
L-Carnitine and Male Fertility: Is Supplementation Beneficial?

 ,
,  ,
,  ,
,  and
and 
Abstract
:
1. Introduction
2. L-Carnitine Is Commonly Used as a Supplement for Professional Athletes: Is It Safe?
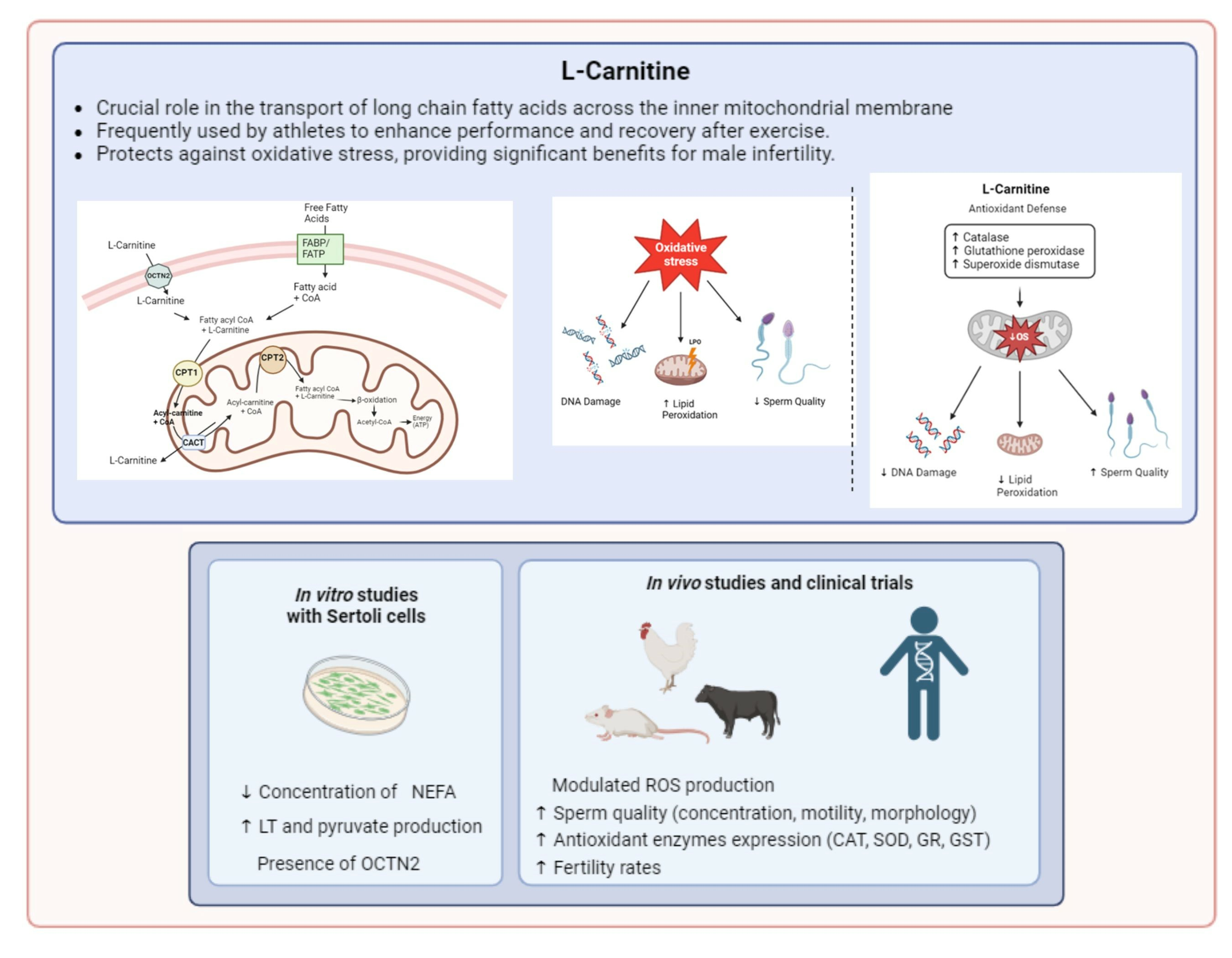
3. Molecular Mechanisms of L-Carnitine Function
4. Signaling Pathways Affected by L-Carnitine
5. L-Carnitine in Male Fertility
5.1. In Vitro Studies
5.2. In Vivo Studies

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Aims of the Study | Period, Dosing, Route of Administration | Model | Main Results | References |
|---|---|---|---|---|
| Examine whether the detrimental effects of long-term copper consumption on sperm quality and testis function of Wistar albino rats could be prevented by LC therapy | 30 days; 200 mg/kg CuSO4 50 and 100 mg/kg LC; oral administration | 3-month-old Wistar rats | Revitalization of sperm quality (motility, viability, number); Restoration of histological alterations (germ cell depletion, sloughing of germ cells, vacuolization, and degeneration); Rejuvenation of spermatogenesis. | M. Khushboo et al., 2017 [93] |
| Study the protective effect of LC on Sertoli testis cells from the damage of chemotherapy | 5 days; 100 mg/kg LC intraperitoneally administration | Testis Sertoli cells from adult male mice | Recovery of the sperm count and sperm motility; Higher expressions of occluding and GDNF; Lower expression of TGF-β3. | Y. Cao et al., 2017 [38] |
| Evaluate the effect of carnitine supplementation of semen extender on fertility parameters of frozen-thawed buffalo sperm | 2.5 and 7.5 mM carnitine | Cryopreserved Buffalo semen | Increased ATP generation; Modulated ROS production. | V. Longobardi et al., 2017 [96] |
| Evaluated whether some spermatic qualitative parameters could be ameliorated by carnitine treatment in adult rats exposed to doxorubicin | A single dose of LC (250 mg/kg body weight) | Pre-pubertal male Wistar rats | Increase in intact acrosome integrity; Decreased MDA and nitrite concentration; Increased fertility and implantation rate; Decreased spermatozoa with damaged DNA. | R. Cabral et al., 2017 [35] |
| Investigate the protective effect of LC and L-arginine on semen quality, OS parameters, and testis cell energy after busulfan treatment | single I.P. injection of busulfan; 1 mL of L-arginine daily by oral gavage; 1 mL of LC by oral gavages | Adult male Wistar rats | Improved sperm morphology, motility, velocity, and count; Increased MDA and ATP. | Abd-Elrazek et al., 2017 [98] |
| Investigate the effects of dietary LC on semen quality, seminal antioxidant activity, and their implications for fertility in aged roosters | 12 weeks; 50 and 150 mg/kg body weight/day of LC | Aged roosters | Increased sperm quality; Increased plasma concentration of testosterone; Increased seminal MDA concentration, CAT, and GSH-PX activity. | Elokil et al., 2019 [37] |
| Investigate the possible protective role of Se and LC against the adverse effects induced by cadmium | 30 days; LC at a dose of 10 mg/kg | Mature adult male albino mice | Increased CAT, GR, SOD, and GST activities; Less histopathological abnormalities; Less DNA damage. | Alharthi et al., 2020 [33] |
| Investigate the possible effect of LC on the mechanisms of reproductive toxicity induced by chronic lead acetate treatment | 40 days by oral gavage; 50 mg/kg of lead acetate; 100 mg/kg of LC | Male Wistar rats | Reduced testicular OS; Improved sperm parameters; Elevated serum FSH, LH, and testosterone. | Abdel-Emam et al., 2021 [95] |
| Evaluate the underlying mechanism of the ameliorative effects of LC | 30 days; diet supplemented with 1.5% cholesterol + LC 150 mg/kg given orally | Premature albino male rats | Histological protection for spermatogenic layers; Better concentration of sperm; Reduced sperm abnormalities. | Karam et al., 2022 [94] |
5.3. Clinical Studies
| Aims of the Study | Period, Dosing, Route of Administration | Population | Main Results | References |
|---|---|---|---|---|
| Determine the effects of LC on sperm parameters in patients with idiopathic sperm abnormalities | 3 months; 1 g of LC orally; 3 times a day | 48 smokers and 122 non-smokers (Aged 20–56 years) | Increased sperm motility; Increased normal-shaped spermatozoa. | Khademi et al., 2005 [99] |
| Explore the clinical effect of LC on infertile males with asthenozoospermia | 3 months; 2 g/day of LC | 135 patients with asthenozoospermia | Increased sperm motility; Raised rate of pregnancy. | Wang et al., 2010 [40] |
| Determine the efficacy of combined LC and ALC therapy in infertile males with oligoasthenoteratozoospermia | 2 months’ wash out, 6 months’ therapy, 2 month follow-up; 10 mL vial containing 2 g/d carnitine orally administered; 500 mg twice per day orally every 12 h of ALC; | 60 infertile men with oligoasthenoteratozoospermia (Aged 20–40 years) | Increased sperm motility. | Lenzi et al., 2004 [28] |
| Investigate the effect of supplementation with LC and ALC on sperm quality in 104 subjects with oligo- and/or astheno- and/or teratozoospermia with or without varicocele. | 6 months; two sachets daily of a supplement containing 1000 mg LC, 500 mg ALC, and other micronutrients | 104 men with oligo and/or astheno and/or teratozoospermia with or without varicocele (Aged 18–48 years) | Increased sperm concentration; Increased total sperm count; Increased total motility. | Busetto et al., 2018 [102] |
| Evaluate the effect of LC on low sperm acrosin activity in infertile men | 3 months; 1 g of LC | 240 male infertility patients with low sperm acrosin activity | Increased sperm motility; Increased sperm acrosin activity; Increased sperm concentration. | Sun et al., 2018 [100] |
| Study the effects of complex ALC, LC, fumarate, and alpha-lipoic acid on oxidative stress, ejaculate quality, and sperm DNA fragmentation in men with infertility. | 6 months; 10 g of the supplement once a day | 80 infertile men with increased levels of sperm DNA fragmentation and oxidative stress (Aged 25–40 years) | Improved sperm mobility and morphology; Decreased free oxygen radicals: Decrease in DNA fragmentation. | Gamidov et al., 2019 [23] |
| Evaluate a combined treatment with LC and acetyl-L-Carnitine with micronutrients as a treatment for infertility in men | 6 months; 1.0 g LC, 0.5 g ALC and other micronutrients | 175 males with idiopathic oligoasthenozoospermia (Aged 19–44 years) | Decreased DNA fragmentation; Increased seminal carnitine concentration; Improved sperm motility. | Micic et al., 2018 [107] |
| Comparing sperm parameters and hormonal levels with LC vs. CoQ10 and vitamin E therapy for patients with asthenozoospermia and teratozoospermia. | 3 months; 15 g/bag LC complex; orally, one bag at a time, twice a day | 143 patients with asthenozoospermia and teratozoospermia. | Increased sperm motility, morphology, and concentration; Improved testosterone levels; Superior to the others in improving sperm quality. | Ma et al., 2020 [39] |
| Study the influence of a multi-component nutrient dietary supplement on sperm parameters and pregnancy rates in idiopathic male infertility | 6 months; supplement containing LC/ALC, l-arginine, glutathione, co-enzyme Q10, zinc, vitamin B9, vitamin B12, selenium, once daily | 83 males aged 21–50 years with oligo-, astheno-, and teratozoospermia (Aged 21–50 years) | Improved sperm quality (concentration, motility, and morphology); Increased fertility rates. | Kopets et al., 2020 [41] |
| Assess the effect of LC and CoQ10 during, before, and after freezing in oligospermia men | 1 h to 2 weeks; G1—LC (100 µM) for 1 h; G3—CoQ10 (100 µM) for 1 h; G4—LC and CoQ10 (100 µM) for 1 h; G5—freezing control group; G6—frozen with CoQ10 (100 µM); G7—frozen with LC (100 µM); G8—frozen with 100 µM (LC + CoQ10) for 2 weeks. | 30 oligospermic men (Aged 25–40 years) | Decreased ROS levels; Improved motility and protamine deficiency. | Nezhad et al., 2021 [101] |
6. Materials and Methods
7. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Infertility. Available online: https://www.who.int/news-room/fact-sheets/detail/infertility (accessed on 9 December 2022).
- Babakhanzadeh, E.; Nazari, M.; Ghasemifar, S.; Khodadadian, A. Some of the Factors Involved in Male Infertility: A Prospective Review. Int. J. Gen. Med. 2020, 13, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Chua, M.E.; Escusa, K.G.; Luna, S.; Tapia, L.C.; Dofitas, B.; Morales, M. Revisiting oestrogen antagonists (clomiphene or tamoxifen) as medical empiric therapy for idiopathic male infertility: A meta-analysis. Andrology 2013, 1, 749–757. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Minhas, S.; Dhillo, W.S.; Jayasena, C.N. Male infertility due to testicular disorders. J. Clin. Endocrinol. Metab. 2021, 106, e442–e459. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, S.F. Spermatogenesis. In Developmental Biology, 6th ed.; Sinauer Associates: Sunderland, MA, USA, 2000. Available online: https://www.ncbi.nlm.nih.gov/books/NBK10095/ (accessed on 7 July 2023).
- Cole, L.A. (Ed.) Human Male Spermatogenesis in Encyclopedia of Reproduction, 2nd ed.; Chapter 18; Biology of Life; Academic Press: Cambridge, MA, USA, 2018; pp. 135–141. [Google Scholar] [CrossRef]
- Rato, L.; Alves, M.G.; Socorro, S.; Duarte, A.I.; Cavaco, J.E.; Oliveira, P.F. Metabolic regulation is important for spermatogenesis. Nat. Rev. Urol. 2012, 9, 330–338. [Google Scholar] [CrossRef]
- Abou-Haila, A.; Tulsiani, D.R. Mammalian Sperm Acrosome: Formation, Contents, and Function. Arch. Biochem. Biophys. 2000, 379, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.Y.; Mruk, D.D. The biology of spermatogenesis: The past, present and future. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 1459–1463. [Google Scholar] [CrossRef]
- Tremellen, K. Oxidative stress and male infertility—A clinical perspective. Hum. Reprod. Updat. 2008, 14, 243–258. [Google Scholar] [CrossRef] [PubMed]
- Martin-Hidalgo, D.; Bragado, M.J.; Batista, A.R.; Oliveira, P.F.; Alves, M.G. Antioxidants and Male Fertility: From Molecular Studies to Clinical Evidence. Antioxidants 2019, 8, 89. [Google Scholar] [CrossRef]
- Wright, C.; Milne, S.; Leeson, H. Sperm DNA damage caused by oxidative stress: Modifiable clinical, lifestyle and nutritional factors in male infertility. Reprod. Biomed. Online 2014, 28, 684–703. [Google Scholar] [CrossRef]
- Smith, M.A.; Perry, G.; Pryor, W.A. Causes and consequences of oxidative stress in Alzheimer’s disease. Free Radic. Biol. Med. 2002, 32, 1049. [Google Scholar] [CrossRef]
- Halliwell, B. Antioxidants in human health and disease. Annu. Rev. Nutr. 1996, 16, 33–50. [Google Scholar] [CrossRef]
- Forman, H.J.; Ursini, F.; Maiorino, M. An overview of mechanisms of redox signaling. J. Mol. Cell. Cardiol. 2014, 73, 2–9. [Google Scholar] [CrossRef]
- Sakkas, D.; Mariethoz, E.; Manicardi, G.; Bizzaro, D.; Bianchi, P.; Bianchi, U. Origin of DNA damage in ejaculated human spermatozoa. Rev. Reprod. 1999, 4, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Dutta, S.; Sengupta, P.; Slama, P.; Roychoudhury, S. Oxidative Stress, Testicular Inflammatory Pathways, and Male Reproduction. Int. J. Mol. Sci. 2021, 22, 10043. [Google Scholar] [CrossRef] [PubMed]
- Hosen, M.B.; Islam, M.R.; Begum, F.; Kabir, Y.; Howlader, M.Z.H. Oxidative Stress Induced Sperm DNA Damage, a Possible Reason for Male Infertility. Iran. J. Reprod. Med. 2015, 13, 525. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4637119/ (accessed on 22 April 2023). [PubMed]
- Gomez, E.; Irvine, D.S.; Aitken, R.J. Evaluation of a spectrophotometric assay for the measurement of malondialdehyde and 4-hydroxyalkenals in human spermatozoa: Relationships with semen quality and sperm function. Int. J. Androl. 1998, 21, 81–94. [Google Scholar] [CrossRef]
- Cilio, S.; Rienzo, M.; Villano, G.; Mirto, B.F.; Giampaglia, G.; Capone, F.; Ferretti, G.; Di Zazzo, E.; Crocetto, F. Beneficial Effects of Antioxidants in Male Infertility Management: A Narrative Review. Oxygen 2022, 2, 1. [Google Scholar] [CrossRef]
- Soumya, R.; Carl, H.; Vani, R. l-carnitine as a Potential Additive in Blood Storage Solutions: A Study on Erythrocytes. Indian J. Hematol. Blood Transfus. 2016, 32, 328. [Google Scholar] [CrossRef]
- Ahmadi, S.; Bashiri, R.; Ghadiri-Anari, A.; Nadjarzadeh, A. Antioxidant supplements and semen parameters: An evidence based review. Int. J. Reprod. Biomed. 2016, 14, 729–736. [Google Scholar] [CrossRef]
- Gamidov, S.G.; Ovchinnikov, R.O.; Popova, A.P. Double-blind, randomized placebo-controlled study of efficiency and safety of complex acetyl-L-carnitine, L-carnitine fumarate and alpha-lipoic acid (Spermactin® forte) for treatment of male infertility. Urologiia 2019, 2019, 62–68. [Google Scholar] [CrossRef]
- Longo, N.; Frigeni, M.; Pasquali, M. Carnitine transport and fatty acid oxidation. Biochim. Biophys. Acta 2016, 1863, 2422. [Google Scholar] [CrossRef] [PubMed]
- Pekala, J.; Patkowska-Sokola, B.; Bodkowski, R.; Jamroz, D.; Nowakowski, P.; Lochynski, S.; Librowski, T. L-Carnitine—Metabolic Functions and Meaning in Humans Life. Curr. Drug Metab. 2011, 12, 667–678. [Google Scholar] [CrossRef] [PubMed]
- Karlic, H.; Lohninger, A. Supplementation of L-Carnitine in athletes: Does it make sense? Nutrition 2004, 20, 709–715. [Google Scholar] [CrossRef] [PubMed]
- Adeva-Andany, M.M.; Calvo-Castro, I.; Fernández-Fernández, C.; Donapetry-García, C.; Pedre-Piñeiro, A.M. Significance of L-Carnitine for Human Health. IUBMB Life 2017, 69, 578–594. [Google Scholar] [CrossRef]
- Lenzi, A.; Sgro, P.; Salacone, P.; Paoli, D.; Gilio, B.; Lombardo, F.; Santulli, M.; Agarwal, A.; Gandini, L. A placebo-controlled double-blind randomized trial of the use of combined L-Carnitine and L-acetyl-carnitine treatment in men with asthenozoospermia. Fertil. Steril. 2004, 81, 1578–1584. [Google Scholar] [CrossRef]
- Jeulin, C.; Lewin, L. Role of free L-Carnitine and acetyl-L-Carnitine in post-gonadal maturation of mammalian spermatozoa. Hum. Reprod. Update 1996, 2, 87–102. [Google Scholar] [CrossRef]
- Mostafa, T.; Abougabal, K.; Mintziori, G.; Nabil, N.; Adel, M.; AboSief, A.F. Seminal L-Carnitine In Infertile Oligoasthenoteratozoospermic Men with Varicocele. J. Reprod. Infertil. 2022, 23, 26–32. [Google Scholar] [CrossRef]
- Lewin, L.M.; Beer, R.; Lunenfeld, B. Epididymis and seminal vesicle as sources of carnitine in human seminal fluid: The clinical significance of the carnitine concentration in human seminal fluid. Fertil. Steril. 1976, 27, 9–13. [Google Scholar] [CrossRef]
- Menchini-Fabris, G.F.; Canale, D.; Izzo, P.L.; Olivieri, L.; Bartelloni, M. Free L-Carnitine in human semen: Its variability in different andrologic pathologies. Fertil. Steril. 1984, 42, 263–267. [Google Scholar] [CrossRef]
- Alharthi, W.A.; Hamza, R.Z.; Elmahdi, M.M.; Abuelzahab, H.S.H.; Saleh, H. Selenium and L-Carnitine Ameliorate Reproductive Toxicity Induced by Cadmium in Male Mice. Biol. Trace Element Res. 2020, 197, 619–627. [Google Scholar] [CrossRef]
- Aliabadi, E.; Mehranjani, M.S.; Borzoei, Z.; Talaei-Khozani, T.; Mirkhani, H.; Tabesh, H. Effects of L-Carnitine and L-acetyl-carnitine on testicular sperm motility and chromatin quality. Iran. J. Reprod. Med. 2012, 10, 77–82. Available online: https://pubmed.ncbi.nlm.nih.gov/25242977/ (accessed on 1 May 2023). [PubMed]
- Cabral, R.E.L.; Mendes, T.B.; Vendramini, V.; Miraglia, S.M. Carnitine partially improves oxidative stress, acrosome integrity, and reproductive competence in doxorubicin-treated rats. Andrology 2018, 6, 236–246. [Google Scholar] [CrossRef] [PubMed]
- Thangasamy, T.; Jeyakumar, P.; Sittadjody, S.; Joyee, A.G.; Chinnakannu, P. L-Carnitine mediates protection against DNA damage in lymphocytes of aged rats. Biogerontology 2009, 10, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Elokil, A.A.; Bhuiyan, A.A.; Liu, H.-Z.; Hussein, M.N.; Ahmed, H.I.; Azmal, S.A.; Yang, L.; Li, S. The capability of L-Carnitine-mediated antioxidant on cock during aging: Evidence for the improved semen quality and enhanced testicular expressions of GnRH1, GnRHR, and melatonin receptors MT 1/2. Poult. Sci. 2019, 98, 4172–4181. [Google Scholar] [CrossRef]
- Cao, Y.; Wang, X.; Li, S.; Wang, H.; Yu, L.; Wang, P. The Effects of L-Carnitine against Cyclophosphamide-Induced Injuries in Mouse Testis. Basic Clin. Pharmacol. Toxicol. 2017, 120, 152–158. [Google Scholar] [CrossRef]
- Ma, L.; Sun, Y. Comparison of L-Carnitine vs. Coq10 and Vitamin E for idiopathic male infertility: A randomized controlled trial. Eur. Rev. Med. Pharmacol. Sci. 2022, 26, 4698–4704. [Google Scholar] [CrossRef]
- Wang, Y.-X.; Yang, S.-W.; Qu, C.-B.; Huo, H.-X.; Li, W.; Li, J.-D.; Chang, X.-L.; Cai, G.-Z. L-Carnitine: Safe and effective for asthenozoospermia. Zhonghua Nan Ke Xue = Natl. J. Androl. 2010, 16, 420–422. Available online: https://pubmed.ncbi.nlm.nih.gov/20684322/ (accessed on 10 April 2023).
- Kopets, R.; Kuibida, I.; Chernyavska, I.; Cherepanyn, V.; Mazo, R.; Fedevych, V.; Gerasymov, S. Dietary supplementation with a novel L-Carnitine multi-micronutrient in idiopathic male subfertility involving oligo-, astheno-, teratozoospermia: A randomized clinical study. Andrology 2020, 8, 1184–1193. [Google Scholar] [CrossRef]
- Agarwal, A.; Virk, G.; Ong, C.; Du Plessis, S.S. Effect of Oxidative Stress on Male Reproduction. World J. Men’s Health 2014, 32, 1–17. [Google Scholar] [CrossRef]
- Fielding, R.; Riede, L.; Lugo, J.P.; Bellamine, A. l-Carnitine Supplementation in Recovery after Exercise. Nutrients 2018, 10, 349. [Google Scholar] [CrossRef]
- Arenas, J.; Ricoy, J.R.; Encinas, A.R.; Pola, P.; D’Iddio, S.; Zeviani, M.; DiDonato, S.; Corsi, M. Carnitine in muscle, serum, and urine of nonprofessional athletes: Effects of physical exercise, training, and L-Carnitine administration. Muscle Nerve 1991, 14, 598–604. [Google Scholar] [CrossRef] [PubMed]
- Broad, E.M.; Maughan, R.J.; Galloway, S.D. Carbohydrate, protein, and fat metabolism during exercise after oral carnitine supplementation in humans. Int. J. Sport Nutr. Exerc. Metab. 2008, 18, 567–584. [Google Scholar] [CrossRef] [PubMed]
- Dragan, G.I.; Pleosteanu, E.; Selejan, V. Studies concerning the ergogenic value of Cantamega-2000® supply in top junior cyclists. Physiologie 1988, 25, 129–132. Available online: https://pubmed.ncbi.nlm.nih.gov/3144009/ (accessed on 27 October 2022).
- Giamberardino, M.A.; Dragani, L.; Valente, R.; Di Lisa, F.; Saggin, R.; Vecchiet, L. Effects of prolonged L-Carnitine administration on delayed muscle pain and CK release after eccentric effort. Int. J. Sports Med. 1996, 17, 320–324. [Google Scholar] [CrossRef] [PubMed]
- DrĂgan, I.G.; Vasiliu, A.; Georgescu, E.; Eremia, N. Studies concerning chronic and acute effects of L-Carnitina in elite athletes. Rev. Roum. Morphol. Physiol.-Ser. Physiol. 1989, 26, 111–129. Available online: https://pubmed.ncbi.nlm.nih.gov/2510191 (accessed on 27 October 2022).
- Parandak, K.; Arazi, H.; Khoshkhahesh, F.; Nakhostin-Roohi, B. The effect of two-week L-Carnitine supplementation on exercise—Induced oxidative stress and muscle damage. Asian J. Sports Med. 2014, 5, 123–128. Available online: https://pubmed.ncbi.nlm.nih.gov/25834706/ (accessed on 27 October 2022).
- Pistone, G.; Marino, A.D.; Leotta, C.; Dell’Arte, S.; Finocchiaro, G.; Malaguarnera, M. Levocarnitine administration in elderly subjects with rapid muscle fatigue: Effect on body composition, lipid profile and fatigue. Drugs Aging 2003, 20, 761–767. [Google Scholar] [CrossRef]
- McGarry, J.D.; Brown, N.F. The mitochondrial carnitine palmitoyltransferase system: From concept to molecular analysis. Eur. J. Biochem. 1997, 244, 1–14. [Google Scholar] [CrossRef]
- Cooper, D.E.; Young, P.A.; Klett, E.L.; Coleman, R.A. Physiological consequences of compartmentalized acyl-CoA metabolism. J. Biol. Chem. 2015, 290, 20023–20031. [Google Scholar] [CrossRef]
- Longo, N.; di San Filippo, C.A.; Pasquali, M. Disorders of carnitine transport and the carnitine cycle. Am. J. Med. Genet. Part C Semin. Med. Genet. 2006, 142C, 77–85. [Google Scholar] [CrossRef]
- Anderson, C.M.; Stahl, A. SLC27 fatty acid transport proteins. Mol. Asp. Med. 2013, 34, 516–528. [Google Scholar] [CrossRef] [PubMed]
- Jain, S.; Singh, S.N. Effect of L-Carnitine Supplementation on Nutritional Status and Physical Performance Under Calorie Restriction. Indian J. Clin. Biochem. 2015, 30, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.-R.; Hong, Y.M.; Boriack, R.L.; Bennett, M.J. Effect of L-Carnitine supplementation on cardiac carnitine palmitoyltransferase activities and plasma carnitine concentrations in adriamycin-treated rats. Pediatr. Res. 2003, 53, 788–792. [Google Scholar] [CrossRef]
- Jeukendrup, A.E. Regulation of fat metabolism in skeletal muscle. Ann. N. Y. Acad. Sci. 2002, 967, 217–235. [Google Scholar] [CrossRef] [PubMed]
- Institute of Medicine (US) Committee on Military Nutrition Research. Food Components to Enhance Performance: An Evaluation of Potential Performance-Enhancing Food Components for Operational Rations; The Role of Carnitine in Enhancing Physical Performance, Marriott, B.M., Eds.; National Academies Press (US): Washington, DC, USA, 1994. Available online: https://www.ncbi.nlm.nih.gov/books/NBK209052/ (accessed on 30 March 2023).
- Siliprandi, N.; Di Lisa, F.; Pieralisi, G.; Ripari, P.; Maccari, F.; Menabo, R.; Giamberardino, M.A.; Vecchiat, L. Metabolic changes induced by maximal exercise in human subjects following L-Carnitine administration. Biochim. Biophys. Acta (BBA)—Gen. Subj. 1990, 1034, 17–21. [Google Scholar] [CrossRef]
- Wanders, R.J.; Visser, G.; Ferdinandusse, S.; Vaz, F.M.; Houtkooper, R.H. Mitochondrial Fatty Acid Oxidation Disorders: Laboratory Diagnosis, Pathogenesis, and the Complicated Route to Treatment. J. Lipid Atheroscler. 2020, 9, 313. [Google Scholar] [CrossRef]
- Virmani, M.A.; Cirulli, M. The Role of l-Carnitine in Mitochondria, Prevention of Metabolic Inflexibility and Disease Initiation. Int. J. Mol. Sci. 2022, 23, 2717. [Google Scholar] [CrossRef]
- Virmani, A.; Binienda, Z. Role of carnitine esters in brain neuropathology. Mol. Asp. Med. 2004, 25, 533–549. [Google Scholar] [CrossRef]
- Hülsmann, W.; Dubelaar, M.-L.; Lamers, J.; Maccari, F. Protection by acyl-carnitines and phenylmethylsulfonyl fluoride of rat heart subjected to ischemia and reperfusion. Biochim. Biophys. Acta (BBA)—Mol. Cell Res. 1985, 847, 62–66. [Google Scholar] [CrossRef]
- Vescovo, G.; Ravara, B.; Gobbo, V.; Sandri, M.; Angelini, A.; Della Barbera, M.; Dona, M.; Peluso, G.; Calvani, M.; Mosconi, L.; et al. L-Carnitine: A potential treatment for blocking apoptosis and preventing skeletal muscle myopathy in heart failure. Am. J. Physiol. Cell Physiol. 2002, 283, C802–C810. [Google Scholar] [CrossRef]
- Afshin-Majd, S.; Bashiri, K.; Kiasalari, Z.; Baluchnejadmojarad, T.; Sedaghat, R.; Roghani, M. Acetyl-L-carnitine protects dopaminergic nigrostriatal pathway in 6-hydroxydopamine-induced model of Parkinson’s disease in the rat. Biomed. Pharmacother. 2017, 89, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Abdelrazik, H.; Sharma, R.; Mahfouz, R.; Agarwal, A. L-Carnitine decreases DNA damage and improves the in vitro blastocyst development rate in mouse embryos. Fertil. Steril. 2009, 91, 589–596. [Google Scholar] [CrossRef] [PubMed]
- Cankorkmaz, L.; Köylüoğlu, G.; Özer, H.; Yıldız, E.; Sümer, Z.; Özdemir, Ö. The role of apoptosis and protective effect of carnitine in contralateral testicular injury in experimental unilateral testicular torsion. Turk. J. Trauma Emerg. Surg. 2009, 15, 529–534. [Google Scholar]
- Ranger, A.M.; Malynn, B.A.; Korsmeyer, S.J. Mouse models of cell death. Nat. Genet. 2001, 28, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Vardiyan, R.; Ezati, D.; Anvari, M.; Ghasemi, N.; Talebi, A. Effect of L-carnitine on the expression of the apoptotic genes Bcl-2 and Bax. Clin. Exp. Reprod. Med. 2020, 47, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Altun, Z.; Olgun, Y.; Ercetin, P.; Aktas, S.; Kirkim, G.; Serbetcioglu, B.; Olgun, N.; Guneri, E.A. Protective effect of acetyl-l-carnitine against cisplatin ototoxicity: Role of apoptosis-related genes and pro-inflammatory cytokines. Cell Prolif. 2014, 47, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Mutomba, M.C.; Yuan, H.; Konyavko, M.; Adachi, S.; Yokoyama, C.B.; Esser, V.; McGarry, J.; Babior, B.M.; Gottlieb, R.A. Regulation of the activity of caspases by L-carnitine and palmitoylcarnitine. FEBS Lett. 2000, 478, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Hirschey, M.D.; Shimazu, T.; Goetzman, E.; Jing, E.; Schwer, B.; Lombard, D.B.; Grueter, C.A.; Harris, C.; Biddinger, S.; Ilkayeva, O.R.; et al. SIRT3 regulates mitochondrial fatty-acid oxidation by reversible enzyme deacetylation. Nature 2010, 464, 121–125. [Google Scholar] [CrossRef]
- Cao, Y.; Li, X.; Wang, C.-J.; Li, P.; Yang, B.; Wang, C.-B.; Wang, L.-X. Role of NF-E2-related factor 2 in neuroprotective effect of L-Carnitine against high glucose-induced oxidative stress in the retinal ganglion cells. Biomed. Pharmacother. 2015, 69, 345–348. [Google Scholar] [CrossRef]
- Morigi, M.; Perico, L.; Rota, C.; Longaretti, L.; Conti, S.; Rottoli, D.; Novelli, R.; Remuzzi, G.; Benigni, A. Sirtuin 3–dependent mitochondrial dynamic improvements protect against acute kidney injury. J. Clin. Investig. 2015, 125, 715–726. [Google Scholar] [CrossRef]
- Dikalova, A.E.; Pandey, A.; Xiao, L.; Arslanbaeva, L.; Sidorova, T.; Lopez, M.G.; Billings, F.T.; Verdin, E.; Auwerx, J.; Harrison, D.G.; et al. Mitochondrial Deacetylase Sirt3 Reduces Vascular Dysfunction and Hypertension While Sirt3 Depletion in Essential Hypertension Is Linked to Vascular Inflammation and Oxidative Stress. Circ. Res. 2020, 126, 439–452. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Wu, Q.; Shi, J.; Zhou, S. Regulation of SIRT3 on mitochondrial functions and oxidative stress in Parkinson’s disease. Biomed. Pharmacother. 2020, 132, 110928. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Chen, S.; Zhang, B.; Liu, J. SIRT3 as a potential therapeutic target for heart failure. Pharmacol. Res. 2021, 165, 105432. [Google Scholar] [CrossRef]
- Zhang, D.-M.; Guo, Z.-X.; Zhao, Y.-L.; Wang, Q.-J.; Gao, Y.-S.; Yu, T.; Chen, Y.-K.; Chen, X.-M.; Wang, G.-Q. L-Carnitine regulated Nrf2/Keap1 activation in vitro and in vivo and protected oxidized fish oil-induced inflammation response by inhibiting the NF-κB signaling pathway in Rhynchocypris lagowski Dybowski. Fish Shellfish. Immunol. 2019, 93, 1100–1110. [Google Scholar] [CrossRef]
- Nguyen, T.; Nioi, P.; Pickett, C.B. The Nrf2-Antioxidant Response Element Signaling Pathway and Its Activation by Oxidative Stress. J. Biol. Chem. 2009, 284, 13291–13295. [Google Scholar] [CrossRef]
- Aydos, O.S.; Yukselten, Y.; Aydos, D.; Sunguroglu, A.; Aydos, K. Relationship between functional Nrf2 gene promoter polymorphism and sperm DNA damage in male infertility. Syst. Biol. Reprod. Med. 2021, 67, 399–412. [Google Scholar] [CrossRef] [PubMed]
- Han, P.; Wang, X.; Zhou, T.; Cheng, J.; Wang, C.; Sun, F.; Zhao, X. Inhibition of ferroptosis attenuates oligospermia in male Nrf2 knockout mice. Free Radic. Biol. Med. 2022, 193 Pt 1, 421–429. [Google Scholar] [CrossRef]
- Wang, Z.-Y.; Liu, Y.-Y.; Liu, G.-H.; Lu, H.-B.; Mao, C.-Y. L-Carnitine and heart disease. Life Sci. 2018, 194, 88–97. [Google Scholar] [CrossRef]
- Ferreira, G.C.; McKenna, M.C. L-Carnitine and Acetyl-l-carnitine Roles and Neuroprotection in Developing Brain. Neurochem. Res. 2017, 42, 1661–1675. [Google Scholar] [CrossRef]
- Fattorossi, A.; Biselli, R.; Casciaro, A.; Tzantzoglou, S.; de Simone, C. Regulation of normal human polyrnorphonuclear leucocytes by carnitine. Mediat. Inflamm. 1993, 2, S37. [Google Scholar] [CrossRef]
- Matalliotakis, I.; Koumantaki, Y.; Evageliou, A.; Matalliotakis, G.; Goumenou, A.; Koumantakis, E. L-Carnitine levels in the seminal plasma of fertile and infertile men: Correlation with sperm quality. Int. J. Fertil. Women’s Med. 2000, 45, 236–240. Available online: https://pubmed.ncbi.nlm.nih.gov/10929687/ (accessed on 27 October 2022).
- Vicari, E.; Calogero, A. Effects of treatment with carnitines in infertile patients with prostato-vesiculo-epididymitis. Hum. Reprod. 2001, 16, 2338–2342. [Google Scholar] [CrossRef] [PubMed]
- Casillas, E.R. The distribution of carnitine in male reproductive tissues and its effect on palmitate oxidation by spermatozoal particles. Biochim. Biophys. Acta (BBA)—Lipids Lipid Metab. 1972, 280, 545–551. [Google Scholar] [CrossRef]
- Casillas, E.R.; Chaipayungpan, S. The distribution of carnitine and acetylcarnitine in the rabbit epididymis and the carnitine content of rabbit spermatozoa during maturation. Reproduction 1979, 56, 439–444. [Google Scholar] [CrossRef]
- Hinton, B.T.; Brooks, D.E.; Dott, H.M.; Setchell, B.P. Effects of carnitine and some related compounds on the motility of rat spermatozoa from the caput epididymidis. J. Reprod. Fertil. 1981, 61, 59–64. [Google Scholar] [CrossRef]
- Palmero, S.; Bottazzi, C.; Costa, M.; Leone, M.; Fugassa, E. Metabolic effects of L-Carnitine on prepubertal rat Sertoli cells. Horm. Metab. Res. 2000, 32, 87–90. [Google Scholar] [CrossRef]
- Caviglia, D.; Scarabelli, L.; Palmero, S. Effects of Carnitines on Rat Sertoli Cell Protein Metabolism. Horm. Metab. Res. 2004, 36, 221–225. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, D.; Goto, A.; Maeda, T.; Nezu, J.-I.; Tsuji, A.; Tamai, I. OCTN2-mediated transport of carnitine in isolated Sertoli cells. Reproduction 2005, 129, 729–736. [Google Scholar] [CrossRef]
- Khushboo, M.; Murthy, M.K.; Devi, M.S.; Sanjeev, S.; Ibrahim, K.S.; Kumar, N.S.; Roy, V.K.; Gurusubramanian, G. Testicular toxicity and sperm quality following copper exposure in Wistar albino rats: Ameliorative potentials of L-Carnitine. Environ. Sci. Pollut. Res. Int. 2018, 25, 1837–1862. [Google Scholar] [CrossRef]
- Karam, K.M.; Alebady, A.S.; Al-Nailey, K.G.C.; Al-Delemi, D.H.J. L-Carnitine effect on induced hyperlipidemia on premature rats: Fertility profile. J. Med. Life 2022, 15, 124–131. [Google Scholar] [CrossRef]
- Abdel-Emam, R.A.; Ahmed, E.A. Ameliorative effect of L-Carnitine on chronic lead-induced reproductive toxicity in male rats. Vet. Med. Sci. 2021, 7, 1426–1435. [Google Scholar] [CrossRef]
- Longobardi, V.; Salzano, A.; Campanile, G.; Marrone, R.; Palumbo, F.; Vitiello, M.; Zullo, G.; Gasparrini, B. Carnitine supplementation decreases capacitation-like changes of frozen-thawed buffalo spermatozoa. Theriogenology 2017, 88, 236–243. [Google Scholar] [CrossRef]
- Fu, L.-L.; Zhang, L.-Y.; An, Q.; Zhou, F.; Tong, Y.; Guo, Y.; Lu, W.-H.; Liang, X.-W.; Chang, B.; Gu, Y.-Q. L-Carnitine protects the motion parameters and mitochondrial function of human sperm in cryopreservation. Zhonghua Nan Ke Xue = Natl. J. Androl. 2018, 24, 1059–1063. [Google Scholar]
- Abd-Elrazek, A.M.; Ahmed-Farid, O.A.H. Protective effect of L-Carnitine and L-Arginine against busulfan-induced oligospermia in adult rat. Andrologia 2017, 50, e12806. [Google Scholar] [CrossRef] [PubMed]
- Khademi, A.; Alleyassin, A.; Safdarian, L.; Hamed, E.A.; Rabiee, E.; Haghaninezhad, H. The effects of L-Carnitine on sperm parameters in smoker and non-smoker patients with idiopathic sperm abnormalities. J. Assist. Reprod. Genet. 2005, 22, 395–399. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.-L.; Wan, X.-X.; Zhang, Y.; Zhang, Y.-H.; Zhao, W.-J.; Wang, D.; Wang, J.-G.; Xie, J.-L.; Ma, H.-G. L-Carnitine improves sperm acrosin activity in male infertility patients. Zhonghua Nan Ke Xue = Natl. J. Androl. 2018, 24, 1064–1068. Available online: https://pubmed.ncbi.nlm.nih.gov/32212483/ (accessed on 10 April 2023).
- Nezhad, N.C.; Vahabzadeh, Z.; Allahveisie, A.; Rahmani, K.; Raoofi, A.; Rezaie, M.J.; Rezaei, M.; Partovyan, M. The Effect of L-Carnitine and Coenzyme Q10 on the Sperm Motility, DNA Fragmentation, Chromatin Structure and Oxygen Free Radicals During, before and after Freezing in Oligospermia Men. Urol. J. 2021, 18, 330–336. [Google Scholar] [CrossRef]
- Busetto, G.M.; Agarwal, A.; Virmani, A.; Antonini, G.; Ragonesi, G.; Del Giudice, F.; Micic, S.; Gentile, V.; De Berardinis, E. Effect of metabolic and antioxidant supplementation on sperm parameters in oligo-astheno-teratozoospermia, with and without varicocele: A double-blind placebo-controlled study. Andrologia 2018, 50, e12927. [Google Scholar] [CrossRef]
- Rebouche, C.J. Carnitine. In Modern Nutrition in Health and Disease, 9th ed.; Shils, M.E., Olson, J.A., Shike, M., Ross, A.C., Eds.; Lippincott Williams & Wilkins: New York, NY, USA, 1999; pp. 505–512. [Google Scholar]
- Alesci, S.; Manoli, I.; Costello, R.; Coates, P.; Gold, P.W.; Chrousos, G.P.; Blackman, M.R. Carnitine: Lessons from one hundred years of research. Ann. N. Y. Acad. Sci. 2004, 1033, ix–xi. [Google Scholar]
- Diao, R.; Gan, H.; Tian, F.; Cai, X.; Zhen, W.; Song, X.; Duan, Y. In vitro antioxidation effect of Quercetin on sperm function from the infertile patients with leukocytospermia. Am. J. Reprod. Immunol. 2019, 82, e13155. [Google Scholar] [CrossRef]
- Pasquariello, R.; Verdile, N.; Brevini, T.A.L.; Gandolfi, F.; Boiti, C.; Zerani, M.; Maranesi, M. The Role of Resveratrol in Mammalian Reproduction. Molecules 2020, 25, 4554. [Google Scholar] [CrossRef] [PubMed]
- Micic, S.; Lalic, N.; Djordjevic, D.; Bojanic, N.; Bogavac-Stanojevic, N.; Busetto, G.M.; Virmani, A.; Agarwal, A. Double-blind, randomised, placebo-controlled trial on the effect of L-Carnitine and L-Acetylcarnitine on sperm parameters in men with idiopathic oligoasthenozoospermia. Andrologia 2019, 51, e13267. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mateus, F.G.; Moreira, S.; Martins, A.D.; Oliveira, P.F.; Alves, M.G.; Pereira, M.d.L. L-Carnitine and Male Fertility: Is Supplementation Beneficial? J. Clin. Med. 2023, 12, 5796. https://doi.org/10.3390/jcm12185796
Mateus FG, Moreira S, Martins AD, Oliveira PF, Alves MG, Pereira MdL. L-Carnitine and Male Fertility: Is Supplementation Beneficial? Journal of Clinical Medicine. 2023; 12(18):5796. https://doi.org/10.3390/jcm12185796
Chicago/Turabian StyleMateus, Filipa G., Silvia Moreira, Ana D. Martins, Pedro F. Oliveira, Marco G. Alves, and Maria de Lourdes Pereira. 2023. "L-Carnitine and Male Fertility: Is Supplementation Beneficial?" Journal of Clinical Medicine 12, no. 18: 5796. https://doi.org/10.3390/jcm12185796