Fenofibrate Facilitates Post-Active Tuberculosis Infection in Macrophages and is Associated with Higher Mortality in Patients under Long-Term Treatment

 , , and
, , and 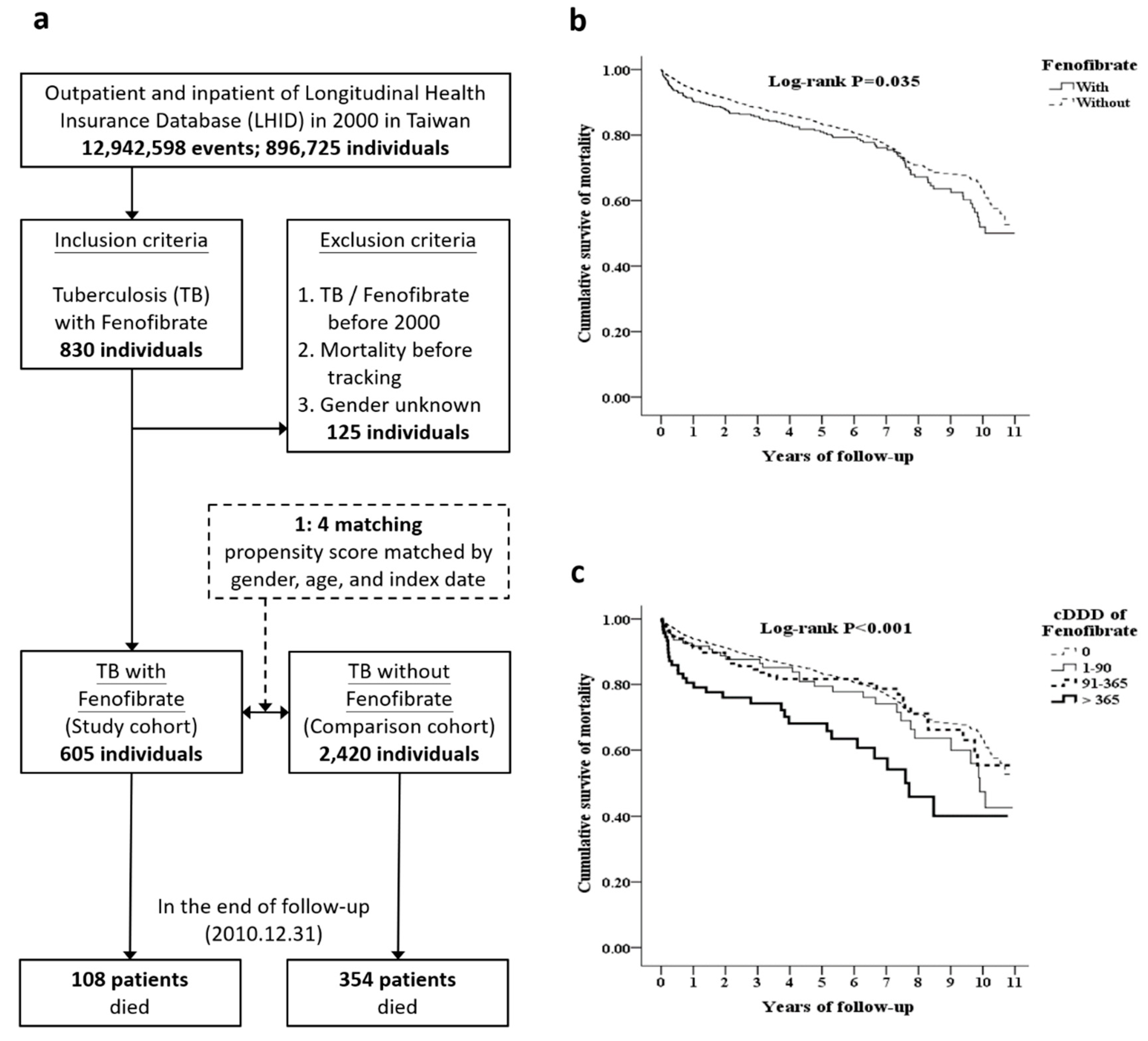{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
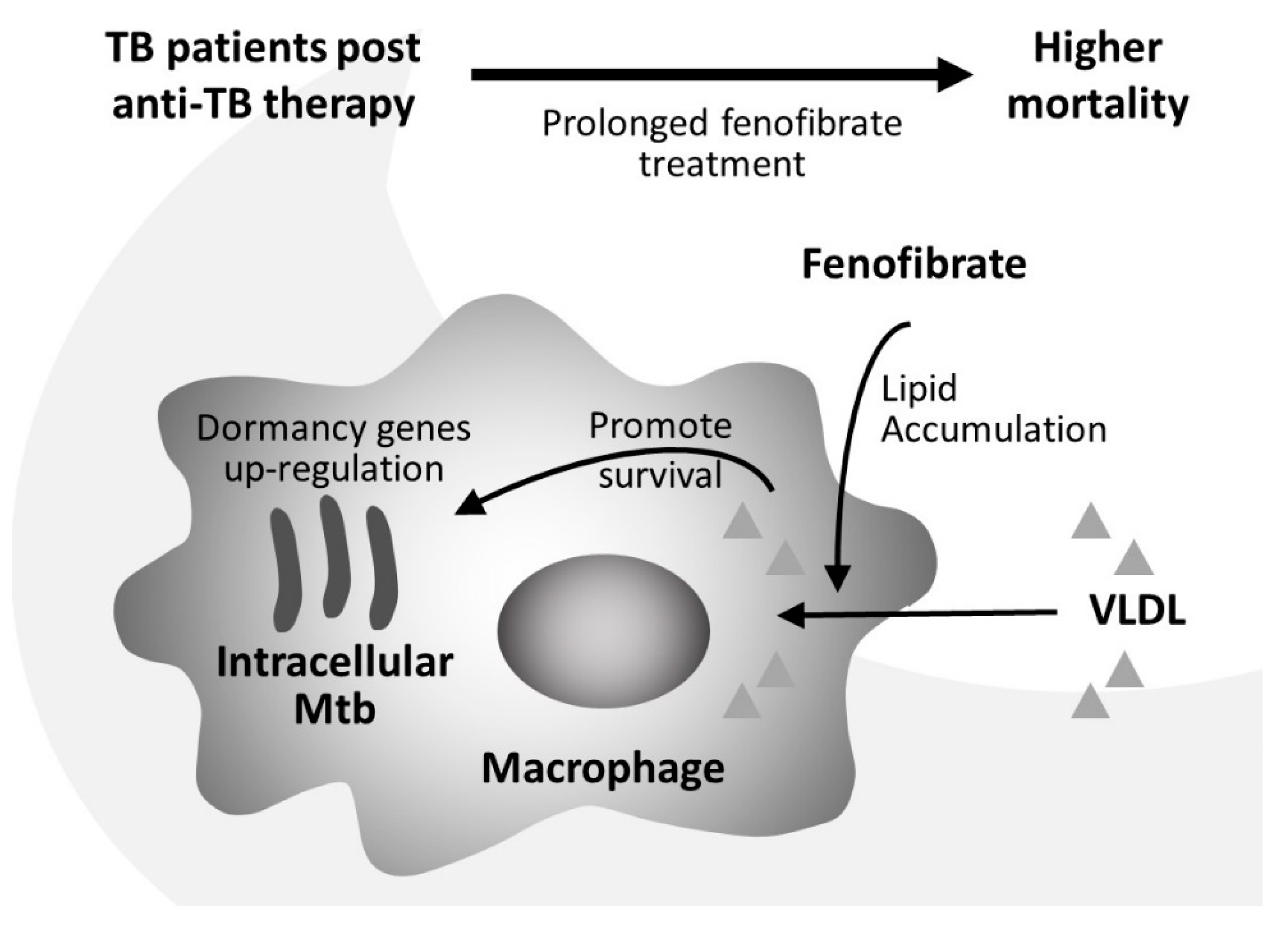
Abstract
:1. Introduction
2. Materials and Methods
2.1. Epidemiological Survey of Outcomes of TB Patients Treated with Fenofibrate
2.2. Cell Lines and Culture
2.3. Mycobacterium Tuberculosis Culture
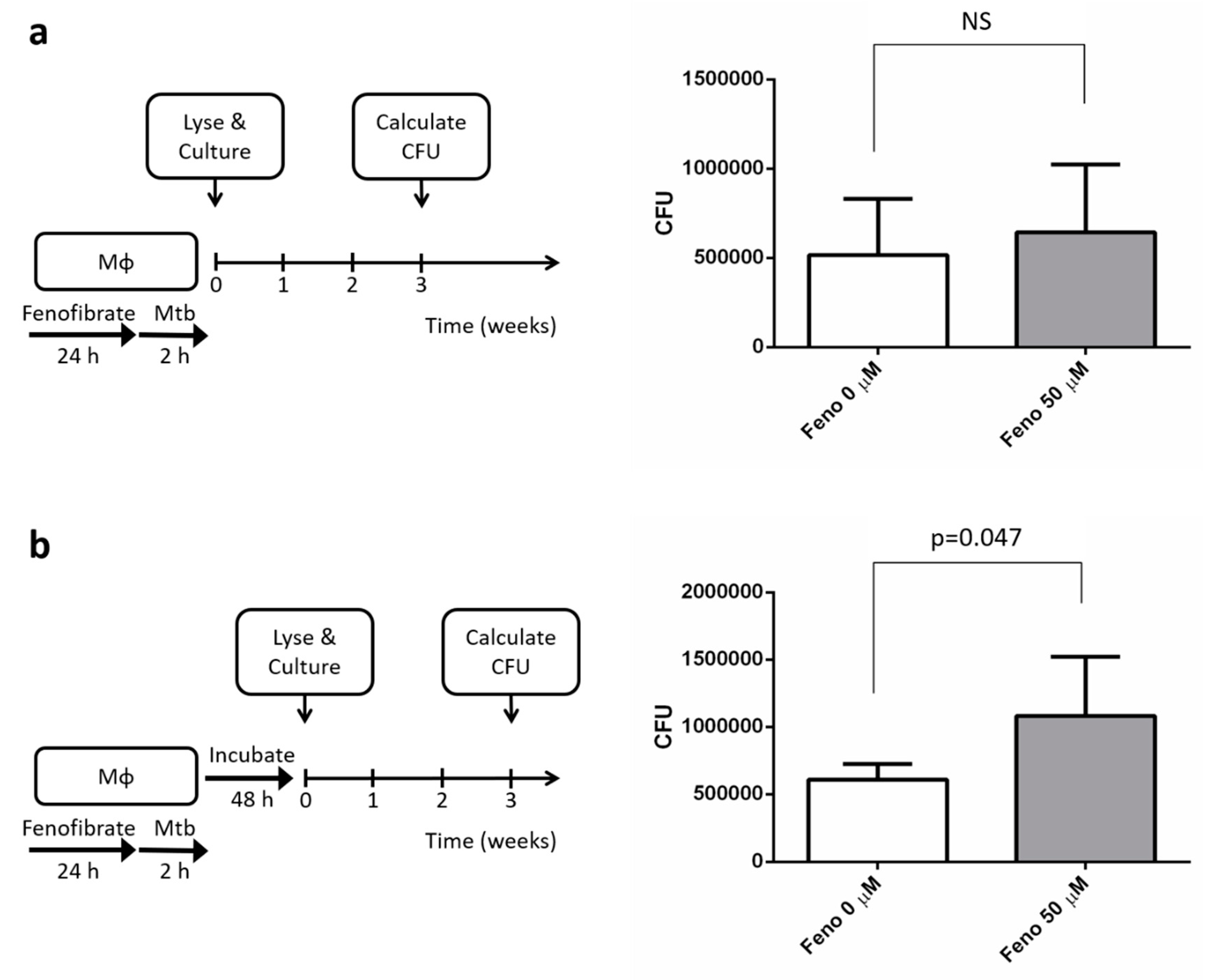
2.4. Macrophage Infection and Mtb Colony-Forming Units
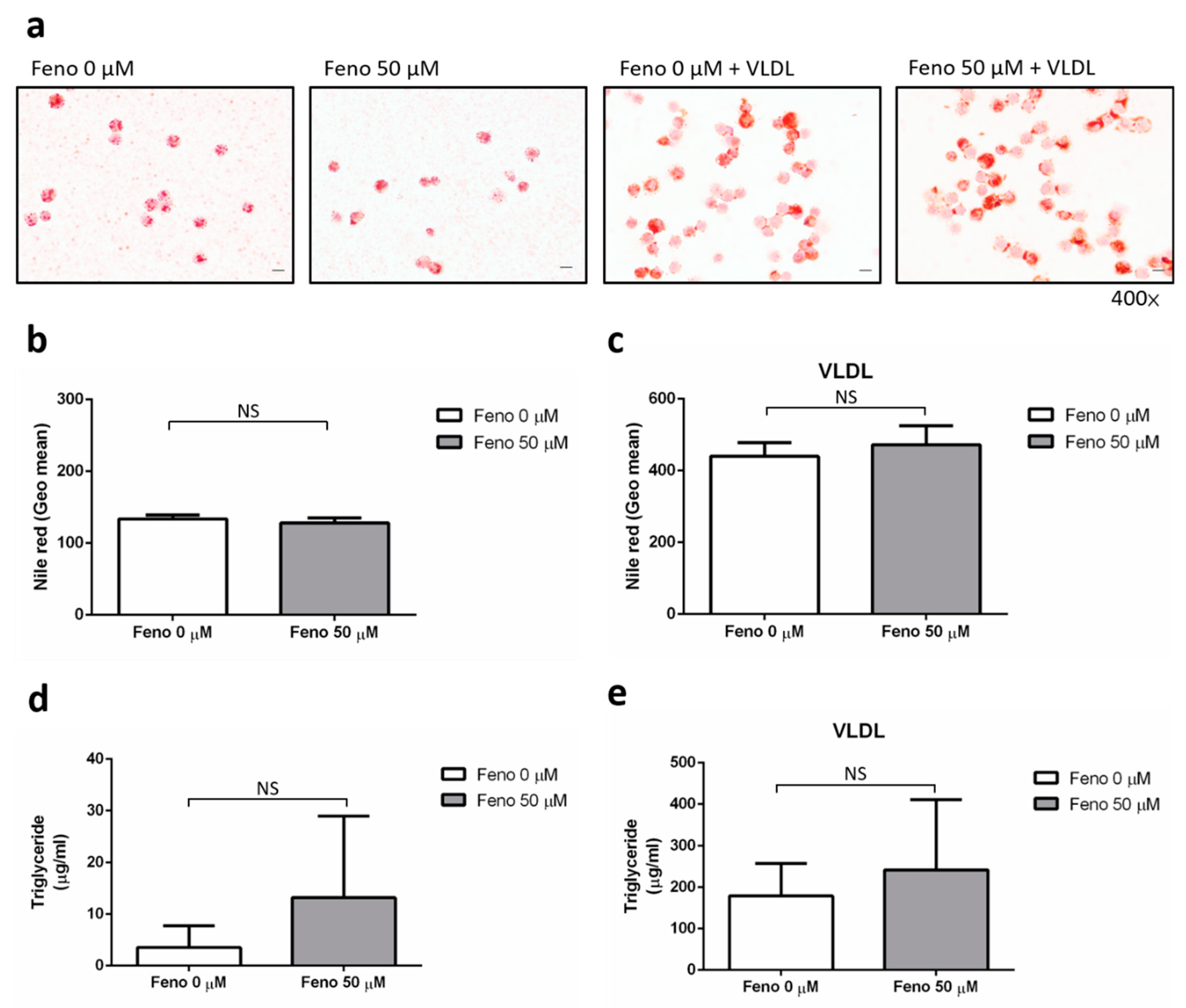
2.5. Oil Red O Staining
2.6. Nile Red Staining
2.7. Triglyceride colorimetric Assay
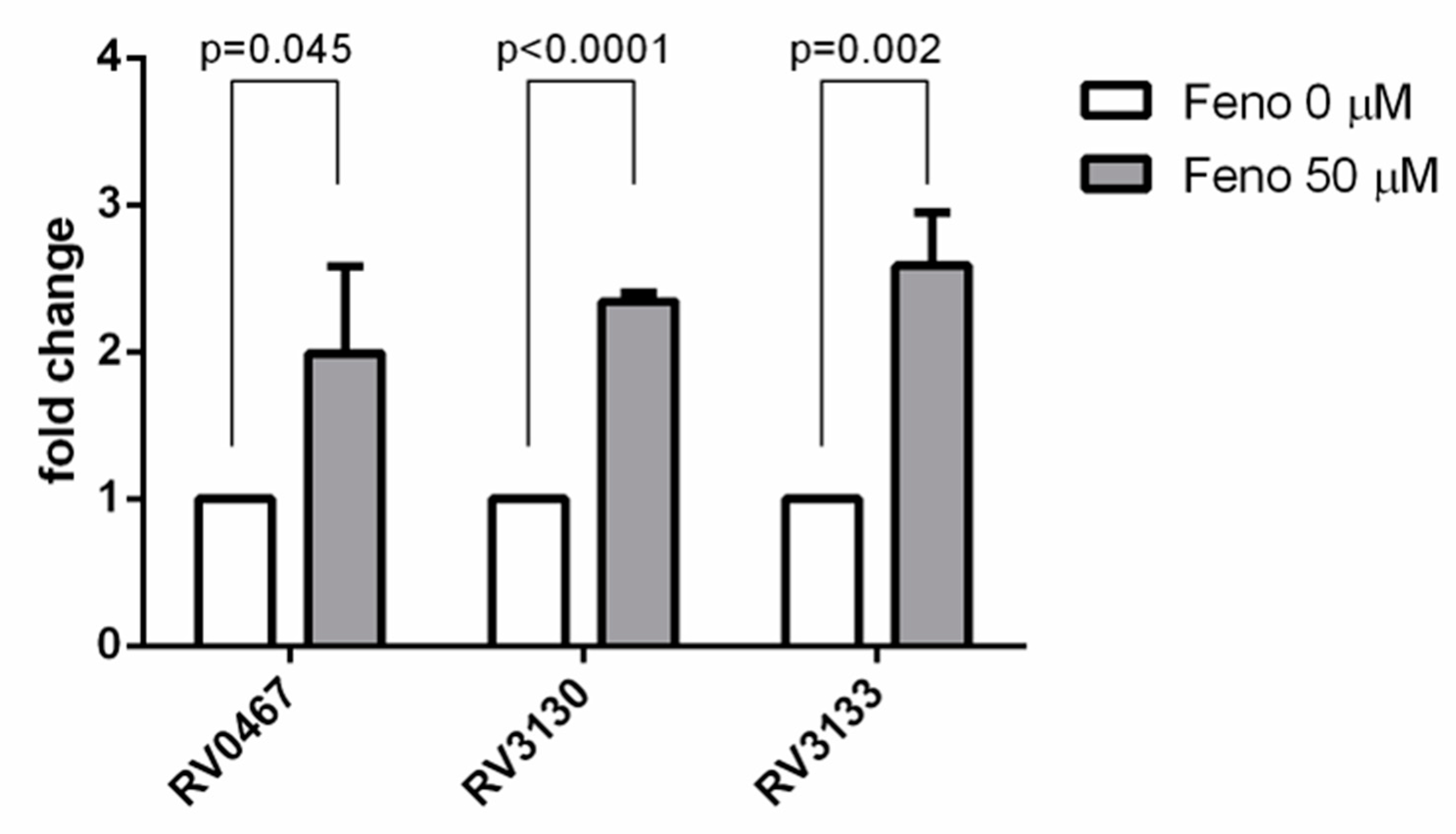
2.8. RNA Isolation and Quantitative Reverse Transcription PCR (qRT-PCR)
2.9. Statistical Analysis
3. Ethics Statement
4. Results
4.1. Fenofibrate Therapy is Associated with Adverse Outcomes in Patients with TB
4.2. The Effect of Fenofibrate on the Intracellular Lipid Content of Human Macrophages
4.3. Treatment with Fenofibrate Increased Mtb’s Intracellular Growth
4.4. The Effect of Fenofibrate on the Expression of Dormant, Intracellular Genes of Mtb
5. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- World Health Organization. Global Tuberculosis Report 2019. Available online: https://www.who.int/tb/publications/global_report/en/ (accessed on 21 October 2019).
- Taiwan CDC. Statistics of Tuberculosis in Taiwan. Available online: https://daily.cdc.gov.tw/stoptb/CareMagChart.aspx (accessed on 21 October 2019).
- Ramakrishnan, L. Revisiting the role of the granuloma in tuberculosis. Nat. Rev. Immunol. 2012, 12, 352–366. [Google Scholar] [CrossRef] [PubMed]
- Cadena, A.M.; Fortune, S.M.; Flynn, J.L. Heterogeneity in tuberculosis. Nat. Rev. Immunol. 2017, 17, 691–702. [Google Scholar] [CrossRef] [PubMed]
- Peddireddy, V.; Doddam, S.N.; Ahmed, N. Mycobacterial dormancy systems and host responses in tuberculosis. Front. Immunol. 2017, 8, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peyron, P.; Vaubourgeix, J.; Poquet, Y.; Levillain, F.; Botanch, C.; Bardou, F.; Daffe, M.; Emile, J.F.; Marchou, B.; Cardona, P.J.; et al. Foamy macrophages from tuberculous patients’ granulomas constitute a nutrient-rich reservoir for M. tuberculosis persistence. PLoS Pathog 2008, 4, e1000204. [Google Scholar] [CrossRef] [PubMed]
- Santucci, P.; Bouzid, F.; Smichi, N.; Poncin, I.; Kremer, L.; De Chastellier, C.; Drancourt, M.; Canaan, S. Experimental Models of Foamy Macrophages and Approaches for Dissecting the Mechanisms of Lipid Accumulation and Consumption during Dormancy and Reactivation of Tuberculosis. Front. Cell. Infect. Microbiol. 2016, 6, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reich-Slotky, R.; Kabbash, C.A.; Della-Latta, P.; Blanchard, J.S.; Feinmark, S.J.; Freeman, S.; Kaplan, G.; Shuman, H.A.; Silverstein, S.C. Gemfibrozil inhibits Legionella pneumophila and Mycobacterium tuberculosis enoyl coenzyme A reductases and blocks intracellular growth of these bacteria in macrophages. J. Bacteriol. 2009, 191, 5262–5271. [Google Scholar] [CrossRef] [Green Version]
- Soto-Ramirez, M.D.; Aguilar-Ayala, D.A.; Garcia-Morales, L.; Rodriguez-Peredo, S.M.; Badillo-Lopez, C.; Rios-Muñiz, D.E.; Mario, A.M.; Rivera-Morales, G.Y.; Rivera-Gutierrez, S.; Leon-Solis, L.; et al. Cholesterol plays a larger role during Mycobacterium tuberculosis in vitro dormancy and reactivation than previously suspected. Tuberculosis 2017, 103, 1–9. [Google Scholar] [CrossRef]
- Baures, P.W. Is RORγ a therapeutic target for treating Mycobacterium tuberculosis infections? Tuberculosis 2012, 92, 95–99. [Google Scholar] [CrossRef]
- Braissant, O.; Foufelle, F.; Scotto, C.; Dauça, M.; Wahli, W. Differential expression of peroxisome proliferator-activated receptors (PPARs): Tissue distribution of PPAR-alpha, -beta, -gamma in the adult rat. Endocrinology 1996, 137, 354–366. [Google Scholar] [CrossRef] [Green Version]
- Chinetti, G.; Griglio, S.; Antonucci, M.; Torra, I.P.; Delerive, P.; Majd, Z.; Fruchart, J.-C.; Chapman, J.; Najib, J.; Staels, B. Activation of proliferator activated receptors α and γ induces apoptosis of human monocyte-derived macrophages. J. Biol. Chem. 1998, 273, 25573–25580. [Google Scholar] [CrossRef] [Green Version]
- Staels, B.; Dallongeville, J.; Auwerx, J.; Schoonjans, K.; Leitersdorf, E.; Fruchart, J.C. Mechanism of action of fibrates on lipid and lipoprotein metabolism. Circulation 1998, 98, 2088–2093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerre-Millo, M.; Gervois, P.; Raspé, E.; Madsen, L.; Poulain, P.; Derudas, B.; Herbert, J.-M.; Winegar, D.A.; Willson, T.M.; Fruchart, J.-C.; et al. Peroxisome proliferator-activated receptor alpha activators improve insulin sensitivity and reduce adiposity. J. Biol. Chem. 2000, 275, 16638–16642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, C.F.; Chang, Y.H.; Chien, S.C.; Lin, Y.H.; Yeh, H.Y. Epidemiology of Dyslipidemia in the Asia Pacific Region. Int. J. Gerontol. 2018, 12, 2–6. [Google Scholar] [CrossRef]
- Saxena, A.; Srivastava, V.; Srivastava, R.; Srivastava, B.S. Identification of genes of Mycobacterium tuberculosis upregulated during anaerobic persistence by fluorescence and kanamycin resistance selection. Tuberculosis 2008, 88, 518–525. [Google Scholar] [CrossRef]
- Kumar, A.; Farhana, A.; Guidry, L.; Saini, V.; Hondalus, M.; Steyn, A.J. Redox homeostasis in mycobacteria: The key to tuberculosis control? Expert. Rev. Mol. Med. 2011, 13, e39. [Google Scholar] [CrossRef] [Green Version]
- Sherman, D.R.; Voskuil, M.; Schnappinger, D.; Liao, R.; Harrell, M.; Schoolnik, G. Regulation of the Mycobacterium tuberculosis hypoxic response gene encoding α-crystallin. Proc. Natl. Acad. Sci. USA 2001, 98, 7534–7539. [Google Scholar] [CrossRef] [Green Version]
- Hsueh, P.R.; Liu, Y.C.; So, J.; Liu, C.Y.; Yang, P.C.; Luh, K.T. Mycobacterium tuberculosis in Taiwan. J. Infect. 2006, 52, 77–85. [Google Scholar] [CrossRef]
- Yin, W.H.; Wu, C.C.; Chen, J.W. Registry of Lipid Control and the Use of Lipid-lowering Drugs for Secondary Prevention of Cardiovascular Events in Patients with Established Atherosclerotic Disease in Taiwan: Rationality and Methods. Int. J. Gerontol. 2012, 6, 241–246. [Google Scholar] [CrossRef] [Green Version]
- Kiran, D.; Podell, B.K.; Chambers, M.; Basaraba, R.J. Host-directed therapy targeting the Mycobacterium tuberculosis granuloma: A review. Semin. Immunopathol. 2016, 38, 167–183. [Google Scholar] [CrossRef] [Green Version]
- Brzostek, A.; Pawelczyk, J.; Rumijowska-Galewicz, A.; Dziadek, B.; Dziadek, J. Mycobacterium tuberculosis is able to accumulate and utilize cholesterol. J. Bacteriol. 2009, 191, 6584–6591. [Google Scholar] [CrossRef] [Green Version]
- Lovewell, R.R.; Sassetti, C.M.; VenderVen, B.C. Chewing the fat: Lipid metabolism and homeostasis during M. tuberculosis infection. Curr. Opin. Microbiol. 2016, 29, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Martens, G.W.; Arikan, M.C.; Lee, J.; Ren, F.; Vallerskog, T.; Kornfeld, H. Hypercholesterolemia impairs immunity to tuberculosis. Infect. Immun. 2008, 76, 3464–3472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deniz, O.; Gumus, S.; Yaman, H.; Ciftci, F.; Ors, F.; Cakir, E.; Tozkoparan, E.; Bilgic, H.; Ekiz, K. Serum total cholesterol, HDL-C and LDL-C concentrations significantly correlate with the radiological extent of disease and the degree of smear positivity in patients with pulmonary tuberculosis. Clin. Biochem. 2007, 40, 162–166. [Google Scholar] [CrossRef] [PubMed]
- Ravnskov, U. High cholesterol may protect against infections and atherosclerosis. Int. J. Med. 2003, 96, 927–934. [Google Scholar] [CrossRef]
- Pérez-Guzmán, C.; Vargas, M.H.; Quiñonez, F.; Bazavilvazo, N.; Aguilar, A. A cholesterol-rich diet accelerates bacteriologic sterilization in pulmonary tuberculosis. Chest 2005, 127, 643–651. [Google Scholar] [CrossRef] [Green Version]
- Su, V.Y.F.; Su, W.J.; Yen, Y.F.; Pan, S.W.; Chuang, P.H.; Feng, J.Y.; Chou, K.T.; Yang, K.Y.; Lee, Y.C.; Chen, T.J. Statin Use is Associated with a Lower Risk of Tuberculosis. Chest 2017, 152, 598–606. [Google Scholar] [CrossRef] [PubMed]
- Ho, H.T.; Tsai, I.F.; Wu, C.L.; Lu, Y.T. Aminopeptidase N facilitates entry and intracellular survival of Mycobacterium tuberculosis in monocytes. Respirology 2014, 19, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Kuo, C.P.; Chang, K.S.; Hsu, J.L.; Tsai, I.F.; Lin, A.B.; Wei, T.Y.; Wu, C.L.; Lu, Y.T. Analysis of the immune response of human dendritic cells to Mycobacterium tuberculosis by quantitative proteomics. Proteome Sci. 2016, 14, 5. [Google Scholar] [CrossRef] [Green Version]
- Tsai, I.F.; Kuo, C.P.; Lin, A.B.; Chien, M.N.; Ho, H.T.; Wei, T.Y.; Wu, C.L.; Lu, Y.T. Potential effect of ezetimibe against Mycobacterium tuberculosis infecting in type II diabetes. Respirology 2017, 22, 559–566. [Google Scholar] [CrossRef]
- Shu, C.C.; Lee, C.H.; Lee, M.C.; Wang, J.Y.; Yu, C.J.; Lee, L.N. Hepatotoxicity due to first-line anti-tuberculosis drugs: A five-year experience in a Taiwan medical centre. Int. J. Tuberc. Lung Dis. 2013, 17, 934–939. [Google Scholar] [CrossRef]
- Ahmad, J.; Odin, J.A.; Hayashi, P.H.; Chalasani, N.; Fontana, R.J.; Barnhart, H.; Cirulli, E.T.; Kleiner, D.E.; Hoofnagle, J.H. Identification and Characterization of Fenofibrate-Induced Liver Injury. Dig. Dis. Sci. 2017, 62, 3596–3604. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Lee, H.M.; Kim, J.K.; Yang, C.S.; Kim, T.S.; Jung, M.; Jin, H.S.; Kim, S.; Jang, J.; Kim, J.M.; et al. PPAR-α Activation Mediates Innate Host Defense through Induction of TFEB and Lipid Catabolism. J. Immunol. 2017, 198, 3283–3295. [Google Scholar] [CrossRef] [PubMed] [Green Version]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, C.-L.; Lu, Y.-T.; Tsai, I.-F.; Wu, L.-C.; Chien, W.-C.; Chung, C.-H.; Ma, K.-H. Fenofibrate Facilitates Post-Active Tuberculosis Infection in Macrophages and is Associated with Higher Mortality in Patients under Long-Term Treatment. J. Clin. Med. 2020, 9, 337. https://doi.org/10.3390/jcm9020337
Liu C-L, Lu Y-T, Tsai I-F, Wu L-C, Chien W-C, Chung C-H, Ma K-H. Fenofibrate Facilitates Post-Active Tuberculosis Infection in Macrophages and is Associated with Higher Mortality in Patients under Long-Term Treatment. Journal of Clinical Medicine. 2020; 9(2):337. https://doi.org/10.3390/jcm9020337
Chicago/Turabian StyleLiu, Ching-Lung, Yen-Ta Lu, I-Fan Tsai, Ling-Chiao Wu, Wu-Chien Chien, Chi-Hsiang Chung, and Kuo-Hsing Ma. 2020. "Fenofibrate Facilitates Post-Active Tuberculosis Infection in Macrophages and is Associated with Higher Mortality in Patients under Long-Term Treatment" Journal of Clinical Medicine 9, no. 2: 337. https://doi.org/10.3390/jcm9020337