Propofol Exposure in Pregnant Rats Induces Neurotoxicity and Persistent Learning Deficit in the Offspring

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Propofol Administration
2.3. Western Blot Analysis for Cleaved Casepase-3 in the Fetal Brain
2.4. Neonatal Brain Histology Studies
2.5. Nissl Staining
2.6. Immunohistochemistry Analysis for NeuN
2.7. Immunofluorescence Staining of Synaptophysin in the Hippocampus
2.8. Animal Behavioral Studies
2.9. Open-Field Test
2.10. Eight-Arm Radial Maze
2.11. Statistical Analysis
3. Results
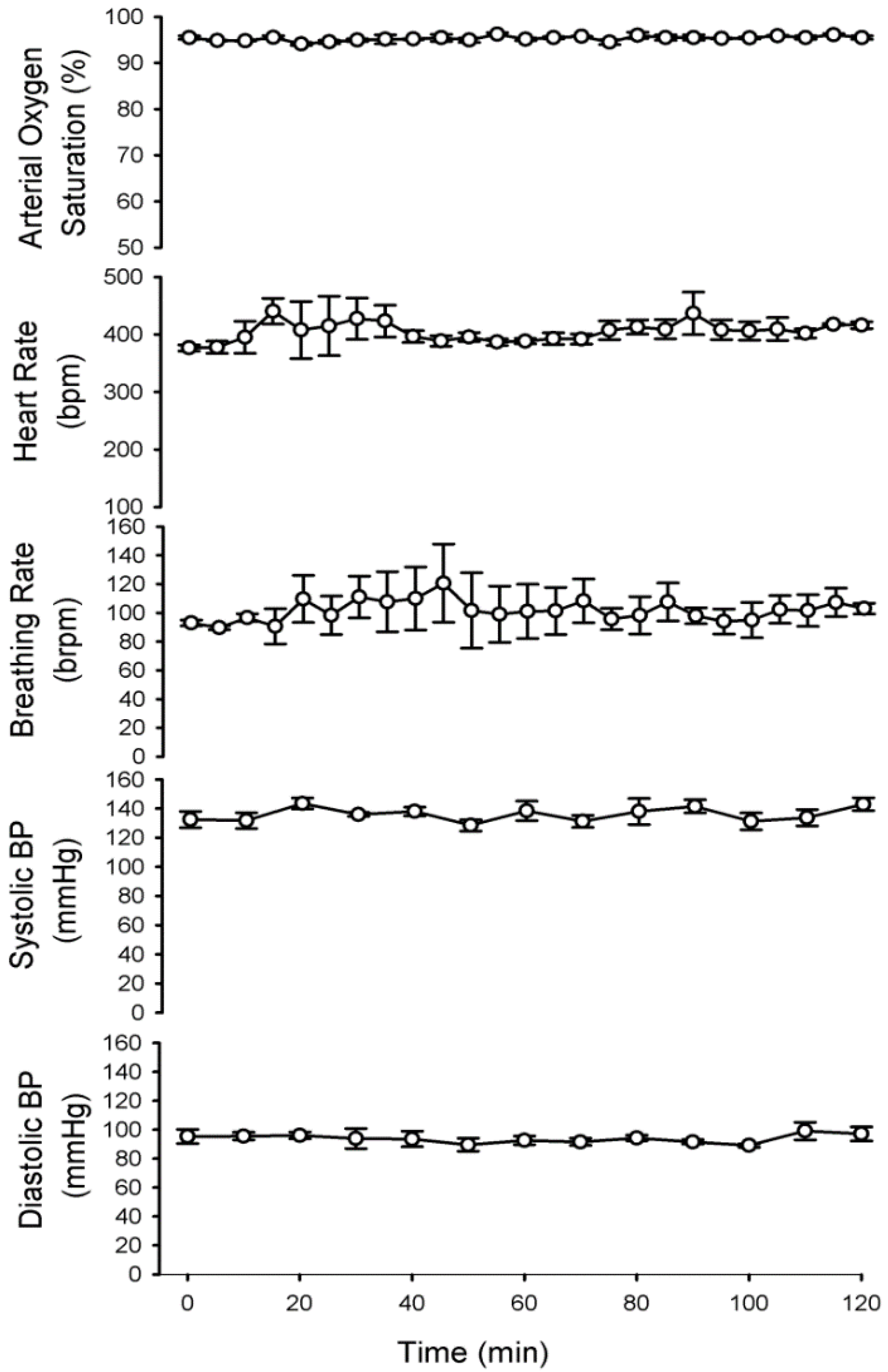
3.1. Physiologic Parameters of Dams during Propofol Infusion


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| VBG | Propofol infusion | ||
|---|---|---|---|
| 0 h (awake) | 1 h | 2 h | |
| pH (venous) | 7.40 ± 0.00 | 7.38 ± 0.03 | 7.33 ± 0.00 |
| PVCO2 (mmHg) | 40.1 ± 4.5 | 42.5 ± 2.5 | 41.6 ± 2.4 |
| PVO2 (mmHg) | 50.3 ± 7.3 | 50.3 ± 2.8 | 73.0 ± 14.1 |
| HCO3− (mmol/L) | 22.7 ± 3.2 | 26.7 ± 1.5 | 24.6 ± 2.4 |
| Na+ (mmol/L) | 146.0 ± 4.0 | 136.6 ± 2.0 | 140.6 ± 2.3 |
| K+ (mmol/L) | 3.7 ± 0.2 | 3.6 ± 0.2 | 3.7 ± 0.3 |
| Glucose (mg/dL) | 89 ± 1.2 | 83.4 ± 1.2 | 87.0 ± 5.5 |
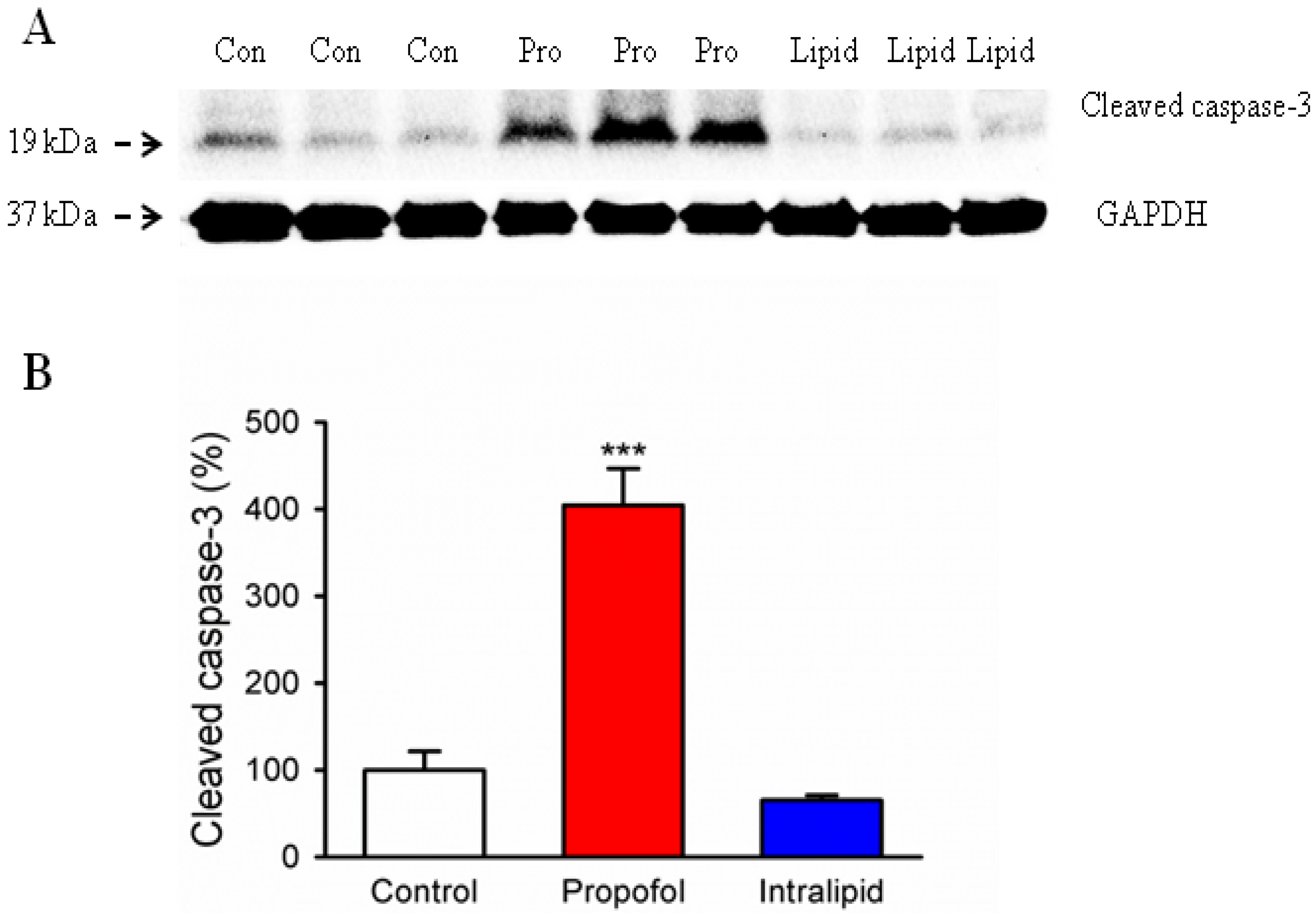
3.2. Cleaved Caspase-3 Levels in the Brain of Fetal Rats

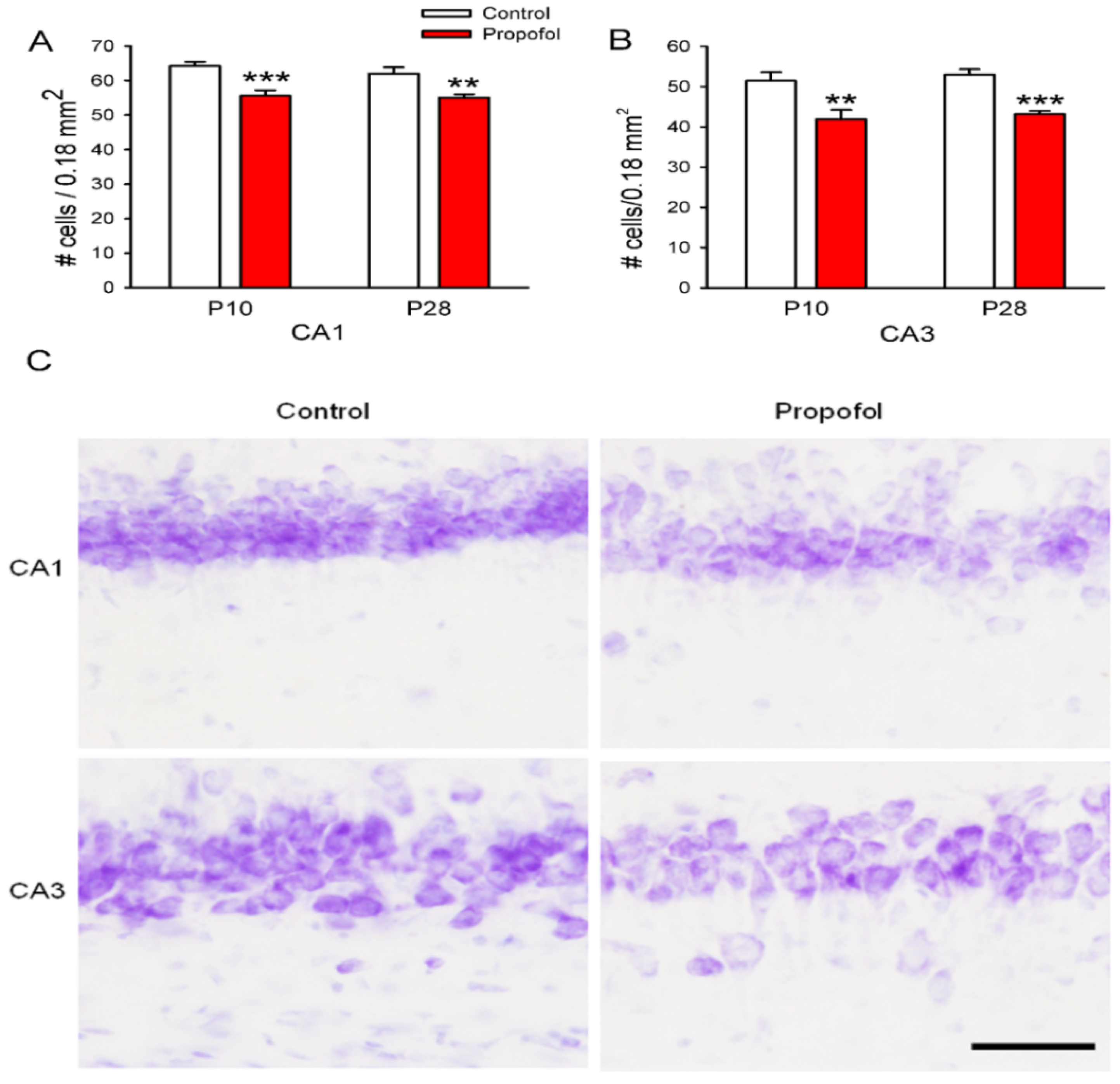
3.3. Neuron Deletion in the Hippocampus of Offspring Rats

3.4. NeuN Staining in the Hippocampus

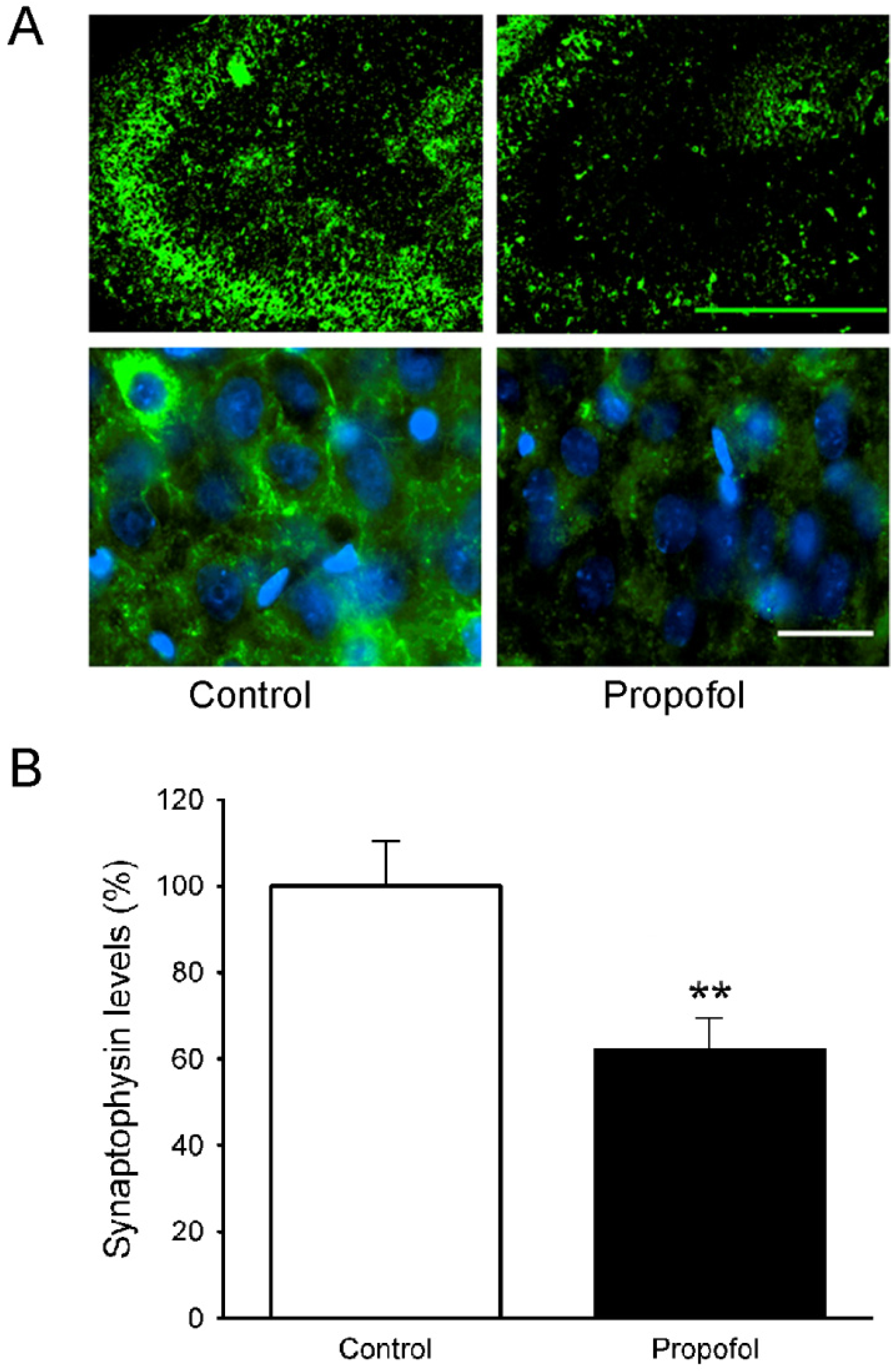
3.5. Synaptophysin Levels in the Hippocampus of Offspring Rats

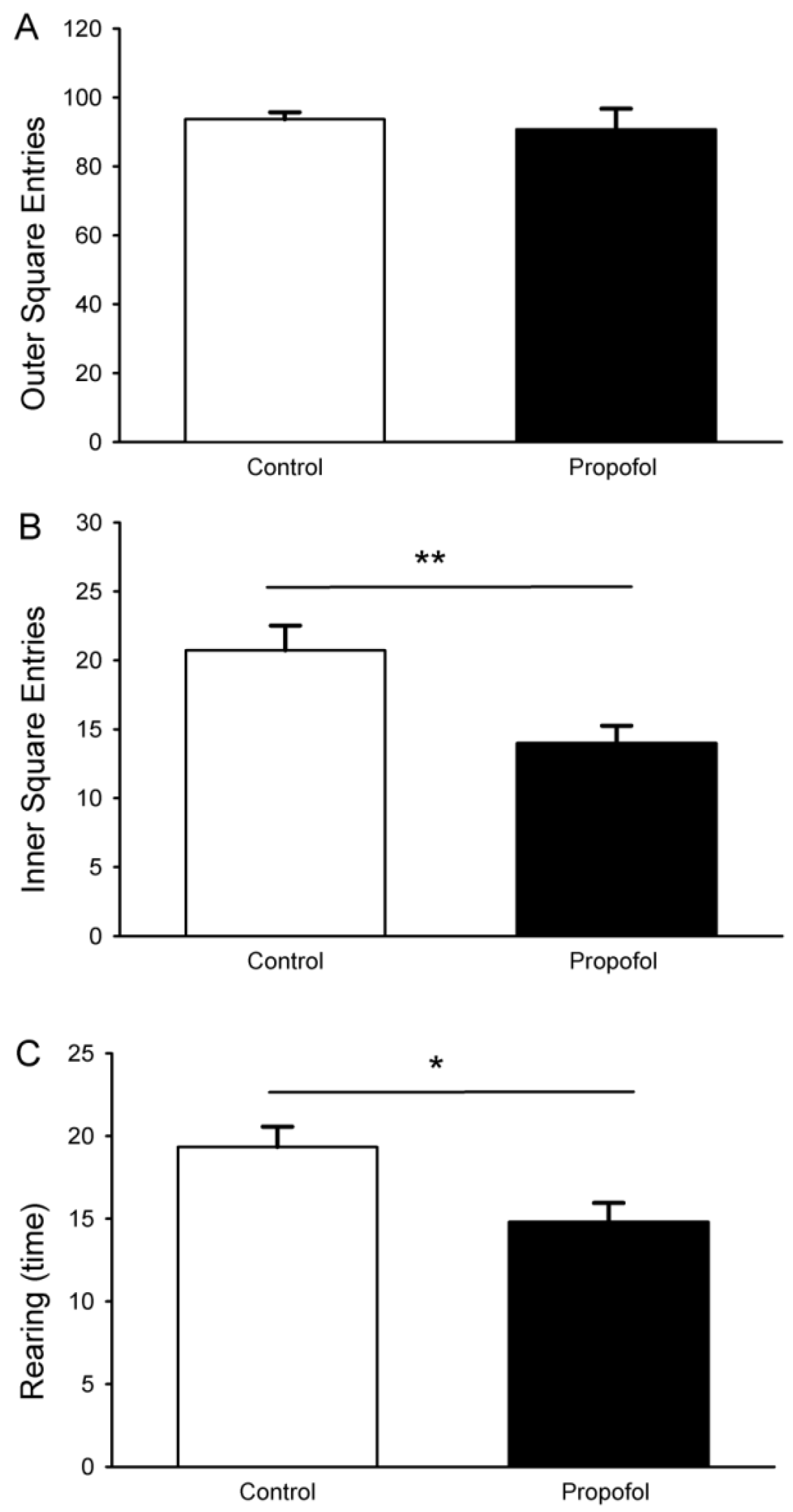
3.6. Open-Field Test

| Open field test | Control (n = 18) | Propofol (n = 16) |
|---|---|---|
| Number of feces | 4.0 ± 0.4 | 3.5 ± 0.4 |
| Time spent in the inner squares (seconds) | 29.0 ± 3.0 | 26.4 ± 3.3 |
| Time spent in the outer squares (seconds) | 271 ± 3.0 | 273.6 ± 3.3 |
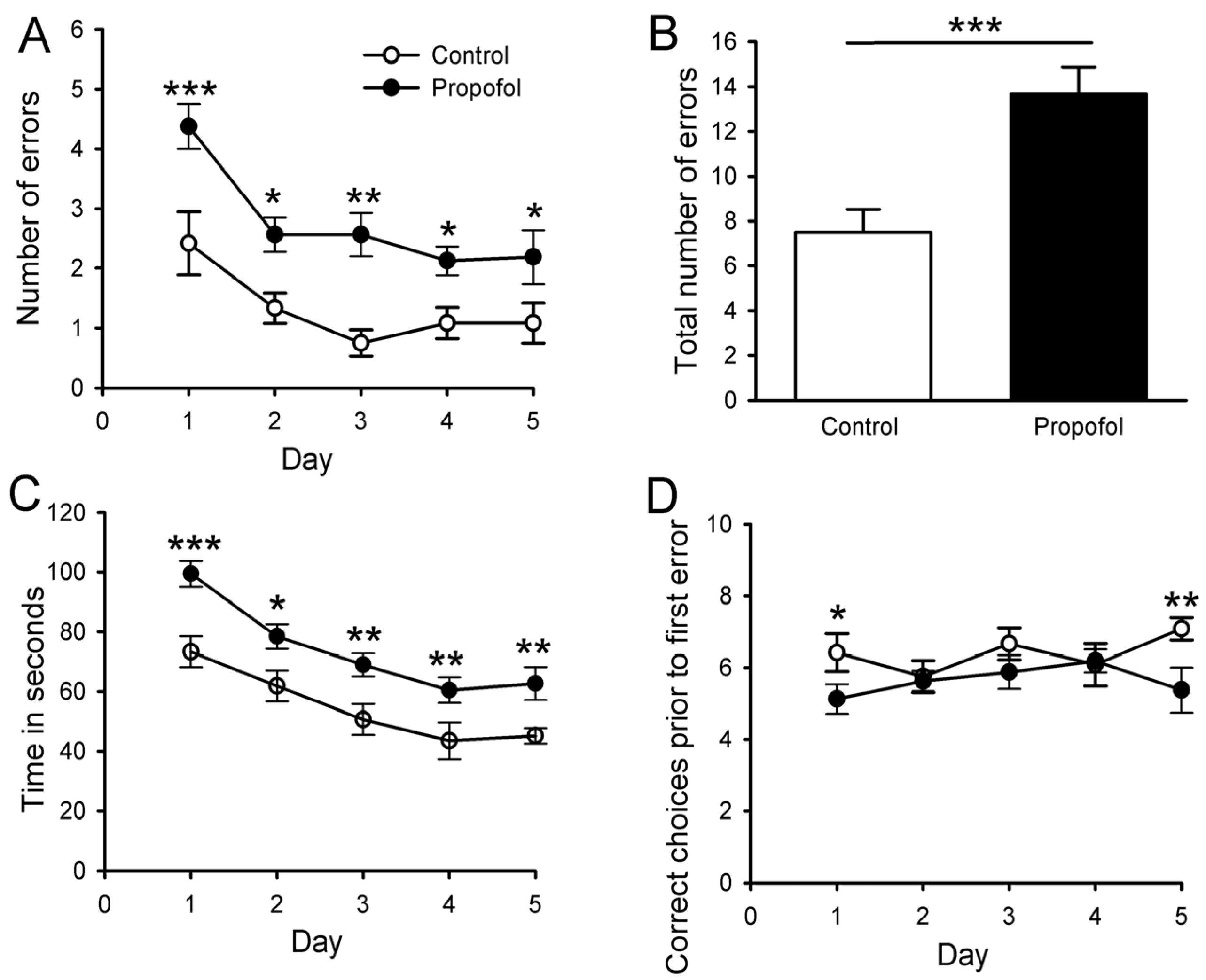
3.7. Eight-Arm Radial Maze

4. Discussion
5. Conclusions
Author Contributions
Conflicts of Interest
References
- Kotani, Y.; Shimazawa, M.; Yoshimura, S.; Iwama, T.; Hara, H. The experimental and clinical pharmacology of propofol, an anesthetic agent with neuroprotective properties. CNS Neurosci. Ther. 2008, 14, 95–106. [Google Scholar] [CrossRef]
- Olsen, E.A.; Brambrink, A.M. Anesthetic neurotoxicity in the newborn and infant. Curr. Opin. Anaesthesiol. 2013. [Google Scholar] [CrossRef]
- Creeley, C.E.; Dikranian, K.T.; Dissen, G.A.; Back, S.A.; Olney, J.W.; Brambrink, A.M. Isoflurane-induced Apoptosis of Neurons and Oligodendrocytes in the Fetal Rhesus Macaque Brain. Anesthesiology 2013. [Google Scholar] [CrossRef]
- Jevtovic-Todorovic, V. Pediatric anesthesia neurotoxicity: An overview of the 2011 SmartTots panel. Anesth. Analg. 2011, 113, 965–968. [Google Scholar] [CrossRef]
- Van de Velde, M.; de Buck, F. Anesthesia for non-obstetric surgery in the pregnant patient. Minerva Anestesiol. 2007, 73, 235–240. [Google Scholar]
- Palanisamy, A.; Baxter, M.G.; Keel, P.K.; Xie, Z.; Crosby, G.; Culley, D.J. Rats exposed to isoflurane in utero during early gestation are behaviorally abnormal as adults. Anesthesiology 2011, 114, 521–528. [Google Scholar] [CrossRef]
- Kong, F.J.; Xu, L.; He, D.; Zhang, X.; Lu, H. Effects of gestational isoflurane exposure on postnatal memory and learning in rats. Eur. J. Pharmacol. 2011, 670, 168–174. [Google Scholar] [CrossRef]
- Dupret, D.; O’Neill, J.; Csicsvari, J. Dynamic reconfiguration of hippocampal interneuron circuits during spatial learning. Neuron 2013, 78, 166–180. [Google Scholar] [CrossRef]
- Dupret, D.; Fabre, A.; Döbrössy, M.D.; Panatier, A.; Rodríguez, J.J.; Lamarque, S.; Lemaire, V.; Oliet, S.H.; Piazza, P.V.; Abrous, D.N. Spatial learning depends on both the addition and removal of new hippocampal neurons. PLoS Biol. 2007, 5, e214. [Google Scholar] [CrossRef]
- Edelmann, L.; Hanson, P.I.; Chapman, E.R.; Jahn, R. Synaptobrevin binding to synaptophysin: A potential mechanism for controlling the exocytotic fusion machine. EMBO J. 1995, 14, 224–231. [Google Scholar]
- Grosse, G.; Tapp, R.; Wartenberg, M.; Sauer, H.; Fox, P.A.; Grosse, J.; Gratzl, M.; Bergmann, M. Prenatal hippocampal granule cells in primary cell culture form mossy fiber boutons at pyramidal cell dendrites. J. Neurosci. Res. 1998, 51, 602–611. [Google Scholar] [CrossRef]
- Voigt, T.; de Lima, A.D.; Beckmann, M. Synaptophysin immunohistochemistry reveals inside-out pattern of early synaptogenesis in ferret cerebral cortex. J. Comp. Neurol. 1993, 330, 48–64. [Google Scholar] [CrossRef]
- Lavezzi, A.M.; Corna, M.F.; Matturri, L. Neuronal nuclear antigen (NeuN): A useful marker of neuronal immaturity in sudden unexplained perinatal death. J. Neurol. Sci. 2013, 329, 45–50. [Google Scholar] [CrossRef]
- Mullen, R.J.; Buck, C.R.; Smith, A.M. NeuN, a neuronal specific nuclear protein in vertebrates. Development 1992, 116, 201–211. [Google Scholar]
- Yang, G.; Chang, P.C.; Bekker, A.; Blanck, T.J.; Gan, W.B. Transient effects of anesthetics on dendritic spines and filopodia in the living mouse cortex. Anesthesiology 2011, 115, 718–726. [Google Scholar] [CrossRef]
- Li, Y.; Liang, G.; Wang, S.; Meng, Q.; Wang, Q.; Weia, H. Effects of fetal exposure to isoflurane on postnatal memory and learning in rats. Neuropharmacology 2007, 53, 942–950. [Google Scholar] [CrossRef]
- Cattano, D.; Young, C.; Straiko, M.M.; Olney, J.W. Subanesthetic doses of propofol induce neuroapoptosis in the infant mouse brain. Anesth. Analg. 2008, 106, 1712–1714. [Google Scholar] [CrossRef]
- Yu, D.; Jiang, Y.; Gao, J.; Liu, B.; Chen, P. Repeated exposure to propofol potentiates neuroapoptosis and long-term behavioral deficits in neonatal rats. Neurosci. Lett. 2013, 534, 41–46. [Google Scholar] [CrossRef]
- Pearn, M.L.; Hu, Y.; Niesman, I.R.; Patel, H.H.; Drummond, J.C.; Roth, D.M.; Akassoglou, K.; Patel, P.M.; Head, B.P. Propofol neurotoxicity is mediated by p75 neurotrophin receptor activation. Anesthesiology 2012, 116, 352–361. [Google Scholar] [CrossRef]
- Creeley, C.; Dikranian, K.; Dissen, G.; Martin, L.; Olney, L.; Brambrink, A. Propofol-induced apoptosis of neurones and oligodendrocytes in fetal and neonatal rhesus macaque brain. Br. J. Anaesth. 2013, 110 (Suppl. 1), i29–i38. [Google Scholar] [CrossRef]
- Vutskits, L.; Gascon, E.; Tassonyi, E.; Kiss, J.Z. Clinically relevant concentrations of propofol but not midazolam alter in vitro dendritic development of isolated γ-aminobutyric acid-positive interneurons. Anesthesiology 2005, 102, 970–976. [Google Scholar] [CrossRef]
- Nunez, J.L.; Alt, J.J.; McCarthy, M.M. A novel model for prenatal brain damage. II. Long-term deficits in hippocampal cell number and hippocampal-dependent behavior following neonatal GABAA receptor activation. Exp. Neurol. 2003, 181, 270–280. [Google Scholar] [CrossRef]
- Olton, D.S.; Walker, J.A.; Gage, F.H. Hippocampal connections and spatial discrimination. Brain Res. 1978, 139, 295–308. [Google Scholar] [CrossRef]
- Xia, L.; Jiang, Z.L.; Wang, G.H.; Hu, B.Y.; Ke, K.F. Treatment with ginseng total saponins reduces the secondary brain injury in rat after cortical impact. J. Neurosci. Res. 2012, 90, 1424–1436. [Google Scholar] [CrossRef]
- Li, J.; Sun, Y.; Ye, J.H. Electroacupuncture decreases excessive alcohol consumption involving reduction of FosB/DeltaFosB levels in reward-related brain regions. PLoS One 2012, 7, e40347. [Google Scholar]
- Li, J.; Cheng, Y.; Bian, W.; Liu, X.; Zhang, C.; Ye, J.H. Region-specific induction of FosB/∆FosB by voluntary alcohol intake: Effects of naltrexone. Alcohol. Clin. Exp. Res. 2010, 34, 1742–1750. [Google Scholar] [CrossRef]
- Zheng, H.; Dong, Y.; Xu, Z.; Crosby, G.; Culley, D.J.; Zhang, Y.; Xie, Z. Sevoflurane anesthesia in pregnant mice induces neurotoxicity in fetal and offspring mice. Anesthesiology 2013, 118, 516–526. [Google Scholar] [CrossRef]
- Liu, P.; Bilkey, D.K. The effect of excitotoxic lesions centered on the perirhinal cortex in two versions of the radial arm maze task. Behav. Neurosci. 1999, 113, 672–682. [Google Scholar] [CrossRef]
- Soylemezoglu, F.; Onder, S.; Tezel, G.G.; Berker, M. Neuronal nuclear antigen (NeuN): A new tool in the diagnosis of central neurocytoma. Pathol. Res. Pract. 2003, 199, 463–468. [Google Scholar] [CrossRef]
- Popic, J.; Pesic, V.; Milanovic, D.; Todorovic, S.; Kanazir, S.; Jevtovic-Todorovic, V.; Ruzdijic, S. Propofol-induced changes in neurotrophic signaling in the developing nervous system in vivo. PLoS One 2012, 7, e34396. [Google Scholar] [CrossRef]
- Kahraman, S.; Zup, S.L.; McCarthy, M.M.; Fiskum, G. GABAergic mechanism of propofol toxicity in immature neurons. J. Neurosurg. Anesthesiol. 2008, 20, 233–240. [Google Scholar] [CrossRef]
- Sanchez-Alcaraz, A.; Quintana, M.B.; Laguarda, M. Placental transfer and neonatal effects of propofol in caesarean section. J. Clin. Pharm. Ther. 1998, 23, 19–23. [Google Scholar] [CrossRef]
- Briner, A.; Nikonenko, I.; de Roo, M.; Dayer, A.; Muller, D.; Vutskits, L. Developmental Stage-dependent persistent impact of propofol anesthesia on dendritic spines in the rat medial prefrontal cortex. Anesthesiology 2011, 115, 282–293. [Google Scholar] [CrossRef]
- Whishaw, I.Q.; Rod, M.R.; Auer, R.N. Behavioral deficits revealed by multiple tests in rats with ischemic damage limited to half of the CA1 sector of the hippocampus. Brain Res. Bull. 1994, 34, 283–289. [Google Scholar] [CrossRef]
- Brambrink, A.M.; Evers, A.S.; Avidan, M.S.; Farber, N.B.; Smith, D.J.; Martin, L.D.; Dissen, G.A.; Creeley, C.E.; Olney, J.W. Ketamine-induced neuroapoptosis in the fetal and neonatal rhesus macaque brain. Anesthesiology 2012, 116, 372–384. [Google Scholar] [CrossRef]
- Stratmann, G. Review article: Neurotoxicity of anesthetic drugs in the developing brain. Anesth. Analg. 2011, 113, 1170–1179. [Google Scholar] [CrossRef]
- Jevtovic-Todorovic, V. Anesthesia and the developing brain: Are we getting closer to understanding the truth? Curr. Opin. Anaesthesiol. 2011, 24, 395–399. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Xiong, M.; Li, J.; Alhashem, H.M.; Tilak, V.; Patel, A.; Pisklakov, S.; Siegel, A.; Ye, J.H.; Bekker, A. Propofol Exposure in Pregnant Rats Induces Neurotoxicity and Persistent Learning Deficit in the Offspring. Brain Sci. 2014, 4, 356-375. https://doi.org/10.3390/brainsci4020356
Xiong M, Li J, Alhashem HM, Tilak V, Patel A, Pisklakov S, Siegel A, Ye JH, Bekker A. Propofol Exposure in Pregnant Rats Induces Neurotoxicity and Persistent Learning Deficit in the Offspring. Brain Sciences. 2014; 4(2):356-375. https://doi.org/10.3390/brainsci4020356
Chicago/Turabian StyleXiong, Ming, Jing Li, Hussain M. Alhashem, Vasanti Tilak, Anuradha Patel, Sergey Pisklakov, Allan Siegel, Jiang Hong Ye, and Alex Bekker. 2014. "Propofol Exposure in Pregnant Rats Induces Neurotoxicity and Persistent Learning Deficit in the Offspring" Brain Sciences 4, no. 2: 356-375. https://doi.org/10.3390/brainsci4020356