
EMT Involved in Migration of Stem/Progenitor Cells for Pituitary Development and Regeneration

Abstract
:1. Introduction
2. Pituitary Stem/Progenitor Cells and Their Niche
2.1. Identification of Pituitary Stem/Progenitor Cells
2.2. Differentiation Ability of SOX2+-Pituitary Stem/Progenitor Cells
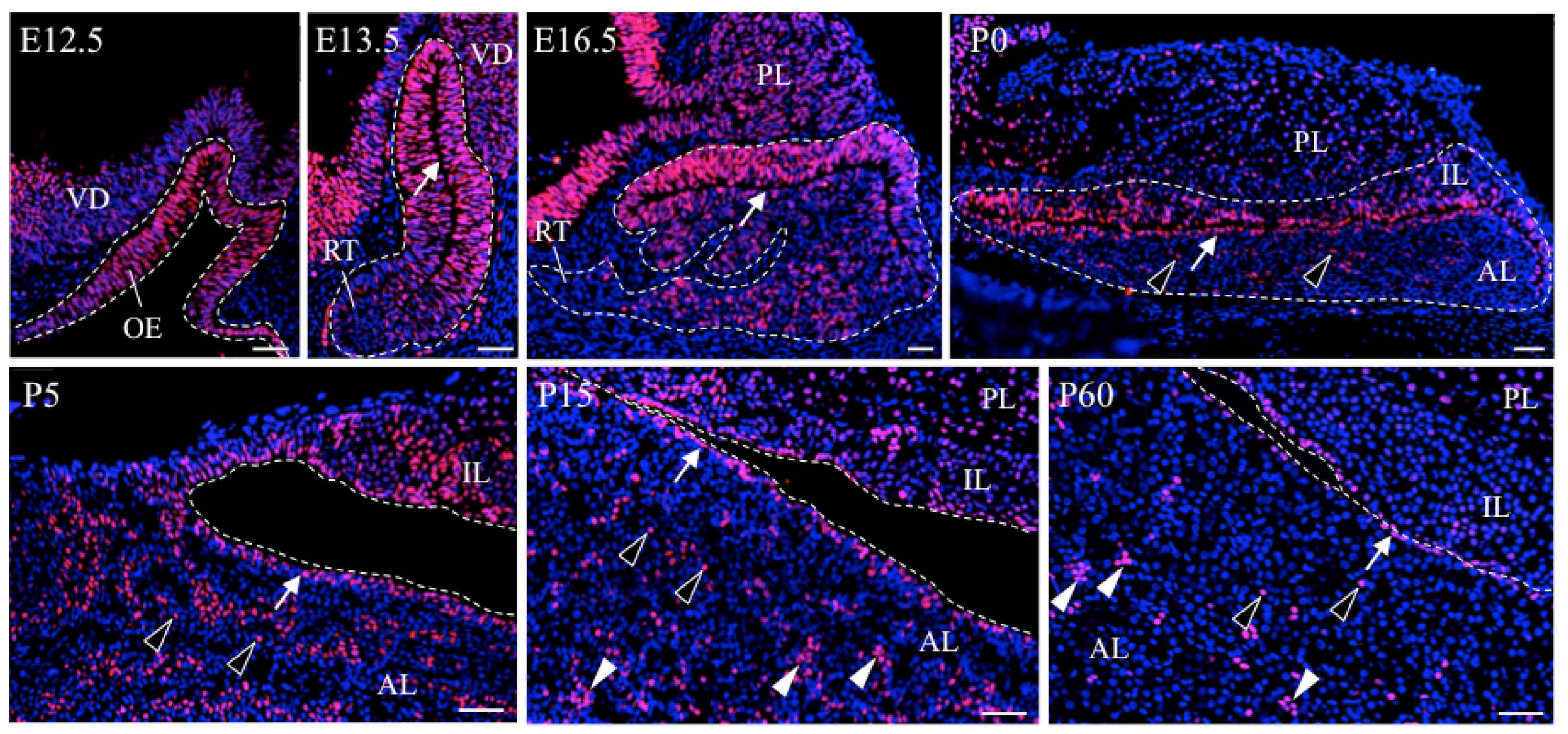
2.3. Characteristic Localization of SOX2+-Pituitary Stem/Progenitor Cells in the Developing and Postnatal Pituitary
2.4. Heterogeneity of SOX2+-Cells in the Adult Pituitary
3. Relationship between Migration of Stem/Progenitor Cells and EMT
3.1. EMT Involved in Migration of Neural Crest Cells during Development
- (I)
- Induction of key transcription factors. In the process of EMT, SNAILs, TWISTs, and Zinc-finger E-box-binding (ZEB) transcription factors play important roles [46]. SNAILs (SNAIL1 and SLUG) are required not only for EMT, but also for specification of neural crest cells, and are induced by signaling pathways such as Wnt [47,48]. In addition, TGFβ (transforming growth factor, beta) plays roles in the regulatory signaling pathway for SNAILs, TWISTs, and ZEBs [49].
- (II)
- Delamination from neural tube. Changes in cell–cell adhesion molecules enable the initiation of cell migration. The most important cell surface molecule in the delamination is E-cadherin, which belongs to a type I cadherin and is known to form strong cell–cell interactions for epithelial stabilization. Therefore, downregulation of E-cadherin and its replacement with a type II cadherin such as Cadherin7 or Cadherin11 (cadherin switching) are important steps for cell migration [14]. In the process of downregulation of the E-cadherin, SNAIL1, SLUG, TWISTs, and ZEB2 act as transcriptional repressors for E-cadherin expression via direct binding to the E-cadherin promoter [46,50].
- (III)
- Acquisition of mesenchymal properties. Neural crest cells lose their polarity and start to migrate across the extracellular matrix (ECM). To digest the ECM, neural crest cells produce MMPs (matrix metalloproteases) and ADAMs (A Disintegrin and Metalloproteases) in a manner similar to invasion and metastasis of cancer cells. Interestingly, SNAIL1, SLUG, and ZEB2 also act as stimulators of MMPs and ADAM proteins [51].
- (IV)
- Directional migration. Orientation of the migration of neural crest cells into the destined areas is hypothesized to be directed by multiplex factors such as cytokines, chemokines, signaling molecules (e.g., TGFβ), and juxtacrine factors (e.g., ephrin/Eph) [52]. Among them, signaling introduced by CXCL12 (stromal cell-derived factor-1; SDF1), a member of the CXC chemokine family, and its receptor CXCR4 promote migration toward the dorsal root ganglia (DRG) [53] and sympathetic ganglia (SG) [54].
3.2. Migration of Stem/Progenitor Cells in the Pituitary Development
3.2.1. Migration of Stem/Progenitor Cells from the MCL Niche during Pituitary Organogenesis
3.2.2. PROP1 as a Candidate Factor for Regulating Cell Migration in Pituitary Organogenesis
3.2.3. Role of PROP1 in EMT Giving Rise to Cell Migration in the Pituitary Organogenesis
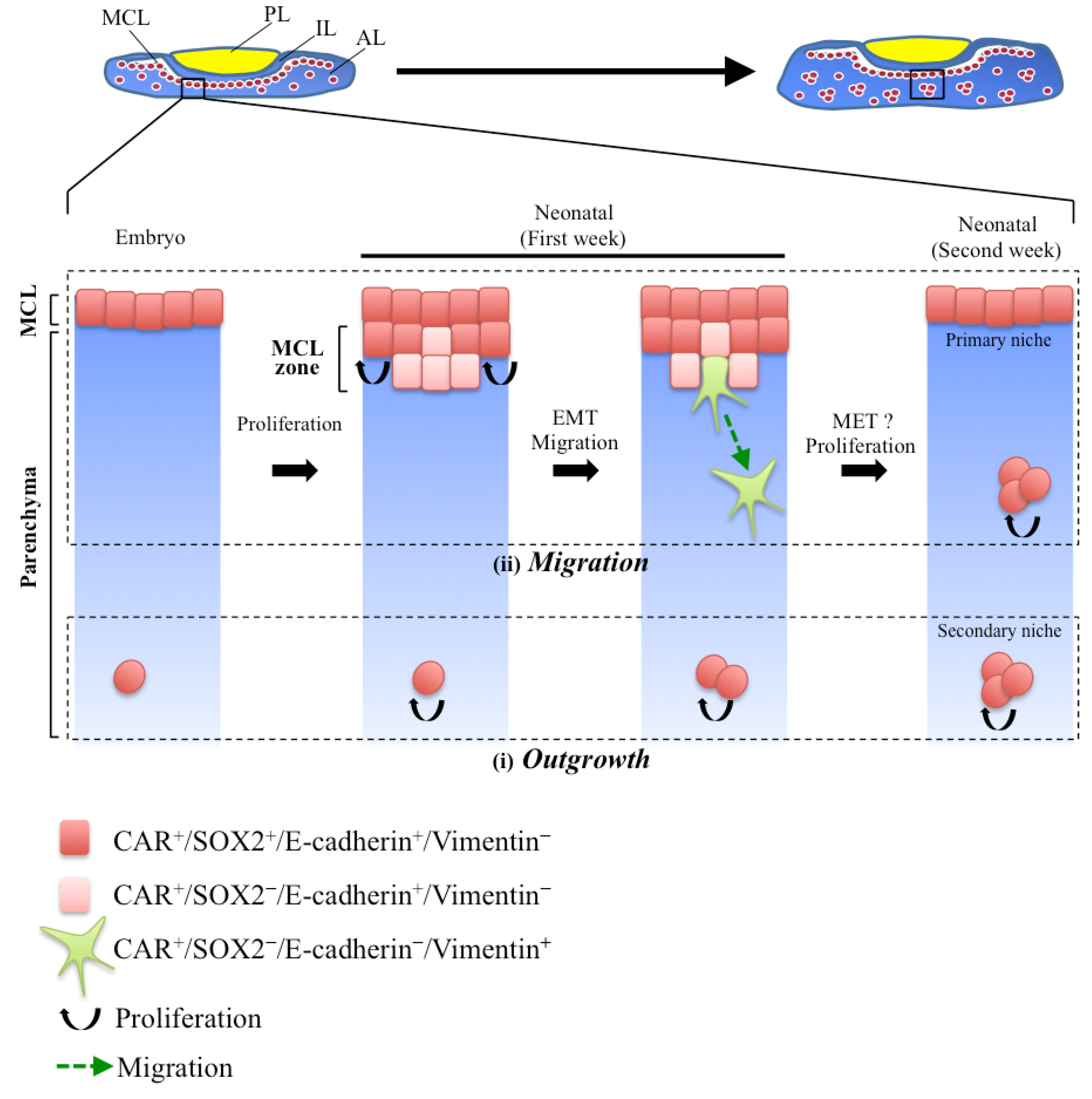
4. Relationship between EMT and Cell Migration in the Pituitary Postnatal Growth Wave
4.1. Secondary Niche Formation in the Early Postnatal Pituitary
4.2. CAR+-Stem/Progenitor Cells Undergoing EMT in the Process of Parenchymal Niche Formation
5. EMT Involved in Cell Migration in the Process of Cell Regeneration in the Adult Pituitary
5.1. Regeneration of Pituitary Cells
5.2. Potential Abilities for EMT in the Adult Pituitary Stem/Progenitor Cells
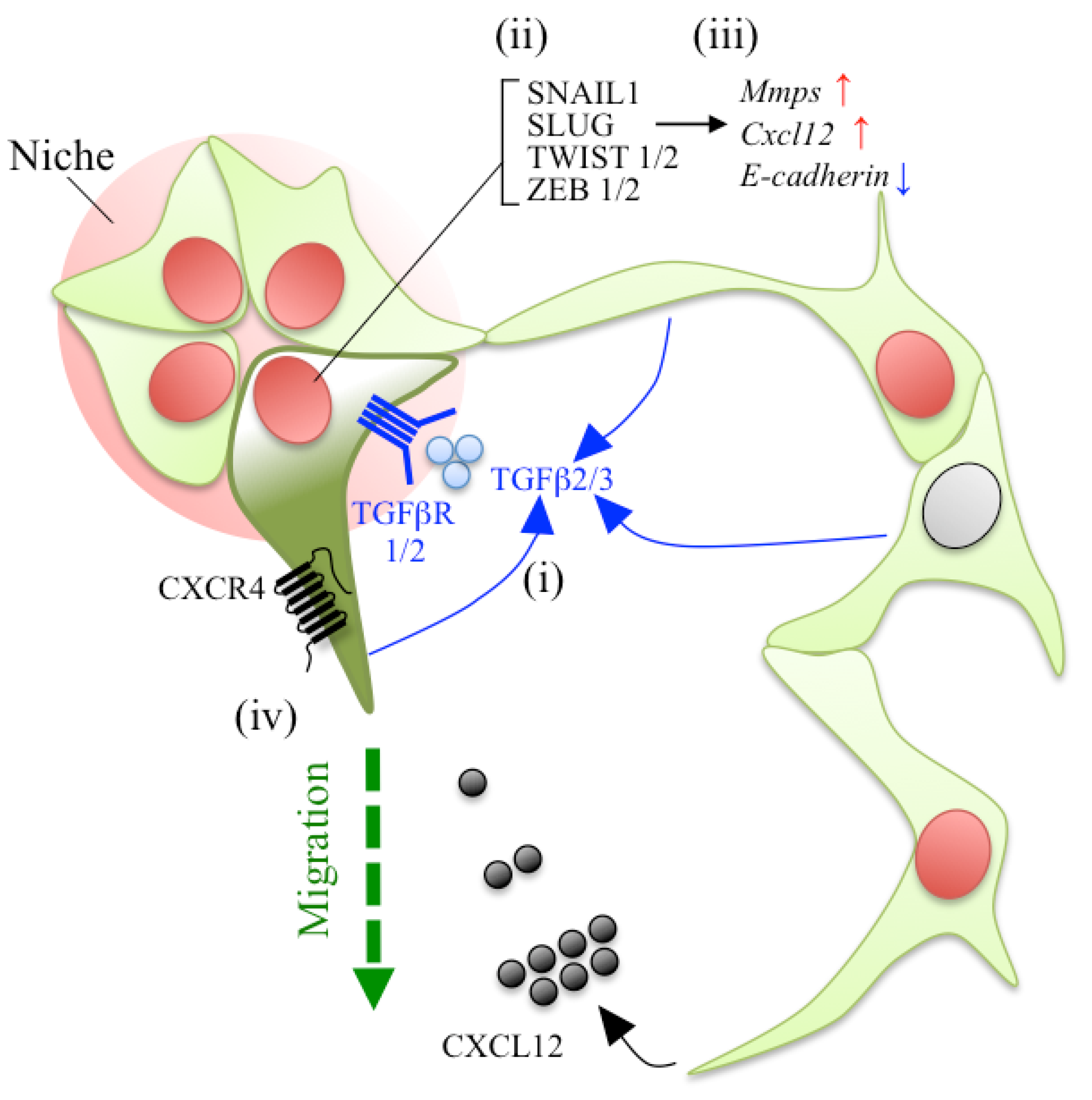
5.3. Chemokines in the Process of EMT and Cell Migration
5.4. A Proposal Model of Stem/Progenitor Cell Migration in the Adult Pituitary
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Zhu, X.; Gleiberman, A.S.; Rosenfeld, M.G. Molecular physiology of pituitary development: Signaling and transcriptional networks. Physiol. Rev. 2007, 87, 933–963. [Google Scholar] [CrossRef] [PubMed]
- Ward, R.D.; Raetzman, L.T.; Suh, H.; Stone, B.M.; Nasonkin, I.O.; Camper, S.A. Role of PROP1 in pituitary gland growth. Mol. Endocrinol. 2005, 19, 698–710. [Google Scholar] [CrossRef] [PubMed]
- Levy, A. Physiological implications of pituitary trophic activity. J. Endocrinol. 2002, 174, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Levy, A. Stem cells, hormones and pituitary adenomas. J. Neuroendocrinol. 2008, 20, 139–140. [Google Scholar] [CrossRef] [PubMed]
- Nolan, L.A.; Kavanagh, E.; Lightman, S.L.; Levy, A. Anterior pituitary cell population control: Basal cell turnover and the effects of adrenalectomy and dexamethasone treatment. J. Neuroendocrinol. 1998, 10, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Castinetti, F.; Davis, S.W.; Brue, T.; Camper, S.A. Pituitary stem cell update and potential implications for treating hypopituitarism. Endocr. Rev. 2011, 32, 453–471. [Google Scholar] [CrossRef] [PubMed]
- Vankelecom, H.; Gremeaux, L. Stem cells in the pituitary gland: A burgeoning field. Gen. Comp. Endocrinol. 2010, 166, 478–488. [Google Scholar] [CrossRef] [PubMed]
- Vankelecom, H. Pituitary stem cells drop their mask. Curr. Stem Cell Res. Ther. 2012, 7, 36–71. [Google Scholar] [CrossRef] [PubMed]
- Vankelecom, H.; Chen, J. Pituitary stem cells: Where do we stand? Mol. Cell. Endocrinol. 2014, 385, 2–17. [Google Scholar] [CrossRef] [PubMed]
- Andoniadou, C.L.; Matsushima, D.; Mousavy Gharavy, S.N.; Signore, M.; Mackintosh, A.I.; Schaeffer, M.; Gaston-Massuet, C.; Mollard, P.; Jacques, T.S.; Le Tissier, P.; et al. Sox2(+) stem/progenitor cells in the adult mouse pituitary support organ homeostasis and have tumor-inducing potential. Cell Stem Cell 2013, 13, 433–445. [Google Scholar] [CrossRef] [PubMed]
- Fauquier, T.; Rizzoti, K.; Dattani, M.; Lovell-Badge, R.; Robinson, I.C. Sox2-expressing progenitor cells generate all of the major cell types in the adult mouse pituitary gland. Proc. Natl. Acad. Sci. USA 2008, 105, 2907–2912. [Google Scholar] [CrossRef] [PubMed]
- Rizzoti, K.; Akiyama, H.; Lovell-Badge, R. Mobilized adult pituitary stem cells contribute to endocrine regeneration in response to physiological demand. Cell Stem Cell 2013, 13, 419–432. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, A.; Hochedlinger, K. The sox family of transcription factors: Versatile regulators of stem and progenitor cell fate. Cell Stem Cell 2013, 12, 15–30. [Google Scholar] [CrossRef] [PubMed]
- Simoes-Costa, M.; Bronner, M.E. Establishing neural crest identity: A gene regulatory recipe. Development 2015, 142, 242–257. [Google Scholar] [CrossRef] [PubMed]
- Davis, F.M.; Stewart, T.A.; Thompson, E.W.; Monteith, G.R. Targeting EMT in cancer: Opportunities for pharmacological intervention. Trends Pharmacol. Sci. 2014, 35, 479–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, Y.C.; Fuchs, E. A family business: Stem cell progeny join the niche to regulate homeostasis. Nat. Rev. Mol. Cell Biol. 2012, 13, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Murry, C.E.; Keller, G. Differentiation of embryonic stem cells to clinically relevant populations: Lessons from embryonic development. Cell 2008, 132, 661–680. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, F.; Harumiya, K.; Ishikawa, H.; Otsuka, Y. Differentiation of isolated chromophobes into acidophils or basophils when transplanted into the hypophysiotrophic area of hypothalamus. Endocrinol. Jpn. 1969, 16, 531–540. [Google Scholar] [CrossRef] [PubMed]
- Castrique, E.; Fernandez-Fuente, M.; Le Tissier, P.; Herman, A.; Levy, A. Use of a prolactin-CRE/rosa-YFP transgenic mouse provides no evidence for lactotroph transdifferentiation after weaning, or increase in lactotroph/somatotroph proportion in lactation. J. Endocrinol. 2010, 205, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Nolan, L.A.; Levy, A. A population of non-luteinising hormone/non-adrenocorticotrophic hormone-positive cells in the male rat anterior pituitary responds mitotically to both gonadectomy and adrenalectomy. J. Neuroendocrinol. 2006, 18, 655–661. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Hersmus, N.; Van Duppen, V.; Caesens, P.; Denef, C.; Vankelecom, H. The adult pituitary contains a cell population displaying stem/progenitor cell and early embryonic characteristics. Endocrinology 2005, 146, 3985–3998. [Google Scholar] [CrossRef] [PubMed]
- Goodell, M.A.; Brose, K.; Paradis, G.; Conner, A.S.; Mulligan, R.C. Isolation and functional properties of murine hematopoietic stem cells that are replicating in vivo. J. Exp. Med. 1996, 183, 1797–1806. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Crabbe, A.; Van Duppen, V.; Vankelecom, H. The notch signaling system is present in the postnatal pituitary: Marked expression and regulatory activity in the newly discovered side population. Mol. Endocrinol. 2006, 20, 3293–3307. [Google Scholar] [CrossRef] [PubMed]
- Pastrana, E.; Silva-Vargas, V.; Doetsch, F. Eyes wide open: A critical review of sphere-formation as an assay for stem cells. Cell Stem Cell 2011, 8, 486–498. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Gremeaux, L.; Fu, Q.; Liekens, D.; Van Laere, S.; Vankelecom, H. Pituitary progenitor cells tracked down by side population dissection. Stem Cells 2009, 27, 1182–1195. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, S.; Kato, T.; Susa, T.; Cai, L.-Y.; Nakayama, M.; Kato, Y. Prop1 coexists with sox2 and induces pit1-commitment cells. Biochem. Biophys. Res. Commun. 2009, 385, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Ward, R.D.; Stone, B.M.; Raetzman, L.T.; Camper, S.A. Cell proliferation and vascularization in mouse models of pituitary hormone deficiency. Mol. Endocrinol. 2006, 20, 1378–1390. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, S.; Kato, T.; Higuchi, M.; Yako, H.; Chen, M.; Kanno, N.; Ueharu, H.; Kato, Y. Rapid transition of nestin-expressing dividing cells from PROP1-positive to PIT1-positive advances prenatal pituitary development. J. Neuroendocrinol. 2013, 25, 779–791. [Google Scholar] [CrossRef] [PubMed]
- Japon, M.A.; Rubinstein, M.; Low, M.J. In situ hybridization analysis of anterior pituitary hormone gene expression during fetal mouse development. J. Histochem. Cytochem. 1994, 42, 1117–1125. [Google Scholar] [CrossRef] [PubMed]
- Gremeaux, L.; Fu, Q.; Chen, J.; Vankelecom, H. Activated phenotype of the pituitary stem/progenitor cell compartment during the early-postnatal maturation phase of the gland. Stem Cells Dev. 2012, 21, 801–813. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Kato, T.; Higuchi, M.; Yoshida, S.; Yako, H.; Kanno, N.; Kato, Y. Coxsackievirus and adenovirus receptor-positive cells compose the putative stem/progenitor cell niches in the marginal cell layer and parenchyma of the rat anterior pituitary. Cell Tissue Res. 2013, 354, 823–836. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.C.; Chuong, C.M. Multi-layered environmental regulation on the homeostasis of stem cells: The saga of hair growth and alopecia. J. Dermatol. Sci. 2012, 66, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Rezza, A.; Sennett, R.; Rendl, M. Adult stem cell niches: Cellular and molecular components. Curr. Top. Dev. Biol. 2014, 107, 333–372. [Google Scholar] [PubMed]
- Wabik, A.; Jones, P.H. Switching roles: The functional plasticity of adult tissue stem cells. EMBO J. 2015, 34, 1164–1179. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Clevers, H. Growing self-organizing mini-guts from a single intestinal stem cell: Mechanism and applications. Science 2013, 340, 1190–1194. [Google Scholar] [CrossRef] [PubMed]
- Gucciardo, E.; Sugiyama, N.; Lehti, K. Eph- and ephrin-dependent mechanisms in tumor and stem cell dynamics. Cell. Mol. Life Sci. 2014, 71, 3685–3710. [Google Scholar] [CrossRef] [PubMed]
- Gattazzo, F.; Urciuolo, A.; Bonaldo, P. Extracellular matrix: A dynamic microenvironment for stem cell niche. Biochim. Biophys. Acta 2014, 1840, 2506–2519. [Google Scholar] [CrossRef] [PubMed]
- Vankelecom, H. Pituitary stem/progenitor cells: Embryonic players in the adult gland? Eur. J. Neurosci. 2010, 32, 2063–2081. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, S.; Kato, T.; Yako, H.; Susa, T.; Cai, L.Y.; Osuna, M.; Inoue, K.; Kato, Y. Significant quantitative and qualitative transition in pituitary stem/progenitor cells occurs during the postnatal development of the rat anterior pituitary. J. Neuroendocrinol. 2011, 23, 933–943. [Google Scholar] [CrossRef] [PubMed]
- Soji, T.; Sirasawa, N.; Kurono, C.; Yashiro, T.; Herbert, D.C. Immunohistochemical study of the post-natal development of the folliculo-stellate cells in the rat anterior pituitary gland. Tissue Cell 1994, 26, 1–8. [Google Scholar] [CrossRef]
- Devnath, S.; Inoue, K. An insight to pituitary folliculo-stellate cells. J. Neuroendocrinol. 2008, 20, 687–691. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Lavandeira, M.; Quereda, V.; Flores, I.; Saez, C.; Diaz-Rodriguez, E.; Japon, M.A.; Ryan, A.K.; Blasco, M.A.; Dieguez, C.; Malumbres, M.; et al. A GRFa2/Prop1/stem (GPS) cell niche in the pituitary. PLoS ONE 2009, 4, e4815. [Google Scholar] [CrossRef] [PubMed]
- Nantie, L.B.; Himes, A.D.; Getz, D.R.; Raetzman, L.T. Notch signaling in postnatal pituitary expansion: Proliferation, progenitors, and cell specification. Mol. Endocrinol. 2014, 28, 731–744. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, M.; Yoshida, S.; Ueharu, H.; Chen, M.; Kato, T.; Kato, Y. PRRX1 and PRRX2 distinctively participate in pituitary organogenesis and cell supply system. Cell Tissue Res. 2014, 357, 323–335. [Google Scholar] [CrossRef] [PubMed]
- Ueharu, H.; Higuchi, M.; Nishimura, N.; Yoshida, S.; Shibuya, S.; Sensui, K.; Kato, T.; Kato, Y. Krüppel-like factor 6, KLF6, is expressed in the rat pituitary stem/progenitor cells and regulates PRRX2 gene. J. Reprod. Dev. 2014, 60, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Lamouille, S.; Xu, J.; Derynck, R. Molecular mechanisms of epithelial-mesenchymal transition. Nat. Rev. Mol. Cell Biol. 2014, 15, 178–196. [Google Scholar] [CrossRef] [PubMed]
- Vallin, J.; Thuret, R.; Giacomello, E.; Faraldo, M.M.; Thiery, J.P.; Broders, F. Cloning and characterization of three Xenopus slug promoters reveal direct regulation by Lef/β-catenin signaling. J. Biol. Chem. 2001, 276, 30350–30358. [Google Scholar] [CrossRef] [PubMed]
- Yook, J.I.; Li, X.Y.; Ota, I.; Hu, C.; Kim, H.S.; Kim, N.H.; Cha, S.Y.; Ryu, J.K.; Choi, Y.J.; Kim, J.; et al. A Wnt-Axin2-GSK3β cascade regulates snail1 activity in breast cancer cells. Nat. Cell Biol. 2006, 8, 1398–1406. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Lamouille, S.; Derynck, R. TGF-β-induced epithelial to mesenchymal transition. Cell Res. 2009, 19, 156–172. [Google Scholar] [CrossRef] [PubMed]
- Gheldof, A.; Hulpiau, P.; van Roy, F.; De Craene, B.; Berx, G. Evolutionary functional analysis and molecular regulation of the ZEB transcription factors. Cell. Mol. Life Sci. 2012, 69, 2527–2541. [Google Scholar] [CrossRef] [PubMed]
- Joseph, M.J.; Dangi-Garimella, S.; Shields, M.A.; Diamond, M.E.; Sun, L.; Koblinski, J.E.; Munshi, H.G. Slug is a downstream mediator of transforming growth factor-β1-induced matrix metalloproteinase-9 expression and invasion of oral cancer cells. J. Cell. Biochem. 2009, 108, 726–736. [Google Scholar] [CrossRef] [PubMed]
- Kulesa, P.M.; Gammill, L.S. Neural crest migration: Patterns, phases and signals. Dev. Biol. 2010, 344, 566–568. [Google Scholar] [CrossRef] [PubMed]
- Belmadani, A.; Tran, P.B.; Ren, D.; Assimacopoulos, S.; Grove, E.A.; Miller, R.J. The chemokine stromal cell-derived factor-1 regulates the migration of sensory neuron progenitors. J. Neurosci. 2005, 25, 3995–4003. [Google Scholar] [CrossRef] [PubMed]
- Kasemeier-Kulesa, J.C.; McLennan, R.; Romine, M.H.; Kulesa, P.M.; Lefcort, F. CXCR4 controls ventral migration of sympathetic precursor cells. J. Neurosci. 2010, 30, 13078–13088. [Google Scholar] [CrossRef] [PubMed]
- Cimadamore, F.; Fishwick, K.; Giusto, E.; Gnedeva, K.; Cattarossi, G.; Miller, A.; Pluchino, S.; Brill, L.M.; Bronner-Fraser, M.; Terskikh, A.V. Human ESC-derived neural crest model reveals a key role for SOX2 in sensory neurogenesis. Cell Stem Cell 2011, 8, 538–551. [Google Scholar] [CrossRef] [PubMed]
- Davis, S.W.; Mortensen, A.H.; Camper, S.A. Birthdating studies reshape models for pituitary gland cell specification. Dev. Biol. 2011, 352, 215–227. [Google Scholar] [CrossRef] [PubMed]
- Himes, A.D.; Raetzman, L.T. Premature differentiation and aberrant movement of pituitary cells lacking both Hes1 and Prop1. Dev. Biol. 2009, 325, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Gage, P.J.; Brinkmeier, M.L.; Scarlett, L.M.; Knapp, L.T.; Camper, S.A.; Mahon, K.A. The Ames dwarf gene, df, is required early in pituitary ontogeny for the extinction of Rpx transcription and initiation of lineage-specific cell proliferation. Mol. Endocrinol. 1996, 10, 1570–1581. [Google Scholar] [PubMed]
- Raetzman, L.T.; Ward, R.; Camper, S.A. Lhx4 and prop1 are required for cell survival and expansion of the pituitary primordia. Development 2002, 129, 4229–4239. [Google Scholar] [PubMed]
- Wu, W.; Cogan, J.D.; Pfaffle, R.W.; Dasen, J.S.; Frisch, H.; O’Connell, S.M.; Flynn, S.E.; Brown, M.R.; Mullis, P.E.; Parks, J.S.; et al. Mutations in prop1 cause familial combined pituitary hormone deficiency. Nat. Genet. 1998, 18, 147–149. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Crenshaw, E.B., III; Rawson, W.; Simmons, D.; Swanson, L.; Rosenfeld, M. Dwarf locus mutants lacking three pituitary cell types result from mutations in the POU-domain gene pit-1. Nature 1990, 347, 528–533. [Google Scholar] [CrossRef] [PubMed]
- Nasonkin, I.O.; Ward, R.D.; Raetzman, L.T.; Seasholtz, A.F.; Saunders, T.L.; Gillespie, P.J.; Camper, S.A. Pituitary hypoplasia and respiratory distress syndrome in Prop1 knockout mice. Hum. Mol. Genet. 2004, 13, 2727–2735. [Google Scholar] [CrossRef] [PubMed]
- Sornson, M.W.; Wu, W.; Dasen, J.S.; Flynn, S.E.; Norman, D.J.; O’Connell, S.M.; Gukovsky, I.; Carriere, C.; Ryan, A.K.; Miller, A.P.; et al. Pituitary lineage determination by the Prophet of Pit-1 homeodomain factor defective in Ames dwarfism. Nature 1996, 384, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Brinkmeier, M.L.; Davis, S.W.; Carninci, P.; MacDonald, J.W.; Kawai, J.; Ghosh, D.; Hayashizaki, Y.; Lyons, R.H.; Camper, S.A. Discovery of transcriptional regulators and signaling pathways in the developing pituitary gland by bioinformatic and genomic approaches. Genomics 2009, 93, 449–460. [Google Scholar] [CrossRef] [PubMed]
- Olson, L.E.; Tollkuhn, J.; Scafoglio, C.; Krones, A.; Zhang, J.; Ohgi, K.A.; Wu, W.; Taketo, M.M.; Kemler, R.; Grosschedl, R.; et al. Homeodomain-mediated beta-catenin-dependent switching events dictate cell-lineage determination. Cell 2006, 125, 593–605. [Google Scholar] [CrossRef] [PubMed]
- Cha, K.B.; Douglas, K.R.; Potok, M.A.; Liang, H.; Jones, S.N.; Camper, S.A. Wnt5a signaling affects pituitary gland shape. Mech. Dev. 2004, 121, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Potok, M.A.; Cha, K.B.; Hunt, A.; Brinkmeier, M.L.; Leitges, M.; Kispert, A.; Camper, S.A. Wnt signaling affects gene expression in the ventral diencephalon and pituitary gland growth. Dev. Dyn. 2008, 237, 1006–1020. [Google Scholar] [CrossRef] [PubMed]
- Niehrs, C. The complex world of wnt receptor signalling. Nat. Rev. Mol. Cell Biol. 2012, 13, 767–779. [Google Scholar] [CrossRef] [PubMed]
- Brabletz, T.; Jung, A.; Reu, S.; Porzner, M.; Hlubek, F.; Kunz-Schughart, L.A.; Knuechel, R.; Kirchner, T. Variable beta-catenin expression in colorectal cancers indicates tumor progression driven by the tumor environment. Proc. Natl. Acad. Sci. USA 2001, 98, 10356–10361. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Wang, J.; Ju, B.G.; Rosenfeld, M.G. Signaling and epigenetic regulation of pituitary development. Curr. Opin. Cell Biol. 2007, 19, 605–611. [Google Scholar] [CrossRef] [PubMed]
- Bergelson, J.M.; Cunningham, J.A.; Droguett, G.; Kurt-Jones, E.A.; Krithivas, A.; Hong, J.S.; Horwitz, M.S.; Crowell, R.L.; Finberg, R.W. Isolation of a common receptor for coxsackie b viruses and adenoviruses 2 and 5. Science 1997, 275, 1320–1323. [Google Scholar] [CrossRef] [PubMed]
- Bowles, K.R.; Gibson, J.; Wu, J.; Shaffer, L.G.; Towbin, J.A.; Bowles, N.E. Genomic organization and chromosomal localization of the human coxsackievirus b-adenovirus receptor gene. Hum. Genet. 1999, 105, 354–359. [Google Scholar] [CrossRef] [PubMed]
- Hotta, Y.; Honda, T.; Naito, M.; Kuwano, R. Developmental distribution of coxsackie virus and adenovirus receptor localized in the nervous system. Brain Res. Dev. Brain Res. 2003, 143, 1–13. [Google Scholar] [CrossRef]
- Vankelecom, H. Stem cells in the postnatal pituitary? Neuroendocrinology 2007, 85, 110–130. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, M.; Yatabe, M.; Kouki, T.; Fujiwara, K.; Takigami, S.; Sakamoto, A.; Yashiro, T. Changes in e- and n-cadherin expression in developing rat adenohypophysis. Anat. Rec. 2007, 290, 486–490. [Google Scholar] [CrossRef] [PubMed]
- Hafner, C.; Meyer, S.; Hagen, I.; Becker, B.; Roesch, A.; Landthaler, M.; Vogt, T. Ephrin-b reverse signaling induces expression of wound healing associated genes in IEC-6 intestinal epithelial cells. World J. Gastroenterol. 2005, 11, 4511–4518. [Google Scholar] [CrossRef] [PubMed]
- Nakada, M.; Anderson, E.M.; Demuth, T.; Nakada, S.; Reavie, L.B.; Drake, K.L.; Hoelzinger, D.B.; Berens, M.E. The phosphorylation of ephrin-B2 ligand promotes glioma cell migration and invasion. Int. J. Cancer 2010, 126, 1155–1165. [Google Scholar] [CrossRef] [PubMed]
- Meyer, S.; Hafner, C.; Guba, M.; Flegel, S.; Geissler, E.K.; Becker, B.; Koehl, G.E.; Orso, E.; Landthaler, M.; Vogt, T. Ephrin-B2 overexpression enhances integrin-mediated ECM-attachment and migration of B16 melanoma cells. Int. J. Oncol. 2005, 27, 1197–1206. [Google Scholar] [CrossRef] [PubMed]
- Cowan, C.A.; Henkemeyer, M. The SH2/SH3 adaptor Grb4 transduces B-ephrin reverse signals. Nature 2001, 413, 174–179. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, S.; Kato, T.; Chen, M.; Higuchi, M.; Ueharu, H.; Nishimura, N.; Kato, Y. Localization of a juxtacrine factor ephrin-B2 in the pituitary stem/progenitor cell niches throughout life. Cell Tissue Res. 2015, 359, 755–766. [Google Scholar] [CrossRef] [PubMed]
- Conboy, I.M.; Rando, T.A. The regulation of notch signaling controls satellite cell activation and cell fate determination in postnatal myogenesis. Dev. Cell 2002, 3, 397–409. [Google Scholar] [CrossRef]
- Fu, Q.; Gremeaux, L.; Luque, R.M.; Liekens, D.; Chen, J.; Buch, T.; Waisman, A.; Kineman, R.; Vankelecom, H. The adult pituitary shows stem/progenitor cell activation in response to injury and is capable of regeneration. Endocrinology 2012, 153, 3224–3235. [Google Scholar] [CrossRef] [PubMed]
- Fu, Q.; Vankelecom, H. Regenerative capacity of the adult pituitary: Multiple mechanisms of lactotroph restoration after transgenic ablation. Stem Cells Dev. 2012, 21, 3245–3257. [Google Scholar] [CrossRef] [PubMed]
- Horiguchi, K.; Fujiwara, K.; Tsukada, T.; Yako, H.; Tateno, K.; Hasegawa, R.; Takegami, S.; Osako, S.; Yashiro, T.; Kato, T.; et al. Expression of slug in s100β protein-positive cells of the postnatal developing rat anterior pituitary gland. Cell Tissue Res. 2015, 363, 513–524. [Google Scholar] [CrossRef] [PubMed]
- Horiguchi, K.; Ilmiawati, C.; Fujiwara, K.; Tsukada, T.; Kikuchi, M.; Yashiro, T. Expression of chemokine CXCL12 and its receptor CXCR4 in folliculostellate (fs) cells of the rat anterior pituitary gland: The CXCL12/CXCR4 axis induces interconnection of fs cells. Endocrinology 2012, 153, 1717–1724. [Google Scholar] [CrossRef] [PubMed]
- Nagasawa, T. CXC chemokine ligand 12 (CXCL12) and its receptor CXCR4. J. Mol. Med. 2014, 92, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Ara, T.; Tokoyoda, K.; Sugiyama, T.; Egawa, T.; Kawabata, K.; Nagasawa, T. Long-term hematopoietic stem cells require stromal cell-derived factor-1 for colonizing bone marrow during ontogeny. Immunity 2003, 19, 257–267. [Google Scholar] [CrossRef]
- Sugiyama, T.; Kohara, H.; Noda, M.; Nagasawa, T. Maintenance of the hematopoietic stem cell pool by CXCL12- CXCR4 chemokine signaling in bone marrow stromal cell niches. Immunity 2006, 25, 977–988. [Google Scholar] [CrossRef] [PubMed]
- Tokoyoda, K.; Egawa, T.; Sugiyama, T.; Choi, B.I.; Nagasawa, T. Cellular niches controlling b lymphocyte behavior within bone marrow during development. Immunity 2004, 20, 707–718. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Kim, J.M.; Lee, E.J. Functional expression of CXCR4 in somatotrophs: CXCL12 activates GH gene, GH production and secretion, and cellular proliferation. J. Endocrinol. 2008, 199, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Krylyshkina, O.; Chen, J.; Mebis, L.; Denef, C.; Vankelecom, H. Nestin-immunoreactive cells in rat pituitary are neither hormonal nor typical folliculo-stellate cells. Endocrinology 2005, 146, 2376–2387. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Lavandeira, M.; Diaz-Rodriguez, E.; Bahar, D.; Garcia-Rendueles, A.R.; Rodrigues, J.S.; Dieguez, C.; Alvarez, C.V. Pituitary cell turnover: From adult stem cell recruitment through differentiation to death. Neuroendocrinology 2015, 101, 175–192. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Gene Title | Species | Characteristic | References |
|---|---|---|---|---|
| Sox9 | SRY-box containing gene 9 | Mouse, rat | Expressed in both sides of the MCL, and the parenchyma of both AL and IL. | [11,12,42,43] |
| Prop1 | Paired like homeodomain factor 1 | Rat | Localized in about 50% of SOX2-positive cells in the adult rat pituitary. Notably, PROP1-positive cells are gradually decreased in the MCL, but maintained in the parenchyma. | [25,38] |
| Lhx3 | LIM homeobox protein 3 | Mouse | Localized in SOX2-positive cells of both the MCL and parenchyma, and also in SOX2-negative cells. | [42] |
| Prrx1 | Paired related homeobox 1 | Rat | Localized in both sides of MCL and parenchyma of AL, and in about 20% of SOX2-positive cells in the adult rat pituitary. Also in SOX2-negative cells. | [44] |
| Prrx2 | Paired related homeobox 2 | Rat | Localized in a very small population of SOX2-positive cells in both sides of the MCL only after P30 but not E20.5. | [44] |
| β-catenin | Ctnnb1 | Rat | Localized mainly in the MCL. | [42] |
| Hes1 | Hairy and enhancer of split 1 | Mouse | Expressed in the anterior side of MCL and parenchyma of AL. | [43] |
| Hey1 | Hairy/enhancer-of-split related with YRPW motif 1 | Mouse | Expressed in the both sides of MCL and parenchyma of AL. | [43] |
| Klf6 | Kruppel-like factor 6 | Rat | Localized in a very small population of SOX2-positive cells in the anterior side of MCL, but not in the parenchyma. | [45] |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoshida, S.; Kato, T.; Kato, Y. EMT Involved in Migration of Stem/Progenitor Cells for Pituitary Development and Regeneration. J. Clin. Med. 2016, 5, 43. https://doi.org/10.3390/jcm5040043
Yoshida S, Kato T, Kato Y. EMT Involved in Migration of Stem/Progenitor Cells for Pituitary Development and Regeneration. Journal of Clinical Medicine. 2016; 5(4):43. https://doi.org/10.3390/jcm5040043
Chicago/Turabian StyleYoshida, Saishu, Takako Kato, and Yukio Kato. 2016. "EMT Involved in Migration of Stem/Progenitor Cells for Pituitary Development and Regeneration" Journal of Clinical Medicine 5, no. 4: 43. https://doi.org/10.3390/jcm5040043