Proteomics Analysis of Gastric Cancer Patients with Diabetes Mellitus

 , , , ,
, , , ,  , and
, and
Abstract
:1. Introduction
2. Experimental Section
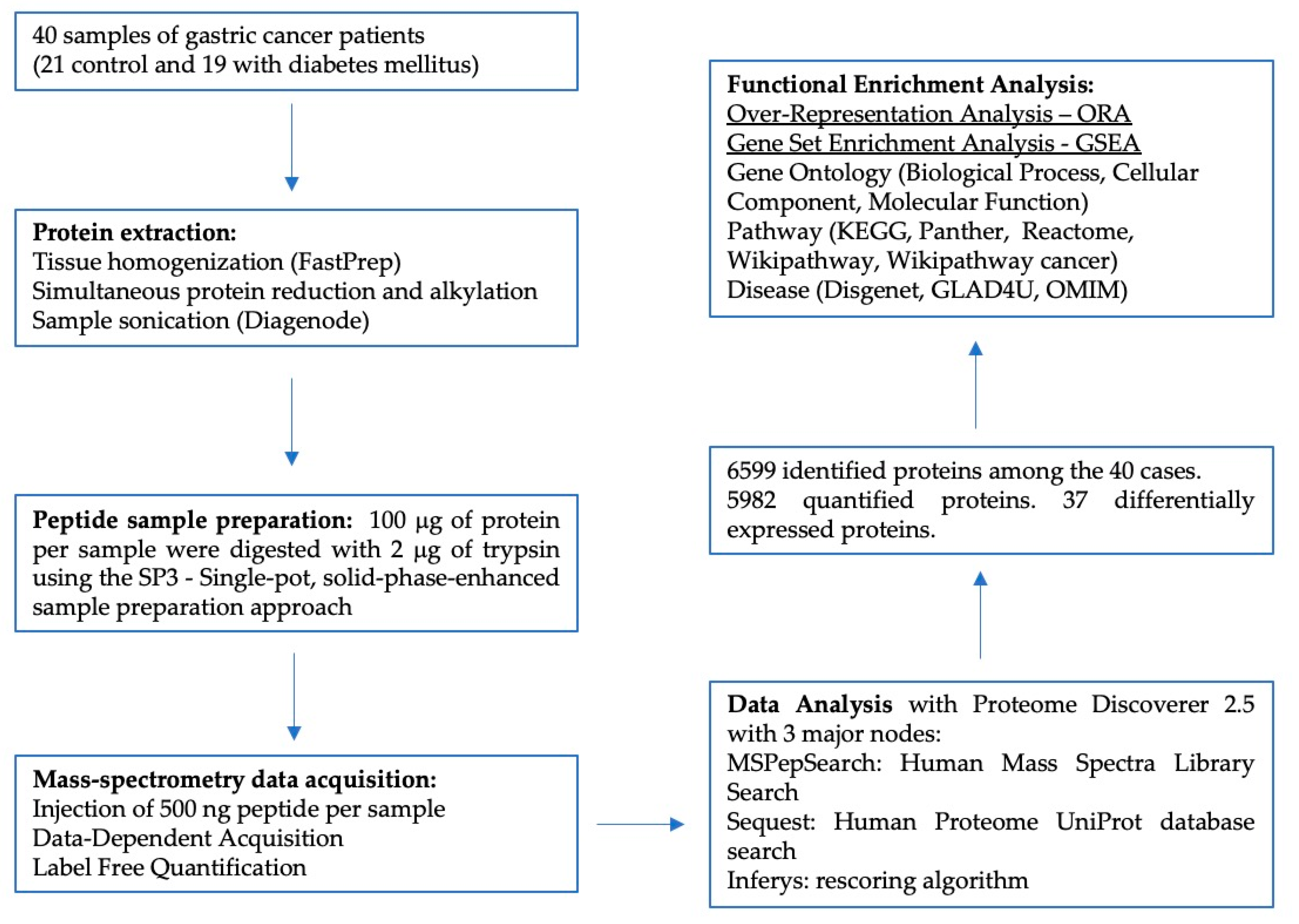
2.1. Sample Selection
2.2. Protein Extraction
2.3. Proteomics Data Acquisition
2.4. Data Analysis
2.5. Protein Functional Enrichment Analysis
3. Results
3.1. Clinical Data
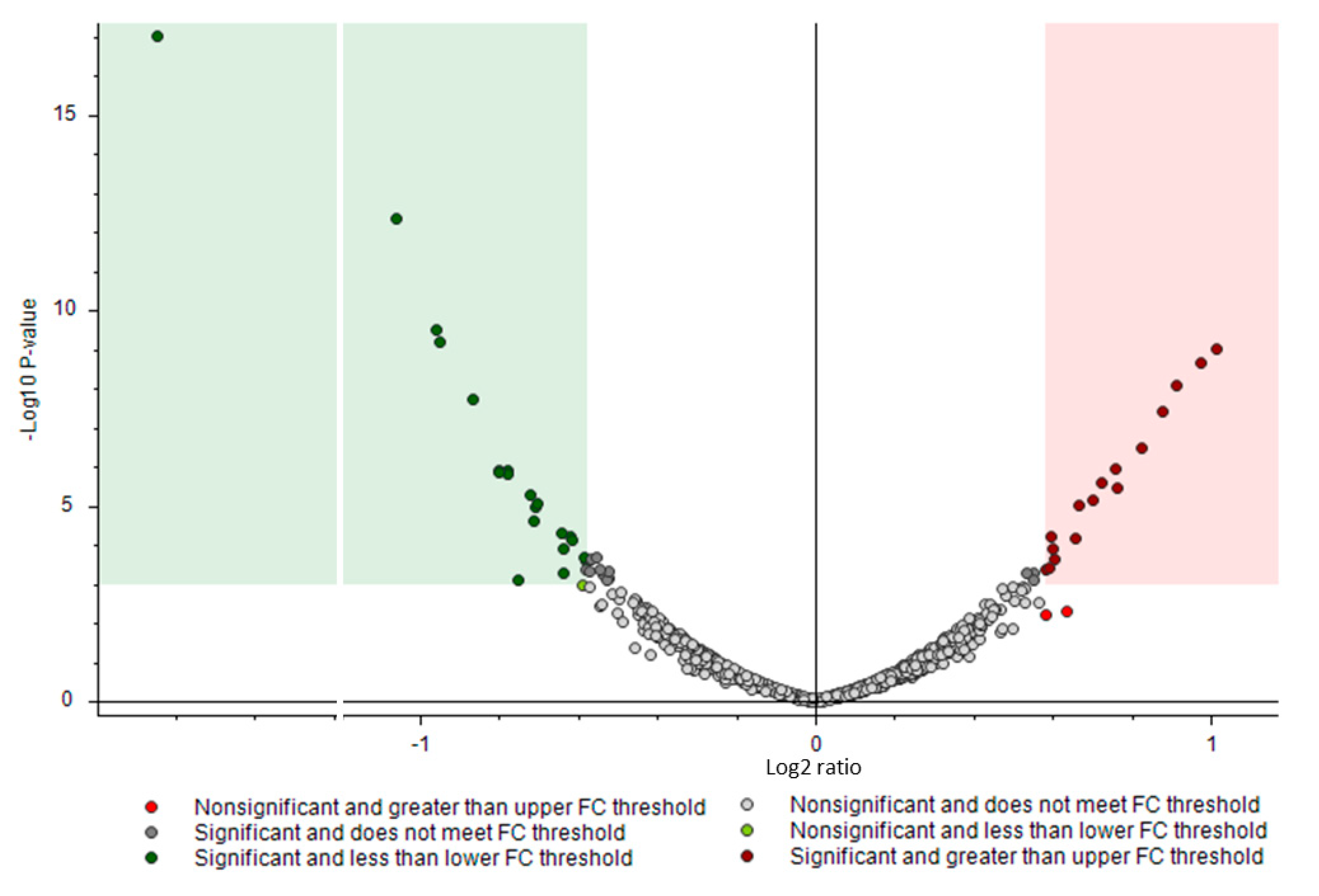
3.2. Proteomics Analysis
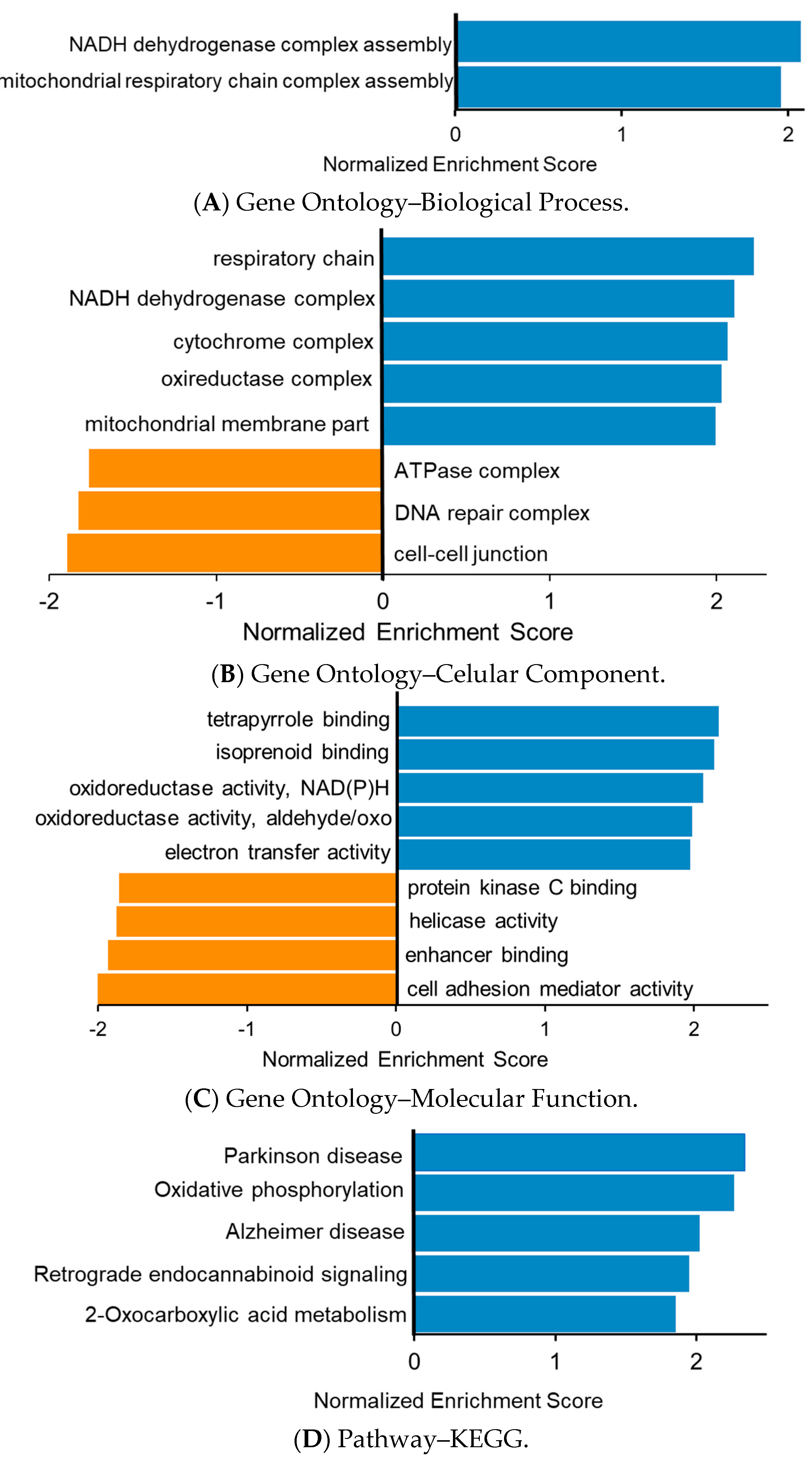
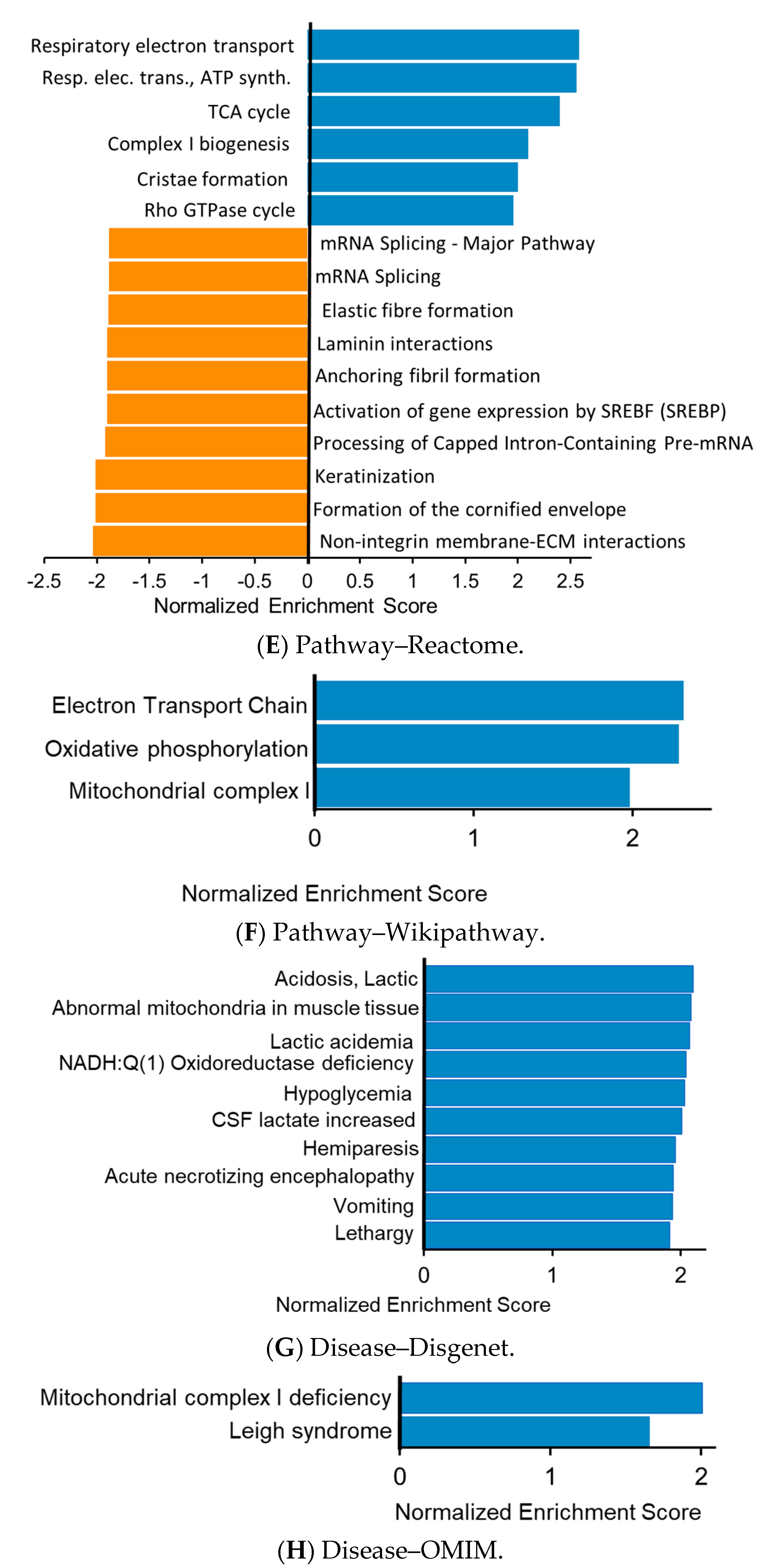
3.3. Functional Enrichment Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| GC | Gastric Cancer |
| DM | Diabetes mellitus |
| MS | Mass spectrometry |
| ORA | Over-Representation Analysis |
| GSEA | Gene Set Enrichment Analysis |
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA A Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karimi, P.; Islami, F.; Anandasabapathy, S.; Freedman, N.D.; Kamangar, F. Gastric Cancer: Descriptive Epidemiology, Risk Factors, Screening, and Prevention. Cancer Epidemiol. Biomark. Prev. 2014, 23, 700–713. [Google Scholar] [CrossRef] [Green Version]
- Guggenheim, D.E.; Shah, M.A. Gastric cancer epidemiology and risk factors. J. Surg. Oncol. 2013, 107, 230–236. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, J.; Oliveira, P.; Senz, J.; José, C.S.; Hansford, S.; Teles, S.P.; Ferreira, M.; Corso, G.; Pinheiro, H.; Lemos, D.; et al. Redefinition of familial intestinal gastric cancer: Clinical and genetic perspectives. J. Med. Genet. 2020, 58, 1–11. [Google Scholar] [CrossRef]
- Giovannucci, E.L.; Harlan, D.M.; Archer, M.C.; Bergenstal, R.M.; Gapstur, S.M.; Habel, L.A.; Pollak, M.; Regensteiner, J.G.; Yee, D. Diabetes and Cancer: A consensus report. Diabetes Care 2010, 33, 1674–1685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimoyama, S. Diabetes mellitus carries a risk of gastric cancer: A meta-analysis. World J. Gastroenterol. 2013, 19, 6902–6910. [Google Scholar] [CrossRef]
- Ge, Z.; Ben, Q.; Qian, J.; Wang, Y.; Li, Y. Diabetes mellitus and risk of gastric cancer. Eur. J. Gastroenterol. Hepatol. 2011, 23, 1127–1135. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.-W.; Freedman, N.D.; Hollenbeck, A.R.; Schatzkin, A.; Abnet, C.C. Prospective Study of Self-Reported Diabetes and Risk of Upper Gastrointestinal Cancers. Cancer Epidemiol. Biomark. Prev. 2011, 20, 954–961. [Google Scholar] [CrossRef] [Green Version]
- Cheung, K.S.; Chan, E.W.; Chen, L.; Seto, W.K.; Wong, I.C.; Leung, W.K. Diabetes Increases Risk of Gastric Cancer AfterHelicobacter pyloriEradication: A Territory-Wide Study With Propensity Score Analysis. Diabetes Care 2019, 42, 1769–1775. [Google Scholar] [CrossRef] [PubMed]
- Mansori, K.; Moradi, Y.; Naderpour, S.; Rashti, R.; Moghaddam, A.B.; Saed, L.; Mohammadi, H. Helicobacter pylori infection as a risk factor for diabetes: A meta-analysis of case-control studies. BMC Gastroenterol. 2020, 20, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Tseng, C.-H.; Tseng, F.-H. Diabetes and gastric cancer: The potential links. World J. Gastroenterol. 2014, 20, 1701–1711. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Rutegård, M.; Santoni, G.; Wallner, B.; Johansson, I.; Sund, M.; Xie, S.-H.; Lagergren, J. Prediabetes and diabetes in relation to risk of gastric adenocarcinoma. Br. J. Cancer 2019, 120, 1147–1152. [Google Scholar] [CrossRef] [PubMed]
- Rostami-Nejad, M.; Rezaei-Tavirani, M.; Mansouri, V.; Akbari, Z.; Abdi, S. Impact of proteomics investigations on gastric cancer treatment and diagnosis. Gastroenterol. Hepatol. Bed Bench 2019, 12, S1–S7. [Google Scholar] [CrossRef] [PubMed]
- Del Puerto-Nevado, L.; Minguez, P.; Corton, M.; Solanes-Casado, S.; Prieto, I.; Mas, S.; Sanz, A.B.; González-Alonso, P.; Villaverde, C.; Portal-Núñez, S.; et al. Molecular evidence of field cancerization initiated by diabetes in colon cancer patients. Mol. Oncol. 2019, 13, 857–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, C.S.; Moggridge, S.; Müller, T.; Sorensen, P.H.; Morin, G.B.; Krijgsveld, J. Single-pot, solid-phase-enhanced sample preparation for proteomics experiments. Nat. Protoc. 2019, 14, 68–85. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Wang, J.; Jaehnig, E.J.; Shi, Z.; Zhang, B. WebGestalt 2019: Gene set analysis toolkit with revamped UIs and APIs. Nucleic Acids Res. 2019, 47, W199–W205. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.-Y.; Baffy, G.; Perret, P.; Krauss, S.; Peroni, O.; Grujic, D.; Hagen, T.; Vidal-Puig, A.J.; Boss, O.; Kim, Y.-B.; et al. Uncoupling Protein-2 Negatively Regulates Insulin Secretion and Is a Major Link between Obesity, β Cell Dysfunction, and Type 2 Diabetes. Cell 2001, 105, 745–755. [Google Scholar] [CrossRef] [Green Version]
- Peng, F.; Wu, D.; Gao, B.; Ingram, A.J.; Zhang, B.; Chorneyko, K.; McKenzie, R.; Krepinsky, J.C. RhoA/Rho-Kinase Contribute to the Pathogenesis of Diabetic Renal Disease. Diabetes 2008, 57, 1683–1692. [Google Scholar] [CrossRef] [Green Version]
- Rivelli, J.F.; Amaiden, M.R.; Monesterolo, N.E.; Previtali, G.; Santander, V.S.; Fernandez, A.; Arce, C.A.; Casale, C.H. High glucose levels induce inhibition of Na,K-ATPase via stimulation of aldose reductase, formation of microtubules and formation of an acetylated tubulin/Na,K-ATPase complex. Int. J. Biochem. Cell Biol. 2012, 44, 1203–1213. [Google Scholar] [CrossRef]
- Sivitz, W.I.; Yorek, M.A. Mitochondrial Dysfunction in Diabetes: From Molecular Mechanisms to Functional Significance and Therapeutic Opportunities. Antioxid. Redox Signal. 2010, 12, 537–577. [Google Scholar] [CrossRef] [Green Version]
- Ruegsegger, G.N.; Creo, A.L.; Cortes, T.M.; Dasari, S.; Nair, K.S. Altered mitochondrial function in insulin-deficient and insulin-resistant states. J. Clin. Investig. 2018, 128, 3671–3681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crescenzo, R.; Bianco, F.; Mazzoli, A.; Giacco, A.; Liverini, G.; Iossa, S. Mitochondrial efficiency and insulin resistance. Front. Physiol. 2015, 6, 512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belosludtsev, K.N.; Belosludtseva, N.V.; Dubinin, M.V. Diabetes Mellitus, Mitochondrial Dysfunction and Ca2+-Dependent Permeability Transition Pore. Int. J. Mol. Sci. 2020, 21, 6559. [Google Scholar] [CrossRef] [PubMed]
- Sergi, D.; Renaud, J.; Simola, N.; Martinoli, M.-G. Diabetes, a Contemporary Risk for Parkinson’s Disease: Epidemiological and Cellular Evidences. Front. Aging Neurosci. 2019, 11, 302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.J.; Seo, H.I.; Cha, H.Y.; Yang, Y.J.; Kwon, S.H.; Yang, S.J. Diabetes and Alzheimer’s Disease: Mechanisms and Nutritional Aspects. Clin. Nutr. Res. 2018, 7, 229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okugawa, T.; Oshima, T.; Chen, X.; Hori, K.; Tomita, T.; Fukui, H.; Watari, J.; Matsumoto, T.; Miwa, H. Down-Regulation of Claudin-3 Is Associated with Proliferative Potential in Early Gastric Cancers. Dig. Dis. Sci. 2012, 57, 1562–1567. [Google Scholar] [CrossRef] [PubMed]
- Fristedt, R.; Gaber, A.; Hedner, C.; Nodin, B.; Uhlen, M.; Eberhard, J.; Jirström, K. Expression and prognostic significance of the polymeric immunoglobulin receptor in esophageal and gastric adenocarcinoma. J. Transl. Med. 2014, 12, 83. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Yang, H.-Q.; Xi, H.-L.; Feng, S.; Qin, R.-H. Inhibition of CDH17 gene expression via RNA interference reduces proliferation and apoptosis of human MKN28 gastric cancer cells. Int. J. Oncol. 2017, 50, 15–22. [Google Scholar] [CrossRef] [Green Version]
- Azizi, S.M.; Hashemi, M.; Falsafi, S.; Azizi, S.M.; Shirkoohi, R. Expression of Villin associated with Epithelial to Mesenchymal Transition in patients with gastric cancer relating to their clinical and morphological specifications. Asian Pac. J. Cancer Biol. 2020, 5, 11–14. [Google Scholar] [CrossRef]
- Szondy, Z.; Korponay-Szabó, I.; Király, R.; Sarang, Z.; Tsay, G.J. Transglutaminase 2 in human diseases. BioMedicine 2017, 7, 15. [Google Scholar] [CrossRef] [Green Version]
- Cho, S.-Y.; Oh, Y.; Jeong, E.M.; Park, S.; Lee, D.; Wang, X.; Zeng, Q.; Qin, H.; Hu, F.; Gong, H.; et al. Amplification of transglutaminase 2 enhances tumor-promoting inflammation in gastric cancers. Exp. Mol. Med. 2020, 52, 854–864. [Google Scholar] [CrossRef] [PubMed]
- Shimamura, T.; Ito, H.; Shibahara, J.; Watanabe, A.; Hippo, Y.; Taniguchi, H.; Chen, Y.; Kashima, T.; Ohtomo, T.; Tanioka, F.; et al. Overexpression of MUC13 is associated with intestinal-type gastric cancer. Cancer Sci. 2005, 96, 265–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biedermann, K.; Vogelsang, H.; Becker, I.; Plaschke, S.; Siewert, J.R.; Höfler, H.; Keller, G. Desmoglein 2 is expressed abnormally rather than mutated in familial and sporadic gastric cancer. J. Pathol. 2005, 207, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.C.; Chan, J.C.N. Evidence for DNA Damage as a Biological Link between Diabetes and Cancer. Chin. Med. J. 2015, 128, 1543–1548. [Google Scholar] [CrossRef]
- Kyuno, D.; Takasawa, A.; Kikuchi, S.; Takemasa, I.; Osanai, M.; Kojima, T. Role of tight junctions in the epithelial-to-mesenchymal transition of cancer cells. Biochim. Biophys. Acta Biomembr. 2020, 183503. [Google Scholar] [CrossRef]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Non-DM Patients | DM Patients | |
|---|---|---|
| Patients (N) | 21 | 19 |
| Gender (male: female) | 7:14 | 7:12 |
| Median age (range) | 75 (57–85) | 75 (58–87) |
| Laurén classification (n—%) | ||
| Intestinal | 13 (61.9) | 10 (52.6) |
| Diffuse | 1 (4.8) | 1 (5.3) |
| Mixed | 7 (33.3) | 6 (31.6) |
| Indeterminate | 0 (0) | 2 (10.5) |
| pT category (N—%) | ||
| pT1a, pT1b | 1 (4.8) | 2 (10.5) |
| pT2 | 2 (9.5) | 1 (5.3) |
| pT3 | 13 (61.9) | 12 (63.1) |
| pT4a, pT4b | 5 (23.8) | 4 (21.1) |
| pN category (N—%) | ||
| pN0 | 6 (28.6) | 9 (47.4) |
| pN ≥ 1 | 15 (71.4) | 10 (52.6) |
| M category (N—%) | ||
| M0 | 20 (95) | 16 (84.2) |
| (p)M1 | 1 (4.8) | 3 (15.8) |
| Protein Name | Accession No. | Gene Symbol | Fold-Change (DM/C) | Unique Peptides |
|---|---|---|---|---|
| Immunoglobulin lambda variable 3–27 | P01718 | IGLV3-27 | 2.023 | 4 |
| Desmin | P17661 | DES | 1.968 | 42 |
| Gastricsin | P20142 | PGC | 1.885 | 3 |
| Protein S100-P | P25815 | S100P | 1.837 | 5 |
| Galectin-10 | Q05315 | CLC | 1.771 | 7 |
| Aldehyde dehydrogenase, dimeric NADP-preferring | P30838 | ALDH3A1 | 1.701 | 21 |
| Caveolae-associated protein 3 | E9PIE3 | CAVIN3 | 1.691 | 8 |
| Gastrokine-2 | Q86XP6 | GKN2 | 1.654 | 6 |
| Eosinophil peroxidase | P11678 | EPX | 1.629 | 15 |
| Marginal zone B- and B1-cell-specific protein | Q8WU39 | MZB1 | 1.586 | 11 |
| Annexin A10 | Q9UJ72 | ANXA10 | 1.578 | 16 |
| Protein S100-A8 | P05109 | S100A8 | 1.523 | 12 |
| Immunoglobulin kappa variable 4-1 | P06312 | IGKV4-1 | 1.517 | 2 |
| Carboxymethylenebutenolidase homolog | Q96DG6 | CMBL | 1.510 | 10 |
| High mobility group protein HMG-I/HMG-Y | P17096 | HMGA1 | 1.507 | 4 |
| Neutrophil collagenase | P22894 | MMP8 | 1.500 | 11 |
| Protein-glutamine gamma-glutamyltransferase 2 | P21980 | TGM2 | 0.667 | 32 |
| Type-1 angiotensin II receptor-associated protein | Q6RW13 | AGTRAP | 0.664 | 2 |
| Glycerol-3-phosphate dehydrogenase [NAD(+)], cytoplasm. | P21695 | GPD1 | 0.654 | 14 |
| Transcription factor BTF3 | P20290 | BTF3 | 0.651 | 7 |
| Monoglyceride lipase | Q99685 | MGLL | 0.643 | 7 |
| Metal cation symporter ZIP14 | Q15043 | SLC39A14 | 0.642 | 4 |
| Desmoglein-2 | Q14126 | DSG2 | 0.642 | 28 |
| Polymeric immunoglobulin receptor | P01833 | PIGR | 0.615 | 54 |
| Fatty acid-binding protein, liver | P07148 | FABP1 | 0.613 | 9 |
| Claudin-3 | O15551 | CLDN3 | 0.610 | 4 |
| Myosin-14 | Q7Z406 | MYH14 | 0.606 | 54 |
| Complement C1q subcomponent subunit A | P02745 | C1QA | 0.595 | 3 |
| Cellular nucleic acid-binding protein | P62633 | CNBP | 0.584 | 5 |
| Villin-1 | P09327 | VIL1 | 0.584 | 35 |
| Protein POF1B | Q8WVV4 | POF1B | 0.575 | 19 |
| CD59 glycoprotein (Fragment) | A0A2U3TZL5 | CD59 | 0.574 | 4 |
| Estradiol 17-beta-dehydrogenase 2 | P37059 | HSD17B2 | 0.549 | 12 |
| Cadherin-17 | Q12864 | CDH17 | 0.518 | 21 |
| Mucin-13 | Q9H3R2 | MUC13 | 0.515 | 6 |
| Carcinoembryonic antigen-related cell adhesion molecule 5 | A0A024R0K5 | CEACAM5 | 0.480 | 8 |
| YTH domain-containing family protein 1 | Q9BYJ9 | YTHDF1 | 0.010 | 2 |
| DEP | Functional Database | GENE Set | Description | Protein Name |
|---|---|---|---|---|
| + | GO—CC | GO:0005576 | Extracellular region | S100 calcium binding protein A8 eosinophil peroxidase desmin progastricsin matrix metallopeptidase 8 S100 calcium binding protein P aldehyde dehydrogenase 3 family member A1 Galectin-10 gastrokine 2 marginal zone B and B1 cell specific protein carboxymethylenebutenolidase homolog |
| − | Disease—GLAD4U | PA444632 | Intestinal Diseases | transglutaminase 2 polymeric immunoglobulin receptor claudin 3 villin 1 cadherin 17 mucin 13, cell surface associated |
| − | Disease—GLAD4U | PA447242 | Epithelial Cancers | transglutaminase 2 desmoglein 2 polymeric immunoglobulin receptor claudin 3 villin 1 mucin 13, cell surface associated |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Osório, H.; Silva, C.; Ferreira, M.; Gullo, I.; Máximo, V.; Barros, R.; Mendonça, F.; Oliveira, C.; Carneiro, F. Proteomics Analysis of Gastric Cancer Patients with Diabetes Mellitus. J. Clin. Med. 2021, 10, 407. https://doi.org/10.3390/jcm10030407
Osório H, Silva C, Ferreira M, Gullo I, Máximo V, Barros R, Mendonça F, Oliveira C, Carneiro F. Proteomics Analysis of Gastric Cancer Patients with Diabetes Mellitus. Journal of Clinical Medicine. 2021; 10(3):407. https://doi.org/10.3390/jcm10030407
Chicago/Turabian StyleOsório, Hugo, Cátia Silva, Marta Ferreira, Irene Gullo, Valdemar Máximo, Rita Barros, Fernando Mendonça, Carla Oliveira, and Fátima Carneiro. 2021. "Proteomics Analysis of Gastric Cancer Patients with Diabetes Mellitus" Journal of Clinical Medicine 10, no. 3: 407. https://doi.org/10.3390/jcm10030407