
Biodiversity of Upwelling Coastal Systems of the Southern Caribbean Sea Adjacent to Guajira Peninsula

Abstract
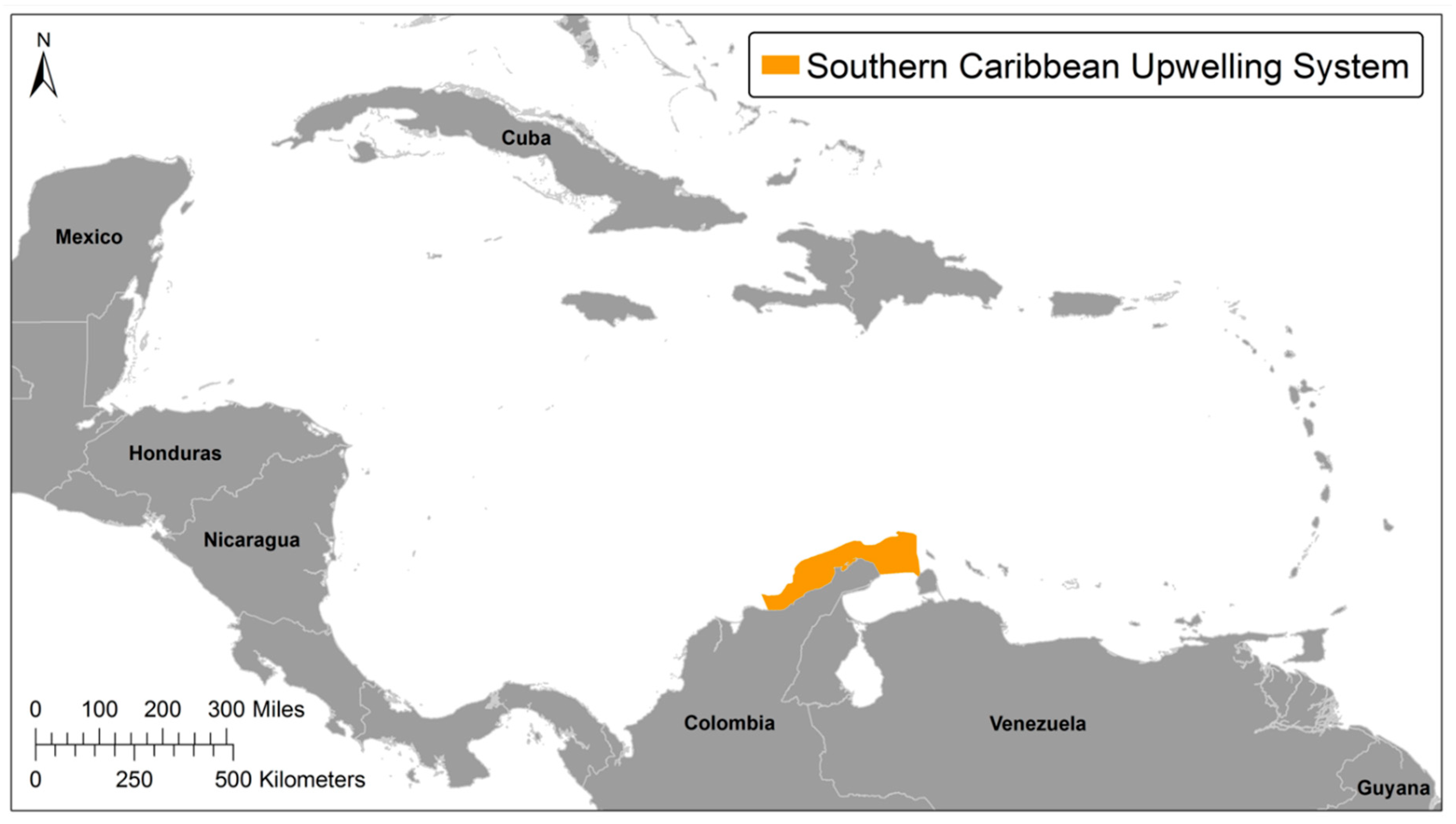
:1. Introduction
2. The Biodiversity of Coastal Guajira
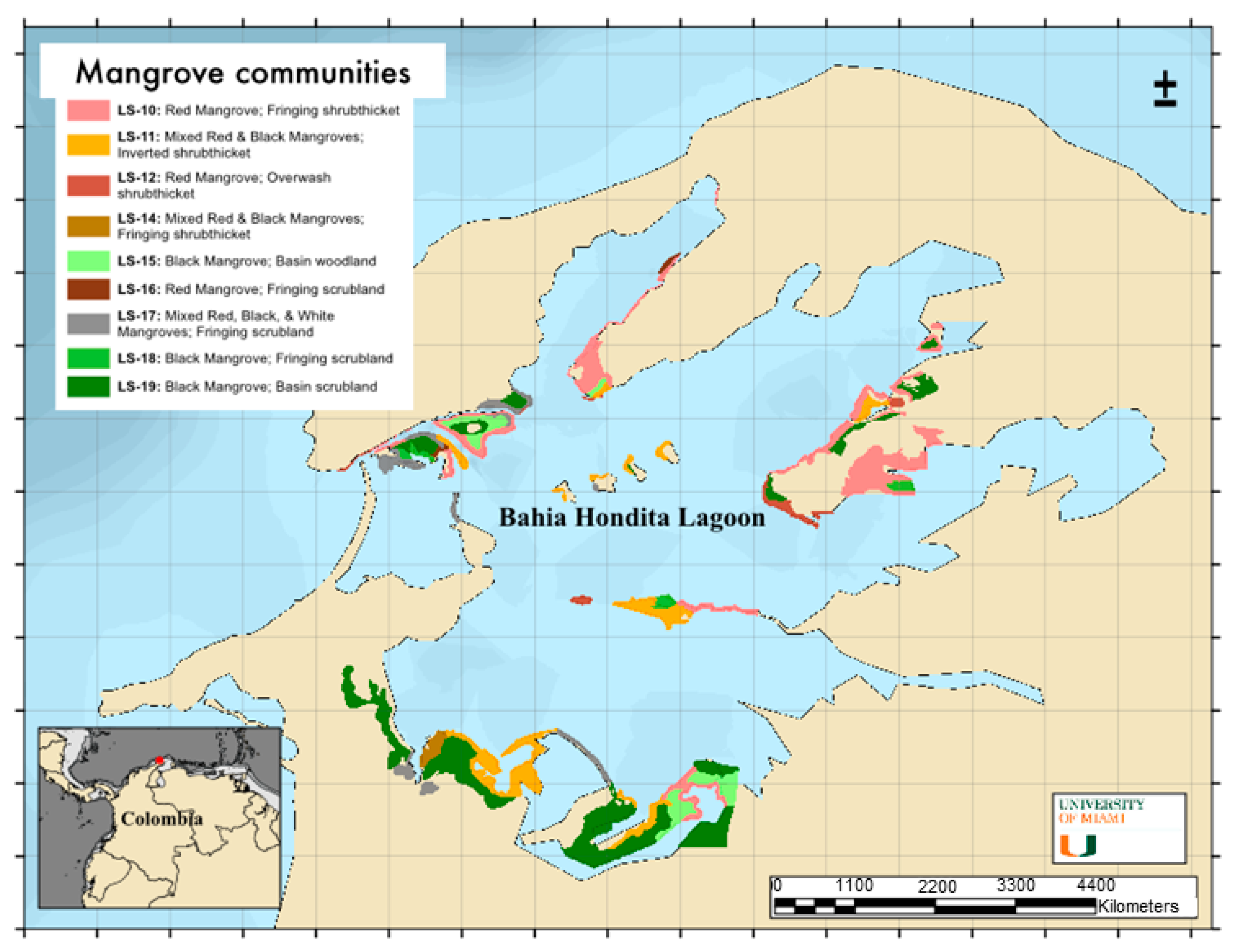
2.1. Biotope Diversity
2.2. Species Diversity of Eastern Guajira
2.3. Genetic Diversity
3. Unique Environments That Support Guajira’s Biodiversity
3.1. Nutrient Enriched Waters
3.2. Complex Structure Seafloors
3.3. Low Anthropogenic Disturbance Seashores
4. Threats to Guajira’s Biodiversity
5. Strategies for Conservation and Management of Guajira’s Coastal Environments
6. Conclusions and Recommendations
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Spalding, M.D.; Fox, H.E.; Allen, G.R.; Davidson, N.; Ferdaña, Z.A.; Finlayson, M.; Halpern, B.S.; Jorge, M.A.; Lombana, A.; Lourie, S.A.; et al. Marine Ecoregions of the World: A Bioregionalization of Coastal and Shelf Areas. BioScience 2007, 57, 573–583. [Google Scholar] [CrossRef] [Green Version]
- Sullivan-Sealey, K.; Bustamante, G. Setting Geographic Priorities for Marine Conservation in Latin America and the Caribbean; The Nature Conservancy: Arlington, VA, USA, 1999. [Google Scholar]
- Burke, L.; Maidens, J. Reefs at Risk in the Caribbean; World Resources Institute: Washington, DC, USA, 2004. [Google Scholar]
- Miloslavich, P.; Díaz, J.M.; Klein, E.; Alvarado, J.J.; Díaz, C.; Gobin, J.; Escobar-Briones, E.; Cruz-Motta, J.J.; Weil, E.; Cortes, J. Marine biodiversity in the Caribbean: Regional estimates and distribution patterns. PLoS ONE 2010, 5, e11916. [Google Scholar] [CrossRef] [Green Version]
- Rueda-Roa, D.T.; Muller-Karger, F.E. The southern Caribbean upwelling system: Sea surface temperature, wind forcing and chlorophyll concentration patterns. Deep Sea Res. Part I Oceanogr. Res. Pap. 2013, 78, 102–114. [Google Scholar] [CrossRef]
- Siung-Chang, A. A review of marine pollution issues in the Caribbean. Environ. Geochem. Health 1997, 19, 45–55. [Google Scholar] [CrossRef]
- Jackson, J.B.; Kirby, M.X.; Berger, W.H.; Bjorndal, K.A.; Botsford, L.W.; Bourque, B.J.; Bradbury, R.H.; Cooke, R.; Erlandson, J.; Estes, J.A. Historical overfishing and the recent collapse of coastal ecosystems. Science 2001, 293, 629–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cramer, K.L. History of human occupation and environmental change in western and central Caribbean Panama. Bull. Mar. Sci. 2013, 89, 955–982. [Google Scholar] [CrossRef]
- Rogers, C.S.; Beets, J. Degradation of marine ecosystems and decline of fishery resources in marine protected areas in the US Virgin Islands. Environ. Conserv. 2001, 28, 312–322. [Google Scholar] [CrossRef]
- Criales-Hernández, M.I.; García, C.B.; Wolff, M. Flujos de biomasa y estructura de un ecosistema de surgencia tropical en La Guajira, Caribe colombiano. Rev. Biol. Trop. 2006, 54, 1257–1282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Instituto de Investigaciones Marinas y Costeras José Benito Vives De Andréis and Corpoguajira. Atlas Marino Costero de La Guajira; Serie de Publicaciones Especiales de Invemar: Santa Marta, Colombia, 2012; p. 188. [Google Scholar]
- Viloria Maestre, E.; Santos Acevedo, M.; Chávez Zabaleta, S.; Romero Arenas, J.A. Pesquería Artesanal del Margen Costero Entre Los Cocos (Magdalena) y Punta Gallinas (La Guajira), Caribe Colombiano; Instituto de Investigaciones Marinas y Costeras José Benito Vives De Andréis: Santa Marta, Colombia, 2016; p. 60. [Google Scholar]
- Vásquez-Carrillo, C.; Peláez-Ossa, M. Insights into the ecology of sea turtles and the fisheries of eastern Guajira from the traditional knowledge of fishermen. Fish. Res. 2021, 238, 105915. [Google Scholar] [CrossRef]
- IDEAM. Mapa Nacional de Degradación de Suelos por Salinización. 2017. Available online: http://www.ideam.gov.co/web/ecosistemas/monitoreo-seguimiento-estado-calidad-suelos/-/document_library_display/9mDFEmPPfxAk/view/95764901 (accessed on 30 May 2021).
- DANE. Censo Nacional de Población y Vivienda (CNPV) de Colombia. 2018. Available online: https://www.dane.gov.co/index.php/estadisticas-por-tema/demografia-y-poblacion/censo-nacional-de-poblacion-y-vivenda-2018 (accessed on 1 May 2021).
- Díaz Merlano, J.M.; Barrios Suárez, L.M.; Gómez-López, D.I. Las Praderas de Pastos Marinos en Colombia: Estructura y Distribución de un Ecosistema Estratégico; Serie Publicaciones Especiales; Instituto de Investigaciones Marinas y Costeras José Benito Vives De Andréis Invemar: Santa Marta, Colombia, 2003; p. 160. [Google Scholar]
- Albis Salas, M.R.; Gómez-López, D.I.; Duque, G. Estructura de las praderas de Thalassia testudinum en un gradiente de profundidad en La Guajira, Caribe colombiano. Bol. Investig. Mar. Costeras-INVEMAR 2010, 39, 381–395. [Google Scholar] [CrossRef]
- Chasqui, L.; Nieto, R.; Rodríguez-Rincón, A.; Gil-Agudelo, D.L. Ambientes marinos de la plataforma somera de la Guajira, Caribe Colombiano. Bol. Investig. Mar. Costeras 2013, 42, 2. [Google Scholar] [CrossRef]
- Federal Geographic Data Committee (FGDC). Coastal and Marine Ecological Classification Standard (CMECS); NOAA: Reston, VA, USA, 2012.
- Posada Posada, B.O.; Henao Pineda, W. Diagnóstico de la Erosión en la Zona Costera del Caribe Colombiano; Instituto de Investigaciones Marinas y Costeras-INVEMAR: Santa Marta, Colombia, 2008. [Google Scholar]
- Vasquez-Carrillo, C.; Sullivan Sealey, K. Diversity and extent of coastal submerged aquatic vegetation in an unexplored coastal upwelling region of the Caribbean Sea. Int. J. Biodivers. Endanger. Species 2018, 2018. [Google Scholar] [CrossRef]
- Díaz-Pulido, G. Vegetación marina de un sector de la plataforma continental de la Guajira (Caribe colombiano). Bol. Investig. Mar. Costeras 2000, 29, 27–34. [Google Scholar]
- Avellaneda, W.J.B.; Patiño, E.A.; Guerra, M.L.; Gouriyu, W.A. New evidence of nesting Dermochelys coriacea (Tortuga Achepa) at Iporoimao-Utareo Beaches, Guajira, Colombia. Mar. Turt. Newsl. 2013, 13, 13–14. [Google Scholar]
- Diaz-Pulido, G.; Garzón-Ferreira, J. Seasonality in algal assemblages on upwelling-influenced coral reefs in the Colombian Caribbean. Bot. Mar. 2002, 45, 284–292. [Google Scholar] [CrossRef] [Green Version]
- Morales-Rozo, A. Monitoreo de Aves Migratorias y Residentes en Siete Humedales de La Guajira, Colombia. In Memorias II Congreso de Ornitología Colombiana; 2006. Available online: http://corpoguajira.gov.co/wp/wp-content/uploads/2017/07/AVES-Costeras.pdf (accessed on 5 August 2021).
- Ceballos-Fonseca, C. Distribución de playas de anidación y áreas de alimentación de tortugas marinas y sus amenazas en el Caribe colombiano. Bol. Investig. Mar. Costeras 2004, 33, 79–99. [Google Scholar]
- Vásquez-Carrillo, C.; Noriega-Hoyos, C.L.; Hernandez-Rivera, L.; Jáuregui-Romero, G.A.; Sullivan Sealey, K. Genetic Diversity and Demographic Connectivity of Atlantic Green Sea Turtles at Foraging Grounds in Northeastern Colombia, Caribbean Sea. Front. Mar. Sci. 2020, 7, 96. [Google Scholar] [CrossRef] [Green Version]
- Puentes, V.; Navia, A.F.; Falla, P.A.M.; Caldas, J.P.; Diazgranados, M.C.; Padilla, L.A.Z. (Eds.) Avances en el Conocimiento de Tiburones, Rayas y Quimeras de Colombia; Cross Market Ltda.: Bogotá, Colombia, 2009; 245p. [Google Scholar]
- Lozano-Duque, Y.; Vidal, L.A.; Navas, S.G.R. La Comunidad Fitoplanctonica en el Mar Caribe Colombiano. In Biodiversidad del Margen Continental del Caribe Colombiano; Navas, G., Segura-Quintero, C., Garrido-Linares, M., Benavides-Serrato, M., Alonso, D., Eds.; Instituto de Investigaciones Marinas y Costeras Invemar: Santa Marta, Colombia, 2010; p. 458. [Google Scholar]
- Urrego-Salinas, M.P.; Peña-Quevedo, H.; Dueñas-Valderrama, F. Equinodermos del Cabo de la Vela (La Guajira, Colombia) enla colección de referencia de la Universidad El Bosque. Biota Colomb. 2016, 17, 124–130. [Google Scholar] [CrossRef] [Green Version]
- Díaz, M.; Zea, S. Distribución de esponjas sobre la plataforma continental de La Guajira, Caribe colombiano. Bol. Investig. Mar. Costeras-INVEMAR 2008, 37, 27–43. [Google Scholar] [CrossRef]
- Polanco, A.; Quintero-Gil, J.A.; Cortés, F.; Duque, G. Contribución al conocimiento de la fauna íctica en dos isóbatas (10 y 50 m) de la región de La Guajira, Caribe colombiano. Bol. Investig. Mar. Costeras 2009, 38, 145–163. [Google Scholar] [CrossRef]
- Almanza-Bernal, M. Análisis Genético Poblacional del Tiburón Cazón Antillano, Rhizoprionodon Porosus (Carcharhinidae), en el Caribe Colombiano; Universidad Nacional de Colombia-Sede Caribe: Santa Marta, Colombia, 2014; p. 62. [Google Scholar]
- Carlson, J.; Charvet, P.; Avalos, C.; Briones Bell-lloch, A.; Cardenosa, D.; Espinoza, E.; Morales-Saldaña, J.M.; Naranjo-Elizondo, B.; Pacoureau, N.; Pilar Blasco, M.; et al. Rhizoprionodon porosus. IUCN Red List Threat. Species 2021. [Google Scholar] [CrossRef]
- Carlson, J.K.; Heupel, M.R.; Bethea, D.M.; Hollensead, L.D. Coastal habitat use and residency of juvenile Atlantic sharpnose sharks (Rhizoprionodon terraenovae). Estuaries Coasts 2008, 31, 931–940. [Google Scholar] [CrossRef]
- Pabón-Quintero, P.E. Estructura genética de las poblaciones de dos especies de haemúlidos (Haemulon aurolineatumy Haemulon steindachneri) en el Caribe colombiano. In Institutio de Estudios de Ciecias del Mar CECIMAR—Sede Caribe; Universidad Nacional de Colombia: Santa Marta, Colombia, 2016; p. 74. [Google Scholar]
- Boström, C.; Pittman, S.J.; Simenstad, C.; Kneib, R.T. Seascape ecology of coastal biogenic habitats: Advances, gaps, and challenges. Mar. Ecol. Prog. Ser. 2011, 427, 191–217. [Google Scholar] [CrossRef] [Green Version]
- Andrade, C.A.; Barton, E.D. The Guajira upwelling system. Cont. Shelf Res. 2005, 25, 1003–1022. [Google Scholar] [CrossRef] [Green Version]
- Muller-Karger, F.E.; Castro, R.A. Mesoscale processes affecting phytoplankton abundance in the southern Caribbean Sea. Cont. Shelf Res. 1994, 14, 199–221. [Google Scholar] [CrossRef]
- Gilbes, F.; Armstrong, R.A. Phytoplankton dynamics in the eastern Caribbean Sea as detected with space remote sensing. Int. J. Remote Sens. 2004, 25, 1449–1453. [Google Scholar] [CrossRef] [Green Version]
- Chollett, I.; Mumby, P.J.; Müller-Karger, F.E.; Hu, C. Physical environments of the Caribbean Sea. Limnol. Oceanogr. 2012, 57, 1233–1244. [Google Scholar] [CrossRef]
- Correa-Ramirez, M.; Rodriguez-Santana, Á.; Ricaurte-Villota, C.; Paramo, J. The Southern Caribbean upwelling system off Colombia: Water masses and mixing processes. Deep Sea Res. Part I Oceanogr. Res. Pap. 2020, 155, 103145. [Google Scholar] [CrossRef]
- Waide, R.; Willig, M.; Steiner, C.; Mittelbach, G.; Gough, L.; Dodson, S.; Juday, G.; Parmenter, R. The relationship between productivity and species richness. Annu. Rev. Ecol. Syst. 1999, 30, 257–300. [Google Scholar] [CrossRef] [Green Version]
- Paramo, J.; Quiñones, R.A.; Ramirez, A.; Wiff, R. Relationship between abundance of small pelagic fishes and environmental factors in the Colombian Caribbean Sea: An analysis based on hydroacoustic information. Aquat. Living Resour. 2003, 16, 239–245. [Google Scholar] [CrossRef]
- Bakun, A. Patterns in the Ocean: Ocean Processes and Marine Population Dynamics; California Sea Grant, in Cooperation with Centro de Investigaciones Biologicas del Noroeste: La Paz, Mexico, 1996. [Google Scholar]
- Hayes, D.B.; Ferreri, C.P.; Taylor, W.W. Linking fish habitat to their population dynamics. Can. J. Fish. Aquat. Sci. 1996, 53, 383–390. [Google Scholar] [CrossRef]
- Wiegand, T.; Revilla, E.; Moloney, K.A. Effects of habitat loss and fragmentation on population dynamics. Conserv. Biol. 2005, 19, 108–121. [Google Scholar] [CrossRef]
- Lutz, P.L.; Musick, J.A.; Wyneken, J. The Biology of Sea Turtles; CRC Press: Boca Raton, FL, USA, 2002. [Google Scholar]
- Meylan, P.A.; Meylan, A.B.; Gray, J.A. The Ecology and Migrations of Sea Turtles 8. Tests of the Developmental Habitat Hypothesis. Bull. Am. Mus. Nat. Hist. 2011, 357, 1–70. [Google Scholar] [CrossRef]
- Bass, A.L.; Lagueux, C.J.; Bowen, B.W. Origin of green turtles, Chelonia mydas, at “Sleeping Rocks” off the northeast coast of Nicaragua. Copeia 1998, 1998, 1064–1069. [Google Scholar] [CrossRef]
- Makowski, C.; Seminoff, J.A.; Salmon, M. Home range and habitat use of juvenile Atlantic green turtles (Chelonia mydas L.) on shallow reef habitats in Palm Beach, Florida, USA. Mar. Biol. 2005, 148, 1167–1179. [Google Scholar] [CrossRef]
- Navas, G.; Segura-Quintero, C.; Garrido-Linares, M.; Benavides-Serrato, M.; Alonso, D. Biodiversidad del Margen Continental del Caribe Colombiano; Serie de Publicaciones Especiales; Invemar: Santa Marta, Colombia, 2010; p. 458. [Google Scholar]
- Martínez, M.L.; Intralawan, A.; Vázquez, G.; Pérez-Maqueo, O.; Sutton, P.; Landgrave, R. The coasts of our world: Ecological, economic and social importance. Ecol. Econ. 2007, 63, 254–272. [Google Scholar] [CrossRef]
- Crain, C.M.; Halpern, B.S.; Beck, M.W.; Kappel, C.V. Understanding and managing human threats to the coastal marine environment. Ann. N. Y. Acad. Sci. 2009, 1162, 39–62. [Google Scholar] [CrossRef]
- Pawar, P.R.; Shirgaonkar, S.S.; Patil, R.B. Plastic marine debris: Sources, distribution and impacts on coastal and ocean biodiversity. PENCIL Publ. Biol. Sci. 2016, 3, 40–54. [Google Scholar]
- Ellison, A.M.; Farnsworth, E.J. Anthropogenic disturbance of Caribbean mangrove ecosystems: Past impacts, present trends, and future predictions. Biotropica 1996, 28, 549–565. [Google Scholar] [CrossRef]
- Dahlgren, C.P.; Kellison, G.T.; Adams, A.J.; Gillanders, B.M.; Kendall, M.S.; Layman, C.A.; Ley, J.A.; Nagelkerken, I.; Serafy, J.E. Marine nurseries and effective juvenile habitats: Concepts and applications. Mar. Ecol. Prog. Ser. 2006, 312, 291–295. [Google Scholar] [CrossRef] [Green Version]
- Gracia, A.; Medellín-Mora, J.; Gil-Agudelo, D.L.; Puentes, V. (Eds.) Guía de las especies introducidas marinas y costeras de Colombia; Serie de Publicaciones Especiales del Invemar No. 23; Instituto de Investigaciones Marinas y Costeras (Invemar) y Ministerio de Ambiente y Desarrollo Sostenible: Bogotá, Colombia, 2011; p. 136. [Google Scholar]
- Marín-Guirao, L.; Lloret, J.; Marin, A. Carbon and nitrogen stable isotopes and metal concentration in food webs from a mining-impacted coastal lagoon. Sci. Total Environ. 2008, 393, 118–130. [Google Scholar] [CrossRef] [PubMed]
- Mensah, A.K.; Mahiri, I.O.; Owusu, O.; Mireku, O.D.; Wireko, I.; Kissi, E.A. Environmental impacts of mining: A study of mining communities in Ghana. Appl. Ecol. Environ. Sci. 2015, 3, 81–94. [Google Scholar]
- Olivares, H.G.; Lagos, N.M.; Gutierrez, C.J.; Kittelsen, R.C.; Valenzuela, G.L.; Lillo, M.E.H. Assessment oxidative stress biomarkers and metal bioaccumulation in macroalgae from coastal areas with mining activities in Chile. Environ. Monit. Assess. 2016, 188, 25. [Google Scholar] [CrossRef]
- Doria Argumedo, C.; Deluque Viloria, H. Niveles y distribución de metales pesados en el agua de la zona de playa de Riohacha, La Guajira, Colombia. Rev. Investig. Agrar. Ambient. 2015, 6, 123–131. [Google Scholar] [CrossRef] [Green Version]
- Vasquez-Carrillo, C. Role of an Upwelled Coastal Area in Northeastern Colombia in the Distribution, Population Dynamics and Genetic Diversity of the Migratory Habitat-Shifting Chelonia Mydas. Ph.D. Thesis, University of Miami, Coral Gables, FL, USA, 2017; p. 128. [Google Scholar]
- Guimaraes dos Santos, R.G.; Silva Martins, A.S.; Torezani, E.; Baptistotte, C.; da Nóbrega Farias, J.; Antunes Horta, P.A.; Work, T.M.; Balazs, G.H. Relationship between fibropapillomatosis and environmental quality: A case study with Chelonia mydas off Brazil. Dis. Aquat. Org. 2010, 89, 87–95. [Google Scholar] [CrossRef] [Green Version]
- da Silva, C.C.; Klein, R.D.; Barcarolli, I.F.; Bianchini, A. Metal contamination as a possible etiology of fibropapillomatosis in juvenile female green sea turtles Chelonia mydas from the southern Atlantic Ocean. Aquat. Toxicol. 2016, 170, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Vanmaercke, M.; Poesen, J.; Maetens, W.; de Vente, J.; Verstraeten, G. Sediment yield as a desertification risk indicator. Sci. Total Environ. 2011, 409, 1715–1725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Restrepo, J.D.; Kettner, A.J.; Syvitski, J.P. Recent deforestation causes rapid increase in river sediment load in the Colombian Andes. Anthropocene 2015, 10, 13–28. [Google Scholar] [CrossRef]
- Middleton, N. Rangeland management and climate hazards in drylands: Dust storms, desertification and the overgrazing debate. Nat. Hazards 2018, 92, 57–70. [Google Scholar] [CrossRef] [Green Version]
- Doney, S.C.; Ruckelshaus, M.; Duffy, J.E.; Barry, J.P.; Chan, F.; English, C.A.; Galindo, H.M.; Grebmeier, J.M.; Hollowed, A.B.; Knowlton, N. Climate change impacts on marine ecosystems. Annu. Rev. Mar. Sci. 2012, 4, 11–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, G.T.; Muller-Karger, F.E.; Thunell, R.C.; Scranton, M.I.; Astor, Y.; Varela, R.; Ghinaglia, L.T.; Lorenzoni, L.; Fanning, K.A.; Hameed, S.; et al. Ecosystem responses in the southern Caribbean Sea to global climate change. Proc. Natl. Acad. Sci. USA 2012, 109, 19315–19320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lacy, R.C. Considering threats to the viability of small populations using individual-based models. Ecol. Bull. 2000, 48, 39–51. [Google Scholar]
- Edgar, G.J.; Stuart-Smith, R.D.; Willis, T.J.; Kininmonth, S.; Baker, S.C.; Banks, S.; Barrett, N.S.; Becerro, M.A.; Bernard, A.T.; Berkhout, J.; et al. Global conservation outcomes depend on marine protected areas with five key features. Nature 2014, 506, 216–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ban, N.C.; Gurney, G.G.; Marshall, N.A.; Whitney, C.K.; Mills, M.; Gelcich, S.; Bennett, N.J.; Meehan, M.C.; Butler, C.; Ban, S.; et al. Well-being outcomes of marine protected areas. Nat. Sustain. 2019, 2, 524–532. [Google Scholar] [CrossRef]
- Agardy, T. Effects of fisheries on marine ecosystems: A conservationist’s perspective. ICES J. Mar. Sci. 2000, 57, 761–765. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez-Moreno, C.; Alonso, C.D.; Segura-Quintero, C. Diseño de un area marina protegida para Bahía Portete—La Guajira, Caribe Colombiano. Bol. Investig. Mar. Costeras 2008, 37, 198–212. [Google Scholar] [CrossRef]
- Delgadillo-Garzón, O.; Newmark, F. Pilot culture of red seaweeds (Rhodophyta) in Portete Bay, La Guajira, Colombia P. Boletín Investig. Mar. Costeras-INVEMAR 2008, 37, 7–26. [Google Scholar]
- León, R.R.; Tirado, D.A.M. Aspectos técnicos y económicos para el establecimiento comercial del maricultivo de algas en Colombia: Experiencias en la Península de La Guajira. Ambiente Desarro. 2011, 15, 123–144. [Google Scholar]
- Carvajal-Romo, G.; Valderrama-Mendoza, M.; Rodríguez-Urrego, D.; Rodríguez-Urrego, L. Assessment of solar and wind energy potential in La Guajira, Colombia: Current status, and future prospects. Sustain. Energy Technol. Assess. 2019, 36, 100531. [Google Scholar] [CrossRef]
- Wali, A.; Alvira, D.; Tallman, P.; Ravikumar, A.; Macedo, M. A new approach to conservation: Using community empowerment for sustainable well-being. Ecol. Soc. 2017, 22, 6. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Formation | Zone | Macrohabitat | Biotope (or Natural Community) |
|---|---|---|---|
| A.01. Marine lagoon | Water Column | Hypersaline (> 38 ppt) shallow water | Ephemeral hypersaline lagoon, dry season due to evaporation |
| Saline (34–38 ppt) coastal embayment connected to coastal ocean | Saline lagoon with a narrow opening to coastal ocean | ||
| Bottom | Unvegetated hardbottom > rocky | Large boulders and angular rocky dominate the bottom. No notable SAV and high turbidity | |
| Vegetated hardbottom > rocky | Small rocks bottom with some macroalgae on the rocks, and some macroalgae between rocks | ||
| Unvegetated softbottom > mud | Shallow muddy softbottom with high turbidity, no SAV, dominated by bivalves | ||
| Unvegetated softbottom > sand | Intertidal to a very shallow sandy bottom, exposed at low tides | ||
| Unvegetated softbottom > sand | Intertidal to a very shallow biogenic (shell) sandy bottom, near mangroves, exposed at low tides | ||
| Vegetated softbottom > mud, sand | Nearshore shallow sparse Thalassia testudinum biogenic (shells) sandy bottom | ||
| Vegetated softbottom > mud, sand | Nearshore shallow sparse mixed seagrass-macroalgae vegetated bottom | ||
| Vegetated softbottom > mud, sand | Nearshore shallow, moderate to dense mixed T. testudinum–Syringodium filiforme–Halodule wrightii vegetated bottom | ||
| Vegetated softbottom > mud, sand | Nearshore shallow, moderate to dense T. testudinum vegetated bottom | ||
| Vegetated softbottom > mud, sand, shells | Nearshore moderate to dense mixed seagrass–macroalgae vegetated bottom and high turbidity | ||
| Vegetated softbottom > mud, sand, shells | Nearshore moderate to dense mixed T. testudinum–S. filiforme–H. wrightii vegetated bottom and high-water turbidity | ||
| Shoreline | Unvegetated rocky shore | Low relief rocky shore filled with human-altered jetties, seawalls | |
| Unvegetated rocky shore | High relief (>4 m) cliffs and ledges | ||
| Vegetated rocky shore/vegetated karst shore | Low relief (<4 m) rocky shore/karst shore | ||
| Unvegetated sediment shore | Fine sandy mud lagoon mudflat, and high turbidity | ||
| Vegetated sediment shore/Mangrove | Rhizophora mangle mangrove shrub thicket fringing in muddy bottom and high turbidity | ||
| Vegetated sediment shore/Mangrove | Mixed R. mangle (red) and Avicennia germinans (black) mangrove shrub thickets fringing, often with red along the water and black inshore in muddy bottom and high turbidity | ||
| Vegetated sediment shore/Mangrove | R. mangle mangrove shrub thicket-overwash, in small, isolated islands in muddy bottom and high turbidity | ||
| Vegetated sediment shore/Mangrove | R. mangle mangrove shrub thicket inverted, with sandy or shelly berm in front of mangroves and high turbidity | ||
| Vegetated sediment shore/Mangrove | Mixed R. mangle–A. germinans mangrove shrub thickets with sandy or shelly berm in front of mangroves and high turbidity | ||
| Vegetated sediment shore/Mangrove | A. germinans mangrove woodland in basin or basin behind coast in muddy bottom and high turbidity | ||
| Vegetated sediment shore/Mangrove | R. mangle mangrove scrubland in muddy bottom and high turbidity | ||
| Vegetated sediment shore/Mangrove | Mixed R. mangle–A. germinans–Laguncularia racemosa (white) mangrove scrubland in muddy bottom and high turbidity | ||
| Vegetated sediment shore/Mangrove | A. germinans mangrove scrublands-fringing in coastal mudflats and high turbidity | ||
| Vegetated sediment shore/Mangrove | A. germinans mangrove scrublands in basins in muddy bottom and high turbidity | ||
| Unvegetated sandy shore | Low energy shoreline, small pocket beach within high relief rocky shore | ||
| Vegetated sandy shore | Low to medium energy Coccoloba uvifera seagrapes vegetated sandy mud beach strand on back beach swale | ||
| C.02 Nearshore Marine Ocean Side | Water Column | Open coastal water | Surface of nearshore coastal water |
| Channels and cuts | Surface of channels and cuts waters | ||
| Shoreline | Unvegetated rocky shore | low relief rocky shore with human-altered jetties, seawalls, filled areas | |
| Unvegetated rocky shore | High relief (>4 m) cliffs and ledges | ||
| Vegetated rocky shore/vegetated karst shore | Low relief rocky shore (<4 m) with sparse xerophytic vegetation | ||
| Unvegetated sandy shore | Low energy small pocket sandy beach within rocky shore | ||
| Vegetated sandy shore | Low to medium energy Coccoloba uvifera seagrapes sparsely vegetated sandy mud beach strand on back beach swale | ||
| Unvegetated sandy shore | Medium to high energy sandy/rubble beach strand with high sand dunes | ||
| Unvegetated rubble shore | High energy rubble/cobble beach with high dunes |
| Group | Number of Species | Species Expected | Reference |
|---|---|---|---|
| Phytoplankton | Unknown | yes | NA |
| Zooplankton | Unknown | yes | NA |
| Seagrasses | 3 | yes | [21] |
| Macroalgae | 113 | yes | [21] |
| Sponges | Unknown | yes | [31] |
| Bony fish | 25 | yes | [13,32] |
| Cartilaginous fish | 15 | yes | [13] |
| Mollusks and Crustaceans | 1 | yes | [13] |
| Marine reptiles | 4 | no | [13] |
| Seabirds and flamingoes | Unknown | yes | NA |
| Corals and jellyfish | 0 | no | NA |
| Echinoderms | Unknown | yes | NA |
| Other invertebrates | Unknown | yes | NA |
| Category | Specifics of Threat | Low | Med | High |
|---|---|---|---|---|
| 1. Development | Small population centers, no large cities, but some areas are vulnerable to migration of people from Venezuela into Colombia. The largest city in the region, Riohacha, is considered a key point of entry and supply to La Guajira, but limited tourism outside this city. | X | ||
| 2. Agriculture and Farming | The area is arid, and with climate change, conditions will be warmer and drier. Water is imported for cattle, and some reservoirs have been built. Over-grazing by goats and cattle is a serious threat to coastal natural communities. | X | ||
| 3. Energy and Mining | Mining of coal via an open pit mine is a large operation in the area but impacts in coastal environments are unknown. Sea turtles display fibropapilloma (FP), which indicates heavy metals pollution in benthic environments. Heavy metals are used in coal mining operations, but the link to sea turtles FP has not been proved. | X | ||
| 4. Transportation and Service corridors | Rail tracks and port for the export of coal create a threat to marine communities in the westernmost Bahia. A large port was built for the export of coal from the region | X | ||
| 5. Biological resource use and harm including within protected areas | Fishing regulations are poorly enforced. Sea turtles and sharks are targeted as high-value target species. Most species are for subsistence and consumed locally. Trawl fisheries in the area have resulted in the collapse of some fish populations. | X | ||
| 6. Human intrusion and disturbance within protected area | There is one national park that includes culturally important dunes and landforms. The park is managed by the park service and protected. | X | ||
| 7. Natural systems modification | In the marine environment, the abundance of species has been altered by an extant commercial fishery that closed at the end of the 20th century. Many fish stocks were locally overfished. It is unknown what damage the trawling might have done to benthic communities. Fishing now is largely for subsistence. Much of the coastal area has been altered by over-grazing. The entire region has experienced desertification with drought and loss of vegetation cover. Many small communities have lost their goats, and cattle are maintained with imported water. | X | ||
| 8. Invasive and other problematic species and genes | Little is known about invasive species in the coastal and marine environments. Few projects have surveyed the entire extent of the coastal ecosystems. | X? | ||
| 9. Pollution entering and generated within the areas | Little pollution monitoring; pollution loading unknown except for sediment and erosion into coastal waters with heavy rains. Coastal desertification and loss of vegetation can result in higher sedimentation rates in the future. Rancheria river waters charged with pollutants from mining and human populations waste may discharge pollution into the ocean nearby Riohacha and other urban areas along the river. | X | ||
| 10. Geological events | No known threats; earthquakes are a remote threat | X | ||
| 11. Climate change and severe weather | Area is suffering from long-term drought, and water is extremely limited in the region. | X | ||
| 12. Culture and social threats | This is an indigenous area under the management of the Wayuu communities. The environmental management of the area is jeopardized by the high poverty and lack of infrastructure in the indigenous communities. | X |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vasquez-Carrillo, C.; Sullivan Sealey, K. Biodiversity of Upwelling Coastal Systems of the Southern Caribbean Sea Adjacent to Guajira Peninsula. J. Mar. Sci. Eng. 2021, 9, 846. https://doi.org/10.3390/jmse9080846
Vasquez-Carrillo C, Sullivan Sealey K. Biodiversity of Upwelling Coastal Systems of the Southern Caribbean Sea Adjacent to Guajira Peninsula. Journal of Marine Science and Engineering. 2021; 9(8):846. https://doi.org/10.3390/jmse9080846
Chicago/Turabian StyleVasquez-Carrillo, Catalina, and Kathleen Sullivan Sealey. 2021. "Biodiversity of Upwelling Coastal Systems of the Southern Caribbean Sea Adjacent to Guajira Peninsula" Journal of Marine Science and Engineering 9, no. 8: 846. https://doi.org/10.3390/jmse9080846