
Electrometric and Electron Paramagnetic Resonance Measurements of a Difference in the Transmembrane Electrochemical Potential: Photosynthetic Subcellular Structures and Isolated Pigment–Protein Complexes

Abstract
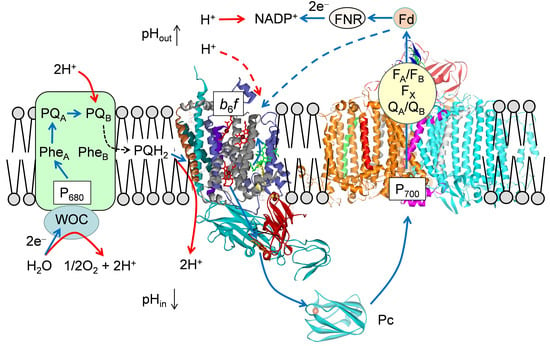
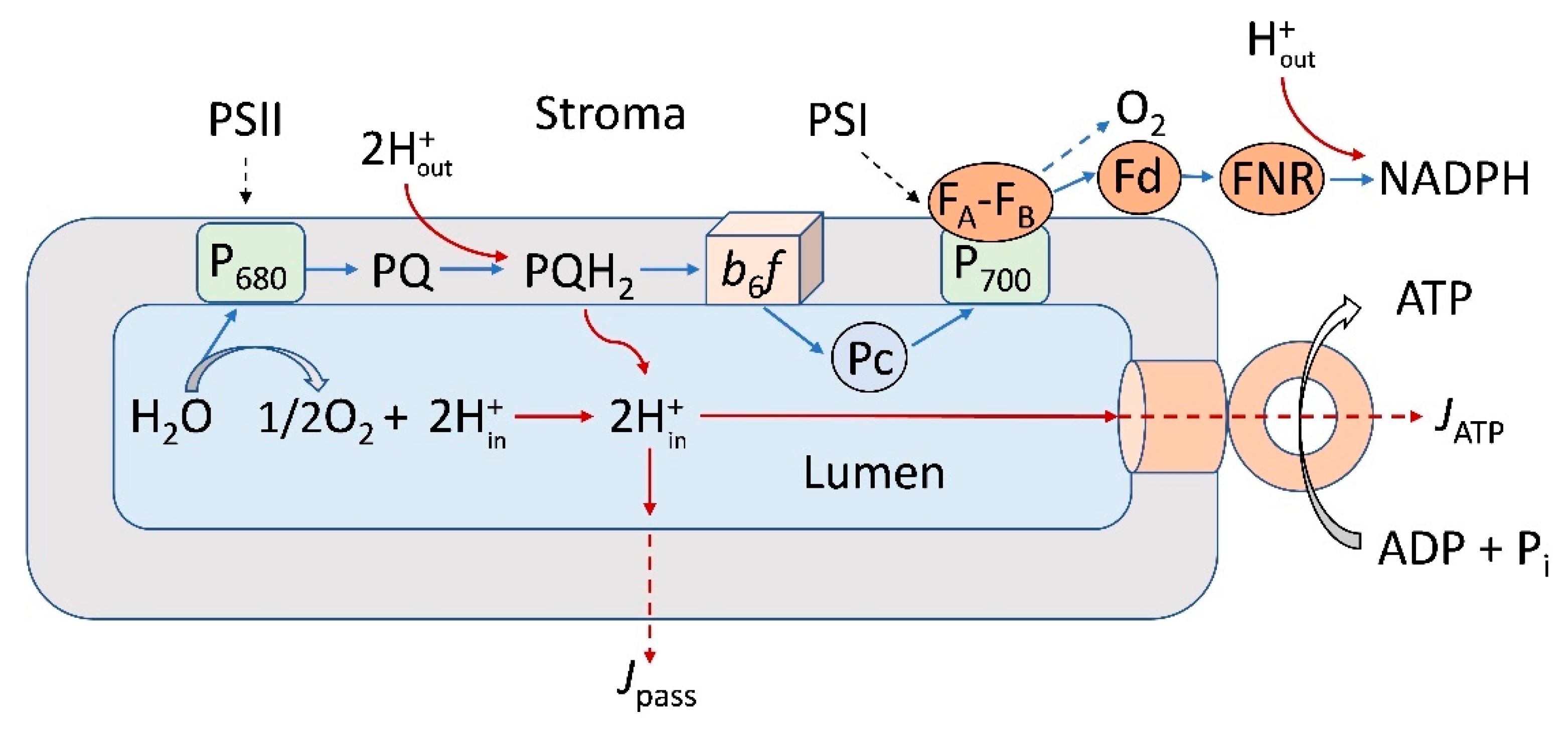
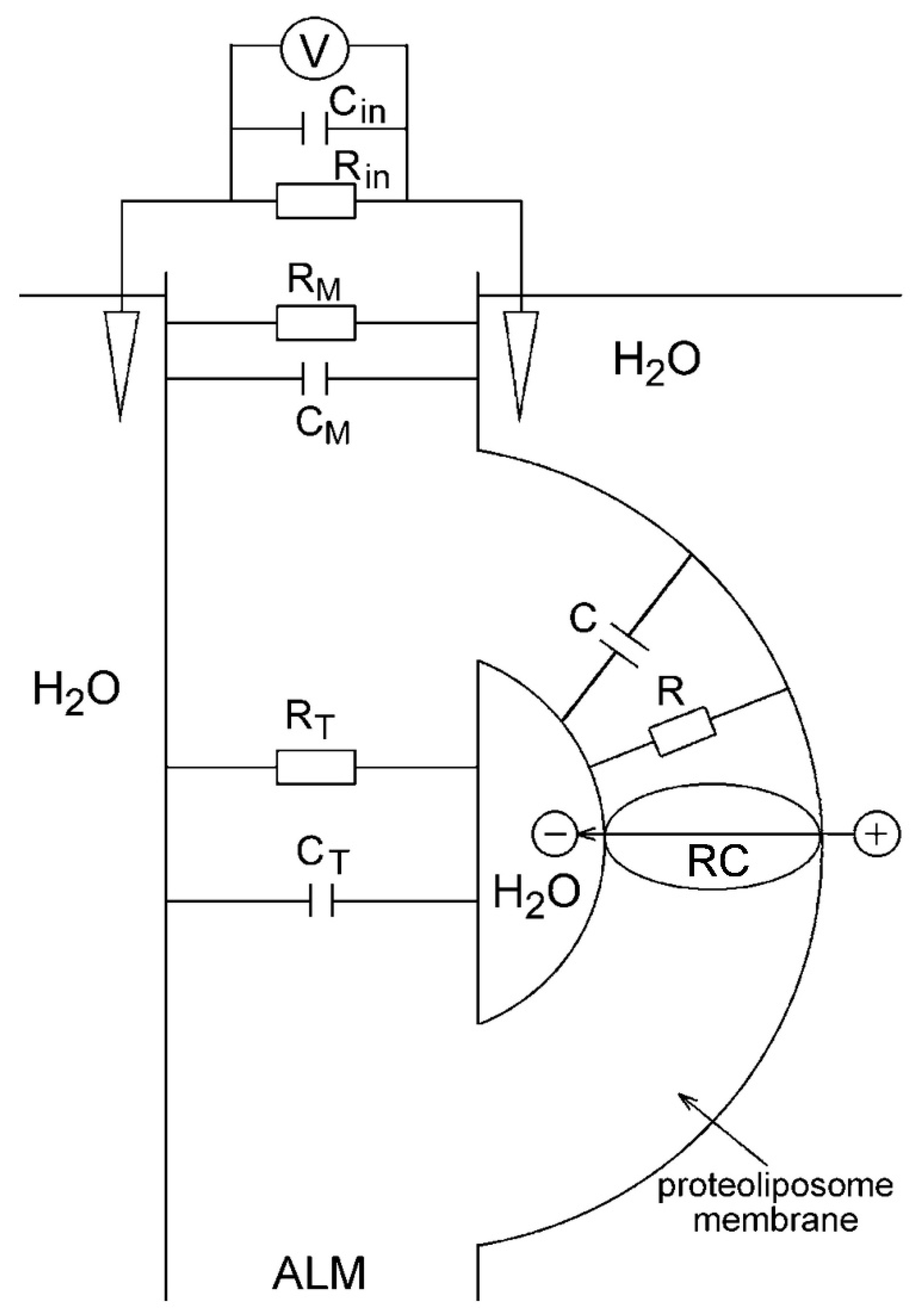
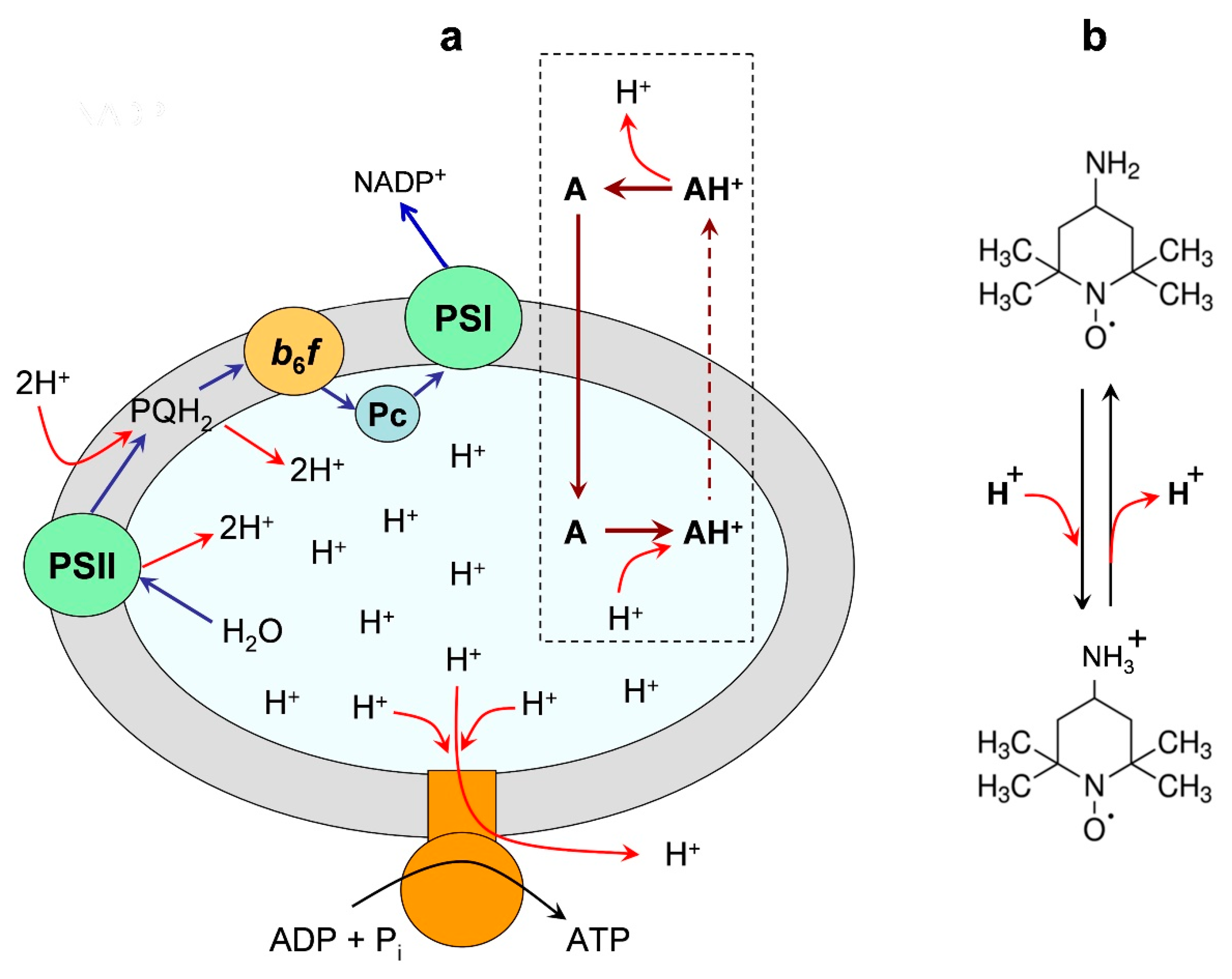
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. The Proton Motive Force in Membrane Bioenergetics
3. Overview of the Methods for Measuring Δψ and ΔpH in Biomembranes
3.1. Synthetic Penetrating Ions
3.2. Electrochromic Band Shift of Carotenoid Spectrum
3.3. The Direct Electrometrical Method of Δψ Measurements
3.4. The Use of pH-Sensitive Molecular Probes
3.4.1. Partitioning the Indicator Molecules between the Lumen and External Volume
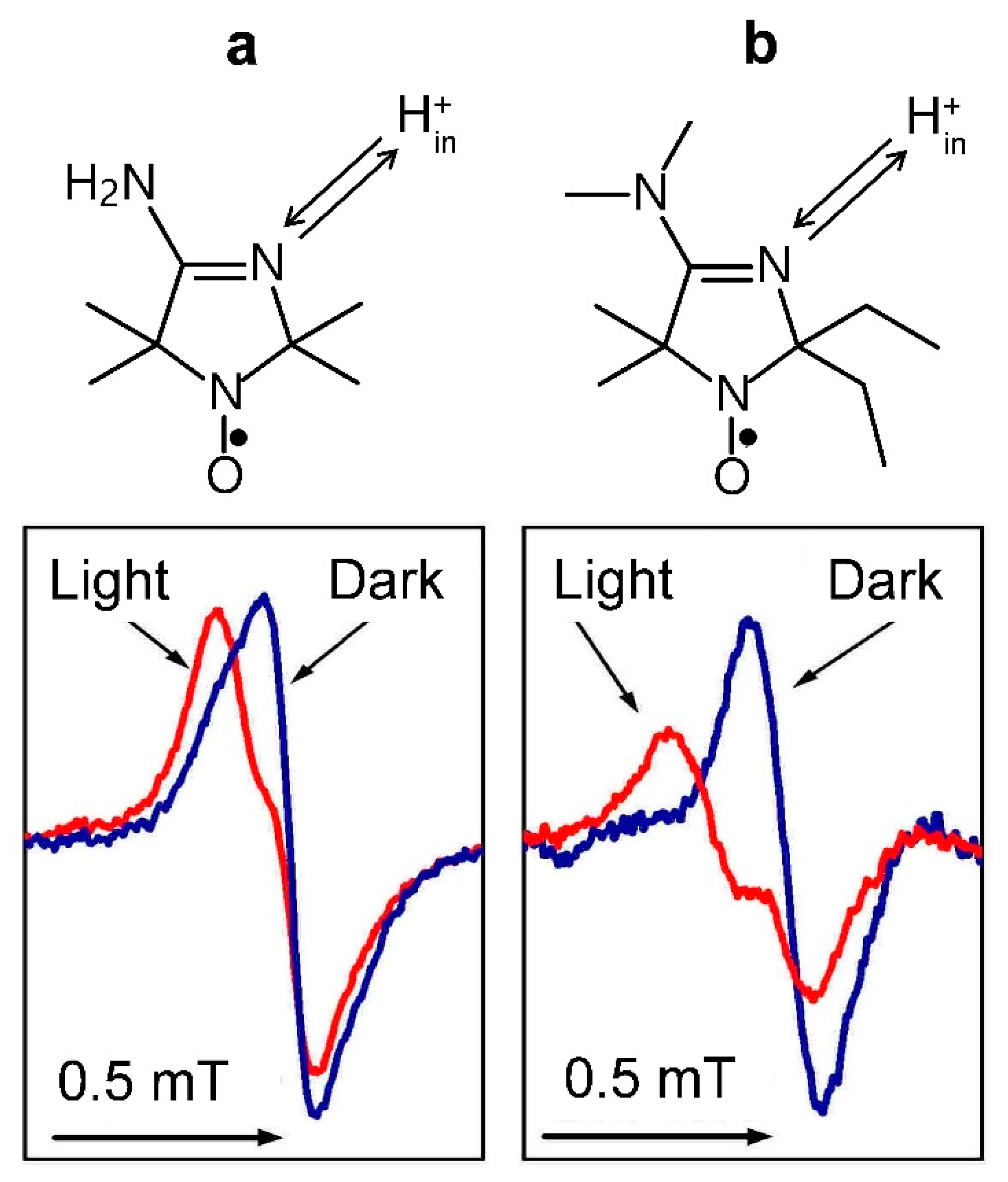
3.4.2. The Light-Induced Changes in the EPR Spectra of pH-Sensitive Spin Probes
3.4.3. Surface Potential Measurements
3.5. pH-Dependent Kinetics of Photosynthetic Electron Transport in Chloroplasts
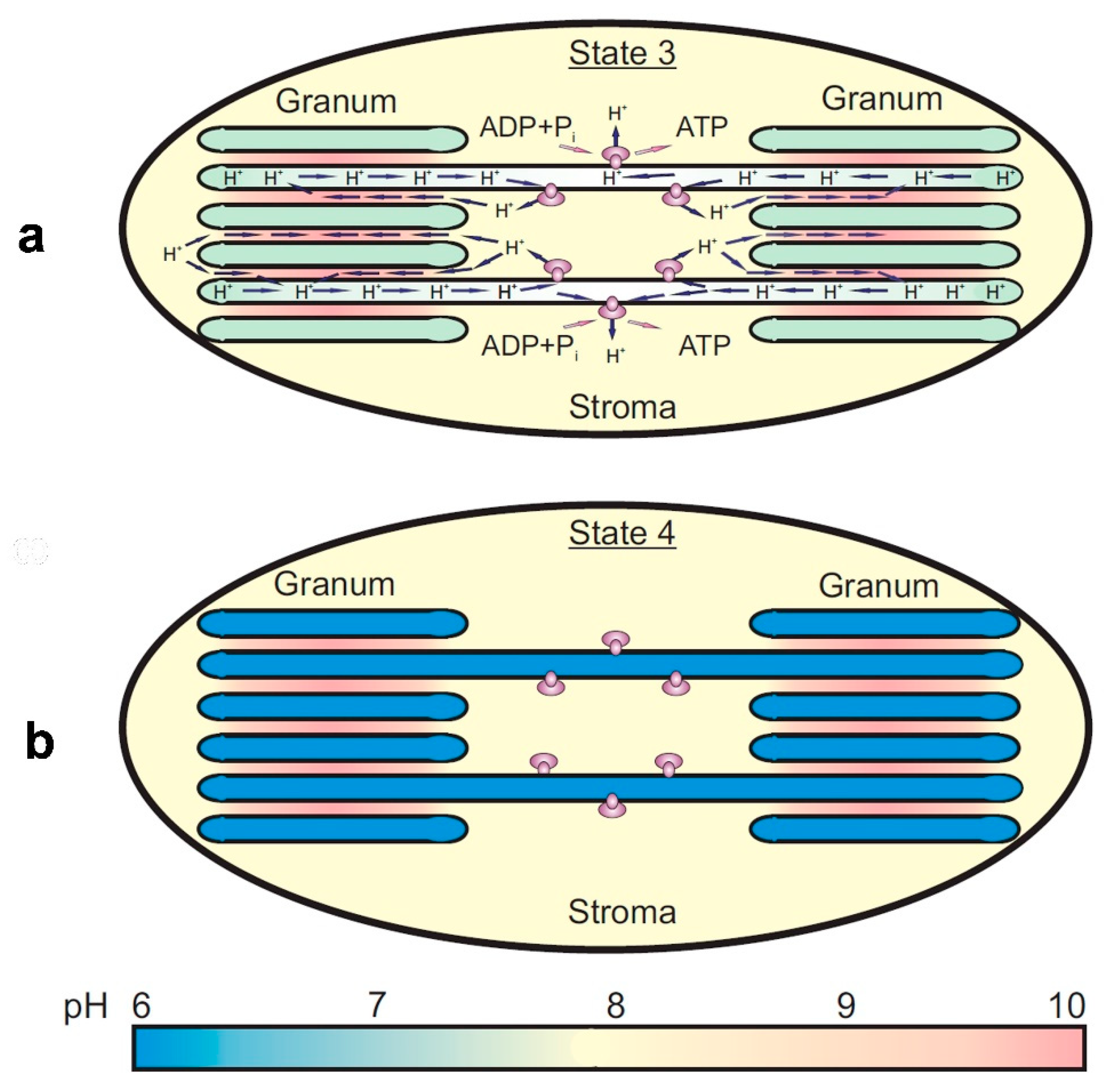
3.6. Lateral Heterogeniety of ΔpH
4. Basic Results of pmf Measurements
4.1. Direct Electrometrical Measurements
4.2. EPR Measurements of ΔpH in Chloroplasts
5. Future Directions
5.1. Fundamental Studies
5.2. Practical Implications
6. Conclusions
Author Contributions
Funding
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Mitchell, P. Coupling of phosphorylation to electron and hydrogen transfer by a chemi-osmotic type of mechanism. Nature 1961, 191, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, P. Chemiosmotic coupling in oxidative and photosynthetic phosphorylation. Biol. Rev. 1966, 41, 445–502. [Google Scholar] [CrossRef] [PubMed]
- Gräber, P. Phosphorylation in chloroplasts: ATP synthesis driven by Δψ and by ΔpH of artificial or light-generated origin. Curr. Top. Membr. Transp. 1982, 16, 215–245. [Google Scholar]
- Hangarter, R.P.; Good, N.E. Energy thresholds for ATP synthesis in chloroplasts. Biochim. Biophys. Acta 1982, 681, 397–404. [Google Scholar] [CrossRef]
- Boyer, P.D. The ATP synthase—A splendid molecular machine. Annu. Rev. Biochem. 1997, 66, 717–749. [Google Scholar] [CrossRef]
- Junge, W.; Nelson, N. ATP synthase. Annu. Rev. Biochem. 2015, 83, 631–657. [Google Scholar] [CrossRef]
- Lai, Y.; Zhang, Y.; Zhou, S.; Xu, J.; Du, Z.; Feng, Z.; Yu, L.; Zhao, Z.; Wang, W.; Tang, Y.; et al. Structure of the human ATP synthase. Mol. Cell 2023, 83, 2137–2147.e4. [Google Scholar] [CrossRef]
- Edwards, G.E.; Walker, D.A. C3, C4: Mechanisms, and Cellular and Environmental Regulation of Photosynthesis; Blackwell: Oxford, UK, 1983. [Google Scholar]
- Blankenship, R.E. Molecular Mechanisms of Photosynthesis; Blackwell Science: Oxford, UK, 2002. [Google Scholar]
- Ruban, A. The Photosynthetic Membrane: Molecular Mechanisms and Biophysics of Light Harvesting; Wiley-Blackwell: Oxford, UK, 2012. [Google Scholar]
- Kramer, D.M.; Avenson, T.J.; Edwards, G.E. Dynamic flexibility in the light reactions of photosynthesis governed by both electron and proton transfer reactions. Trends Plant. Sci. 2004, 9, 349–357. [Google Scholar] [CrossRef]
- Skulachev, V.P.; Vyssokikh, M.Y.; Chernyak, B.V.; Mulkidjanian, A.Y.; Skulachev, M.V.; Shilovsky, G.A.; Lyamzaev, K.G.; Borisov, V.B.; Severin, F.F.; Sadovnichii, V.A. Six functions of respiration: Isn’t it time to take control over ROS production in mitochondria, and aging along with it? Int. J. Mol. Sci. 2023, 24, 12540. [Google Scholar] [CrossRef]
- Eberhard, S.; Finazzi, G.; Wollman, F.A. The dynamics of photosynthesis. Annu. Rev. Genet. 2008, 42, 463–515. [Google Scholar] [CrossRef]
- Haehnel, W. Photosynthetic electron transport in higher plants. Annu. Rev. Plant Physiol. 1984, 35, 659–693. [Google Scholar] [CrossRef]
- Cramer, W.A.; Hasan, S.S. Structure-function of the cytochrome b6f lipoprotein complex. In cytochrome complexes: Evolution, structures, energy transduction, and signaling. Adv. Photosynth. Respir. 2016, 41, 177–207. [Google Scholar]
- Mulkidjanian, A.Y. Proton translocation by the cytochrome bc1 complexes of phototrophic bacteria: Introducing the activated Q-cycle. Photochem. Photobiol. Sci. 2007, 6, 19–34. [Google Scholar] [CrossRef] [PubMed]
- Tikhonov, A.N. pH-Dependent regulation of electron transport and ATP synthesis in chloroplasts. Photosynth. Res. 2013, 116, 511–534. [Google Scholar] [CrossRef] [PubMed]
- Sarewicz, M.; Pintscher, S.; Pietras, R.; Borek, A.; Bujnowicz, Ł.; Hanke, G.; Cramer, W.A.; Finazzi, G.; Osyczka, A. Catalytic reactions and energy conservation in the cytochrome bc1 and b6f complexes of energy-transducing membranes. Chem. Rev. 2021, 121, 2020–2108. [Google Scholar] [CrossRef]
- Malone, L.A.; Proctor, M.S.; Hitchcock, A.; Hunter, C.N.; Johnson, M.P. Cytochrome b6f—Orchestrator of photosynthetic electron transfer. Biochim. Biophis. Acta 2021, 1862, 148380. [Google Scholar] [CrossRef]
- Li, Z.; Wakao, S.; Fischer, B.B.; Niyogi, K.K. Sensing and responding to excess light. Annu. Rev. Plant Biol. 2009, 60, 239–260. [Google Scholar] [CrossRef]
- Gurrieri, L.; Fermani, S.; Zaffagnini, M.; Sparla, F.; Trost, P. Calvin–Benson cycle regulation is getting complex. Trends Plant Sci. 2021, 26, 898–912. [Google Scholar] [CrossRef]
- Pfalz, J.; Liebers, M.; Hirth, M.; Grübler, B.; Holtzegel, U.; Schröter, Y.; Dietzel, L.; Pfannschmidt, T. Environmental control of plant nuclear gene expression by chloroplasts redox signals. Front. Plant Sci. 2012, 3, 129–137. [Google Scholar] [CrossRef]
- Yamori, W.; Hikosaka, K.; Way, D.A. Temperature response of photosynthesis in C3, C4, and CAM plants: Temperature acclimation and temperature adaptation. Photosynth. Res. 2014, 119, 101–117. [Google Scholar] [CrossRef]
- Tikhonov, A.N.; Vershubskii, A.V. Temperature-dependent regulation of electron transport and ATP synthesis in chloroplasts in vitro and in silico. Photosynth. Res. 2020, 146, 299–329. [Google Scholar] [CrossRef] [PubMed]
- Kromdijk, J.; Glowacka, K.; Leonelli, L.; Gabilly, S.T.; Iwai, M.; Niyogi, K.K.; Long, S.P. Improving photosynthesis and crop productivity by accelerating recovery from photoprotection. Science 2016, 354, 857–861. [Google Scholar] [CrossRef] [PubMed]
- Armbruster, U.; Galvis, V.C.; Kunz, H.-H.; Strand, D.D. The regulation of the chloroplast proton motive force plays a key role for photosynthesis in fluctuating light. Curr. Opin. Plant Biol. 2017, 37, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Turina, P.; Petersen, J.; Gräber, P. Thermodynamics of proton transport coupled ATP synthesis. Biochim. Biophys. Acta 2016, 1857, 653–664. [Google Scholar] [CrossRef]
- Turina, P. Influence of transmembrane electrochemical proton gradient on catalysis and regulation of the H+-ATP synthase from Rhodobacter capsulatus. Bioelectrochem. Bioenerg. 1994, 33, 31–43. [Google Scholar] [CrossRef]
- Junesch, U.; Gräber, P. The rate of ATP-synthesis as a function of ΔpH and Δψ catalyzed by the active, reduced H+-ATPase from chloroplasts. FEBS Lett. 1991, 294, 275–278. [Google Scholar] [CrossRef]
- Soga, N.; Kinosita, K., Jr.; Yoshida, M.; Suzuki, T. Kinetic equivalence of transmembrane pH and electrical potential differences on ATP synthesis. J. Biol. Chem. 2012, 287, 9633–9639. [Google Scholar] [CrossRef]
- Cruz, J.A.; Sacksteder, C.A.; Kanazawa, A.; Kramer, D.M. Contribution of electric field (Δψ) to steady-state transthylakoid proton motive force (pmf) in vivo and in vitro. Control of pmf parsing into Δψ and ΔpH by ionic strength. Biochemistry 2001, 40, 1226–1237. [Google Scholar] [CrossRef]
- Kramer, D.M.; Sacksteder, C.A.; Cruz, J.A. Balancing the central roles of the thylakoid proton gradient. Trends Plant Sci. 2003, 8, 27–32. [Google Scholar] [CrossRef]
- Johnson, M.P.; Ruban, A.V. Rethinking the existence of a steady-state Δψ component of the proton motive force across plant thylakoid membranes. Photosynth. Res. 2014, 119, 233–242. [Google Scholar] [CrossRef]
- Tikhonov, A.N.; Agafonov, R.V.; Grigor’ev, I.A.; Kirilyuk, I.A.; Ptushenko, V.V.; Trubitsin, B.V. Spin-probes designed for measuring the intrathylakoid pH in chloroplasts. Biochim. Biophys. Acta 2008, 1777, 285–294. [Google Scholar] [CrossRef] [PubMed]
- Barber, J. Influence of surface charges on thylakoid structure and function. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1982, 33, 261–295. [Google Scholar] [CrossRef]
- Nickolls, D.G.; Ferguson, S.J. Bioenergetics 3; Academic Press, Inc.: New York, NY, USA, 2002. [Google Scholar]
- Skulachev, V.P.; Bogachev, A.V.; Kasparinsky, F.O. Principles of Bioenergetics; Springer: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- van Meer, G.; Voelker, D.R.; Feigenson, G.W. Membrane lipids: Where they are and how they behave. Nat. Rev. Mol. Cell Biol. 2008, 9, 112–124. [Google Scholar] [CrossRef] [PubMed]
- Horvath, S.E.; Daum, G. Lipids of mitochondria. Prog. Lipid Res. 2013, 52, 590–614. [Google Scholar] [CrossRef]
- Osman, C.; Voelker, D.R.; Langer, T. Making heads or tails of phospholipids in mitochondria. J. Cell Biol. 2011, 192, 7–16. [Google Scholar] [CrossRef]
- Mejia, E.M.; Hatch, G.M. Mitochondrial phospholipids: Role in mitochondrial function. J. Bioenerg. Biomembr. 2016, 48, 99–112. [Google Scholar] [CrossRef]
- Mårtensson, C.U.; Doan, K.N.; Becker, T. Effects of lipids on mitochondrial functions. Biochim. Biopys. Acta 2017, 1862, 102–113. [Google Scholar] [CrossRef]
- Wada, H.; Murata, N. Lipids in thylakoid membranes and photosynthetic cells. In Lipids in Photosynthesis: Essentials and Regulatory Functions; Springer: Berlin/Heidelberg, Germany, 2009; pp. 1–9. [Google Scholar]
- Zhou, Y.; vom Dorp, K.; Dörmann, P.; Hölz, G. Chloroplast Lipids. In Chloroplasts. Current Research and Future Trends; Kirchhoff, H., Ed.; Caister Academic Press: Poole, UK, 2016; pp. 1–24. [Google Scholar]
- Wada, H.; Murata, N. The essential role phosphatidylglycerol in photosynthesis. Photosynth. Res. 2007, 92, 205–215. [Google Scholar] [CrossRef]
- de Gier, J. Permeability barriers formed by membrane lipids. Bioelectrochem. Bioenerg. 1992, 27, 1–10. [Google Scholar] [CrossRef]
- Fuks, B.; Homble, F. Permeability and electrical properties of planar lipid membranes from thylakoid lipids. Biophys. J. 1994, 66, 1404–1414. [Google Scholar] [CrossRef]
- Hauser, H.; Oldani, D.; Phillips, M.C. Mechanism of ion escape from phosphatidylcholine and phosphatidylserine single bilayer vesicles. Biochemistry 1973, 12, 4507–4517. [Google Scholar] [CrossRef] [PubMed]
- Deamer, D.W. Proton permeation of lipid bilayers. J. Bioenerg. Biomembr. 1987, 19, 457–479. [Google Scholar] [CrossRef] [PubMed]
- Gutknecht, J. Proton/hydroxide conductance and permeability through phospholipid bilayer membranes. Proc. Natl. Acad. Sci. USA 1987, 84, 6443–6446. [Google Scholar] [CrossRef]
- Marrink, S.J.; Jahnig, F.; Berendsen, H.J. Proton transport across transient single file water pores in a lipid membrane studied by molecular dynamics simulations. Biophys. J. 1996, 71, 632–647. [Google Scholar] [CrossRef] [PubMed]
- Mulkidjanian, A.Y.; Heberle, J.; Cherepanov, D.A. Protons @ interfaces: Implications for biological energy conversion. Biochim. Biophys. Acta 2006, 1757, 913–930. [Google Scholar] [CrossRef] [PubMed]
- Mulkidjanian, A.Y. Proton in the well and through the desolvation barrier. Biochem. Biophys. Acta 2006, 1757, 415–427. [Google Scholar] [CrossRef]
- Springer, A.; Hagen, V.; Cherepanov, D.A.; Antonenko, Y.N.; Pohl, P. Protons migrate along interfacial water without significant contributions from jumps between ionizable groups on the membrane surface. Proc. Natl. Acad. Sci. USA 2011, 108, 14461–14466. [Google Scholar] [CrossRef]
- Bozdaganyan, M.E.; Lokhmatikov, A.V.; Voskoboynikova, N.; Cherepanov, D.A.; Steinhoff, H.-J.; Shaitan, K.V.; Mulkidjanian, A.Y. Proton leakage across lipid bilayers: Oxygen atoms of phospholipid ester linkers align water molecules into transmembrane water wires. Biochim. Biophys. Acta 2019, 1860, 439–451. [Google Scholar] [CrossRef]
- Hasan, S.S.; Yamashita, E.; Baniulis, D.; Cramer, W.A. Quinone-dependent proton transfer pathways in the photosynthetic cytochrome b6f complex. Proc. Natl. Acad. Sci. USA 2013, 110, 4297–4302. [Google Scholar] [CrossRef]
- Herdean, A.; Teardo, E.; Nilsson, A.K.; Pfeil, B.E.; Johansson, O.N.; Unnep, R.; Nagy, G.; Zsiros, O.; Dana, S.; Solymosi, K.; et al. A voltage-dependent chloride channel fine-tunes photosynthesis in plants. Nat. Commun. 2016, 7, 11654. [Google Scholar] [CrossRef]
- Armbruster, U.; Carrillo, L.R.; Venema, K.; Pavlovic, L.; Schmidtmann, E.; Kornfeld, A.; Jahns, P.; Berry, J.A.; Kramer, D.M.; Jonikas, M.C. Ion antiport accelerates photosynthetic acclimation in fluctuating light environments. Nat. Commun. 2014, 5, 5439. [Google Scholar] [CrossRef] [PubMed]
- Kunz, H.H.; Gierth, M.; Herdean, A.; Satoh-Cruz, M.; Kramer, D.M.; Spetea, C.; Schroeder, J.I. Plastidial transporters KEA1, -2, and -3 are essential for chloroplast osmoregulation, integrity, and pH regulation in Arabidopsis. Proc. Natl. Acad. Sci. USA 2014, 111, 7480–7485. [Google Scholar] [CrossRef] [PubMed]
- Suorsa, M.; Jarvi, S.; Grieco, M.; Nurmi, M.; Pietrzykowska, M.; Rantala, M.; Kangasjarvi, S.; Paakkarinen, V.; Tikkanen, M.; Jansson, S.; et al. PROTON GRADIENT REGULATION5 is essential for proper acclimation of Arabidopsis photosystem I to naturally and artificially fluctuating light conditions. Plant Cell 2012, 24, 2934–2948. [Google Scholar] [CrossRef]
- Suorsa, M.; Grieco, M.; Jarvi, S.; Gollan, P.J.; Kangasjarvi, S.; Tikkanen, M.; Aro, E.M. PGR5 ensures photosynthetic control to safeguard photosystem I under fluctuating light conditions. Plant Signal. Behav. 2013, 8, e22741. [Google Scholar] [CrossRef]
- Davis, G.A.; Kanazawa, A.; Schottler, M.A.; Kohzuma, K.; Froehlich, J.E.; Rutherford, A.W.; Satoh-Cruz, M.; Minhas, D.; Tietz, S.; Dhingra, A.; et al. Limitations to photosynthesis by proton motive force-induced photosystem II photodamage. eLife 2016, 5, e16921. [Google Scholar] [CrossRef]
- Peschek, G.A. Respiratory electron transport. In The Cyanobacteria; Fay, P., Van Baalen, C., Eds.; Elsevier: Amsterdam, The Netherlands, 1987; pp. 119–161. [Google Scholar]
- Schmetterer, G. Cyanobacterial respiration. In The Molecular Biology of Cyanobacteria; Bryant, D.A., Ed.; Kluwer: Dordrecht, The Netherlands, 1994; pp. 409–435. [Google Scholar]
- Kobayashi, K.; Endo, K.; Wada, H. Roles of lipids in photosynthesis. In Lipids in Plant and Algae Development. Subcellular Biochemistry; Nakamura, Y., Li-Beisson, Y., Eds.; Springer: Berlin/Heidelberg, Germany, 2016; Volume 86, pp. 21–49. [Google Scholar]
- Grinius, L.L.; Jasaitis, A.A.; Kadziauskas, P.; Liberman, E.A.; Skulachev, V.P.; Topali, V.P.; Tsofina, L.M.; Vladimirova, M.A. Conversion of biomembrane-produced energy into electric form. I. Submitochondrial particles. Biochim. Biophys. Acta 1970, 216, 1–12. [Google Scholar] [CrossRef]
- Isaev, P.I.; Liberman, E.A.; Samuilov, V.D.; Skulachev, V.P.; Tsofina, L.M. Conversion of biomembrane-produced energy into electric form. III. Chromatophores of Rhodospirillum rubrum. Biochim. Biophys. Acta 1970, 216, 22–29. [Google Scholar] [CrossRef]
- Kayushin, L.P.; Skulachev, V.P. Bacteriorhodopsin as an electrogenic proton pump: Reconstitution of bacteriorhodopsin proteoliposomes generating Δψ and ΔpH. FEBS Lett. 1974, 39, 39–42. [Google Scholar] [CrossRef]
- Drachev, L.A.; Kaulen, A.D.; Semenov, A.; Severina, I.I.; Skulachev, V.P. Lipid-impregnated filters as a tool for studying the electric current-generating proteins. Analyt. Biochem. 1979, 96, 250–262. [Google Scholar] [CrossRef]
- Junge, W.; Witt, H.T. On the ion transport system of photosynthesis. Investigations on a molecular level. Z. Naturforsch. 1968, 23, 244–246. [Google Scholar] [CrossRef]
- Jackson, J.B.; Crofts, A.R. The high energy state in chromatophores from Rhodopseudomonas sphaeroides. FEBS Lett. 1969, 4, 185–189. [Google Scholar] [CrossRef] [PubMed]
- Joliot, P.; Joliot, A. Electron transfer between the two photosystems. I. Flash excitation under oxidizing conditions. Biochim. Biophys. Acta 1984, 765, 210–218. [Google Scholar] [CrossRef]
- Kurashov, V.; Gorka, M.; Milanovsky, G.E.; Johnson, W.; Cherepanov, D.A.; Semenov, A.; Golbeck, J.H. Critical evaluation of electron transfer kinetics in P700–FA/FB, P700–FX, and P700–A1 Photosystem I core complexes in liquid and in trehalose glass. Biochim. Biophys. Acta 2018, 1859, 1288–1301. [Google Scholar] [CrossRef] [PubMed]
- Drachev, L.A.; Jasaitis, A.A.; Kaulen, A.D.; Kondrashin, A.A.; Liberman, E.A.; Nemecek, I.B.; Ostroumov, S.A.; Semenov, A.Y.; Skulachev, V.P. Direct measurement of electric current generation by cytochrome oxidase, H+-ATPase and bacteriorhodopsin. Nature 1974, 249, 321–324. [Google Scholar] [CrossRef]
- Drachev, L.A.; Frolov, V.N.; Kaulen, A.D.; Liberman, E.A.; Ostroumov, S.A.; Plakunova, V.G.; Semenov, A.; Skulachev, V.P. Reconstitution of biological molecular generators of electric current. Bacteriorhodopsin. J. Biol. Chem. 1976, 251, 7059–7065. [Google Scholar] [CrossRef]
- Drachev, L.A.; Kaulen, A.D.; Khitrina, L.V.; Skulachev, V.P. Fast stages of photoelectric processes in biological membranes. I. Bacteriorhodopsin. Eur. J. Biochem. 1981, 117, 461–470. [Google Scholar] [CrossRef]
- Kramer, D.M.; Sacksteder, C.A.; Cruz, J.A. How acidic is the lumen? Photosynth. Res. 1999, 60, 151–163. [Google Scholar] [CrossRef]
- Junge, W.; Ausländer, W.; McGeer, A.J.; Runge, T. The buffering capacity of the internal phase of thylakoids and the magnitude of the pH changes inside under flashing light. Biochim. Biophys. Acta 1979, 546, 121–141. [Google Scholar] [CrossRef]
- Junge, W. Electrogenic reactions and proton pumping in green plant photosynthesis. Curr. Top. Membr. Transp. 1982, 16, 431–463. [Google Scholar]
- Grzesiek, S.; Dencher, N.A. The ‘pH’-probe 9-aminoacridine: Response time, binding behaviour and dimerization at the membrane. Biochim. Biophys. Acta 1988, 938, 411–424. [Google Scholar] [CrossRef]
- Lolkema, J.S.; Hellingwerf, K.J.; Konings, W.N. The effect of ‘probe binding’ on the quantitative determination of the proton-motive force in bacteria. Biochim. Biophys. Acta 1982, 681, 85–94. [Google Scholar] [CrossRef]
- Azzone, G.F.; Piewboron, D.; Zoratti, M. Determination of the proton electrochemical gradient across biological membranes. Curr. Top. Bioenerg. 1984, 13, 1–77. [Google Scholar]
- Rottenberg, H.; Grünwald, T.; Avron, M. Determination of ΔpH in chloroplasts: I. Distribution of (14C) methylamine. Eur. J. Biochem. 1972, 25, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Schuldiner, S.; Rottenberg, H.; Avron, M. Determination of ΔpH in chloroplasts: II. Fluorescent amines as a probe for the determination of ΔpH in chloroplasts. Eur. J. Biochem. 1972, 25, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Rottenberg, H.; Grünwald, T. Determination of ΔpH in chloroplasts: III. Ammonium uptake as a measure of pH in chloroplasts and subchloroplast particles. Eur. J. Biochem. 1972, 25, 71–74. [Google Scholar] [CrossRef]
- Cafiso, D.S.; Hubbel, W.L. Estimation of transmembrane pH gradients from phase equilibria of spin-labeled amines. Biochemistry 1978, 17, 3871–3877. [Google Scholar] [CrossRef]
- Quintanilha, A.T.; Mehlhorn, R.J. pH gradient across thylakoid membranes measured with a spin labeled amine. FEBS Lett. 1978, 91, 104–108. [Google Scholar] [CrossRef]
- Tikhonov, A.N. Photosynthetic electron and proton transport in chloroplasts: EPR study of ΔpH generation, an Overview. Cell Biochem. Biophys. 2017, 75, 421–432. [Google Scholar] [CrossRef]
- Trubitsin, B.V.; Tikhonov, A.N. Determination of a transmembrane pH difference in chloroplasts with a spin label Tempamine. J. Magn. Reson. 2003, 163, 257–269. [Google Scholar] [CrossRef]
- Rumberg, B.; Siggel, U. pH changes in the inner phase of the thylakoids during photosynthesis. Naturwissenschaften 1969, 56, 130–132. [Google Scholar] [CrossRef]
- Tikhonov, A.N.; Khomutov, G.B.; Ruuge, E.K.; Blumenfeld, L.A. Electron transport control in chloroplasts. Effects of photosynthetic control monitored by the intrathylakoid pH. Biochem. Biophys. Acta 1981, 637, 321–333. [Google Scholar] [CrossRef]
- Tikhonov, A.N.; Khomutov, G.B.; Ruuge, E.K. Electron transport control in chloroplasts. Effects of magnesium ions on the electron flow between two photosystems. Photobiochem. Photobiophys. 1984, 8, 261–269. [Google Scholar]
- Khramtsov, V.V.; Volodarsky, L.B. Use of imidazoline nitroxides in studies of chemical reactions: ESR measurement of concentration and reactivity of protons, thiols and nitric oxide. In Biological Magnetic Resonance; Berliner, L.J., Ed.; Plenum Press: London, UK, 1998; Volume 4, pp. 109–180. [Google Scholar]
- Pogoryelov, D.; Reichen, C.; Klyszejko, A.L.; Brunisholz, R.; Muller, D.J.; Dimroth, P.; Meier, T. The oligomeric state of c rings from cyanobacterial F-ATP synthases varies from 13 to 15. J. Bacteriol. 2007, 189, 5895–5902. [Google Scholar] [CrossRef] [PubMed]
- Pogoryelov, D.; Klyszejko, A.L.; Krasnoselska, G.O.; Heller, E.M.; Leone, V.; Langer, J.D.; Vonck, J.; Müller, D.J.; Faraldo-Gómez, J.D.; Meier, T. Engineering rotor ring stoichiometries in the ATP synthase. Proc. Natl. Acad. Sci. USA 2012, 109, E1599–E1608. [Google Scholar] [CrossRef]
- Silverstein, T.P. An exploration of how the thermodynamic efficiency of bioenergetic membrane systems varies with c-subunit stoichiometry of F1F0 ATP synthases. J. Bioenerg. Biomembr. 2014, 46, 229–241. [Google Scholar] [CrossRef]
- Hahn, A.; Vonck, J.; Mills, D.J.; Meier, T.; Kühlbrandt, W. Structure, mechanism, and regulation of the chloroplast ATP synthase. Science 2018, 360, eaat4318. [Google Scholar] [CrossRef]
- Davis, G.A.; Kramer, D.M. Optimization of ATP synthase c–rings for oxygenic photosynthesis. Front. Plant Sci. 2020, 10, 1778. [Google Scholar] [CrossRef]
- Castle, J.D.; Hubbell, W.L. Estimation of membrane surface potential and charge density from the phase equilibrium of a paramagnetic amphiphile. Biochemistry 1976, 15, 4818–4831. [Google Scholar] [CrossRef]
- Cafiso, S.; Hubbell, W.L. EPR determination of membrane potentials. Annu. Rev. Biophys. Bioeng. 1981, 10, 217–244. [Google Scholar] [CrossRef]
- Khomutov, G.B.; Gilmiarova, S.G. A surface potential of the thylakoid membrane and energy transduction processes in chloroplasts. II. A surface potential and buffer properties chloroplast membranes. Biol. Membr. 1989, 6, 705–719. [Google Scholar]
- Vershubskii, A.V.; Trubitsin, B.V.; Priklonskii, V.I.; Tikhonov, A.N. Lateral heterogeneity of the proton potential along the thylakoid membranes of chloroplasts. Biochim. Biophys. Acta 2017, 1859, 388–401. [Google Scholar] [CrossRef] [PubMed]
- Haraux, F. Localized or delocalizes protons and ATP synthesis in biomembranes. Physiol. Vég. 1985, 23, 397–410. [Google Scholar]
- Sigalat, C.; de Kouchkovsky, Y.; Haraux, F.; de Kouchkovsky, F. Shift from localized to delocalized energy coupling in thylakoids by permeant amines. Biochim. Biophys. Acta 1988, 934, 375–388. [Google Scholar] [CrossRef]
- Drachev, L.A.; Frolov, V.N.; Kaulen, A.D.; Kondrashin, A.A.; Samuilov, V.D.; Semenov, A.Y.; Skulachev, V.P. Generation of electric current by chromatophores of Rhodospirillum rubrum and reconstitution of electrogenic function in subchromatophore pigment–protein complexes. Biochim. Biophys. Acta 1976, 440, 637–660. [Google Scholar] [CrossRef] [PubMed]
- Jordan, P.; Fromme, P.; Witt, H.T.; Klukas, O.; Saenger, W.; Krauss, N. Three-dimensional structure of cyanobacterial photosystem I at 2.5 angstroms resolution. Nature 2001, 411, 909–917. [Google Scholar] [CrossRef]
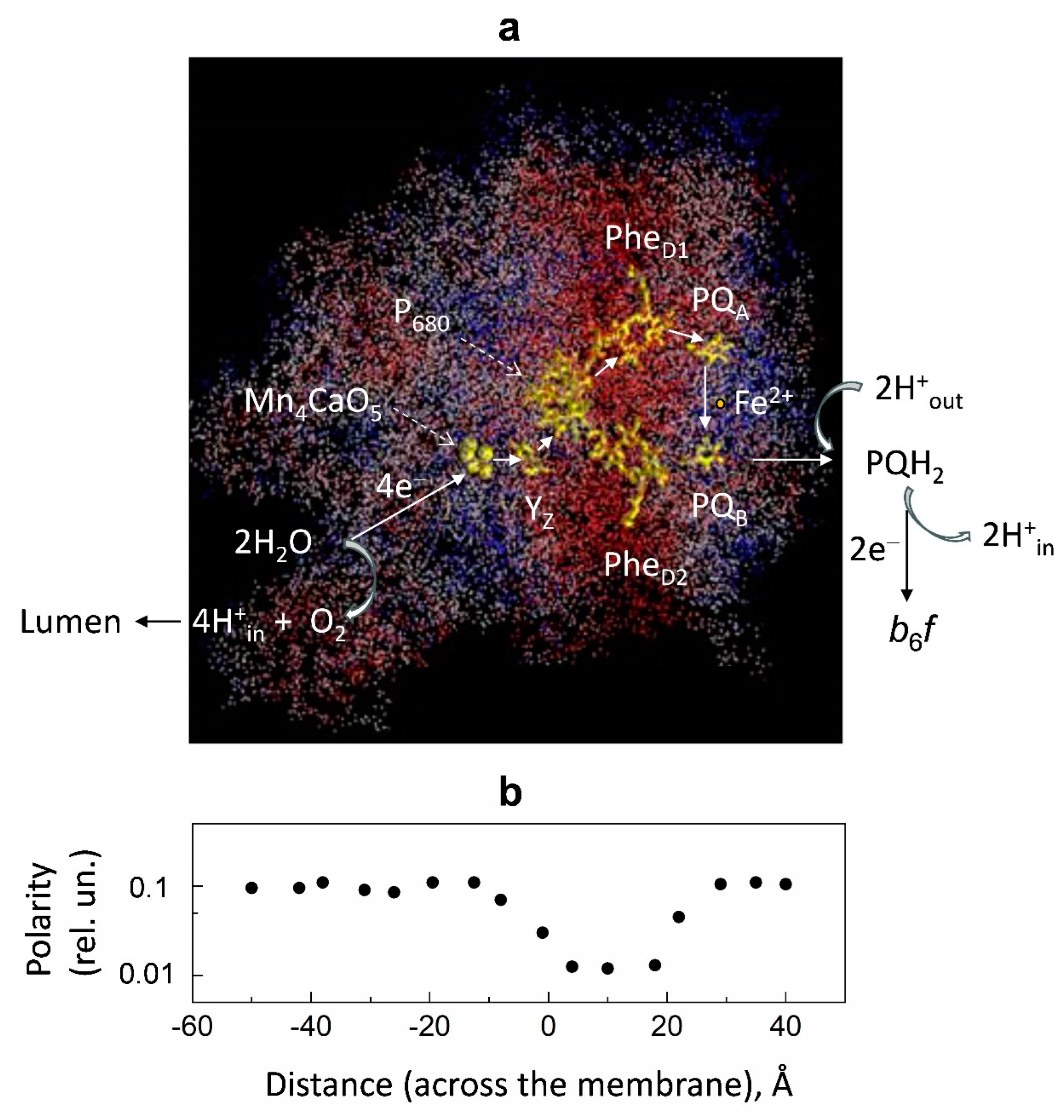
- Loll, B.; Kern, J.; Saenger, W.; Zouni, A.; Biesiadka, J. Towards complete cofactor arrangement in the 3.0 Å resolution structure of photosystem II. Nature 2005, 438, 1040–1044. [Google Scholar] [CrossRef]
- Semenov, A.; Cherepanov, D.A.; Mamedov, M.D. Electrogenic reactions and dielectric properties of photosystem II. Photosynth. Res. 2008, 98, 121–130. [Google Scholar] [CrossRef]
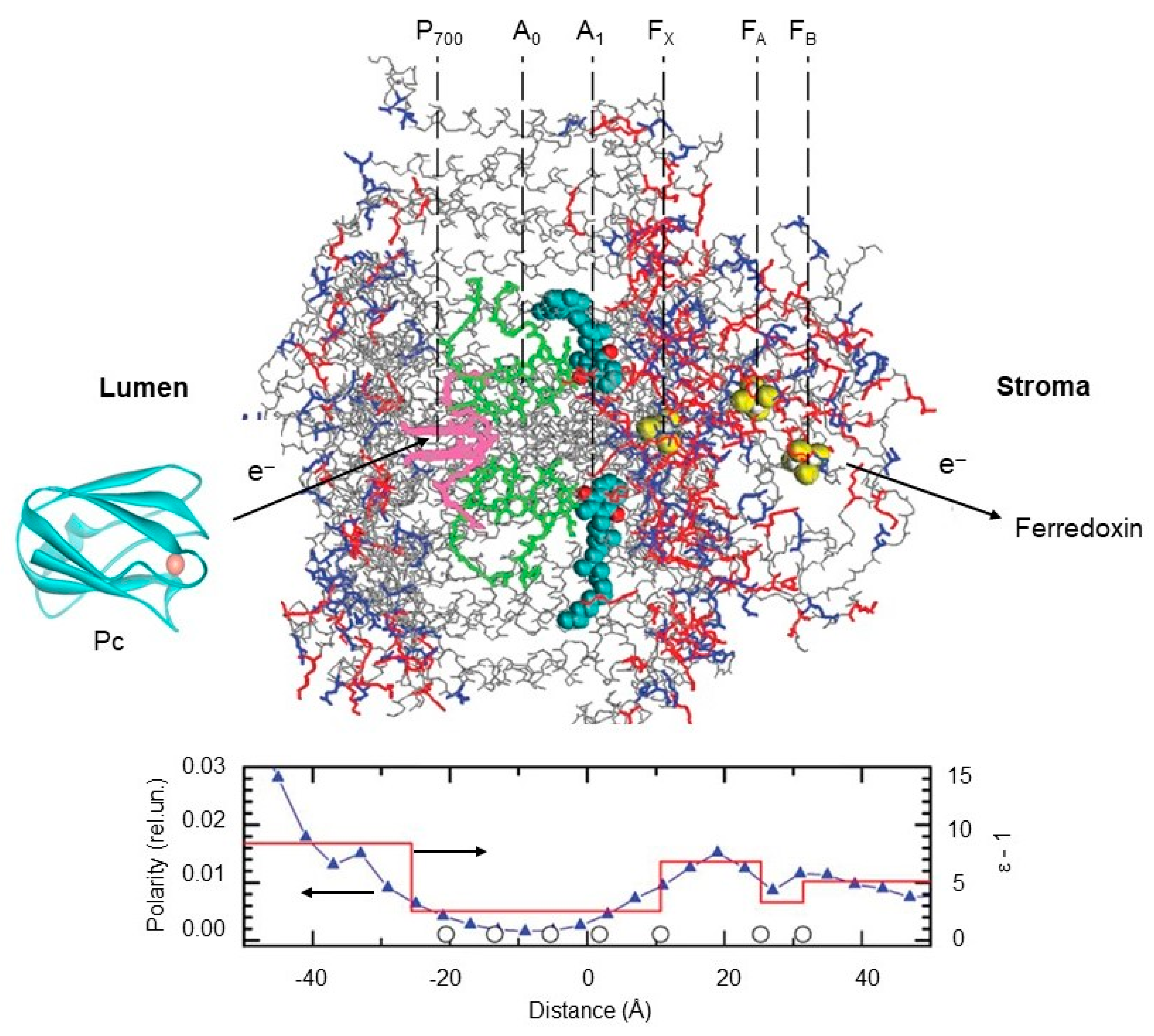
- Ptushenko, V.V.; Cherepanov, D.A.; Krishtalik, L.I.; Semenov, A. Semi-continuum electrostatic calculations of redox potentials in photosystem I. Photosynth. Res. 2008, 97, 55–74. [Google Scholar] [CrossRef]
- Gorka, M.; Cherepanov, D.A.; Semenov, A.; Golbeck, J.H. Control of Electron Transfer by Protein Dynamics in Photosynthetic Reaction Centers. Crit. Rev. Biochem. Mol. Biol. 2020, 55, 425–468. [Google Scholar] [CrossRef]
- Kell, D.B. Localized protonic coupling: Overview and critical evaluation of techniques. Methods Enzymol. 1986, 127, 538–557. [Google Scholar]
- Dilley, R.A.; Theg, S.M.; Beard, W.A. Membrane–proton interactions in chloroplast bioenergetics: Localized proton domains. Annu. Rev. Plant Physiol. 1987, 37, 348–389. [Google Scholar]
- Ewy, R.G.; Dilley, R.A. Distinguishing between luminal and localized proton buffering pools in thylakoid membranes. Plant Physiol. 2000, 122, 583–595. [Google Scholar] [CrossRef] [PubMed]
- Dilley, R.A. On why thylakoids energize ATP formation using either delocalized or localized proton gradients—A Ca2+ mediated role in thylakoid stress responses. Photosynth. Res. 2004, 80, 245–263. [Google Scholar] [CrossRef] [PubMed]
- Wolf, S.; Freier, E.; Gerwert, K. A delocalized proton-binding site within a membrane protein. Biophys. J. 2014, 107, 174–184. [Google Scholar] [CrossRef] [PubMed]
- Haraux, F.; de Kouchkovsky, Y. Measurement of chloroplast internal protons with 9-AA. Probe binding, dark proton gradient, and salt effects. Biochim. Biophys. Acta 1980, 592, 158–168. [Google Scholar]
- Hong, Y.Q.; Junge, W. Localized or delocalized protons in photophosphorylation? On the accessibility of the thylakoid lumen for ions and buffers. Biochim. Biophys. Acta 1983, 722, 197–208. [Google Scholar] [CrossRef]
- Hope, A.B.; Matthews, D.B. Adsorption of amines to thylakoid surfaces and estimation of ΔpH. Aust. J. Plant Physiol. 1985, 12, 9–19. [Google Scholar] [CrossRef]
- Chudakov, D.M.; Matz, M.V.; Lukyanov, S.; Lukyanov, K.A. Fluorescent proteins and their applications in imaging living cells and tissues. Physiol. Rev. 2010, 90, 1103–1163. [Google Scholar] [CrossRef]
- Rodriguez, E.A.; Campbell, R.E.; Lin, J.Y.; Lin, M.Z.; Miyawaki, A.; Palmer, A.E.; Shu, X.; Zhang, J.; Tsien, R.Y. The Growing and Glowing Toolbox of Fluorescent and Photoactive Proteins. Trends Biochem. Sci. 2017, 42, 111–129. [Google Scholar] [CrossRef]
- Rieger, B.; Junge, W.; Busch, K.B. Lateral pH gradient between OXPHOS complex IV and F0F1 ATP-synthase in folded mitochondrial membranes. Nature Commun. 2014, 5, 3103. [Google Scholar] [CrossRef]
- Toth, A.; Meyrat, A.; Stoldt, S.; Santiago, R.; Wenzel, D.; Jakobs, S.; von Ballmoos, C.; Ott, M. Kinetic coupling of the respiratory chain with ATP synthase, but not proton gradients, drives ATP production in cristae membranes. Proc. Natl. Acad. Sci. USA 2020, 117, 2412–2421. [Google Scholar] [CrossRef] [PubMed]
- Sarewicz, M.; Szwalec, M.; Pintscher, S.; Indyka, P.; Rawski, M.; Pietras, R.; Mielecki, B.; Koziej, Ł.; Jaciuk, M.; Glatt, S.; et al. High-resolution cryo-EM structures of plant cytochrome b6f at work. Sci. Adv. 2023, 9, eadd9688. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.Z.; Reisner, E. Advancing photosystem II photoelectrochemistry for semi-artificial photosynthesis. Nat. Rev. Chem. 2020, 4, 6–21. [Google Scholar] [CrossRef]
- Szewczyk, S.; Białek, R.; Burdziński, G.; Gibasiewicz, K. Photovoltaic activity of electrodes based on intact photosystem I electrodeposited on bare conducting glass. Photosyn. Res. 2020, 144, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Teodor, A.H.; Bruce, B.D. Putting photosystem I to work: Truly green energy. Trends Biotechnol. 2020, 38, 1329–1342. [Google Scholar] [CrossRef] [PubMed]
- Ros, C.; Andreu, T.; Morante, J.R. Photoelectrochemical water splitting: A road from stable metal oxides to protected thin film solar cells. J. Mater. Chem. A 2020, 8, 10625. [Google Scholar] [CrossRef]
- Vitukhnovskaya, L.A.; Zaspa, A.A.; Semenov, A.; Mamedov, M.D. Conversion of light into electricity in a semi-synthetic system based on photosynthetic bacterial chromatophores. Biochim. Biophys. Acta 2023, 1864, 148975. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Semenov, A.Y.; Tikhonov, A.N. Electrometric and Electron Paramagnetic Resonance Measurements of a Difference in the Transmembrane Electrochemical Potential: Photosynthetic Subcellular Structures and Isolated Pigment–Protein Complexes. Membranes 2023, 13, 866. https://doi.org/10.3390/membranes13110866
Semenov AY, Tikhonov AN. Electrometric and Electron Paramagnetic Resonance Measurements of a Difference in the Transmembrane Electrochemical Potential: Photosynthetic Subcellular Structures and Isolated Pigment–Protein Complexes. Membranes. 2023; 13(11):866. https://doi.org/10.3390/membranes13110866
Chicago/Turabian StyleSemenov, Alexey Yu., and Alexander N. Tikhonov. 2023. "Electrometric and Electron Paramagnetic Resonance Measurements of a Difference in the Transmembrane Electrochemical Potential: Photosynthetic Subcellular Structures and Isolated Pigment–Protein Complexes" Membranes 13, no. 11: 866. https://doi.org/10.3390/membranes13110866