FBXL7 Upregulation Predicts a Poor Prognosis and Associates with a Possible Mechanism for Paclitaxel Resistance in Ovarian Cancer

 , , ,
, , ,
Abstract
:
1. Introduction
2. Experimental Section
2.1. Cell Culture
2.2. Reverse Transcriptase-Polymerase Chain Reaction (RT-PCR)
2.3. Cell Tranfection
2.4. MTT Assay
2.5. Microarray and RNA Sequencing Data Processing
2.6. Immunohistochemistry Staining Analysis
2.7. Kaplan-Meier Analyses
2.8. Univariate and Multivariate Analyses
2.9. In Silico Analysis
2.10. Statistical Analyses
3. Results
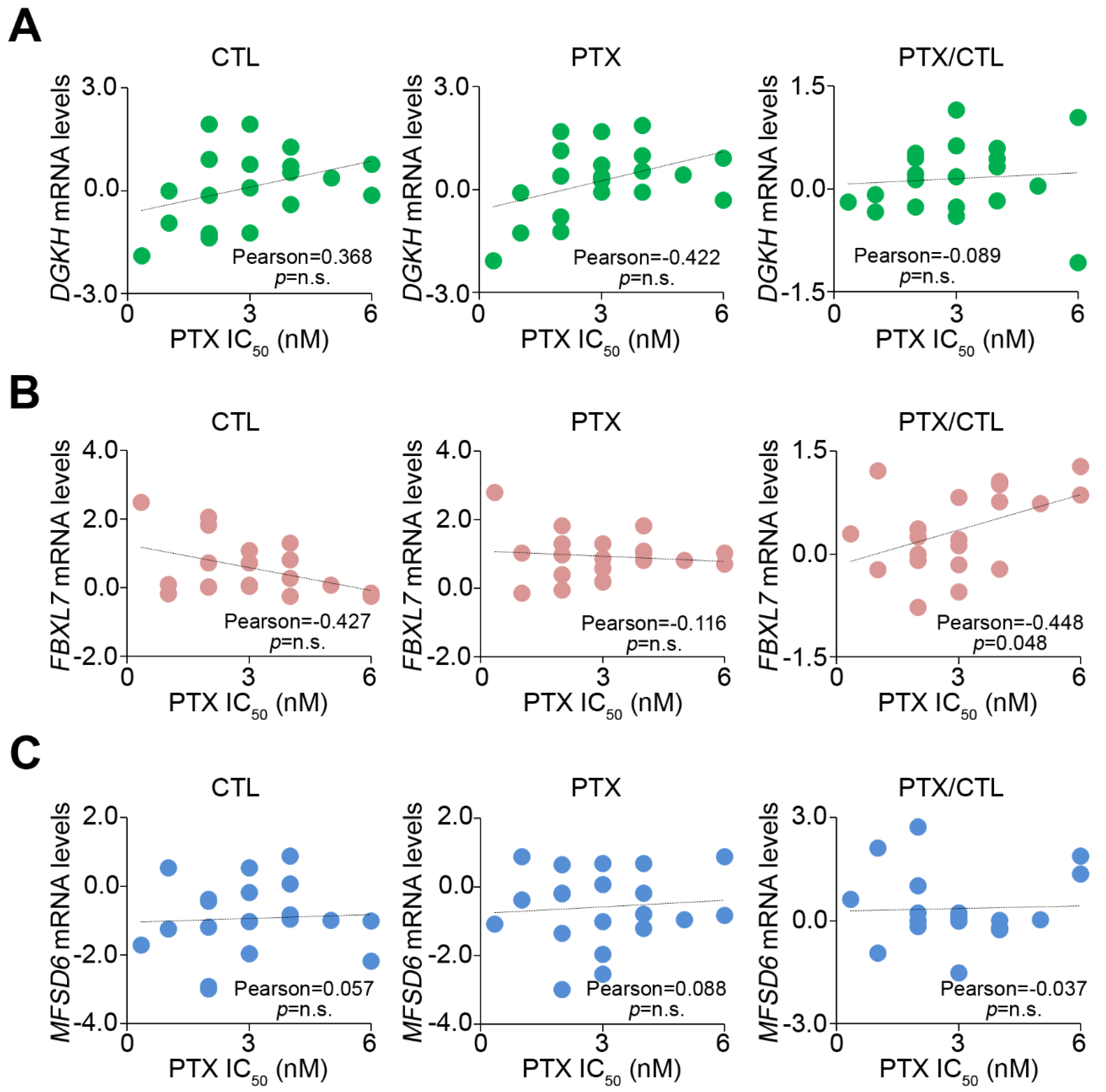
3.1. Upregulation of DGKH, FBXL7 and MFSD6 Is Involved in the Mechanism Underlying PTX Resistance
3.2. FBXL7 Upregulation Is Associated with Poor Progression-Free Survival (PFS) Rates in Ovarian Cancer Patients
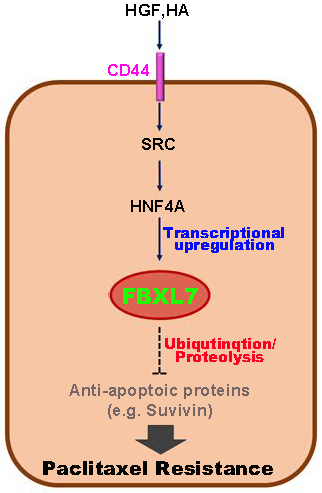
3.3. Possible Mechanism Underlying PTX Resistance in Ovarian Cancer Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Dahiya, N.; Morin, P.J. Micrornas in ovarian carcinomas. Endocr. Relat. Cancer 2010, 17, F77–F89. [Google Scholar] [CrossRef] [PubMed]
- De Wolf, E.; Abdullah, M.I.; Jones, S.M.; Menezes, K.; Moss, D.M.; Drijfhout, F.P.; Hart, S.R.; Hoskins, C.; Stronach, E.A.; Richardson, A. Dietary geranylgeraniol can limit the activity of pitavastatin as a potential treatment for drug-resistant ovarian cancer. Sci. Rep. 2017, 7, 5410. [Google Scholar] [CrossRef] [PubMed]
- Mihanfar, A.; Fattahi, A.; Nejabati, H.R. Microrna-mediated drug resistance in ovarian cancer. J. Cell. Physiol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Scartozzi, M.; Pistelli, M.; Bittoni, A.; Giampieri, R.; Galizia, E.; Berardi, R.; Faloppi, L.; Del Prete, M.; Cascinu, S. Novel perspectives for the treatment of gastric cancer: From a global approach to a personalized strategy. Curr. Oncol. Rep. 2010, 12, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Maryam, A.; Qazi, J.I.; Ma, T. Targeting apoptosis and multiple signaling pathways with icariside II in cancer cells. Int. J. Biol. Sci. 2015, 11, 1100–1112. [Google Scholar] [CrossRef] [PubMed]
- Qiao, L.; Wong, B.C. Targeting apoptosis as an approach for gastrointestinal cancer therapy. Drug Resist. Updates 2009, 12, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Kamran, M.; Long, Z.J.; Xu, D.; Lv, S.S.; Liu, B.; Wang, C.L.; Xu, J.; Lam, E.W.; Liu, Q. Aurora kinase a regulates survivin stability through targeting FBXL7 in gastric cancer drug resistance and prognosis. Oncogenesis 2017, 6, e298. [Google Scholar] [CrossRef] [PubMed]
- Bosch, J.A.; Sumabat, T.M.; Hafezi, Y.; Pellock, B.J.; Gandhi, K.D.; Hariharan, I.K. The drosophila f-box protein FBXL7 binds to the protocadherin fat and regulates dachs localization and hippo signaling. eLife 2014, 3, e03383. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Kon, M.; DeLisi, C. Pathway-based classification of cancer subtypes. Biol. Direct. 2012, 7, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues-Campos, M.; Thompson, B.J. The ubiquitin ligase FBXL7 regulates the dachsous-fat-dachs system in drosophila. Development 2014, 141, 4098–4103. [Google Scholar] [CrossRef] [PubMed]
- Hershko, A.; Ciechanover, A. The ubiquitin system. Annu. Rev. Biochem. 1998, 67, 425–479. [Google Scholar] [CrossRef] [PubMed]
- Cardozo, T.; Pagano, M. The SCF ubiquitin ligase: Insights into a molecular machine. Nat. Rev. Mol. Cell Biol. 2004, 5, 739–751. [Google Scholar] [CrossRef] [PubMed]
- Skaar, J.R.; Pagan, J.K.; Pagano, M. Mechanisms and function of substrate recruitment by f-box proteins. Nat. Rev. Mol. Cell Biol. 2013, 14, 369–381. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Lear, T.; Zhao, Y.; Zhao, J.; Zou, C.; Chen, B.B.; Mallampalli, R.K. F-box protein fbxl18 mediates polyubiquitylation and proteasomal degradation of the pro-apoptotic scf subunit fbxl7. Cell Death Dis. 2015, 6, e1630. [Google Scholar] [CrossRef] [PubMed]
- Kathawala, R.J.; Gupta, P.; Ashby, C.R., Jr.; Chen, Z.S. The modulation of abc transporter-mediated multidrug resistance in cancer: A review of the past decade. Drug Resist. Updates 2015, 18, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Majidinia, M.; Alizadeh, E.; Yousefi, B.; Akbarzadeh, M.; Zarghami, N. Downregulation of notch signaling pathway as an effective chemosensitizer for cancer treatment. Drug Res. 2016, 66, 571–579. [Google Scholar] [CrossRef] [PubMed]
- Baumeister, W.; Walz, J.; Zuhl, F.; Seemuller, E. The proteasome: Paradigm of a self-compartmentalizing protease. Cell 1998, 92, 367–380. [Google Scholar] [CrossRef]
- Zhang, W.; Sidhu, S.S. Development of inhibitors in the ubiquitination cascade. FEBS Lett. 2014, 588, 356–367. [Google Scholar] [CrossRef] [PubMed]
- Matsuura, K.; Huang, N.J.; Cocce, K.; Zhang, L.; Kornbluth, S. Downregulation of the proapoptotic protein moap-1 by the ubr5 ubiquitin ligase and its role in ovarian cancer resistance to cisplatin. Oncogene 2017, 36, 1698–1706. [Google Scholar] [CrossRef] [PubMed]
- MacKay, C.; Carroll, E.; Ibrahim, A.F.M.; Garg, A.; Inman, G.J.; Hay, R.T.; Alpi, A.F. E3 ubiquitin ligase hoip attenuates apoptotic cell death induced by cisplatin. Cancer Res. 2014, 74, 2246–2257. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Pankratz, V.S.; Fredericksen, Z.; Tarrell, R.; Karaus, M.; McGuffog, L.; Pharaoh, P.D.; Ponder, B.A.; Dunning, A.M.; Peock, S.; et al. Common variants associated with breast cancer in genome-wide association studies are modifiers of breast cancer risk in brca1 and brca2 mutation carriers. Hum. Mol. Genet. 2010, 19, 2886–2897. [Google Scholar] [CrossRef] [PubMed]
- King, M.C.; Marks, J.H.; Mandell, J.B. Breast and ovarian cancer risks due to inherited mutations in brca1 and brca2. Science 2003, 302, 643–646. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Lear, T.; Iannone, O.; Shiva, S.; Corey, C.; Rajbhandari, S.; Jerome, J.; Chen, B.B.; Mallampalli, R.K. The proapoptotic f-box protein fbxl7 regulates mitochondrial function by mediating the ubiquitylation and proteasomal degradation of survivin. J. Biol. Chem. 2015, 290, 11843–11852. [Google Scholar] [CrossRef] [PubMed]
- Apte, S.M.; Bucana, C.D.; Killion, J.J.; Gershenson, D.M.; Fidler, I.J. Expression of platelet-derived growth factor and activated receptor in clinical specimens of epithelial ovarian cancer and ovarian carcinoma cell lines. Gynecol. Oncol. 2004, 93, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Dabrow, M.B.; Francesco, M.R.; McBrearty, F.X.; Caradonna, S. The effects of platelet-derived growth factor and receptor on normal and neoplastic human ovarian surface epithelium. Gynecol. Oncol. 1998, 71, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Henriksen, R.; Funa, K.; Wilander, E.; Backstrom, T.; Ridderheim, M.; Oberg, K. Expression and prognostic significance of platelet-derived growth factor and its receptors in epithelial ovarian neoplasms. Cancer Res. 1993, 53, 4550–4554. [Google Scholar] [PubMed]
- Naor, D.; Sionov, R.V.; Ish-Shalom, D. Cd44: Structure, function, and association with the malignant process. Adv. Cancer Res. 1997, 71, 241–319. [Google Scholar] [PubMed]
- Mali, A.V.; Joshi, A.A.; Hegde, M.V.; Kadam, S.S. Enterolactone modulates the erk/nf-kappab/snail signaling pathway in triple-negative breast cancer cell line mda-mb-231 to revert the tgf-beta-induced epithelial-mesenchymal transition. Cancer Biol. Med. 2018, 15, 137–156. [Google Scholar] [PubMed]
- Shah, V.; Taratula, O.; Garbuzenko, O.B.; Taratula, O.R.; Rodriguez-Rodriguez, L.; Minko, T. Targeted nanomedicine for suppression of cd44 and simultaneous cell death induction in ovarian cancer: An optimal delivery of sirna and anticancer drug. Clin. Cancer Res. 2013, 19, 6193–6204. [Google Scholar] [CrossRef] [PubMed]
- Miletti-Gonzalez, K.E.; Chen, S.; Muthukumaran, N.; Saglimbeni, G.N.; Wu, X.; Yang, J.; Apolito, K.; Shih, W.J.; Hait, W.N.; Rodriguez-Rodriguez, L. The cd44 receptor interacts with p-glycoprotein to promote cell migration and invasion in cancer. Cancer Res. 2005, 65, 6660–6667. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Rodriguez, L.; Sancho-Torres, I.; Mesonero, C.; Gibbon, D.G.; Shih, W.J.; Zotalis, G. The cd44 receptor is a molecular predictor of survival in ovarian cancer. Med. Oncol. 2003, 20, 255–263. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Recurrence-Free Survival (n = 578) | ||||
|---|---|---|---|---|
| Variables | Crude HR (95% CI) | p | Adjusted HR (95% CI) | p |
| Age | ||||
| 59< | 1 | NA | 1 | NA |
| 59 | 1.00 (0.79–1.26) | 0.987 | 1.00 (0.79–1.26) | 0.987 |
| Pathologic stage | ||||
| IA-IIIB | 1 | NA | 1 | NA |
| IIIC-IV | 1.76 (1.22–2.53) | 0.002 | 1.69 (1.17–2.45) | 0.005 |
| Histologic grade | ||||
| G1–G2 | 1 | NA | 1 | NA |
| G3–G4 | 1.30 (0.94–1.79) | 0.109 | 1.25 (0.91–1.74) | 0.174 |
| FBXL7 expression | ||||
| low | 1 | NA | 1 | NA |
| high | 1.52 (1.14–2.04) | 0.005 | 1.51 (1.13–2.02) | 0.006 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiu, H.-W.; Chang, J.-S.; Lin, H.-Y.; Lee, H.-H.; Kuei, C.-H.; Lin, C.-H.; Huang, H.-M.; Lin, Y.-F. FBXL7 Upregulation Predicts a Poor Prognosis and Associates with a Possible Mechanism for Paclitaxel Resistance in Ovarian Cancer. J. Clin. Med. 2018, 7, 330. https://doi.org/10.3390/jcm7100330
Chiu H-W, Chang J-S, Lin H-Y, Lee H-H, Kuei C-H, Lin C-H, Huang H-M, Lin Y-F. FBXL7 Upregulation Predicts a Poor Prognosis and Associates with a Possible Mechanism for Paclitaxel Resistance in Ovarian Cancer. Journal of Clinical Medicine. 2018; 7(10):330. https://doi.org/10.3390/jcm7100330
Chicago/Turabian StyleChiu, Hui-Wen, Jeng-Shou Chang, Hui-Yu Lin, Hsun-Hua Lee, Chia-Hao Kuei, Che-Hsuan Lin, Huei-Mei Huang, and Yuan-Feng Lin. 2018. "FBXL7 Upregulation Predicts a Poor Prognosis and Associates with a Possible Mechanism for Paclitaxel Resistance in Ovarian Cancer" Journal of Clinical Medicine 7, no. 10: 330. https://doi.org/10.3390/jcm7100330