1. Introduction
Tetrodotoxin (TTX), one of the most potent natural neurotoxins [
1,
2], was first detected in pufferfish [
3,
4]. Subsequently, TTX was found in various organisms, including vertebrates and invertebrates [
5,
6,
7,
8], and even in bacteria [
9,
10,
11]. Besides TTX, paralytic shellfish toxins (PSTs) such as saxitoxin and its related compounds are also potent natural neurotoxins, distributed not only to shellfish but also fish, and are known to originate from the toxic products of dinoflagellate [
12]. It is interesting that some pufferfish have both TTX and PSTs in their body [
12,
13,
14].
We have recently developed a novel polyclonal antibody against TTX using its haptenic antigen [
15]. This newly developed polyclonal antibody with analytical procedures using direct one-step ELISA (TTX-ELISA kit) has been demonstrated to be useful to detect TTX and its analogs (TTXs) in toxic organisms [
15,
16]. This assay kit is comparably efficient with our previously developed ELISA kit, which recognizes PSTs [
17].
TTX is abundant in internal organs, such as the hepatopancreas (frequently written as liver for pufferfish in many references in place of the correct organ name) and ovaries of most pufferfish. It is also frequently distributed into skin and less often into muscle [
8]. Such tissue distribution has been well demonstrated anatomically in pufferfish, such as
Tetraodon steindachneri and
Takifugu niphobles, by employing monoclonal antibodies raised against TTX [
18,
19,
20]. This strategy was also successful for the detection of TTX in the secretory glands in the skin of the Japanese newt
Cynops pyrrhogaster [
21], and the red-spotted newt
Notophthalmus viridescens [
22]. Interestingly, such TTX secretion from the secretory gland well confirms the previous observation by Kodama and his colleagues, who found specific organs secreting TTX in pufferfish [
23].
Recently, various analogs of TTX have been identified, and some of them are contained in TTX-bearing animals at appreciable amounts [
24,
25]. Interestingly, 5,6,11-trideoxy (TDT), one of the analogs of TTX, was dominant in various tissues of
Takifugu pardalis pufferfish [
25], and it was exclusively found in the liver (hepatopancreas), testis and even muscle of
Lagocephlus spadiceus pufferfish [
16]. Although the immunohistochemical approach is very powerful in investigating the anatomical localization of TTX, as mentioned above, its monoclonal antibody so far prepared [
26,
27,
28,
29] has not been investigated for its reactivity with its analogs. On the other hand, our polyclonal antibody very recently raised against TTX was conjugated with keyhole limpet hemocyanin (KLH) via a sophisticated method, where 4,9-anhydroTTX prepared from TTX was reacted with 1,2-ethanedithiol, and was found to react not only with TTX, but also well with its analogs, except for 4,9-anhydroTTX [
15]. It is noted that this system was established by referring to our development of a novel polyclonal antibody raised against PSTs based on the unique reactions between C11-O-sulfate toxins, such as gonyautoxins 2 and 3 (GTX2,3), and various thiol compounds, followed by coupling to KLH [
17]. Although TDT is predicted to be a precursor of TTX and converted to TTX by endogenous and/or exogenous factors [
25], exact pathway(s) have remained unknown. Therefore, our polyclonal antibody, which recognizes various TTX analogs, seems a promising tool to survey the metabolic pathway(s) of TTX and the anatomical localization of possible precursors of TTX.
The main objective of the present study was to analyze TTXs in various tissues of newt using the specific polyclonal antibody against TTXs, and to compare the obtained results with those measured predominantly by well-established high-performance liquid chromatography with fluorescence detection (HPLC-FLD). Liquid chromatography/mass spectrometry (LC-MS) was employed only for the determination of TDT. These values were first measured in toxin-bearing pufferfish to confirm the availability of the antibody. The anatomical localization of TTXs in these animals was also demonstrated via an immunohistochemical procedure using the polyclonal antibody. Furthermore, TTXs as well as PSTs were traced for their dynamic changes in tissue distribution, when these animals were fed toxic animal tissues exogenously.
2. Materials and Methods
2.1. Materials
One individual globefish puffer Takifugu flavipterus (female, body weight 42.9 g, total length 125 mm) and one individual brown-lined puffer Canthigaster revulata (male, body weight 148.8 g, total length 193 mm) caught in the Pacific Ocean off the Miura Peninsula, Kanagawa Prefecture, Japan, were kindly supplied in 27 February 2019 by Dr. Seiji Akimoto (Kanagawa Prefectural Fisheries Technology Center, Japan). These specimens were collected before spawning by licensed fishermen and transported to the laboratory at the School of Marine Biosciences, Kitasato University. Alive specimens were placed on ice for the sake of anesthetization and dissected into several tissues, including hepatopancreas, intestine, muscle, skin and gonads.
Several wild specimens of Japanese fire-bellied newt Cynops pyrrhogaster were collected from rice farm fields in Kan-nami, Shizuoka Prefecture, Japan, on 3 October 2018, prior to hibernation (adults, about 10 cm; juveniles, about 4 cm), and on 24 May 2019, during spawning season (adults, about 10 cm). Twenty-eight adult specimens were reared by feeding artificial food specially designed for newts (JAN code No. 4971453054017) (Itosui, Tokyo, Japan) in the laboratory before experiments. The eggs of the newt and larva immediately after hatching were also obtained by rearing specimens in the laboratory. All live specimens (adult, juvenile and larva) of newt were placed on ice for anesthesia, and adults were dissected into several tissues, including the hepatopanreas, intestine, skin, tail, and gonads. Only the tail was subjected to the experiments for juveniles (9 specimens) and the whole body was used for hatched larva (10 specimens). Jelly layers and yolks were dissected from the eggs spawned by 10 matured specimens, pooled respectively, and subjected to toxin analysis. The whole bodies of 10 hatched larvae were also pooled and subjected to the toxin analysis.
Decarbamoylsaxitoxin (dcSTX) prepared previously from the toxic Yezo giant scallop
Mizuhopecten yessoensis [
30] was used as the authentic PST for feeding experiments of the newt. The globefish puffer hepatopancreas was also used for feeding experiments of the newt. While adult specimens collected on 24 May 2019 were employed for feeding experiments with dcSTX, juveniles collected on 3 October 2018 were used for those with the globefish puffer hepatopancreas and TDT.
Animal treatment in this study was conducted in compliance with the codes of ethics at Kitasato University, MB1800510 and MB190510.
All the reagents were of analytical grade and purchased from FUJIFILM Wako Pure Chemical Co., Osaka, Japan, unless specified otherwise.
2.2. Preparation of the Extract and Assay Procedures for TTXs
TTXs were extracted from 2 and 0.1 g of sample tissues from pufferfish and newts, respectively, with 0.1% (
v/
v) acetic acid by heating in a boiling water bath for 10 min, according to the method for TTX described in the Standard Methods of Analysis in Food Safety, Japan [
31].
The total concentrations of TTXs in tissue extracts were determined by ELISA, essentially according to Sato et al. [
15]. Briefly, the polyclonal antibody prepared by Sato et al. was diluted 100 times with 0.9% (
w/
v) NaCl in 0.01 M Tris-HCl buffer (pH 8.2), and 100 µL portions were added to each well of a 96-well ELISA plate (Maxisorp, Thermo Fisher Scientific, Waltham, MA, USA). After stirring at 4 °C overnight, the plate was washed with phosphate-buffered saline free of Ca
2+ and Mg
2+ (PBS(−)), added each with 350 µL Block Ace (4%
w/
v in water; KAC Co., Ltd., Kyoto, Japan) solution, and left to stand at 4 °C overnight. The plate was washed with PBS(−) containing 0.05% (
v/
v) Tween 20 (PBSTw) twice. Then, the sample extract containing 0.2 g tissue/mL was diluted 10–1000-fold with 0.1 M sodium phosphate buffer (pH 7.4) and 50 µL of solution was added to each well of the ELISA plate coated with the antibody specific to the TTXs. Next, 50 µL Biotin-TTX (2 nM, diluted with the same buffer) was added to each well and the plate was incubated at 37 °C for 15 min. After the solution was discarded, the wells were washed with PBSTw and added with 50 μL horseradish peroxidase (HRP)–streptavidin solution (2000 times dilution with PBS(−); Funakoshi, Tokyo, Japan). The plate was incubated at 37 °C for 15 min and the solution was discarded. After washing the wells three times with PBSTw, 100 µL of a solution of Sigma-Fast tablets (OPD-H
2O
2, Sigma-Aldrich, St. Louis, MO, USA) dissolved in 10 mL pure water was added to each well. The plate was incubated at 37 °C for 5 min, and to the wells was added 100 µL HCl (2 M). The plate was subjected to measurement of absorbance at 490 nm with a plate reader (iMark, Bio-Rad, Herculus, CA, USA). The toxin solution samples were analyzed in triplicate. The limit of detection (LOD) and working range of ELISA were approximately 3 nM and 10–300 nM, respectively [
15].
TTX, TTX-6-epimer (6-
epiTTX), TTX- 4-epimer (4-
epiTTX) and 4,9-anhydroTTX were analyzed by HPLC-FLD according to Yotsu et al. [
32] with minor modifications. Briefly, HPLC-FLD was performed as follows: HPLC column, J-Pak Symphonia C18 (5 µm, 4.6 × 150 mm; Jasco, Tokyo, Japan); mobile phase, 0.06 M heptafluorobutiric acid (HFBA, 98%; Sigma-Aldrich)/0.05 M ammonium acetate buffer (pH 5.0), 0.4 mL/min; reaction reagent, 4 M sodium hydroxide, 0.4 mL/min; detector, FP-2020 Plus (Jasco, Ex 365 nm, Em 510 nm, Gain ×1000); pump for mobile phase, PU-2080 Plus (Jasco, Tokyo, Japan); pump for reaction reagent, PU-2080 Plus (Jasco, Tokyo, Japan); reaction coil, i.d. 0.5 mm × 200 cm (stainless, 120 °C in dry oven); integrator, Chromatocorder 21 (SIC, Tokyo, Japan); injection volume, 10 µL (overload injection). The authentic standards for HPLC-FLD consisted of 15.2 µM TTX, 9.5 µM 6-
epiTTX, 12.8 µM 4-
epiTTX and 11.8 µM 4,9-anhydroTTX in 0.03 M acetic acid (>96%, as acetic acid salt, calculated from freeze-dried weight), which were prepared as described in the following
Section 2.3. The LODs of TTX, 6-
epiTTX, 4-
epiTTX and 4,9-anhydroTTX were approximately 30 nM each, and their good linearity [Y = 1.31 × 10
3X + 0.00 (R
2 = 0.9987) for TTX, Y = 3.07 × 10
3X + 0.01 (R
2 = 0.9908) for 6-
epiTTX, Y = 1.69 × 10
3X + 0.00 (R
2 = 0.9956) for 4-
epiTTX, Y = 1.53 × 10
3X + 0.00 (R
2 = 0.9902) for 4,9-anhydroTTX; X = conc. (µM), Y = peak area (mVsec)] was in the range of 0.1–100 μM
TDT was identified according to Sato et al. [
15] by LC-MS (Triple TOF
TM 5600
+; SCIEX, Framingham, MA, USA) as follows: column, Atlantis HILIC (Waters, Milford, MA, USA), 3 µm, 2.1 × 150 mm; mobile phase A, acetonitrile; mobile phase B, 10 mM ammonium formate buffer (pH 4.0); flow rate, 0.2 mL/min; injection volume, 5 µL; CE values, 30 ± 10 eV; gradient, 0 min (B: 20%) → > 15 min (B: 60%) → > 16 min (B: 60%) → > 16.1 min (B: 20%) → > 19 min (B: 20%); mode, TOF-MS, operated in a positive ion mode via electrospray ionization.
2.3. Preparation of Standard TTX and Its Analogs
TTX and its analogs 4-
epiTTX, 4,9-anhydroTTX and TDT were prepared previously from the visceral parts of globefish puffer [
15].
2.4. Analytical Procedures of dcSTX
dcSTX fed on animals and recovered in various tissues of the newt was quantitated by HPLC-FLD according to Oshima [
33].
2.5. Feeding Experiments
The globefish puffer hepatopancreas described in
Section 2.1 was used for feeding experiments of the newt, employing 5 larvae collected on 3 October 2018, with a supposed age of 5 months. Two specimens were reared individually in plastic tanks for one month with non-toxic artificial food specially designed for newt and then fed toxic samples, whereas three individuals were fed non-toxic food throughout. Each of two specimens was provided with three small pieces of the globefish puffer hepatopancreas (ca. 50 mg in total) once a week and subjected to a determination of toxin components and immunohistochemical procedures after the 10-fold dosage.
One specimen of juvenile newt collected on 3 October 2018 was also fed three pellets of diet containing 2.0 mM TDT (24 mg in total, corresponding to 35 nmol TDT) five days a week and reared for 4 weeks. Authentic TDT isolated from globefish puffer [
15] was used in this experiment.
dcSTX prepared previously from the toxic Yezo giant scallop
Mizuhopecten yessoensis [
30] was also used as the authentic PST for feeding experiments of the newt. Two adult specimens collected on 25 May 2019 were individually provided with three pellets each, five days a week, and reared for 4 weeks. The three pellets contained 24 mg in total, corresponding to 31 nmol dcSTX.
2.6. Immunohistochemistry
Each pufferfish specimen was dissected into different organs, whereas newt bodies were divided into 2 or 3 parts (according to body size) from the head to the tail. These organs and parts were fixed with freshly prepared Bouin’s fluid at 4 °C for 24 h. Bouin’s fluid was prepared by mixing saturated 2,4,6-trinitrophenol (picric acid), neutral buffered formalin, and acetic acid in the ratio of 15:5:1 (v/v/v). Fixed samples were subsequently rinsed in cold 70% ethanol, dehydrated through a graded series of ethanol concentrations until 100%, and then embedded in paraplast (Monoject, Sherwood Medical, St. Louis, MO, USA). Sagittal sections were cut at 8 μm with a microtome (RM2125 RTS, Leica Biosystems Nussloch GmbH, Nussloch, Germany), and mounted on MAS-GP-coated slides (Matsunami, Osaka, Japan). Sections were stored at 4 °C until use.
Immunohistochemistry was performed according to the method described by Amano et al. [
34] using a Histofine immunostaining kit (SAB-PO (R), Nichirei Biosciences, Tokyo, Japan). The paraplast sections were deparaffinized in clear plus (Falma, Tokyo, Japan) twice for 10 min each. Next, they were rehydrated through graded ethanol concentrations and twice in distilled water for 5 min each. Endogenous peroxidase activity in the sections was blocked with 3% (
w/
v) hydrogen peroxide in PBS containing 0.3% (
v/
v) Triton-X (PBSTr) for 30 min at room temperature. After washing the sections three times with PBSTr for 10 min each, a blocking solution with 10% normal goat serum was applied to the sections for 15 min at room temperature. After washing the sections three times with PBSTr for 10 min each, the sections were incubated with the antibody against TTX at 4 °C overnight. Anti-TTX antibody was diluted 500-fold with PBSTr containing 0.02% (
w/
v) bovine serum albumin (Vector Laboratories Inc, Burlingame, CA, USA). After washing the sections three times with PBSTr for 10 min each, the sections were added with biotin-labeled anti-rabbit IgG and incubated for 15 min at room temperature. After washing the sections three times with PBSTr for 10 min each, peroxidase-labeled streptavidin was added to the sections for 15 min at room temperature. After again washing the sections three times with PBSTr for 10 min each, peroxidase activity was visualized in 0.1 M phosphate buffer (pH 7.4) containing 0.05% (
w/
v) 3′,3′-diaminobenzidine tetrahydrochloride (DAB), and 0.01% (
w/
v) hydrogen peroxide. Immunoreaction was stopped in distilled water. The sections were counterstained with Mayer’s hematoxylin. Finally, the sections were dehydrated through a graded series of ethanol concentrations, cleared in clear plus, and coverslipped in Permount (Fischer, Fair Lawn, NJ, USA).
To test the specificity of the immunoreactions, the control sections were incubated in the anti-TTX antibody that had been pre-absorbed at 4 °C overnight with excessive amounts of TTXs, which were used as the authentic standards for HPLC-FLD (see
Section 2.2) and diluted 5-fold (
v/
v) with the antibody solution. The subsequent procedure was identical to that used for the abovementioned section preparation.
Sections were observed with a microscope (BX50, Olympus, Tokyo, Japan) and photographs were taken by digital camera (FX380, Olympus). For the terminology of the skin of the newt, we referred to Tsuruda et al. [
21].
4. Discussion
The present study was undertaken to analyze TTXs in various tissues of toxin-bearing pufferfish and newt using a specific polyclonal antibody against TTXs and to compare the obtained results with those derived mainly by HPLC-FLD. TDT was measured by LC-MS, because TDT cannot be detected by HPLC-FLD. The anatomical localization of TTXs in these animals was also demonstrated immunohistochemically using the above-mentioned antibody. Furthermore, TTXs and dcSTX were traced for their possible metabolic changes in various tissues when the newts were administered orally with authentic toxins or toxic viscera.
Both sodium channel-blocking natural neurotoxin groups, TTXs and PSTs, consist of several TTX-and STX-analogs, and their metabolic pathways have remained largely unknown. Previously, TDT was identified, and presumed to be a starting material to be converted finally into TTX in pufferfish [
25]. Sato and his colleagues have also recently claimed the same possibility using a polyclonal antibody raised against TTX and toxic and non-toxic
Lagocephalus pufferfish [
15,
16]. The antibody easily detected TTXs in non-toxic
Lagocephalus pufferfish that contained almost exclusively TDT without using LC-MS, which requires complicated procedures, while other fish belonging to this genus contained large amounts of both TTX and TDT [
16].
In the present study, we attempted at first to determine the tissue distribution of TTXs in pufferfish and newt using the immunoassay with the specific antibody and LC-FLD, but not using LC-MS. As a result, the ratios of the amount of TTXs in pufferfish determined by ELISA to that determined by HPLC-FLD were changed depending on individuals. These results suggest that pufferfish contained TTX analogs other than 6-
epiTTX, 4-
epiTTX and 1,9-anhydroTTX—presumably TDT. There are several TTX analogs reported so far, in addition to the above-mentioned analogs, such as 4,9-anhydroTTX, 5-deoxyTTX, 11-deoxyTTX and 11-oxoTTX [
35]. Alternatively, HPLC-FLD would have detected contaminants with the same elution times as those of TTXs. In the case of the adult newt, several individuals showed an amount of TTXs determined by ELISA lower than that determined by HPLC-FLD. Thus, our hypothesis that ELISA would detect the presence of a large amount of TDT in toxin-bearing animals was not necessarily verified. In contrast to the adult newt, juvenile, hatched larva and eggs contained very low amounts of TTXs (
Table 4 and
Table 5), suggesting that the newt accumulates exogenous TTXs or synthesizes them endogenously, although the mechanisms involved are uncertain. Considerable individual variations in toxin levels have been reported within and among populations of red-spotted newt,
Notophthalmus viridescens, inhabiting North America, where the exogenous source of TTX derived either via the food chain or via the synthesis of symbiotic bacteria has been suggested to explain such high variability and the lack of TTX in certain populations [
36]. The TTX-producing bacterial strains, isolated as bacterial symbionts from toxic newts, consisted of four genera, including
Aeromonas,
Pseudomonas,
Shewanella, and
Sphingopyxis, from rough-skinned newts,
Taricha granulosa [
37].
In spite of such uncertainty, we attempted experiments to determine the metabolism of endogenous and exogenous toxins, using newt specimens the tails of which were cut and then regenerated naturally. After 6 months rearing, the regenerated tails, though they were much shorter than their original counterparts, accumulated about half the amount of TTXs originally observed before the removal of the tail, as determined by ELISA (
Table 6). These results suggest that not only symbionts, but also biosynthesis, in the newt are responsible for the production of TTXs.
We supplied a non-toxic diet during rearing, and the TTXs found in the regenerated tail are likely to have been transferred from somewhere in the body, or biosynthesized internally. Therefore, feeding experiments were performed with larval newts using the globefish puffer hepatopancreas, resulting in the accumulation of TTX and 4,9-anhydroTTX predominantly in the tail (
Table 7). Further experiments using adult newts for four weeks with authentic TDT in diets demonstrated that TDT was recovered at a yield of 85% in the tail, but no TTX or its analogs were detected. These results suggest that TDT could not be metabolized to TTX or its other analogs under the present experimental conditions. Additional experiments were further performed using dcSTX, another highly neurotoxic component comparable to TTXs. When dcSTX was introduced as a diet in newt-specific food, almost all the dcSTX was recovered, and the accumulation was the highest in the tail, followed by the spermatic duct, skin and intestine (
Table 8).
Previously, Kodama et al. discovered TTX-secreting glands in pufferfish which have gland-like structures in the skin [
23]. These cells are completely enclosed by epithelial cells with developed microfilmaments, except at their opening to the lumen. Tanu et al. immunohistochemically localized TTX in the skin of a brackishwater puffer
Tetraodon steindachneri using monoclonal antibody against TTX, although they observed TTX localization only on the periphery of the TTX-secreting glands [
18]. In the present study, secretory glands were apparent in both globefish and brown-lined puffers, and showed a positive reaction with the polyclonal antibody together with the epidermis layers (
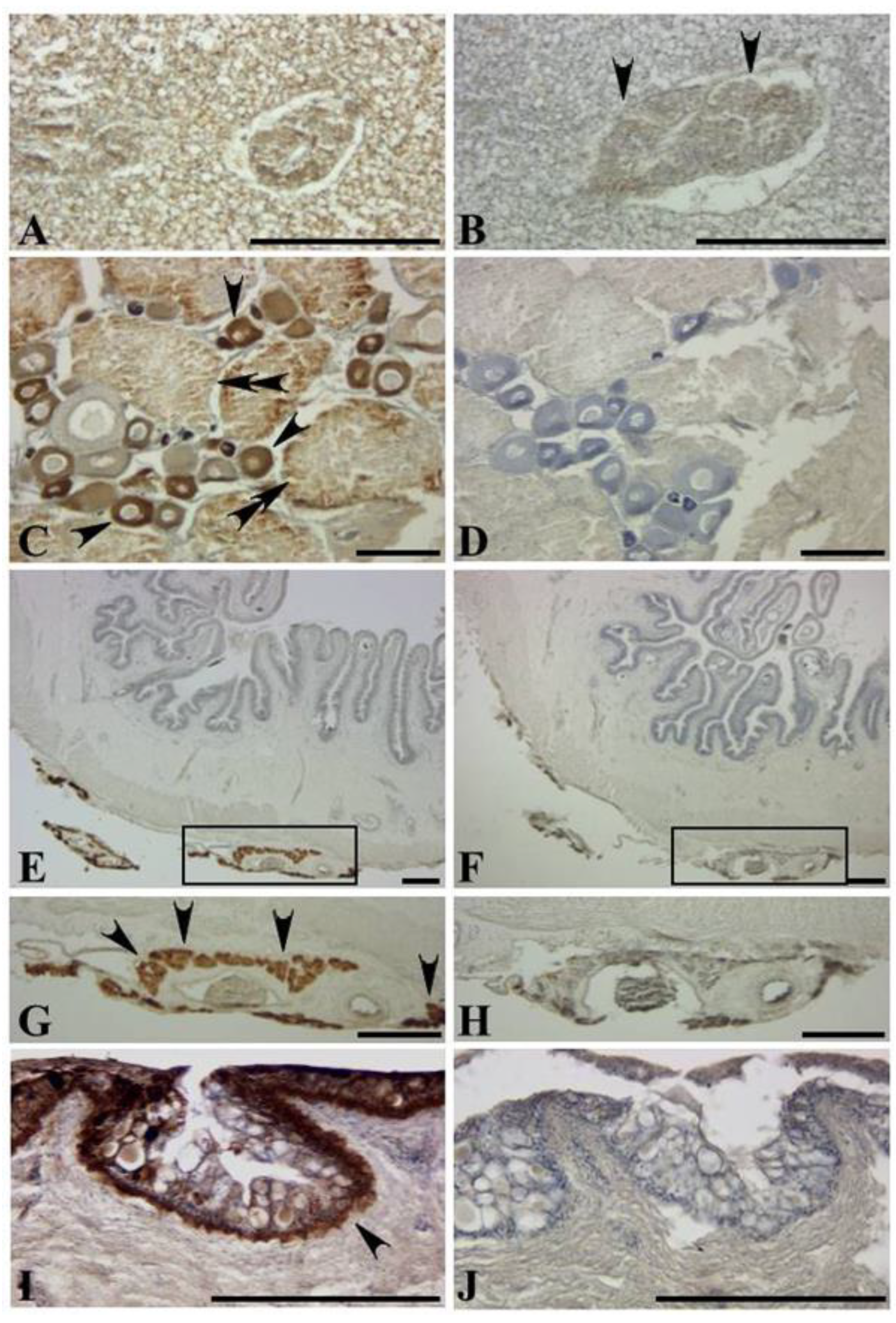
Figure 1 and
Figure 2). The antibody reaction was also observed in the hepatic region, ovary and sac-like tissues just outside the serous membrane of the intestine (
Figure 1). Pufferfish are known to have a hepatopancreas in place of the liver, where the pancreas is surrounded by and bound to the hepatic cells [
38]. Interestingly, the immunoreaction was observed with hepatic cells, but almost nothing was observed in the pancreas (
Figure 1A).
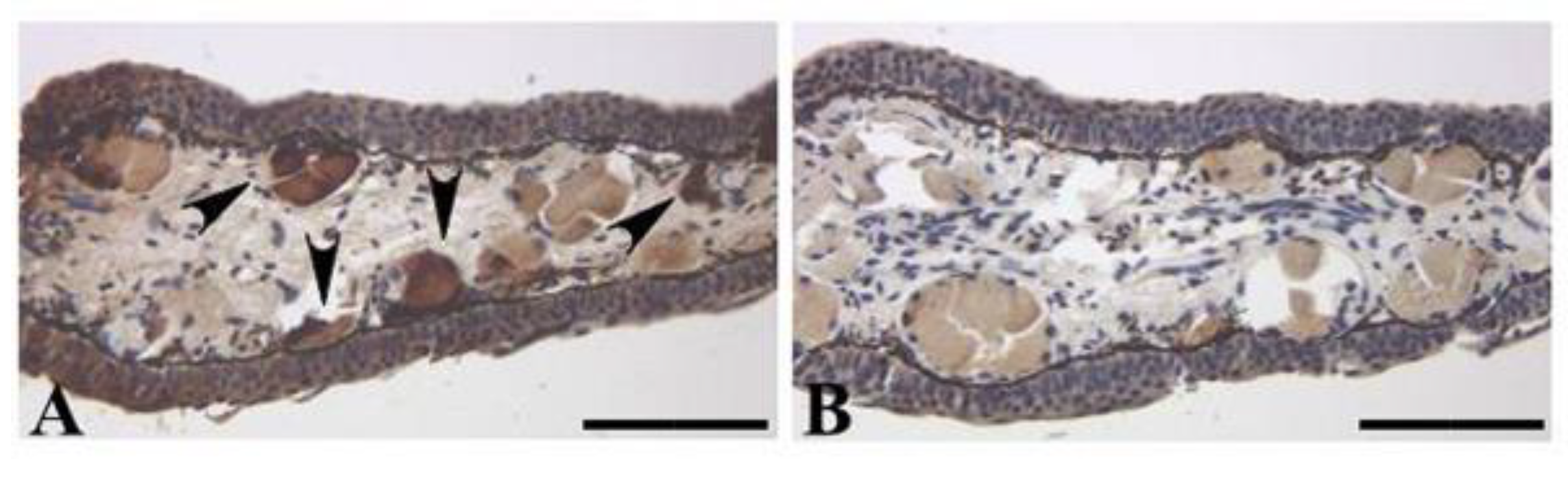
The adult newt tail and the regenerated tail showed positive reactions in the mature glands in their dermis, as reported by Tsuruda et al. [
21]; the newt secreted an appreciable amount of TTX and 6-
epiTTX from the skin following handling stimulus, suggesting that the newt releases TTX via the mature glands in its dermis. Interestingly, the newt dosed with toxic tissues displayed staining not only in the intestine, the ovary, and the skin, but also in the tail (
Figure 5G). Thus, it is indicated that TTX transfers from the body to the tail.
In conclusion, the present study analyzed TTXs in various tissues of toxin-bearing pufferfish and newt using the specific polyclonal antibody against TTXs, and compared the obtained results with those obtained by HPLC-FLD and LC-MS. The anatomical localization of TTXs in these animals was also demonstrated by an immunohistochemical procedure using the polyclonal antibody. Furthermore, TTXs as well as dcSTX were traced for their dynamic changes in tissue distribution when these animals were fed toxic animal tissues exogenously to provide toxins. Here, the newt was the predominant target, and pufferfish, which are well-studied in terms of toxin distribution, were used as the reference, although we only subjected one each of two species of pufferfish to analyses. The obtained results suggest that TTXs absorbed from the environment are transferred to and accumulated in various organs or tissues in a species-specific manner. In spite of our extensive work, however, the synthetic or metabolic pathway(s) of TTXs have remained largely unknown. To cope with such mysterious and tantalizing questions, we need to perform further experiments on the environmental symbionts and/or endogenous factors that may interact with toxin-bearing animals for toxin synthesis and metabolism.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}