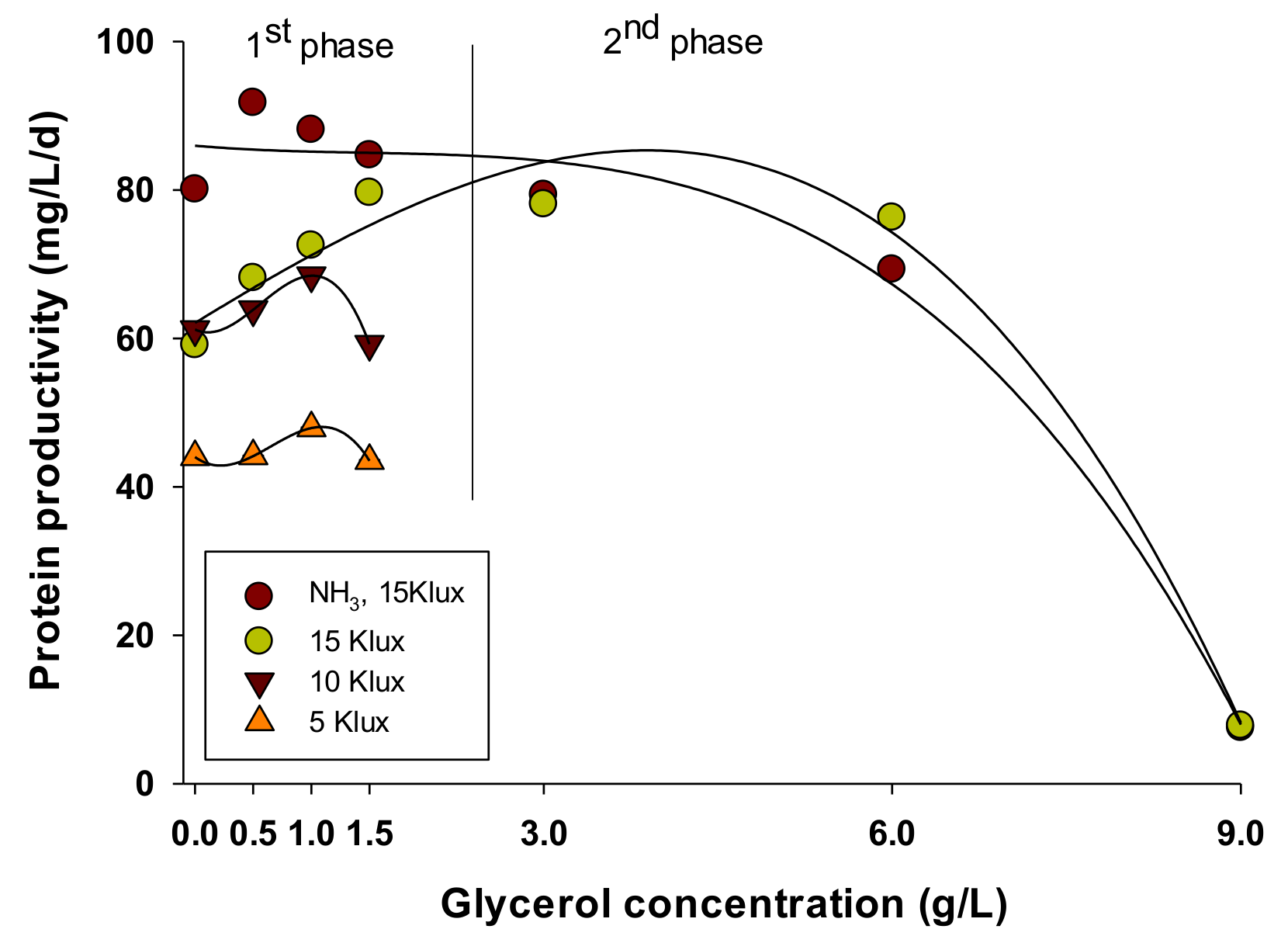
3.2.1. Proteins
3.2.2. Carbohydrates
3.2.3. Lipids
3.2.4. Pigments
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Borowitzka, M.A.; Vonshak, A. Scaling up microalgal cultures to commercial scale. Eur. J. Phycol. 2017, 52, 407–418. [Google Scholar] [CrossRef]
- Chojnacka, K.; Noworyta, A. Evaluation of Spirulina sp. growth in photoautotrophic, heterotrophic and mixotrophic cultures. Enzyme Microb. Technol. 2004, 34, 461–465. [Google Scholar] [CrossRef]
- Monteiro, M.R.; Kugelmeier, C.L.; Pinheiro, R.S.; Batalha, M.O.; da Silva César, A. Glycerol from biodiesel production: Technological paths for sustainability. Renew. Sustain. Energy Rev. 2018, 88, 109–122. [Google Scholar] [CrossRef]
- Chen, F. High cell density culture of microalgae in heterotrophic growth. Trends Biotechnol. 1996, 14, 421–426. [Google Scholar] [CrossRef]
- Marquez, F.J.; Sasaki, K.; Kakizono, T.; Nishio, N.; Nagai, S. Growth characteristics of Spirulina platensis in mixotrophic and heterotrophic conditions. J. Ferment. Bioeng. 1993, 76, 408–410. [Google Scholar] [CrossRef]
- Mühling, M.; Belay, A.; Whitton, B.A. Screening Arthrospira (Spirulina) strains for heterotrophy. J. Appl. Phycol. 2005, 17, 129–135. [Google Scholar] [CrossRef]
- Narayan, M.; Manoj, G.; Vatchravelu, K.; Bhagyalakshmi, N.; Mahadevaswamy, M. Utilization of glycerol as carbon source on the growth, pigment and lipid production in Spirulina platensis. Int. J. Food Sci. Nutr. 2005, 56, 521–528. [Google Scholar] [CrossRef]
- Morais, E.G.; Druzian, J.I.; Nunes, I.L.; Morais, M.G.; Costa, J.A.V. Glycerol increases growth, protein production and alters the fatty acids profile of Spirulina (Arthrospira) sp LEB 18. Process Biochem. 2019, 76, 40–45. [Google Scholar] [CrossRef]
- Shiraishi, H. Association of heterotrophic bacteria with aggregated Arthrospira platensis exopolysaccharides: Implications in the induction of axenic cultures. Biosci. Biotechnol. Biochem. 2015, 79, 331–341. [Google Scholar] [CrossRef]
- Deng, X.; Chen, B.; Xue, C.; Li, D.; Hu, X.; Gao, K. Biomass production and biochemical profiles of a freshwater microalga Chlorella kessleri in mixotrophic culture: Effects of light intensity and photoperiodicity. Bioresour. Technol. 2019, 273, 358–367. [Google Scholar] [CrossRef]
- Pastore, M.; Santaeufemia, S.; Bertucco, A.; Sforza, E. Light intensity affects the mixotrophic carbon exploitation in Chlorella protothecoides: Consequences on microalgae-bacteria based wastewater treatment. Water Sci. Technol. 2018, 78, 1762–1771. [Google Scholar] [CrossRef] [PubMed]
- Markou, G.; Vandamme, D.; Muylaert, K. Ammonia inhibition on Arthrospira platensis in relation to the initial biomass density and pH. Bioresour. Technol. 2014, 166, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, M.J.; Garcin, C.; van Hille, R.P.; Harrison, S.T.L. Interference by pigment in the estimation of microalgal biomass concentration by optical density. J. Microbiol. Methods 2011, 85, 119–123. [Google Scholar] [CrossRef] [PubMed]
- Kochert, G. Carbohydrate determination by phenol-sulfuric acid method. In Handbook of Phycological Methods. Physiological and Biochemical Methods; Hellebust, J.A., Craige, J.S., Eds.; Cambridge University Press: London, UK, 1978; pp. 95–97. [Google Scholar]
- Izard, J.; Limberger, R.J. Rapid screening method for quantitation of bacterial cell lipids from whole cells. J. Microbiol. Methods 2003, 55, 411–418. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [PubMed]
- Sarada, R.; Pillai, M.G.; Ravishankar, G.A. Phycocyanin from Spirulina sp: Influence of processing of biomass on phycocyanin yield, analysis of efficacy of extraction methods and stability studies on phycocyanin. Process Biochem. 1999, 34, 795–801. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]
- Fernandez, E.; Galvan, A. Inorganic nitrogen assimilation in Chlamydomonas. J. Exp. Bot. 2007, 58, 2279–2287. [Google Scholar] [CrossRef]
- Boussiba, S.; Gibson, J. Ammonia translocation in cyanobacteria. FEMS Microbiol. Lett. 1991, 88, 1–14. [Google Scholar] [CrossRef]
- Vílchez, C.; Vega, J.M. Nitrite uptake by Chlamydomonas reinhardtii cells immobilized in calcium alginate. Appl. Microbiol. Biotechnol. 1994, 41, 137–141. [Google Scholar] [CrossRef]
- Choi, G.-G.; Bae, M.-S.; Ahn, C.-Y.; Oh, H.-M. Induction of axenic culture of Arthrospira (Spirulina) platensis based on antibiotic sensitivity of contaminating bacteria. Biotechnol. Lett. 2008, 30, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Mogale, M. Identification and Quantification of Bacteria Associated with Cultivated Spirulina and Impact of Physiological Factors. Ph.D. Thesis, University of Cape Town, Cape Town, South Africa, 2016. [Google Scholar]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Wang, J.; Gong, H.; Wen, X.; Ren, H.; Lu, C. Effects of heat stress on PSII photochemistry in a cyanobacterium Spirulina platensis. Plant Sci. 2008, 175, 556–564. [Google Scholar] [CrossRef]
- Gong, H.; Tang, Y.; Wang, J.; Wen, X.; Zhang, L.; Lu, C. Characterization of photosystem II in salt-stressed cyanobacterial Spirulina platensis cells. Biochim. Biophys. Acta (BBA)-Bioenerg. 2008, 1777, 488–495. [Google Scholar] [CrossRef]
- Markou, G.; Depraetere, O.; Muylaert, K. Effect of ammonia on the photosynthetic activity of Arthrospira and Chlorella: A study on chlorophyll fluorescence and electron transport. Algal Res. 2016, 16, 449–457. [Google Scholar] [CrossRef]
- Perez-Garcia, O.; Escalante, F.M.E.; de-Bashan, L.E.; Bashan, Y. Heterotrophic cultures of microalgae: Metabolism and potential products. Water Res. 2011, 45, 11–36. [Google Scholar] [CrossRef]
- Matson, M.M.; Atsumi, S. Photomixotrophic chemical production in cyanobacteria. Curr. Opin. Biotechnol. 2018, 50, 65–71. [Google Scholar] [CrossRef]
- Neilson, A.; Lewin, R. The uptake and utilization of organic carbon by algae: An essay in comparative biochemistry. Phycologia 1974, 13, 227–264. [Google Scholar] [CrossRef]
- Swamy, M.A. Chapter 6—Marine algal sources for treating bacterial diseases. In Advances in Food and Nutrition Research; Kim, S.-K., Ed.; Academic Press: Cambridge, MA, USA, 2011; Volume 64, pp. 71–84. [Google Scholar]
- Markou, G.; Angelidaki, I.; Georgakakis, D. Microalgal carbohydrates: An overview of the factors influencing carbohydrates production, and of main bioconversion technologies for production of biofuels. Appl. Microbiol. Biotechnol. 2012, 96, 631–645. [Google Scholar] [CrossRef]
- Wang, Y.; Chiu, S.Y.; Ho, S.H.; Liu, Z.; Hasunuma, T.; Chang, T.T.; Chang, K.F.; Chang, J.S.; Ren, N.Q.; Kondo, A. Improving carbohydrate production of Chlorella sorokiniana NIES-2168 through semi-continuous process coupled with mixotrophic cultivation. Biotechnol. J. 2016, 11, 1072–1081. [Google Scholar] [CrossRef]
- Galasso, C.; Gentile, A.; Orefice, I.; Ianora, A.; Bruno, A.; Noonan, D.M.; Sansone, C.; Albini, A.; Brunet, C. Microalgal derivatives as potential nutraceutical and food supplements for human health: A focus on cancer prevention and interception. Nutrients 2019, 11, 1226. [Google Scholar] [CrossRef] [PubMed]
- Tedesco, M.A.; Duerr, E.O. Light, temperature and nitrogen starvation effects on the total lipid and fatty acid content and composition ofSpirulina platensis UTEX 1928. J. Appl. Phycol. 1989, 1, 201–209. [Google Scholar] [CrossRef]
- Humphrey, A. Chlorophyll as a color and functional ingredient. J. Food Sci. 2004, 69, C422–C425. [Google Scholar] [CrossRef]
- Hosikian, A.; Lim, S.; Halim, R.; Danquah, M.K. Chlorophyll extraction from microalgae: A review on the process engineering aspects. Int. J. Chem. Eng. 2010, 2010, 391632. [Google Scholar] [CrossRef]
- Romay, C.; Gonzalez, R.; Ledon, N.; Remirez, D.; Rimbau, V. C-phycocyanin: A biliprotein with antioxidant, anti-inflammatory and neuroprotective effects. Curr. Protein Pept. Sci. 2003, 4, 207–216. [Google Scholar] [CrossRef]
- Liu, X.; Duan, S.; Li, A.; Xu, N.; Cai, Z.; Hu, Z. Effects of organic carbon sources on growth, photosynthesis, and respiration of Phaeodactylum tricornutum. J. Appl. Phycol. 2009, 21, 239–246. [Google Scholar] [CrossRef]
- Wilken, S.; Schuurmans, J.M.; Matthijs, H.C. Do mixotrophs grow as photoheterotrophs? Photophysiological acclimation of the chrysophyte Ochromonas danica after feeding. New Phytol. 2014, 204, 882–889. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Glycerol Concentration g/L | 15 Klux | 10 Klux | 5 Klux | NH3-15 Klux |
|---|---|---|---|---|
| 0 | 0.52 | 0.49 | 0.50 | 0.52 |
| 0.5 | 0.54 | 0.51 | 0.56 | 0.53 |
| 1 | 0.54 | 0.50 | 0.56 | 0.53 |
| 1.5 | 0.53 | 0.51 | 0.54 | 0.53 |
| 3 | 0.49 | 0.50 | ||
| 6 | 0.48 | 0.49 | ||
| 9 | 0.49 | 0.49 |
| Glycerol Concentration g/L | 5 Klux | 10 Klux | 15 Klux | NH3-15 Klux |
|---|---|---|---|---|
| 0 | 39.84 ± 5.44 | 44.81 ± 3.85 | 41.96 ± 3.52 | 53.25 ± 1.95 |
| 0.5 | 48.01 ± 9.43 | 51.65 ± 6.26 | 49.50 ± 3.40 | 59.66 ± 2.25 |
| 1 | 58.14 ± 6.55 | 55.01 ± 6.77 | 53.20 ± 1.60 | 57.86 ± 3.66 |
| 1.5 | 54.80 ± 12.77 | 50.58 ± 2.96 | 50.96 ± 1.73 | 56.70 ± 4.30 |
| 3 | 60.70 ± 3.62 | 57.82 ± 4.85 | ||
| 6 | 64.30 ± 6.85 | 38.36 ± 3.88 | ||
| 9 | 15.93 ± 2.33 | 52.85 ± 5.31 |
| Glycerol Concentration g/L | 5 Klux | 10 Klux | 15 Klux | NH3-15 Klux |
|---|---|---|---|---|
| 0 | 40.53 ± 2.50 | 35.18 ± 1.52 | 35.78 ± 2.23 | 40.48 ± 1.82 |
| 0.5 | 32.24 ± 2.14 | 35.04 ± 1.89 | 33.17 ± 3.01 | 41.75 ± 2.04 |
| 1 | 32.22 ± 3.11 | 32.78 ± 2.77 | 29.82 ± 1.92 | 37.98 ± 0.84 |
| 1.5 | 32.15 ± 1.98 | 31.20 ± 1.55 | 25.48 ± 0.99 | 34.28 ± 3.55 |
| 3 | 25.79 ± 1.24 | 24.20 ± 2.61 | ||
| 6 | 28.65 ± 2.33 | 19.17 ± 2.24 | ||
| 9 | - | - |
| Glycerol Concentration g/L | 5 Klux | 10 Klux | 15 Klux | NH3-15 Klux |
|---|---|---|---|---|
| 0 | 7.22 ± 0.49 | 7.64 ± 1.00 | 5.89 ± 0.58 | 4.96 ± 1.27 |
| 0.5 | 9.23 ± 0.27 | 7.32 ± 0.20 | 6.40 ± 0.43 | 5.84 ± 0.41 |
| 1 | 8.76 ± 0.33 | 8.32 ± 0.43 | 6.36 ± 0.26 | 5.77 ± 0.23 |
| 1.5 | 9.22 ± 0.38 | 8.44 ± 0.85 | 5.70 ± 0.43 | 5.40 ± 0.48 |
| 3 | 9.46 ± 1.05 | 9.16 ± 0.97 | ||
| 6 | 11.13 ± 1.45 | 10.03 ± 0.20 | ||
| 9 | - | - |
| Glycerol Concentration g/L | 5 Klux | 10 Klux | 15 Klux | NH3-15 Klux |
|---|---|---|---|---|
| 0 | 12.99 ± 0.55 | 14.71 ± 0.25 | 14.20 ± 1.38 | 15.73 ± 1.45 |
| 0.5 | 11.22 ± 0.37 | 12.86 ± 0.1 | 12.84 ± 2.95 | 14.21 ± 2.58 |
| 1 | 12.10 ± 0.75 | 10.64 ± 0.14 | 14.83± 0.25 | 14.07 ± 1.27 |
| 1.5 | 11.42 ± 1.11 | 10.55 ± 0.46 | 12.85 ± 0.05 | 12.60 ± 0.79 |
| 3 | 6.60 ± 0.25 | 6.85 ± 0.5 | ||
| 6 | 8.19 ± 2.12 | 6.13 ± 0.34 | ||
| 9 | 1.36 ± 1.58 | 2.85 ± 0.55 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Markou, G.; Kougia, E.; Kefalogianni, I.; Tsagou, V.; Arapoglou, D.; Chatzipavlidis, I. Effect of Glycerol Concentration and Light Intensity on Growth and Biochemical Composition of Arthrospira (Spirulina) Platensis: A Study in Semi-Continuous Mode with Non-Aseptic Conditions. Appl. Sci. 2019, 9, 4703. https://doi.org/10.3390/app9214703
Markou G, Kougia E, Kefalogianni I, Tsagou V, Arapoglou D, Chatzipavlidis I. Effect of Glycerol Concentration and Light Intensity on Growth and Biochemical Composition of Arthrospira (Spirulina) Platensis: A Study in Semi-Continuous Mode with Non-Aseptic Conditions. Applied Sciences. 2019; 9(21):4703. https://doi.org/10.3390/app9214703
Chicago/Turabian StyleMarkou, Giorgos, Eleni Kougia, Io Kefalogianni, Vasiliki Tsagou, Dimitrios Arapoglou, and Iordanis Chatzipavlidis. 2019. "Effect of Glycerol Concentration and Light Intensity on Growth and Biochemical Composition of Arthrospira (Spirulina) Platensis: A Study in Semi-Continuous Mode with Non-Aseptic Conditions" Applied Sciences 9, no. 21: 4703. https://doi.org/10.3390/app9214703
APA StyleMarkou, G., Kougia, E., Kefalogianni, I., Tsagou, V., Arapoglou, D., & Chatzipavlidis, I. (2019). Effect of Glycerol Concentration and Light Intensity on Growth and Biochemical Composition of Arthrospira (Spirulina) Platensis: A Study in Semi-Continuous Mode with Non-Aseptic Conditions. Applied Sciences, 9(21), 4703. https://doi.org/10.3390/app9214703