
The Key Element Role of Metallophores in the Pathogenicity and Virulence of Staphylococcus aureus: A Review


Department of Laboratory Sciences, Faculty of Public Health, Islamic University of Lebanon, Khalde P.O. Box 30014, Lebanon
*
Authors to whom correspondence should be addressed.
Biology 2022, 11(10), 1525; https://doi.org/10.3390/biology11101525
Submission received: 20 August 2022
/
Revised: 17 September 2022
/
Accepted: 14 October 2022
/
Published: 18 October 2022
(This article belongs to the Special Issue Secondary Metabolites from Microorganisms, or Microorganism-Host Interaction?)
Abstract
:Simple Summary
Metallophores, which are secondary metabolites secreted by bacteria, play an important role in their virulence. The bacterium Staphylococcus aureus produces several types of metallophores, such as staphyloferrin A, staphyloferrin B, and staphylopine, which are responsible for metal ion sequestering. The former is specific for iron chelating, while the latter is a wide-spectrum metallophore, but it mainly chelates zinc. The detailed description of the biosynthesis, export and import processes of each metallophore type is highlighted in this review. Moreover, the genetic regulation is explained as well. Previous studies that provide evidence about the crucial part of metallophores in Staphylococcus aureus pathogenesis were also mentioned herein.
Abstract
The ubiquitous bacterium Staphylococcus aureus causes many diseases that sometimes can be fatal due to its high pathogenicity. The latter is caused by the ability of this pathogen to secrete secondary metabolites, enabling it to colonize inside the host causing infection through various processes. Metallophores are secondary metabolites that enable bacteria to sequester metal ions from the surrounding environment since the availability of metal ions is crucial for bacterial metabolism and virulence. The uptake of iron and other metal ions such as nickel and zinc is one of these essential mechanisms that gives this germ its virulence properties and allow it to overcome the host immune system. Additionally, extensive interactions occur between this pathogen and other bacteria as they compete for resources. Staphylococcus aureus has high-affinity metal import pathways including metal ions acquisition, recruitment and metal–chelate complex import. These characteristics give this bacterium the ability to intake metallophores synthesized by other bacteria, thus enabling it to compete with other microorganisms for the limited nutrients. In scarce host conditions, free metal ions are extremely low because they are confined to storage and metabolic molecules, so metal ions are sequestered by metallophores produced by this bacterium. Both siderophores (iron chelating molecules) and staphylopine (wide- spectrum metallophore) are secreted by Staphylococcus aureus giving it infectious properties. The genetic regulation of the synthesis and export together with the import of metal loaded metallophores are well established and are all covered in this review.
1. Introduction
Staphylococcus aureus (S. aureus) is considered one of the most widespread infectious bacteria. It is found in the environment as well as being part of the human skin and nasal microbiota [1]. Normally, S. aureus is harmless on healthy skin, but once it enters the blood or internal tissues, diverse infections occur including pneumonia, infection of surgical site and nosocomial bacteremia [2]. Systemic S. aureus infection depends on the bacteria breaking through the epithelial protective layer. The incidence rate of this serious medical condition is between 20 and 50 cases/100,000 per year, with fatality rate ranging from 10% to 30% [3]. Moreover, S. aureus forms biofilms that are associated with medical device infections such as prosthetic joints and endocarditis [4]. The prevalence of antibiotic resistance S. aureus isolates, methicillin-resistant S. aureus (MRSA), is posing a serious problem for combating infectious diseases caused by this pathogen [5].
This bacterium is Gram-positive, its cell wall composed of a lipid membrane surrounded by a thick peptidoglycan and teichoic and lipoteichoic acid layer [6]. The rigidity of the cell wall is determined by these peptidoglycan chains that gives the bacterium its shape and protects it from osmotic lysis [7]. The staphylococcal cell surface is negatively charged due to the presence of teichoic acids, which contain phosphate and play a role in the localization and acquisition of metal ions [8]. S. aureus produces notable virulence factors, contrary to some other bacteria that promote disease only by secreting toxins. These factors include an excessive amount of immune-evasion substances and a wide range of small-molecule mediators, also called secondary metabolites, that enable host colonization during infection and give S. aureus the ability of successful competition with other microorganisms in nutrient-poor conditions. The biosynthesis of such metabolites involves complex enzymatic cascades, and they are considered vital components of staphylococcal interactions, with microbiome occupying the same niche e.g., human nares [9]. Such mediators belong to a large variety of protein and non-protein compounds that have attractive properties for future drug development. Examples of these molecules include bacteriocins (lanthipeptides, thiopeptides, fibupeptides) that inhibit bacterial competitor species; signaling molecules such as thiolactone peptides that induce or inhibit sensory cascades in other bacteria; or metallophores (staphyloferrins and staphylopine) that are transition metal ions binding molecule [10].
This review focuses on metallophores due to their crucial role during both infections and bacteria–microbiome interactions. These low-molecular-weight molecules are produced by bacteria during scant availability of essential transition metal ions, like iron or zinc. In S. aureus, metal ion cofactors are required in various biochemical processes such as nucleic acid and protein synthesis, virulence factor expression regulation, DNA replication and reactive oxidative species (ROS) metabolism [11]. Metallophores can import metal ions into the bacteria even when the amount in the surrounding environment is extremely low. This enables the invading bacterium to obtain essential metallic nutrients from the human body and endure “nutritional immunity” even when the immune system is acting in full force to limit the availability of metal ions [12].
Metal transport systems of S. aureus are essential for its virulence, especially those required for iron and manganese accretion [13]. The uptake of iron can be conducted by the secretion of siderophores that form soluble Fe3+ complexes in order to be actively taken up via specific receptors [14]. Four siderophores synthesized and secreted by S. aureus were previously described in the literature. The most studied are the two high-affinity siderophores (staphyloferrin A and staphyloferrin B) that bind to free iron and compete with host iron sequestering proteins lactoferrin and transferrin [15]. Then HtsABC and SirABC transporters import iron bound to staphyloferrin A and B, respectively [16]. However, S. aureus has metal requirements other than iron. This pathogen also produces staphylopine, which is another metal chelator that allows it to uptake several metals, including nickel and zinc, from the host during infection [17]. In previous studies, an ABC metal transport system (ATP-binding cassette) named CntABC (Cnt, cobalt-nickel transporter) was identified in S. aureus [18]. This ABC transporter allows S. aureus to internalize nickel and cobalt under low-zinc conditions through CntA (solute-binding protein) and contributed to S. aureus virulence of urinary tract infection in mice [19]. In addition, it produces hemolysins that rupture red blood cells membranes causing hemoglobin release which is then degraded to heme and free iron by a series of processing and import steps brought about by the Iron-Regulated Surface Determinant (Isd) staphylococcal system [20].
The presence of metalloregulators (metal-dependent regulatory proteins) in S. aureus as in other organisms allows it to sense the metals’ bioavailability and maintain metal ion homeostasis. This metal regulation is achieved through the transcription control of the genes involved in metal ion import, storage, distribution and efflux. The metal-sensing mechanism requires the reversible interaction of one or more specific metal ions with the regulator protein, so that the affinity of the regulatory protein for the specific DNA sequences located in target promoter operons will be altered. Such alterations in the metalloregulator occupation on the operator regulate target gene transcription [21]. These processes maintain suitable transition metal ion levels, thus allowing S. aureus adjustment to changing host environment conditions and shifting metabolic demands.
2. Metallophores Produced by Staphylococcus aureus
2.1. Siderophores
Iron, which has multiple oxidative states, is needed in many vital life processes such as electron transfer and DNA replication [22]. Bacteria require iron as an enzyme cofactor in the catalysis of redox reactions included in their basic cellular processes [23]. In the human body the level of free iron ions is extremely low since most iron is confined to storage, metabolic molecules and transport. Ferrous Fe2+ ions are exceedingly toxic due to their association with the Fenton reaction that produces harmful hydroxyl radicals [24]. On the other hand, ferric Fe3+ ions are insoluble at physiological pH and thus not readily bioavailable [25]. In order to endure this iron shortage iron, bacteria developed several mechanisms for obtaining iron from the host since it is important for their colonization during infection. These mechanisms include acquiring heme-bound iron, absorption by membrane-bound intake systems and siderophore secretion [26]. Siderophores are iron high-affinity metallophores and are essential pathogenicity factors in bacteria including S. aureus. The latter produces and secretes two staphyloferrins (siderophores) into the extracellular environment to scavenge iron. In addition, S. aureus has specific uptake systems for these staphyloferrins and for siderophores produced by other microorganisms as well [27].
There are four distinguished types of siderophores, catecholate, phenolate, hydroxamate and carboxylate, classified according to their iron chelation moieties [28,29]. The synthesis of siderophores is achieved either by non-ribosomal peptide synthesis (NRPS) or by NRPS-independent siderophore (NIS) synthesis (polyketide synthase (PKS) domains) that function together with NRPS units [30]. Also, a small quantity of siderophores is produced independent of these two pathways [31]. The NRPS pathway is the most common while that of NIS is less characterized.
NRPS siderophores have peptidic scaffolds, often incorporating nonproteinogenic amino acids and their derivatives, which are assembled stepwise with covalently bound intermediates [32]. In the NIS pathway, the covalent attachment of intermediates to the enzymes was not noted [30]. In the first synthesis route, siderophores are manufactured by the assembly of individual enzymes where dicarboxylic acids are condensed with diamines, amino alcohols, and alcohols in alternating subunits. Further subunit modifications (decarboxylation, oxidation or isomerization) are performed by distinct enzymes encoded by clusters of genes located near those related to synthetases encoding. Composite pathways using both assembly types were also reported [33]. S. aureus uses the NIS pathway in the synthesis of its staphyloferrins. Siderophores secretion is an active process (energy driven) and is flowed out through transport pumps [28].
The intake of iron chelated by siderophores varies between Gram-negative and Gram-positive bacteria due to the presence of an outer membrane in Gram-negative bacteria through which they should be transported [34,35]. In Gram-negative bacteria, the loaded siderophores are recognized specifically by receptors (β-barrel) found in the outer membrane. The change in the receptors conformation once the ligand is bound allows the translocation of loaded siderophores into the periplasm. [36]. Then, the transport into the cytoplasm is mediated by an ABC transporter located in the inner membrane [37]. The iron is reduced in the periplasm in some cases, and only Fe2+ ion is brought into the cytosol [38]. Concerning Gram-positive bacteria, the import of siderophores is directly achieved by an ABC transporter extending across the cell membrane because there is no outer-membrane receptors [39]. After iron release, siderophores may be either recycled [40] or hydrolyzed [41].
2.1.1. Ferric Uptake Regulator (Fur)
In the host iron depleted environment, S. aureus alters its profile of protein expression. Such change is conveyed by the iron-dependent ferric uptake regulator (Fur) [42]. When iron is present, Fur binds to a DNA consensus sequence noted as the Fur box located upstream the Fur-regulated genes and stops their transcription. On the contrary, when iron is scarce, Fur is released from the DNA, reducing Fur-mediated transcriptional suppression. [43]. The imaging of mice infected with S. aureus revealed the expression of Fur-regulated genes in abscesses found in heart and kidneys, which suggested that the bacterium was iron starved in these organs [44,45]. Upon comparing the protein profiles in the cytoplasm of wild type S. aureus and Fur mutant type, twenty staphylococcal proteins were found to be more plentiful when Fur was absent, indicating that Fur negatively regulates these proteins [13]. Furthermore, this investigation showed increases in fermentative and glycolytic enzymes, demonstrating that S. aureus regulates its metabolism so that it can adjust to iron-scant host environment. This lactate secretion decreases the microenvironment pH and thus the transferrin affinity for iron [46]. These data reveal that the host environment is altered by S. aureus so that iron is released from the host proteins assuming the increase in its bioavailability. Fur also regulates the virulence factors expression involved in host cell attachment and formation of biofilm [47,48].
Fur is considered a suppressor that uses Fe2+, it prevents transcription when iron is available through binding to the siderophores operon promoter region. This suppression stops when iron concentration decreases and siderophores biosynthesis and transport genes are allowed [49]. Three Fur homologues (Fur, PerR, and Zur) are present in S. aureus. Fur represses both the hydroxamate intake system fhuD2 genes and the sirABC and sstABCD transport operons for siderophores whenever iron is available. On the other hand, the storage of iron and the resistance of oxidative stress are regulated by PerR. The latter expresses variable genes whenever iron is available such katA, ahpCF, bcp, trxB, ftn, which encode catalase, alkyl hydroperoxide reductase, bacterioferritin comigratory protein, thioredoxin reductase and ferritin respectively [50]. Also, PerR was shown to be important for S. aureus virulence in mice. Concerning Zur, it is responsible for zinc regulation.
2.1.2. Staphyloferrin A
This highly hydrophilic, carboxylate-type siderophore (480 Da) is fabricated by the condensation of two citrate molecules to a D-ornithine backbone [51]. The products of sfaABCD operon regulated by Fur are responsible for both the synthesis and export of Staphyloferrin A [52]. In S. aureus, the two synthetases SfaD and SfaB, which belong to the NIS class and encoded by its genome, perform the catalysis of condensation reactions producing apo- Staphyloferrin A. These reactions proceed when SfaD form an amide bond between first citrate and D-ornithine δ-amine by using it as a nucleophile. Then, SfaB catalyzes the formation of a new bond between the second citrate and the D-ornithine δ -amine through nucleophilic substitution [53]. Staphyloferrin A is exported and carried out from the cell by SfaA that is analogous to known efflux transmembrane proteins and belongs to the principal facilitator superfamily [54]. The production of this siderophore depends on its precursor’s availability. It is supposed that SfaC, a recognized racemase, converts D-ornithine from L-ornithine found in the cytoplasm [55]. The availability of citrate is temporary, where TCA cycle (Tricarboxylic Acid Cycle) is needed for Staphyloferrin A production in vivo [56].
The import of iron Fe3+ loaded Staphyloferrin A is arbitrated by the heme transport system ABC transporter denoted as HtsABC. [52]. Fur regulates the operon of htsABC which is located next to the Staphyloferrin A biosynthesis operon sfa. HtsA is spontaneous bacterial peritonitis (SBP) and HtsBC is the heterodimeric permease integrated in the membrane. The Hts system depends on FhuC (an unselective ATPase) because the ATP-binding protein gene is absent in htsABC operon. On the other hand, S. aureus has ntrA gene, regulated by Fur, that encodes a nitroreductase needed for iron loaded Staphyloferrin A utilization [57].
This bacterium does not produce glutathione but uses bacillithiol instead for its redox homeostasis [58]. This metabolite might have a comparable function with NtrA. Moreover, it was suggested that NtrA is essential for Fe3+-Staphyloferrin A usage since ntrA mutant strain, lacking Staphyloferrin B as well, had reduced growth in iron-depleted media [57]. Contrary, NtrA is not needed for Fe3+- Staphyloferrin B or Fe3+-desferrioxamine utilization proposing a unidirectional iron transfer from desferrioxamine to Staphyloferrin A and then released by NtrA [57].
2.1.3. Staphyloferrin B
Staphyloferrin B (448 Da) is also a carboxylate-type siderophore. Its biosynthesis and export is mediated by enzymes encoded by Fur regulated gene cluster sbn (sbnABCDEFGHI) [59]. This siderophore is assembled from a molecule of citrate and another of α-ketoglutarate with two molecules of L-Dap (L-2,3-diamopropionic acid). Such assembly is achieved by SbnC, SbnE, SbnF, which are NIS-synthetases, and SbnH (a decarboxylase) [60]. Contrary to the Staphyloferrin A, the gene cluster sbn encodes enzymes that synthesize the three precursors of Staphyloferrin B, in addition to the proteins responsible for its assembly [61]. Citrate is produced by SbnG, while SbnA and SbnB work in conjunction in order to produce the two building blocks (α-ketoglutarate and L-Dap) needed for the synthesis of Staphyloferrin B [62]. Concerning sbnI, the cluster last gene, it has a role both in the regulation and the precursor biosynthesis. This gene is needed for sbnD-H full expression, consequently regulating in vivo iron acquiring by Staphyloferrin B [63]. Furthermore, SbnI generates SbnA substrate [64]. Staphyloferrin B is produced by S. aureus in a wide range of mediums, for example infections of bones and blood, which differ in nutrient availability (oxygen and glucose), so the bacterium metabolism will be altered. Thus, Staphyloferrin B can be produced by SbnA, B, G, and I apart from the TCA cycle or glycolysis [64].
When the assembly is complete, the export from the cell is mediated by SbnD [59,60]. The latter is a facilitator protein having ten transmembrane domains. However, there exist additional exporters, which is not the case for Staphyloferrin A [54]. It was found that the extracellular levels of Staphyloferrin B decreased while the intracellular levels increased in sbnD S. aureus deletion strain after 14 h under iron depleted growth conditions. However, after a lag period of 36 h, the extracellular and intracellular levels were similar to those in wild-type bacterium, indicating the activation of an exporter independent of SbnD [54].
S. aureus takes iron-loaded Staphyloferrin B by the staphylococcal iron regulated system (Sir), encoded by the operon sirABC, which is regulated by Fur [16]. The disruption of SirA (lipoprotein for substrate binding) or SirB (membrane permease) causes weakened growth in iron-confined conditions [16]. Similar to the Hts system, the FhuC ATPase connects with SirABC forming a complete ABC transporter [65]. The iron release mechanism from Staphyloferrin B when it reaches the cytoplasm is still not known.
The binding orientations for SirA and HtsA are so different since only few siderophore binding residues are conserved between them despite of the resemblance in their structure. Tthe specificity of SirA for iron loaded Staphyloferrin B and HtsA for iron loaded Staphyloferrin A, is attributed to these differences in the residues responsible for each siderophore binding [66].
The structures of Staphyloferrin A and Staphyloferrin B are illustrated in Figure 1.
2.1.4. Staphylobactin
This siderophore was discovered upon studying the virulence of siderophores in S. aureus. In an infected mouse model, the siderophore-producing strains revealed higher virulence than those that lacked siderophores [59]. The operon responsible for the siderophore production in the study was named sbn as well. The researchers found that sbnE mutants were avirulent and termed the siderophore extracted staphylobactin; they later found that it is a hydroxamate type having citric acid residues [67]. Like other S. aureus siderophores, staphylobactin was transported via ABC transporter. sirB or sirA mutants were not able to uptake Fe-staphylobactin [59].
2.1.5. Aureochelin
Aureochelin (577 Da) is another siderophore produced by S. aureus. Its structure is still unknown, but it might be a phenolate catecholate type due to its reactivity with the Swain reagent that is used to detect phenolic moieties. Two surface-associated proteins of molecular weight 120 and 88 kDa were found to be expressed in iron depleted conditions and were related with the production of Aureochelin. Moreover, these proteins were also detected as antigenic [15].
2.1.6. Xenosiderophores Uptake by S. aureus
S. aureus is able to uptake siderophores produced by other microorganisms and it has both hydroxamate and catechol exogenous siderophores uptake systems thus enabling it to compete with other bacteria. Concerning the hydroxamate system, it is moderated by an ABC transporter named Fhu (ferric hydroxamate uptake) system and consists of the operon fhuCBG and FhuD1 and FhuD2 proteins regulated by Fur [68]. FhuB and FhuG are very hydrophobic and bound to the membrane, while an ATP-binding protein is coded by fhuC [69]. The genes fhuD1 and fhuD2 are outside the operon fhuCBG and code for lipoprotein receptors [70]. FhuD2 has a wider hydroxamate range and higher affinity than FhuD1 [68]. The import of exogenous hydroxamate siderophores ferrichrome, aerobactin, ferrioxamine B and coprogen involves FhuD2. On the other hand, FhuD1 transports only ferrioxamine B and ferrichrome. Unlike other known transport systems, no conformational changes were shown for FhuD2 during iron-hydroxamate binding [71].
The staphylococcal siderophore transporter (sstABCD) operon encodes the ABC transporter responsible for iron-catechol siderophores uptake. Heterodimeric permease is formed by SstA and SstB. SstC is an ATP-binding homodimeric protein, and SstD is the lipid-anchored substrate-binding proteins (SBP) [72,73]. SstD is expressed mostly in iron depleted conditions, [72]. The uptake of ferric enterobactin, 2,3-dihydroxybenzoic acid (DHBA) and bacillibactin complexes strictly requires SstABCD. Although the Sst system can import also salmochelin 4 and petrobactin, there exists another transporter involved in their in vivo uptake, but it is still unidentified [51]. Furthermore, the catecholamine- iron complexes import requires SstABCD. Epinephrine, for example, which is a catecholamine hormone, is known to reduce iron bound to transferrin thus forming iron complexes that S. aureus can use as a source of iron [74].
2.1.7. The Virulence of Staphylococcal Siderophores
Greater siderophore production in S. aureus is associated with its virulence manifested through additional bacterial counts, larger abscesses and further inflammation in infected mice [75]. Moreover, the strains that showed higher resistance to the neutrophils in vitro were found to produce more siderophores [75]. Mutant types that were unable to produce staphylobactin could not persevere in mice. This finding suggests that uptake of iron by sources other than non-siderophores might be necessary in the early infection stages, while siderophores are essential in later infection stages. The deletion of the chromosomal sbn operon impaired the growth of S. aureus in serum [59], emphasizing the significance of iron uptake schemes for the pathogenesis of this bacterium.
2.2. Additional Metal Acquiring Systems
In addition to the iron uptake system, S. aureus have other various systems of transportation for transitional metal ions, such as Cnt, Adc, NixA and Nik [18,76,77]. The latter is an ABC transporter and is essential for bacterial acquisition of nickel. This system (Nik) is effective in delivering nickel by the means of small chelating molecules (e.g., L-histidine), determining the activity of urease, and having a crucial role in the mouse urinary tract colonization [78,79]. Another nickel-acquiring system in S. aureus is NixA, which is a secondary transporter of NiCoT (nickel-cobalt transporter) membrane protein family. Along with Nik, NixA is also critical for the activity of urease and colonization in kidney [19]. Adc system is responsible for zinc uptake in Gram-positive bacteria including S. aureus and it is composed of AdcA that is a metal acquiring unit and AdcBC which is an ABC transporter [17]. Cnt, on the other hand, can transport several metals such as nickel, zinc, copper, cobalt, zinc, and manganese [17] but at zinc scarce conditions, it serves as a zinc uptake system [17].
2.2.1. Metallophore Staphylopine
In the first place, S. aureus utilizes Adc for importing zinc. Cnt system will be aroused when Adc alone becomes unable to meet zinc cellular requirement. A distinctive characteristic of Cnt is utilizing staphylopine, which is a nicotianamine-like metallophore [17]. This last contains imidazole ring and three carboxylic groups. It is an opine metallophore and can chelate several metal ions (nickel, zinc, cobalt, iron and copper), so it is considered a broad-spectrum metallophore [80]. The import of these wide range of metal ions via staphylopine depends on the metal nature and concentration along with the S. aureus growth status [79]. The structure of staphylopine is shown in Figure 2.
2.2.2. Staphylopine Synthesis
Nine genes in the operon cnt (cntKLMABCDFE) encodes the multiple functions needed for the synthesis and the exportof staphylopine in addition to the import of the complexe (Staphylopine-metal ion). The three genes, cntKLM, encode the needed enzymes for staphylopine biosynthesis CntK, CntL and CntM respectively. The five genes cntABCDF encode the transporter ABC implicated in metal loaded staphylopine import [81,82]. Concerning cntE, it is involved in staphylopine export by encoding transport protein located in the bacterium membrane. All Cnt genes are most expressed in metal scant medium [80]. The importer protein CntA plays a main role in initiation of metal loaded staphylopine recognition and importation [18,78,79]. The mechanism of recognition and transportation of metal loaded staphylopine at the molecular level was verified by the interdomain change that occurs in CntA conformation upon binding to metal loaded staphylopine loaded [83]. The two CntB and CntC proteins located in membrane form a channel that may have a role in staphylopine loaded transportation [18,79,80]. As for ATP-binding CntD and CntF membrane proteins, they supply the energy needed for transportation [18,78]. The uptake of iron, nickel, zinc and cobalt decreases in S. aureus cntL and cntA-F mutant strains [79]. Zinc represses the transcription from the promoter cntA [18]. S. aureus also has Zur which represses the operon that encodes the two proteins related to ABC transporter [84]. If iron is available, Fur represses cnt genes [82]. These data designate that the expression of cnt gene is limited in zinc and iron rich environment and that Cnt system is controlled by both Fur and Zur [85] so, the synthesis of staphylopine is under negative control by Fur/Zur binding. On the other hand, staphylopine export and staphylopine metal recovery is less repressed by cooperative Fur/Zur repression [85].
Three steps are involved in the biosynthesis of staphylopine [80]. Firstly, D-histidine is produced via CntK which is a histidine racemase. Then, the enzyme CntL, that resembles nicotianamine-synthase, uses D-histidine as a substrate and catalyzes the production of xNA (the name comes from its nicotianamine correlation). The latter in an intermediate that is produced through the addition of aminobutyrate (an S-adenosyl methionine moiety). The last step involves the enzyme CntM that condensates pyruvate with xNA producing staphylopine [79]. CntM has the biochemical characteristics of opine synthase enzyme members [79]. A study done in vitro has found that metals employ several effects on the CntM catalyzed reaction [86]. They noticed that at low concentration of copper and zinc, the reaction was moderately activated but totally inhibited at high concentration. Manganese, on the other hand, was an activator only while nickel and cobalt were inhibitors only so it was proposed that the metal affinity toward xNA and an enzyme inhibitory binding site controlled the activation or inhibition according to the concentration of metals. This regulation of the enzyme involved in staphylopine synthesis is dependent on metal may happen in vivo as well and can could help in the adjustment of the production of metallophore [86].
2.2.3. Staphylopine as Zincophore
Nutritional immunity drastically limits the bioavailability of zinc during bacterial infection [87]. In spite of this essential nutrient restriction, S. aureus remains capable of causing severe disease because it is able to compete for zinc with the host [88]. As previously mentioned, S. aureus has two distinct ABC permease types involved in zinc acquisition, AdcABC and CntABCDF. AdcABC is homologous to ABC permeases associated with direct zinc recruitment, while CntABCDF belongs to the NikA/Opp family of ABC permeases. CntABCDF functions in conjunction with staphylopine to specifically promote zinc acquisition. This indicates that staphylopine functions as a staphylococcal zincophore although it can bind various metals in vitro. In a study performed by Grim et al. [17], they found that in zinc depleted medium, strains lacking the Cnt-staphylopine system and Adc permease had major growth defects and failed specifically in zinc accumulation. These results demonstrated that both systems serve as the major zinc importers of this bacterium. Concerning other metal ions such as Co, Ni, and Cu, they found that they are not physiological substrates of the Cnt-staphylopine system, and it is modestly responsive to Mn and Fe [82] which only exert transcriptional influence in the absence of zinc. These findings suggest that the abundance of zinc is the main regulatory factor that controls the system expression [17].
2.2.4. Important Features of Cnt-Staphylopine System
The Cnt system, as previously mentioned, is essential for the optimal metals import metal-limiting conditions and contributes to S. aureus virulence. The failure to efflux staphylopine results in its intracellular accumulation thus impairing the fitness of S. aureus [89]. A recent study has shown that CntE loss resulted in a stronger virulence defect than other components of the Cnt-staphylopine system, even in zinc restricted tissues. The toxicity associated with intracellular staphylopineaccumulation contributed to the virulence defect of strains lacking CntE, even when S. aureus is zinc starved during infection. Moreover, they noticed that the intracellular accumulation of staphylopine did not increase metal importer expression or altered cellular metal concentrations, suggesting that contrary to prevailing models, the toxicity associated with staphylopine is not strictly due to intracellular chelation of metals [90]. CntK catalyzes the first step of staphylopine synthesis by converting L -histidine to D -histidine in order to provide an essential building block of staphylopine. It was found, by structural modeling, that CntK is specific for histidine, whereas other proteinogenic amino acids, with the exception of arginine, do not show any binding with it. These findings helped in developing powerful antibiotics targeting the staphylopine-mediated metal acquisition process in bacteria via designing irreversible inhibitors [91]. Another study confirmed that during the synthesis of staphylopine, CntL stereoselectively carries out the catalysis of D-histidine and not L-histidine. These findings provided critical structural and mechanistic insights into CntL for a better understanding of of nicotianamine-like metallophores biosynthesis and the discovery of inhibitors of this process [92]. Concerning CntA, responsible for the recognition and transport of diverse solutes, a study was performed to investigate the structural conformation upon staphylopine binding. CntA has a fork-like structure formed by three domains (Ia and Ib and II). It uses a bi-domain architectural form of domain II assisted by inter-domain hinge cluster residues. Important clustered communities regulat the conformational changes in CntA. In addition to open (without staphylopine) and close states (with staphylopine) [83], the fluctuating regions sampled two additional intermediate states that were considered closed or open previously. CntA prefers fluctuating the non-conserved regions rather than conserved where domain II turned out to be rigid and maintains a stable fold. Such findings are important to the researcher in field of drug-designing [93].
As for the regulation of cnt operon, a novel regulator (Rsp) was identified that activates the system, in addition to the metal-dependent Fur and Zur repressors. This regulator is an AraC-type regulator. Rsp activation in S. aureus may act to maintain basal cellular levels of staphylopine to scavenge free metals when needed [94]. It is worth mentioning that the AraC family regulators are an abundant group of transcriptional regulators in bacteria, acting mostly as gene expression activators, that controls diverse cellular functions such as virulence and stress response [95]. A study has reported the establishment of a fast and efficient method for directly converting adenine to guanine in bacterial genomes. A systematic screening that targets the possibly editable adenine sites of S. aureus cntBC locates key residues for metal importation, demonstrating that the application of the system might greatly facilitate the bacterial genomic engineering [96].
3. Conclusions
During bacterial infection, essential nutrients, such as metal ions, are needed by the host in order to fight invading pathogens. The Gram-positive bacterium S. aureus can compete successfully with the host for iron and other metal ions. This is mainly achieved through the secretion of metal chelators, known as metallophores. Four siderophores (staphyloferrin A, staphyloferrin B, staphylobactin and Aureochelin) are synthesized and secreted by S. aureus to sequester iron from the extracellular environment. This bacterium has intake systems specific for these siderophores (ABC transporters), and it has the ability to uptake other microorganisms’ siderophores. Staphylopine is a nicotianamine-like wide spectrum metallophore produced by S. aureus as well. Staphylopine is synthesized by a three-step pathway and is exported and imported by specific membrane transporters. Siderophores are genetically regulated by Fur, while staphylopine is regulated by both Fur and Zur. The identification of the metal ion-acquiring systems utilized by S. aureus provides novel opportunities to distort this pathogen’s ability to compete with the host metal ions and thus minimize its production, enabling the development of new strategies to improve the therapeutic approaches.
Author Contributions
G.G. conceived this work, managed the manuscript writing and corrected it; Z.E. wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The study did not report any data.
Acknowledgments
This work was supported by Islamic University of Lebanon, Center of Research and Development.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Coates, R.; Moran, J.; Horsburgh, M.J. Staphylococci: Colonizers and pathogens of human skin. Future Microbiol. 2014, 9, 75–91. [Google Scholar] [CrossRef]
- Tong, S.Y.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.G., Jr. Staphylococcus aureus infections: Epidemiology, pathophysiology, clinical manifestations, and management. Clin. Microbiol. Rev. 2015, 28, 603–661. [Google Scholar] [CrossRef] [Green Version]
- Van Hal, S.J.; Jensen, S.O.; Vaska, V.L.; Espedido, B.A.; Paterson, D.L.; Gosbell, I.B. Predictors of mortality in Staphylococcus aureus Bacteremia. Clin. Microbiol. Rev. 2012, 25, 362–386. [Google Scholar] [CrossRef] [Green Version]
- Otto, M. Staphylococcal Biofilms. Microbiol. Spectr. 2018, 322, 207–228. [Google Scholar] [CrossRef] [PubMed]
- Turner, N.A.; Sharma-Kuinkel, B.K.; Maskarinec, S.A.; Eichenberger, E.M.; Shah, P.P.; Carugati, M.; Holland, T.L.; Fowler, V.G., Jr. Methicillin-resistant Staphylococcus aureus: An overview of basic and clinical research. Nat. Rev. Microbiol. 2019, 17, 203–218. [Google Scholar] [CrossRef] [PubMed]
- Shockman, G.D.; Barren, J.F. Structure, Function, and Assembly of Cell Walls of Gram-Positive Bacteria. Annu. Rev. Microbiol. 1983, 37, 501–527. [Google Scholar] [CrossRef]
- Lowy, F.D. Staphylococcus aureus infections. N. Engl. J. Med. 1998, 339, 520–532. [Google Scholar] [CrossRef]
- Oliveira, D.; Borges, A.; Simões, M. Staphylococcus aureus Toxins and Their Molecular Activity in Infectious Diseases. Toxins 2018, 10, 252. [Google Scholar] [CrossRef] [Green Version]
- Mulcahy, M.E.; McLoughlin, R.M. Host–bacterial crosstalk determines Staphylococcus aureus nasal colonization. Trends Microbiol. 2016, 24, 872–886. [Google Scholar] [CrossRef]
- Torres Salazar, B.O.; Heilbronner, S.; Peschel, A.; Krismer, B. Secondary Metabolites Governing Microbiome Interaction of Staphylococcal Pathogens and Commensals. Microb. Physiol. 2021, 31, 198–216. [Google Scholar] [CrossRef]
- Garcia, Y.M.; Barwinska-Sendra, A.; Tarrant, E.; Skaar, E.P.; Waldron, K.J.; Kehl-Fie, T.E. A Superoxide Dismutase Capable of Functioning with Iron 472 or Manganese Promotes the Resistance of Staphylococcus aureus to Calprotectin and 473 Nutritional Immunity. PLoS Pathog. 2017, 13, e1006125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hood, M.I.; Skaar, E.P. Nutritional immunity: Transition metals at the pathogen-host interface. Nat. Rev. Microbiol. 2012, 10, 525–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammer, N.D.; Skaar, E.P. Molecular mechanisms of Staphylococcus aureus iron acquisition. Annu. Rev. Microbiol. 2011, 65, 129–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerinot, M.L. Microbial iron transport. Annu. Rev. Microbiol. 1994, 48, 743–772. [Google Scholar] [CrossRef] [PubMed]
- Courcol, R.J.; Trivier, D.; Bissinger, M.C.; Martin, G.R.; Brown, M.R. Siderophore production by Staphylococcus aureus and identification of iron-regulated proteins. Infect. Immun. 1997, 65, 1944–1948. [Google Scholar] [CrossRef] [Green Version]
- Dale, S.E.; Sebulsky, M.T.; Heinrichs, D.E. Involvement of SirABC in iron-siderophore import in Staphylococcus aureus. J. Bacteriol. 2004, 186, 8356–8362. [Google Scholar] [CrossRef] [Green Version]
- Grim, K.P.; San Francisco, B.; Radin, J.N.; Brazel, E.B.; Kelliher, J.L.; Párraga Solórzano, P.K.; Kim, P.C.; McDevitt, C.A.; Kehl-Fie, T.E. The Metallophore Staphylopine Enables Staphylococcus aureus To Compete with the Host for Zinc and Overcome Nutritional Immunity. mBio 2017, 8, e01281-17. [Google Scholar] [CrossRef] [Green Version]
- Remy, L.; Carrière, M.; Derré-Bobillot, A.; Martini, C.; Sanguinetti, M.; Borezée-Durant, E. The Staphylococcus aureus Opp1 ABC transporter imports nickel and cobalt in zinc-depleted conditions and contributes to virulence. Mol. Microbiol. 2013, 87, 730–743. [Google Scholar] [CrossRef]
- Hiron, A.; Posteraro, B.; Carrière, M.; Remy, L.; Delporte, C.; La Sorda, M.; Sanguinetti, M.; Juillard, V.; Borezée-Durant, E. A nickel ABC-transporter of Staphylococcus aureus is involved in urinary tract infection. Mol. Microbiol. 2010, 77, 1246–1260. [Google Scholar] [CrossRef]
- Skaar, E.P.; Schneewind, O. Iron-regulated surface determinants (Isd) of 520 Staphylococcus aureus: Stealing iron from heme. Microbes Infect. 2004, 6, 390–397. [Google Scholar] [CrossRef]
- Mascher, T.; Helmann, J.D.; Unden, G. Stimulus Perception in Bacterial Signal-Transducing 529 Histidine Kinases. Microbiol. Mol. Biol. Rev. 2006, 70, 910–938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gkouvatsos, K.; Papanikolaou, G.; Pantopoulos, K. Regulation of iron transport and the role of transferrin. Biochim. Biophys. Acta 2012, 1820, 188–202. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Wang, W.; Wu, S.; Gao, H. Recent Advances in the Siderophore Biology of Shewanella. Front. Microbiol. 2022, 13, 823758. [Google Scholar] [CrossRef] [PubMed]
- Winterbourn, C.C. Toxicity of iron and hydrogen peroxide: The Fenton reaction. Toxicol. Lett. 1995, 82, 969–974. [Google Scholar] [CrossRef]
- Emerson, D.; Roden, E.; Twining, B.S. The microbial ferrous wheel: Iron cycling in terrestrial, freshwater, and marine environments. Front. Microbiol. 2012, 3, 383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kramer, J.; Özkaya, O.; Kümmerli, R. Bacterial siderophores in community and host interactions. Nat. Rev. Microbiol. 2020, 18, 152–163. [Google Scholar] [CrossRef]
- Sebulsky, M.T.; Hohnstein, D.; Hunter, M.D.; Heinrichs, D.E. Identification and characterization of a membrane permease ivolved in iron-hydroxamate transport in Staphylococcus aureus. J. Bacteriol. 2000, 182, 4394–4400. [Google Scholar] [CrossRef] [Green Version]
- Miethke, M.; Marahiel, M.A. Siderophore-based iron acquisition and pathogen control. Microbiol. Mol. Biol. Rev. 2007, 71, 413–451. [Google Scholar] [CrossRef] [Green Version]
- Cassat, J.E.; Skaar, E.P. Iron in infection and immunity. Cell Host Microbe 2013, 13, 509–519. [Google Scholar] [CrossRef] [Green Version]
- Challis, G.L. A widely distributed bacterial pathway for siderophore biosynthesis independent of nonribosomal peptide synthetases. Chem. Bio. Chem. 2005, 6, 601–661. [Google Scholar] [CrossRef]
- Carroll, C.S.; Moore, M.M. Ironing out siderophore biosynthesis: A review of non-ribosomal peptide synthetase (NRPS)-independent siderophore synthetases. Crit. Rev. Biochem. Mol. Biol. 2018, 53, 356–381. [Google Scholar] [CrossRef] [PubMed]
- Crosa, J.H.; Walsh, C.T. Genetics and assembly line enzymology of siderophore biosynthesis in bacteria. Microbiol. Mol. Biol. Rev. 2002, 66, 223–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.Y.; Janes, B.K.; Passalacqua, K.D.; Pfleger, B.F.; Bergman, N.H.; Liu, H.; Håkansson, K.; Somu, R.V.; Aldrich, C.C.; Cendrowski, S.; et al. Biosynthetic analysis of the petrobactin siderophore pathway from Bacillus anthracis. J. Bacteriol. 2007, 189, 1698–1710. [Google Scholar] [CrossRef] [Green Version]
- Andrews, S.C.; Robinson, A.K.; Rodriguez-Quinones, F. Bacterial iron homeostasis. FEMS Microbiol Rev. 2003, 27, 215–237. [Google Scholar] [CrossRef] [Green Version]
- Sheldon, J.R.; Heinrichs, D.E. Recent developments in understanding the iron acquisition strategies of gram positive pathogens. FEMS Microbiol. Rev. 2015, 39, 592–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faraldo-Gómez, J.D.; Sansom, M.S.P. Acquisition of siderophores in gram-negative bacteria. Nat. Rev. Mol. Cell Biol. 2003, 4, 105–116. [Google Scholar] [CrossRef]
- Schalk, I.J.; Guillon, L. Fate of ferrisiderophores after import across bacterial outer membranes: Different iron release strategies are observed in the cytoplasm or periplasm depending on the siderophore pathways. Amino Acids 2013, 44, 1267–1277. [Google Scholar] [CrossRef]
- Ganne, G.; Brillet, K.; Basta, B.; Roche, B.; Hoegy, F.; Gasser, V.; Schalk, I.J. Iron release from the siderophore pyoverdine in Pseudomonas aeruginosa involves three new actors: FpvC, FpvG, and FpvH. ACS Chem. Biol. 2017, 12, 1056–1065. [Google Scholar] [CrossRef]
- Krewulak, K.D.; Vogel, H.J. Structural biology of bacterial iron uptake. Biochim. Biophys. Acta 2008, 1778, 1781–1804. [Google Scholar] [CrossRef] [Green Version]
- Imperi, F.; Tiburzi, F.; Visca, P. Molecular basis of pyoverdine siderophore recycling in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2009, 106, 20440–20445. [Google Scholar] [CrossRef]
- Lin, H.; Fischbach, M.A.; Liu, D.R.; Walsh, C.T. In vitro characterization of salmochelin and enterobactin trilactone hydrolases IroD, IroE, and Fes. J. Am. Chem. Soc. 2005, 127, 11075–11084. [Google Scholar] [CrossRef] [Green Version]
- Torres, V.J.; Pishchany, G.; Humayun, M.; Schneewind, O.; Skaar, E.P. Staphylococcus aureus IsdB is a hemoglobin receptor required for heme iron utilization. J. Bacteriol. 2006, 188, 8421–8429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baichoo, N.; Wang, T.; Ye, R.; Helmann, J.D. Global analysis of the Bacillus subtilis Fur regulon and the iron starvation stimulon. Mol. Microbiol. 2002, 45, 1613–1629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pishchany, G.; Dickey, S.E.; Skaar, E.P. Subcellular localization of the Staphylococcus aureus heme iron transport components IsdA and IsdB. Infect. Immun. 2009, 77, 2624–2634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reniere, M.L.; Skaar, E.P. Staphylococcus aureus haem oxygenases are differentially regulated by iron and haem. Mol. Microbiol. 2008, 69, 1304–1315. [Google Scholar] [CrossRef] [Green Version]
- Friedman, D.B.; Stauff, D.L.; Pishchany, G.; Whitwell, C.W.; Torres, V.J.; Skaar, E.P. Staphylococcus aureus Redirects Central Metabolism to Increase Iron Availability. PLoS Pathog. 2006, 2, e87. [Google Scholar] [CrossRef]
- Chavakis, T.; Hussain, M.; Kanse, S.M.; Peters, G.; Bretzel, R.G.; Flock, J.-I.; Herrmann, M.; Preissner, K.T. Staphylococcus aureus extracellular adherence protein serves as anti-inflammatory factor by inhibiting the recruitment of host leukocytes. Nat. Med. 2002, 8, 687–693. [Google Scholar] [CrossRef]
- Johnson, M.; Cockayne, A.; Morrissey, J.A. Iron-regulated biofilm formation in Staphylococcus aureus Newman requires ica and the secreted protein Emp. Infect. Immun. 2008, 76, 1756–1765. [Google Scholar] [CrossRef] [Green Version]
- Bagg, A.; Neilands, J.B. Ferric uptake regulation protein acts as a repressor, employing iron(II) as a cofactor to bind the operator in an iron transport operon in Escherichia coli. Biochemistry 1987, 26, 5471–5477. [Google Scholar] [CrossRef]
- Horsburgh, M.J.; Clements, M.O.; Crossley, H.; Ingram, E.; Foster, S.J. PerR controls oxidative stress resistance and iron storage proteins and is required for virulence in Staphylococcus aureus. Infect. Immun. 2001, 69, 744–3754. [Google Scholar] [CrossRef]
- Conroy, B.S.; Grigg, J.C.; Kolesnikov, M.; Morales, L.D.; Murphy, M.E. Staphylococcus aureus heme and siderophore-iron acquisition pathways. Biometals 2019, 32, 409–424. [Google Scholar] [CrossRef] [PubMed]
- Beasley, F.C.; Vinés, E.D.; Grigg, J.C.; Zheng, Q.; Liu, S.; Lajoie, G.A.; Murphy, M.E.P.; Heinrichs, D.E. Characterization of staphyloferrin A biosynthetic and transport mutants in Staphylococcus aureus. Mol. Microbiol. 2009, 72, 947–963. [Google Scholar] [CrossRef] [PubMed]
- Beasley, F.C.; Heinrichs, D.E. Siderophore-mediated iron acquisition in the staphylococci. J. Inorg. Biochem. 2010, 104, 282–288. [Google Scholar] [CrossRef] [PubMed]
- Hannauer, M.; Sheldon, J.R.; Heinrichs, D.E. Involvement of major facilitator superfamily proteins SfaA and SbnD in staphyloferrin secretion in Staphylococcus aureus. FEBS Lett. 2015, 589, 730–737. [Google Scholar] [CrossRef] [Green Version]
- Cotton, J.L.; Tao, J.; Balibar, C.J. Identification and characterization of the Staphylococcus aureus gene cluster coding for staphyloferrin A. Biochemistry 2009, 48, 1025–1035. [Google Scholar] [CrossRef]
- Sheldon, J.R.; Marolda, C.L.; Heinrichs, D.E. TCA cycle activity in Staphylococcus aureus is essential for ironregulated synthesis of staphyloferrin A, but not staphyloferrin B: The benefit of a second citrate synthase. Mol. Microbiol. 2014, 92, 824–839. [Google Scholar] [CrossRef]
- Hannauer, M.; Arifin, A.J.; Heinrichs, D.E. Involvement of reductases IruO and NtrA in iron acquisition by Staphylococcus aureus. Mol. Microbiol. 2015, 96, 1192–1210. [Google Scholar] [CrossRef]
- Perera, V.R.; Newton, G.L.; Pogliano, K. Bacillithiol: A key protective thiol in Staphylococcus aureus. Expert Rev. Anti-infect. Ther. 2015, 13, 1089–1107. [Google Scholar] [CrossRef] [Green Version]
- Dale, S.E.; Doherty-Kirby, A.; Lajoie, G.; Heinrichs, D.E. Role of siderophore biosynthesis in virulence of Staphylococcus aureus: Identification and characterization of genes involved in production of a siderophore. Infect. Immun. 2004, 72, 29–37. [Google Scholar] [CrossRef] [Green Version]
- Cheung, J.; Beasley, F.C.; Liu, S.; Lajoie, G.A.; Heinrichs, D.E. Molecular characterization of staphyloferrin B biosynthesis in Staphylococcus aureus. Mol. Microbiol. 2009, 74, 594–608. [Google Scholar] [CrossRef]
- Allard, M.; Moisan, H.; Brouillette, E.; Gervais, A.L.; Jacques, M.; Lacasse, P.; Diarra, M.S.; Malouin, F. Transcriptional modulation of some Staphyloccocus aureus iron-regulated genes during growth in vitro and in a tissue cage model in vivo. Microbes Infect. 2006, 8, 1679–1690. [Google Scholar] [CrossRef] [PubMed]
- Kobylarz, M.J.; Grigg, J.C.; Liu, Y.; Lee, M.S.F.; Heinrichs, D.E.; Murphy, M.E.P. Deciphering the substrate specificity of SbnA, the enzyme catalyzing the first step in Staphyloferrin B biosynthesis. Biochemistry 2016, 55, 927–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laakso, H.A.; Marolda, C.L.; Pinter, T.B.; Stillman, M.J.; Heinrichs, D.E. A heme-responsive regulator controls synthesis of staphyloferrin B in Staphylococcus aureus. J. Biol. Chem. 2016, 291, 29–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verstraete, M.M.; Perez-Borrajero, C.; Brown, K.L.; Heinrichs, D.E.; Murphy, M.E.P. SbnI is a free serine kinase that generates O-phospho-L-serine for staphyloferrin B biosynthesis in Staphylococcus aureus. J. Biol. Chem. 2018, 293, 6147–6160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Speziali, C.D.; Dale, S.E.; Henderson, J.A.; Vine’s, E.D.; Heinrichs, D.E. Requirement of Staphylococcus aureus ATP-binding cassette-ATPase FhuC for iron-restricted growth and evidence that it functions with more than one iron transporter. J. Bacteriol. 2006, 188, 2048–2055. [Google Scholar] [CrossRef] [Green Version]
- Grigg, J.C.; Cheung, J.; Heinrichs, D.E.; Murphy, M.E.P. Specificity of staphyloferrin B recognition by the SirA receptor from Staphylococcus aureus. J. Biol. Chem. 2010, 285, 34579–34588. [Google Scholar] [CrossRef] [Green Version]
- Wysocki, P.; Lisiecki, P.; Mikucki, J. Receptors for endogenous and heterogeneous siderophores in Staphylococcus aureus B 471. Pol. J. Microbiol. 2005, 54, 97–103. [Google Scholar]
- Sebulsky, M.T.; Speziali, C.D.; Shilton, B.H.; Edgell, D.R.; Heinrichs, D.E. FhuD1, a ferric hydroxamate-binding lipoprotein in Staphylococcus aureus: A case of gene duplication and lateral transfer. J. Biol. Chem. 2004, 279, 53152–53159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabrera, G.; Xiong, A.; Uebel, M.; Singh, V.K.; Jayaswal, R.K. Molecular characterization of the iron-hydroxamate uptake system in Staphylococcus aureus. Appl. Environ. Microbiol. 2001, 67, 1001–1003. [Google Scholar] [CrossRef] [Green Version]
- Sebulsky, M.T.; Heinrichs, D.E. Identification and characterization of fhuD1 and fhuD2, two genes involved in iron-hydroxamate uptake in Staphylococcus aureus. J. Bacteriol. 2001, 183, 4994–5000. [Google Scholar] [CrossRef] [Green Version]
- Sebulsky, M.T.; Shillton, B.H.; Speziali, D.; Heinrichs, D.E. The role of FhuD2 in iron(III)-hydroxamate transport in Staphylococcus aureus. Demonstration that FhuD2 binds iron(III)-hydroxamamtes but with minimal conformational change and implications of mutations on transport. J. Biol. Chem. 2003, 278, 49890–49900. [Google Scholar] [CrossRef] [PubMed]
- Beasley, F.C.; Marolda, C.L.; Cheung, J.; Buac, S.; Heinrichs, D.E. Staphylococcus aureus transporters Hts, Sir, and Sst capture iron liberated from human transferrin by staphyloferrin A, staphyloferrin B, and catecholamine stress hormones, respectively, and contribute to virulence. Infect. Immun. 2011, 79, 2345–2355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrissey, J.A.; Cockayne, A.; Hill, P.J.; Williams, P. Molecular cloning and analysis of a putative siderophore ABC transporter from Staphylococcus aureus. Infect. Immun. 2000, 68, 6281–6288. [Google Scholar] [CrossRef] [PubMed]
- Beasley, F.C.; Cheung, J.; Heinrichs, D.E. Mutation of L-2,3- diaminopropionic acid synthase genes blocks staphyloferrin B synthesis in Staphylococcus aureus. BMC Microbiol. 2011, 11, 199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rozalska, B.; Lisiecki, P.; Sadowska, B.; Mikucki, J.; Rudnicka, W. The virulence of Staphylococcus aureus isolates differing by siderophore production. Acta Microbiol. Pol. 1998, 47, 185–194. [Google Scholar] [PubMed]
- Lebrette, H.; Brochier-Armanet, C.; Zambelli, B.; de Reuse, H.; Borezée-Durant, E.; Ciurli, S.; Cavazza, C. Promiscuous nickel import in human pathogens: Structure, thermodynamics, and evolution of extracytoplasmic nickel-binding proteins. Structure 2014, 22, 1421–1432. [Google Scholar] [CrossRef] [Green Version]
- Eitinger, T.; Suhr, J.; Moore, L.; Smith, J.A. Secondary transporters for nickel and cobalt ions: Theme and variations. Biometals 2005, 18, 399–405. [Google Scholar] [CrossRef]
- Lebrette, H.; Borezée-Durant, E.; Martin, L.; Richaud, P.; Erba, E.B.; Cavazza, C. Novel insights into nickel import in Staphylococcus aureus: The positive role of free histidine and structural characterization of a new thiazolidine- type nickel chelator. Metallomics 2015, 7, 613–621. [Google Scholar] [CrossRef]
- Ghssein, G.; Brutesco, C.; Ouerdane, L.; Fojcik, C.; Izaute, A.; Wang, S.; Hajjar, C.; Lobinski, R.; Lemaire, D.; Richaud, P.; et al. Biosynthesis of a broad-spectrum nicotianamine-like metallophore in Staphylococcus aureus. Science 2016, 352, 1105–1109. [Google Scholar] [CrossRef]
- Ghssein, G.; Matar, S.F. Chelating Mechanisms of Transition Metals by Bacterial Metallophores “Pseudopaline and Staphylopine”: A Quantum Chemical Assessment. Computation 2018, 6, 56. [Google Scholar] [CrossRef] [Green Version]
- Hiron, A.; Borezee-Durant, E.; Piard, J.C.; Juillard, V. Only one of four oligopeptide transport systems mediates nitrogen nutrition in Staphylococcus aureus. J. Bacteriol. 2007, 189, 5119–5129. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Fu, Y.; Lee, J.C.; Hooper, D.C. Staphylococcus aureus NorD, a putative efflux pump coregulated with the Opp1 oligopeptide permease, contributes selectively to fitness in vivo. J. Bacteriol. 2012, 194, 6586–6593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, L.; Zhang, Y.; Chen, W.; Gu, T.; Zhang, S.Y.; Ji, Q. Mechanistic insights into staphylopine-mediated metal acquisition. Proc. Natl. Acad. Sci. USA 2018, 115, 3942–3947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindsay, J.A.; Foster, S.J. zur: A Zn(21)-responsive regulatory element of Staphylococcus aureus. Microbiology 2001, 147, 1259–1266. [Google Scholar] [CrossRef] [Green Version]
- Fojcik, C.; Arnoux, P.; Ouerdane, L.; Aigle, M.; Alfonsi, L.; Borezée-Durant, E. Independent and Cooperative Regulation of Staphylopine Biosynthesis and Trafficking by Fur and Zur. Mol. Microbiol. 2018, 108, 159–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hajjar, C.; Fanelli, R.; Laffont, C.; Brutesco, C.; Cullia, G.; Tribout, M.; Nurizzo, D.; Borezée-Durant, E.; Voulhoux, R.; Pignol, D.; et al. Control by Metals of Staphylopine Dehydrogenase Activity during Metallophore Biosynthesis. Am. Chem. Soc. 2019, 141, 5555–5562. [Google Scholar] [CrossRef]
- Corbin, B.D.; Seeley, E.H.; Raab, A.; Feldmann, J.; Miller, M.R.; Torres, V.J.; Anderson, K.L.; Dattilo, B.M.; Dunman, P.M.; Gerads, R.; et al. Metal chelation and inhibition of bacterial growth tissue abscesses. Science 2008, 319, 962–965. [Google Scholar] [CrossRef]
- Kehl-Fie, T.E.; Zhang, Y.; Moore, J.L.; Farrand, A.J.; Hood, M.I.; Rathi, S.; Chazin, W.J.; Caprioli, R.M.; Skaar, E.P. MntABC and MntH contribute to systemic Staphylococcus aureus infection by competing with calprotectin for nutrient manganese. Infect. Immun. 2013, 81, 3395–3405. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Hooper, D.C. Intracellular accumulation of staphylopine impairs the fitness of Staphylococcus aureus cntE mutant. FEBS Lett. 2019, 593, 1213–1222. [Google Scholar] [CrossRef]
- Grim, K.P.; Radin, J.N.; Solórzano, P.K.P.; Morey, J.R.; Frye, K.A.; Ganio, K.; Neville, S.L.; McDevitt, C.A.; Kehl-Fie, T.E. Intracellular Accumulation of Staphylopine Can Sensitize Staphylococcus aureus to Host-Imposed Zinc Starvation by Chelation-Independent Toxicity. J. Bacteriol. 2020, 202, 00014–00020. [Google Scholar] [CrossRef]
- Luo, S.; Ju, Y.; Zhou, J.; Gu, Q.; Xu, J.; Zhou, H. Crystal structure of CntK, the cofactor-independent histidine racemase in staphylopine-mediated metal acquisition of Staphylococcus aureus. Int. J. Biol. Macromol. 2019, 135, 725–733. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Luo, S.; Ju, Y.; Ding, P.; Xu, J.; Gu, Q.; Zhou, H. Structural insights into the ligand recognition and catalysis of the key aminobutanoyltransferase CntL in staphylopine biosynthesis. FASEB J. 2021, 5, 21575. [Google Scholar] [CrossRef] [PubMed]
- Abideen, Z.U.; Ahmad, A.; Usman, M.; Majaz, S.; Ali, W.; Noreen, S.; Mahmood, T.; Nouroz, F. Dynamics and conformational propensities of staphylococcal CntA. J. Biomol. Struct. Dyn. 2021, 39, 4923–4935. [Google Scholar] [CrossRef] [PubMed]
- Vinué, L.; Hooper, D.C. Rsp activates expression of the Cnt system in Staphylococcus aureus. BMC Microbiol. 2020, 20, 327. [Google Scholar] [CrossRef]
- Kotecka, K.; Kawalek, A.; Kobylecki, K.; Bartosik, A.A. The AraC-Type Transcriptional Regulator GliR (PA3027) Activates Genes of Glycerolipid Metabolism in Pseudomonas aeruginosa. Int. J. Mol. Sci. 2021, 22, 5066. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, H.; Wang, Z.; Wu, Z.; Wang, Y.; Tang, N.; Xu, X.; Zhao, S.; Chen, W.; Ji, Q. Programmable adenine deamination in bacteria using a Cas9-adenine-deaminase fusion. Chem. Sci. 2020, 6, 1657–1664. [Google Scholar] [CrossRef]
Figure 1.
The structure of the two siderophores: Staphyloferrin A and B.
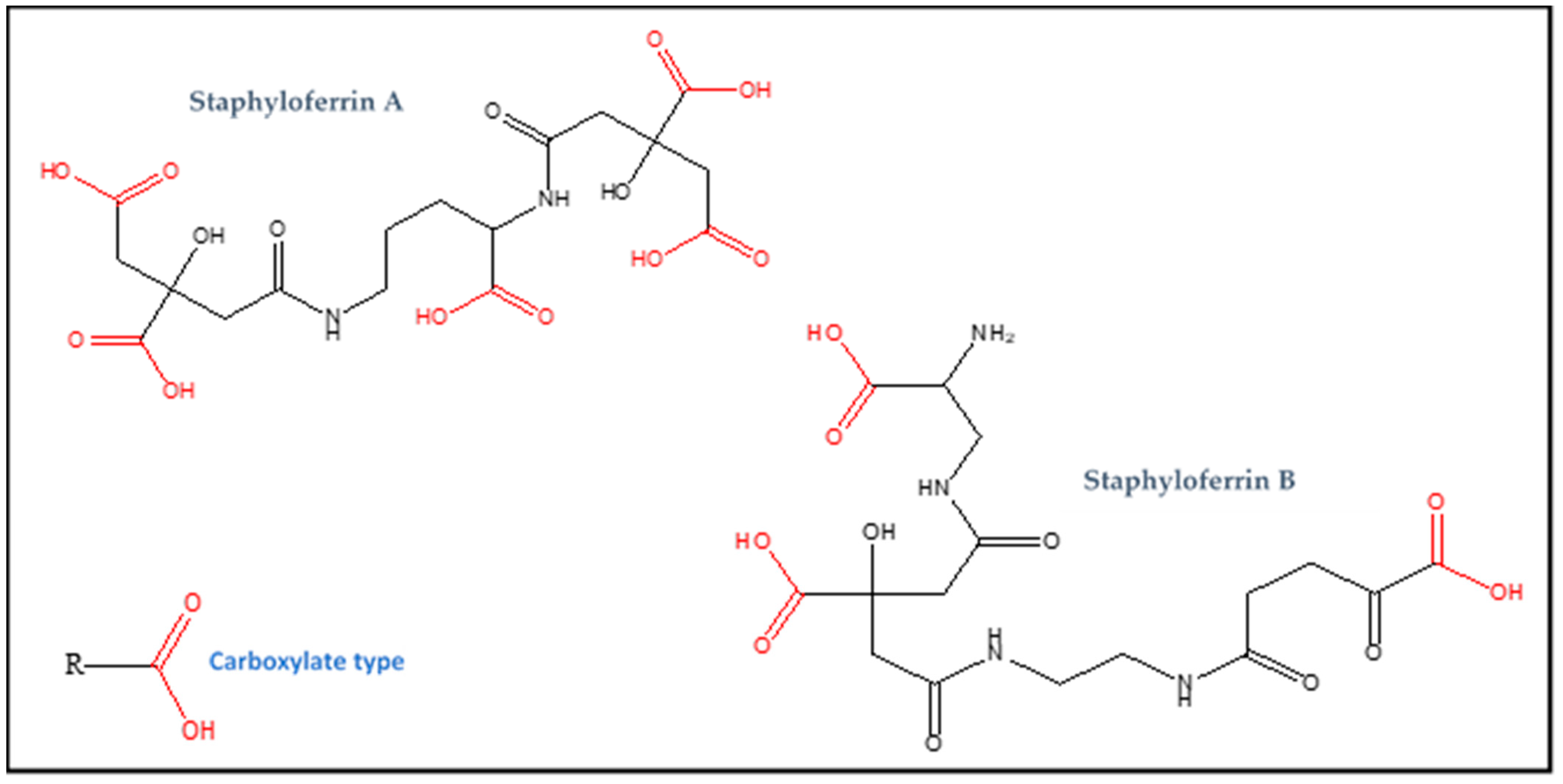
Figure 2.
The structure of metallophore staphylopine.


{kind=link}
{kind=link}
Table 1.
Different types of metallophores produced by S. aureus.
| Name | Type (Functional Group) | Molecular Formula | Molecular Weight (Da) | Specific for |
|---|---|---|---|---|
| Staphyloferrin A | Carboxylate (RCOOH) | C17H24N2O14 | 480 | Iron |
| Staphyloferrin B | Carboxylate (RCOOH) | C16H24N4O11 | 448 | Iron |
| Staphylobactin | Hydroxamate (R-CO-NH-OH) | unknown | 822 | Iron |
| Aureochelin | hydroxamate and catechol (R-CO-NH-OH and C6H4(OH)2) | unknown | 577 | Iron |
| Staphylopine | opine (amine and carboxylic acid) | C13H19N4O6− | 327 | broad-spectrum metallophore (nickel, zinc, cobalt, iron and copper) |
Table 2.
S. aureus metallophores regulation and transportation.
| Name | Operon | Metal Uptake Regulator | Membrane Transporter |
|---|---|---|---|
| Staphyloferrin A | sfaABCD | Fur | HtsABC |
| Staphyloferrin B | sbn (sbnABCDEFGHI) | Fur | SirABC |
| Staphylobactin | sbn | Fur | SirABC |
| Staphylopine | cnt (cntKLMABCDFE) | Fur and Zur | CntB and CntC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ghssein, G.; Ezzeddine, Z. The Key Element Role of Metallophores in the Pathogenicity and Virulence of Staphylococcus aureus: A Review. Biology 2022, 11, 1525. https://doi.org/10.3390/biology11101525
AMA Style
Ghssein G, Ezzeddine Z. The Key Element Role of Metallophores in the Pathogenicity and Virulence of Staphylococcus aureus: A Review. Biology. 2022; 11(10):1525. https://doi.org/10.3390/biology11101525
Chicago/Turabian StyleGhssein, Ghassan, and Zeinab Ezzeddine. 2022. "The Key Element Role of Metallophores in the Pathogenicity and Virulence of Staphylococcus aureus: A Review" Biology 11, no. 10: 1525. https://doi.org/10.3390/biology11101525
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.