

Recording Sodium Self-Inhibition of Epithelial Sodium Channels Using Automated Electrophysiology in Xenopus Oocytes


 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plasmids and cRNA Synthesis
2.2. Site-Directed Mutagenesis
2.3. ENaC Expression in Xenopus laevis Oocytes
2.4. Automated Two-Electrode Voltage-Clamp (TEVC) Electrophysiology
2.5. Chemicals and Reagents
2.6. Molecular Modelling
2.7. Data Analyses and Statistics
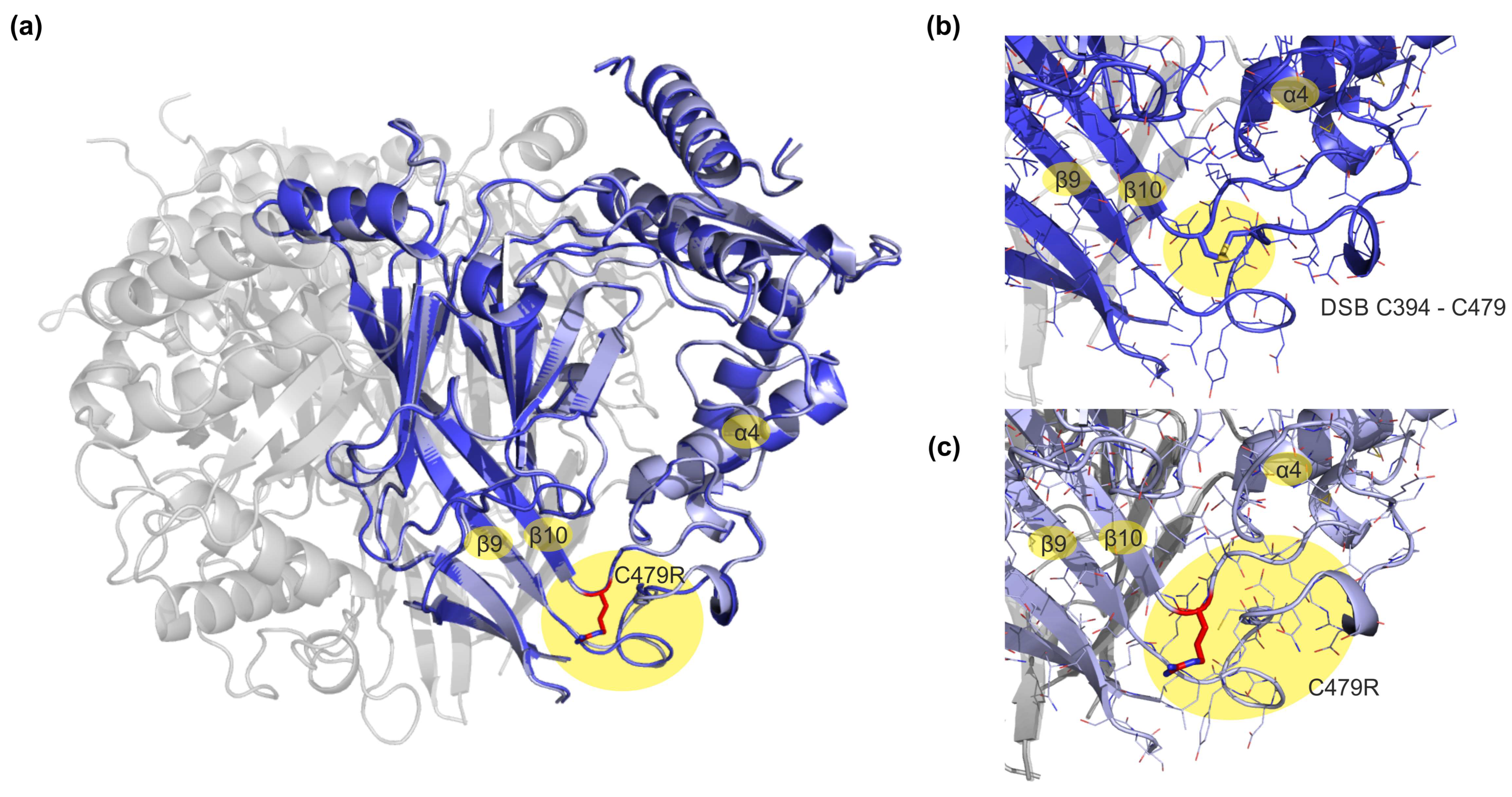
3. Results
3.1. Evaluating Automated SSI Recordings Using Mammalian and Amphibian ENaC Orthologs
3.2. Recording SSI in a Human ENaC Mutation Associated with Liddle Syndrome
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van den Bosch, J.J.O.N.; Hessels, N.R.; Visser, F.W.; Krikken, J.A.; Bakker, S.J.L.; Riphagen, I.J.; Navis, G.J. Plasma Sodium, Extracellular Fluid Volume, and Blood Pressure in Healthy Men. Physiol. Rep. 2021, 9, e15103. [Google Scholar] [CrossRef]
- Firsov, D.; Schild, L.; Gautschi, I.; Mérillat, A.M.; Schneeberger, E.; Rossier, B.C. Cell Surface Expression of the Epithelial Na Channel and a Mutant Causing Liddle Syndrome: A Quantitative Approach. Proc. Natl. Acad. Sci. USA 1996, 93, 15370–15375. [Google Scholar] [CrossRef] [PubMed]
- Shimkets, R.A.; Warnock, D.G.; Bositis, C.M.; Nelson-Williams, C.; Hansson, J.H.; Schambelan, M.; Gill, J.R.; Ulick, S.; Milora, R.V.; Findling, J.W.; et al. Liddle’s Syndrome: Heritable Human Hypertension Caused by Mutations in the β Subunit of the Epithelial Sodium Channel. Cell 1994, 79, 407–414. [Google Scholar] [CrossRef]
- Pitzer, A.L.; Van Beusecum, J.P.; Kleyman, T.R.; Kirabo, A. ENaC in Salt-Sensitive Hypertension: Kidney and Beyond. Curr. Hypertens. Rep. 2020, 22, 69. [Google Scholar] [CrossRef] [PubMed]
- Ray, E.C.; Chen, J.; Kelly, T.N.; He, J.; Hamm, L.L.; Gu, D.; Shimmin, L.C.; Hixson, J.E.; Rao, D.C.; Sheng, S.; et al. Human Epithelial Na+ Channel Missense Variants Identified in the GenSalt Study Alter Channel Activity. Am. J. Physiol. Renal. Physiol. 2016, 311, F908–F914. [Google Scholar] [CrossRef]
- Blobner, B.M.; Kirabo, A.; Kashlan, O.B.; Sheng, S.; Arnett, D.K.; Becker, L.C.; Boerwinkle, E.; Carlson, J.C.; Gao, Y.; Gibbs, R.A.; et al. Rare Variants in Genes Encoding Subunits of the Epithelial Na+ Channel Are Associated With Blood Pressure and Kidney Function. Hypertension 2022, 79, 2573–2582. [Google Scholar] [CrossRef]
- Gu, X.; Gu, D.; He, J.; Rao, D.C.; Hixson, J.E.; Chen, J.; Li, J.; Huang, J.; Wu, X.; Rice, T.K.; et al. Resequencing Epithelial Sodium Channel Genes Identifies Rare Variants Associated With Blood Pressure Salt-Sensitivity: The GenSalt Study. Am. J. Hypertens. 2018, 31, 205–211. [Google Scholar] [CrossRef]
- Rotin, D.; Staub, O. Function and Regulation of the Epithelial Na+ Channel ENaC. Compr. Physiol. 2021, 11, 2017–2045. [Google Scholar] [CrossRef]
- Noreng, S.; Bharadwaj, A.; Posert, R.; Yoshioka, C.; Baconguis, I. Structure of the Human Epithelial Sodium Channel by Cryo-Electron Microscopy. eLife 2018, 7, e39340. [Google Scholar] [CrossRef] [PubMed]
- Noreng, S.; Posert, R.; Bharadwaj, A.; Houser, A.; Baconguis, I. Molecular Principles of Assembly, Activation, and Inhibition in Epithelial Sodium Channel. eLife 2020, 9, e59038. [Google Scholar] [CrossRef]
- Canessa, C.M.; Schild, L.; Buell, G.; Thorens, B.; Gautschi, I.; Horisberger, J.D.; Rossier, B.C. Amiloride-Sensitive Epithelial Na+ Channel Is Made of Three Homologous Subunits. Nature 1994, 367, 463–467. [Google Scholar] [CrossRef] [PubMed]
- Waldmann, R.; Champigny, G.; Bassilana, F.; Voilley, N.; Lazdunski, M. Molecular Cloning and Functional Expression of a Novel Amiloride-Sensitive Na+ Channel. J. Biol. Chem. 1995, 270, 27411–27414. [Google Scholar] [CrossRef] [PubMed]
- Haerteis, S.; Krueger, B.; Korbmacher, C.; Rauh, R. The δ-Subunit of the Epithelial Sodium Channel (ENaC) Enhances Channel Activity and Alters Proteolytic ENaC Activation. J. Biol. Chem. 2009, 284, 29024–29040. [Google Scholar] [CrossRef] [PubMed]
- Wichmann, L.; Vowinkel, K.S.; Perniss, A.; Manzini, I.; Althaus, M. Incorporation of the δ-Subunit into the Epithelial Sodium Channel (ENaC) Generates Protease-Resistant ENaCs in Xenopus Laevis. J. Biol. Chem. 2018, 293, 6647–6658. [Google Scholar] [CrossRef] [PubMed]
- Gettings, S.M.; Maxeiner, S.; Tzika, M.; Cobain, M.R.D.; Ruf, I.; Benseler, F.; Brose, N.; Krasteva-Christ, G.; Vande Velde, G.; Schönberger, M.; et al. Two Functional Epithelial Sodium Channel Isoforms Are Present in Rodents despite Pronounced Evolutionary Pseudogenization and Exon Fusion. Mol. Biol. Evol. 2021, 38, 5704–5725. [Google Scholar] [CrossRef] [PubMed]
- Kashlan, O.B.; Blobner, B.M.; Zuzek, Z.; Tolino, M.; Kleyman, T.R. Na+ Inhibits the Epithelial Na+ Channel by Binding to a Site in an Extracellular Acidic Cleft. J. Biol. Chem. 2015, 290, 568–576. [Google Scholar] [CrossRef]
- Kleyman, T.R.; Eaton, D.C. Regulating ENaC’s Gate. Am. J. Physiol. Cell Physiol. 2020, 318, C150–C162. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, W.; Larsen, E.H.; Lindemann, B. Current-Voltage Curve of Sodium Channels and Concentration Dependence of Sodium Permeability in Frog Skin. J. Physiol. 1977, 267, 137–166. [Google Scholar] [CrossRef]
- Leisgen, C.; Kuester, M.; Methfessel, C. The Roboocyte: Automated Electrophysiology Based on Xenopus Oocytes. Methods Mol. Biol. 2007, 403, 87–109. [Google Scholar] [CrossRef]
- Babini, E.; Geisler, H.-S.; Siba, M.; Gründer, S. A New Subunit of the Epithelial Na+ Channel Identifies Regions Involved in Na+ Self-Inhibition. J. Biol. Chem. 2003, 278, 28418–28426. [Google Scholar] [CrossRef]
- Ji, H.-L.; Su, X.-F.; Kedar, S.; Li, J.; Barbry, P.; Smith, P.R.; Matalon, S.; Benos, D.J. δ-Subunit Confers Novel Biophysical Features to Aβγ-Human Epithelial Sodium Channel (ENaC) via a Physical Interaction. J. Biol. Chem. 2006, 281, 8233–8241. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.; Carattino, M.D.; Kleyman, T.R. Role of the Wrist Domain in the Response of the Epithelial Sodium Channel to External Stimuli. J. Biol. Chem. 2012, 287, 44027–44035. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, M.P.; Friesner, R.A.; Xiang, Z.; Honig, B. On the Role of the Crystal Environment in Determining Protein Side-Chain Conformations. J. Mol. Biol. 2002, 320, 597–608. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, M.P.; Pincus, D.L.; Rapp, C.S.; Day, T.J.F.; Honig, B.; Shaw, D.E.; Friesner, R.A. A Hierarchical Approach to All-Atom Protein Loop Prediction. Proteins 2004, 55, 351–367. [Google Scholar] [CrossRef] [PubMed]
- Prime, 2017; Schrödinger, L.L.C.: New York, NY, USA, 2021.
- Schrödinger, L.L.C. MacroModel|Schrödinger. MacroModel. Schrödinger Release 2018-1 (2018); Schrödinger, Inc.: New York, NY, USA, 2021; Available online: https://www.schrodinger.com/products/macromodel (accessed on 30 March 2023).
- Lu, C.; Wu, C.; Ghoreishi, D.; Chen, W.; Wang, L.; Damm, W.; Ross, G.A.; Dahlgren, M.K.; Russell, E.; Von Bargen, C.D.; et al. OPLS4: Improving Force Field Accuracy on Challenging Regimes of Chemical Space. J. Chem. Theory Comput. 2021, 17, 4291–4300. [Google Scholar] [CrossRef] [PubMed]
- Salih, M.; Gautschi, I.; Van Bemmelen, M.X.; Di Benedetto, M.; Brooks, A.S.; Lugtenberg, D.; Schild, L.; Hoorn, E.J. A Missense Mutation in the Extracellular Domain of Aenac Causes Liddle Syndrome. J. Am. Soc. Nephrol. 2017, 28, 3291–3299. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Chen, J.; Shi, S.; Sheng, S.; Kleyman, T.R. Analyses of Epithelial Na+ Channel Variants Reveal That an Extracellular β-Ball Domain Critically Regulates ENaC Gating. J. Biol. Chem. 2019, 294, 16765–16775. [Google Scholar] [CrossRef]
- Wichmann, L.; Dulai, J.S.; Marles-Wright, J.; Maxeiner, S.; Szczesniak, P.P.; Manzini, I.; Althaus, M. An Extracellular Acidic Cleft Confers Profound H+-Sensitivity to Epithelial Sodium Channels Containing the δ-Subunit in Xenopus Laevis. J. Biol. Chem. 2019, 294, 12507–12520. [Google Scholar] [CrossRef]
- Collier, D.M.; Tomkovicz, V.R.; Peterson, Z.J.; Benson, C.J.; Snyder, P.M. Intersubunit Conformational Changes Mediate Epithelial Sodium Channel Gating. J. Gen. Physiol. 2014, 144, 337–348. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, X.; Chen, J.; Sheng, S.; Kleyman, T.R. Extracellular Intersubunit Interactions Modulate Epithelial Na+ Channel Gating. J. Biol. Chem. 2023, 299, 102914. [Google Scholar] [CrossRef]
- Sheng, S.; Chen, J.; Mukherjee, A.; Yates, M.E.; Buck, T.M.; Brodsky, J.L.; Tolino, M.A.; Hughey, R.P.; Kleyman, T.R. Thumb Domains of the Three Epithelial Na+ Channel Subunits Have Distinct Functions. J. Biol. Chem. 2018, 293, 17582–17592. [Google Scholar] [CrossRef] [PubMed]
- Yoder, N.; Yoshioka, C.; Gouaux, E. Gating Mechanisms of Acid-Sensing Ion Channels. Nature 2018, 555, 397–401. [Google Scholar] [CrossRef] [PubMed]
- Chraïbi, A.; Horisberger, J.D. Na Self Inhibition of Human Epithelial Na Channel: Temperature Dependence and Effect of Extracellular Proteases. J. Gen. Physiol. 2002, 120, 133–145. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lawong, R.Y.; May, F.; Etang, E.C.; Vorrat, P.; George, J.; Weder, J.; Kockler, D.; Preller, M.; Althaus, M. Recording Sodium Self-Inhibition of Epithelial Sodium Channels Using Automated Electrophysiology in Xenopus Oocytes. Membranes 2023, 13, 529. https://doi.org/10.3390/membranes13050529
Lawong RY, May F, Etang EC, Vorrat P, George J, Weder J, Kockler D, Preller M, Althaus M. Recording Sodium Self-Inhibition of Epithelial Sodium Channels Using Automated Electrophysiology in Xenopus Oocytes. Membranes. 2023; 13(5):529. https://doi.org/10.3390/membranes13050529
Chicago/Turabian StyleLawong, Rene Y., Fabian May, Etang C. Etang, Philipp Vorrat, Jonas George, Julia Weder, Dagmar Kockler, Matthias Preller, and Mike Althaus. 2023. "Recording Sodium Self-Inhibition of Epithelial Sodium Channels Using Automated Electrophysiology in Xenopus Oocytes" Membranes 13, no. 5: 529. https://doi.org/10.3390/membranes13050529