
Susceptibility of Beavers to Chronic Wasting Disease

 ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
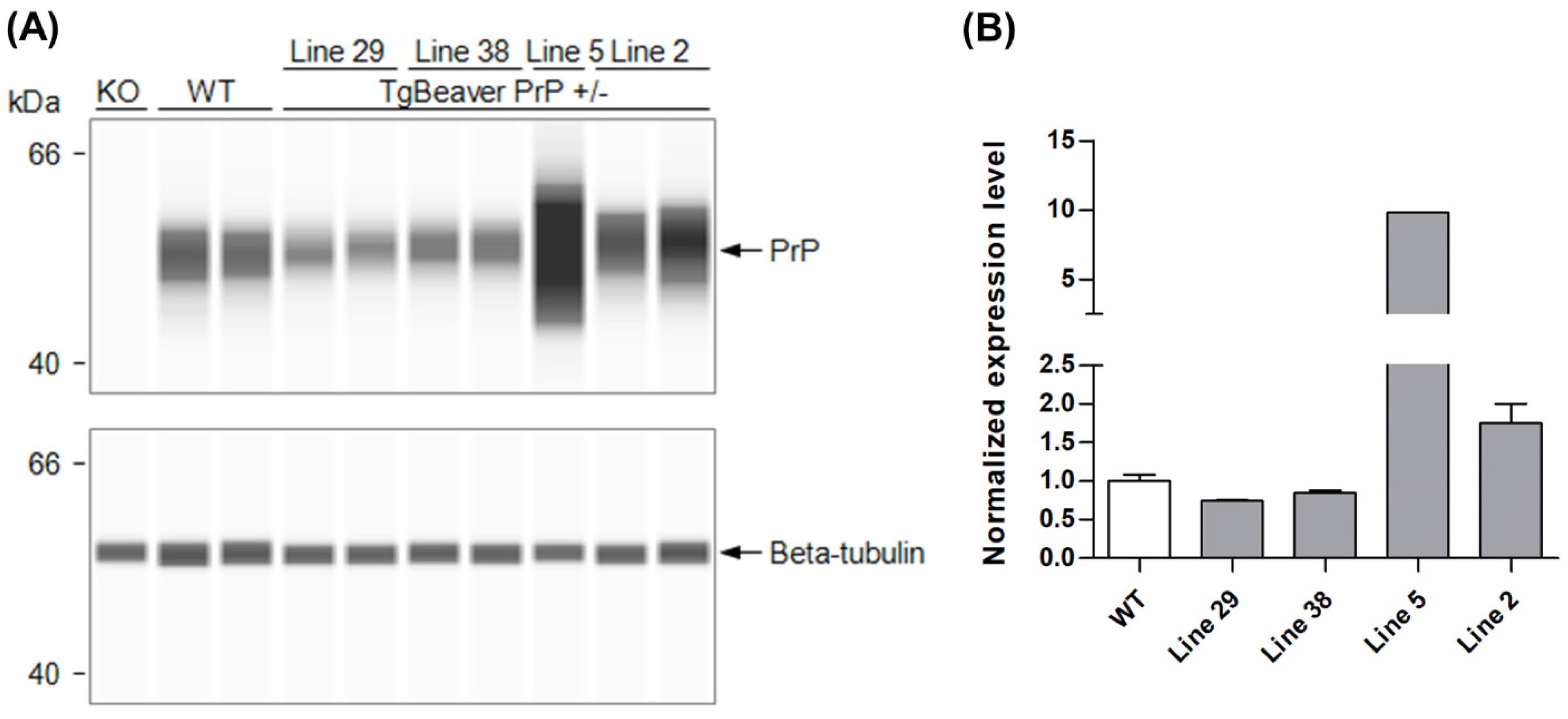
2.1. Creation of Beaver Prnp Transgenic Mice (tgBeaver)
2.2. Bioassay of CWD and Other Prions in tgBeaver Mice
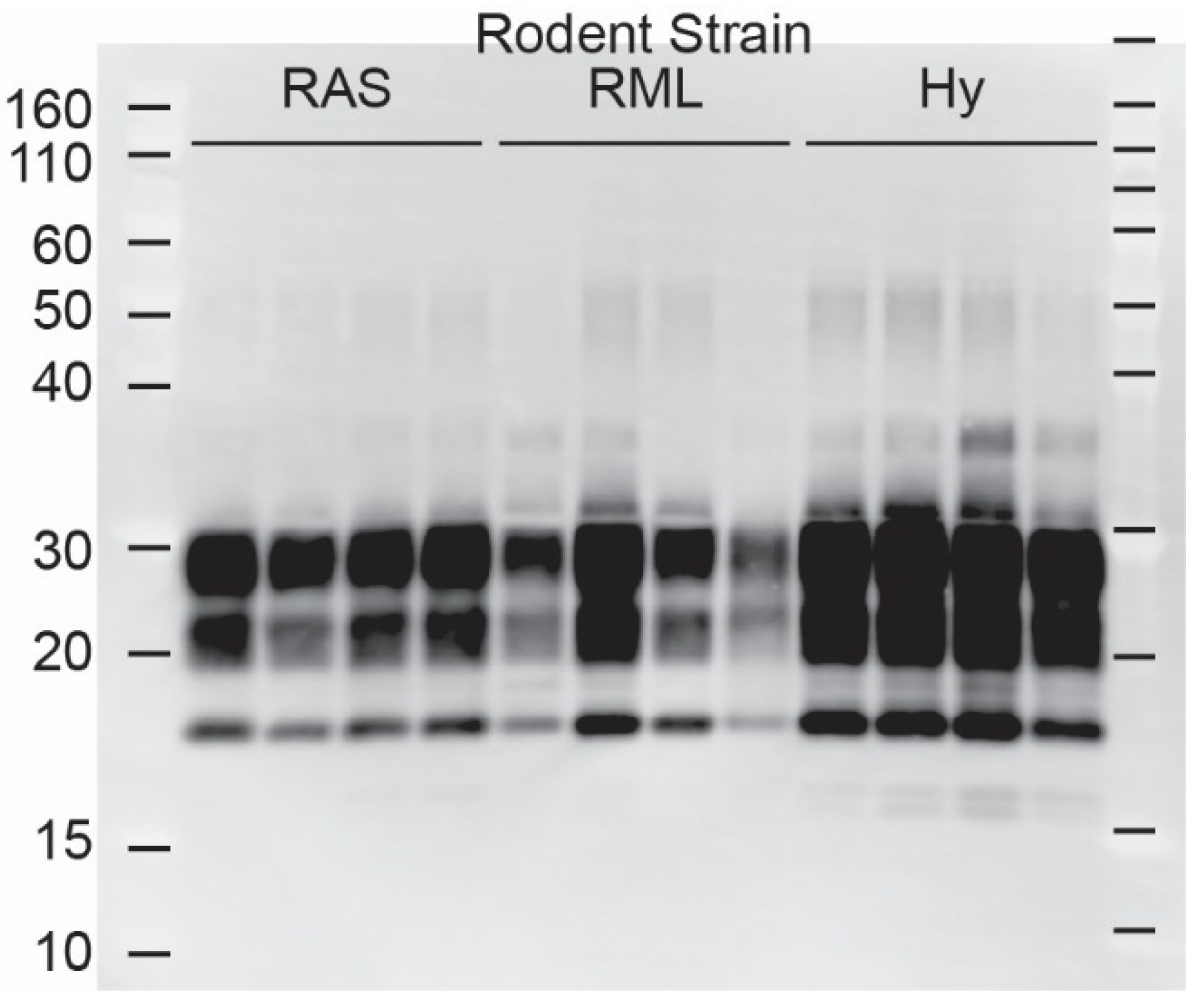
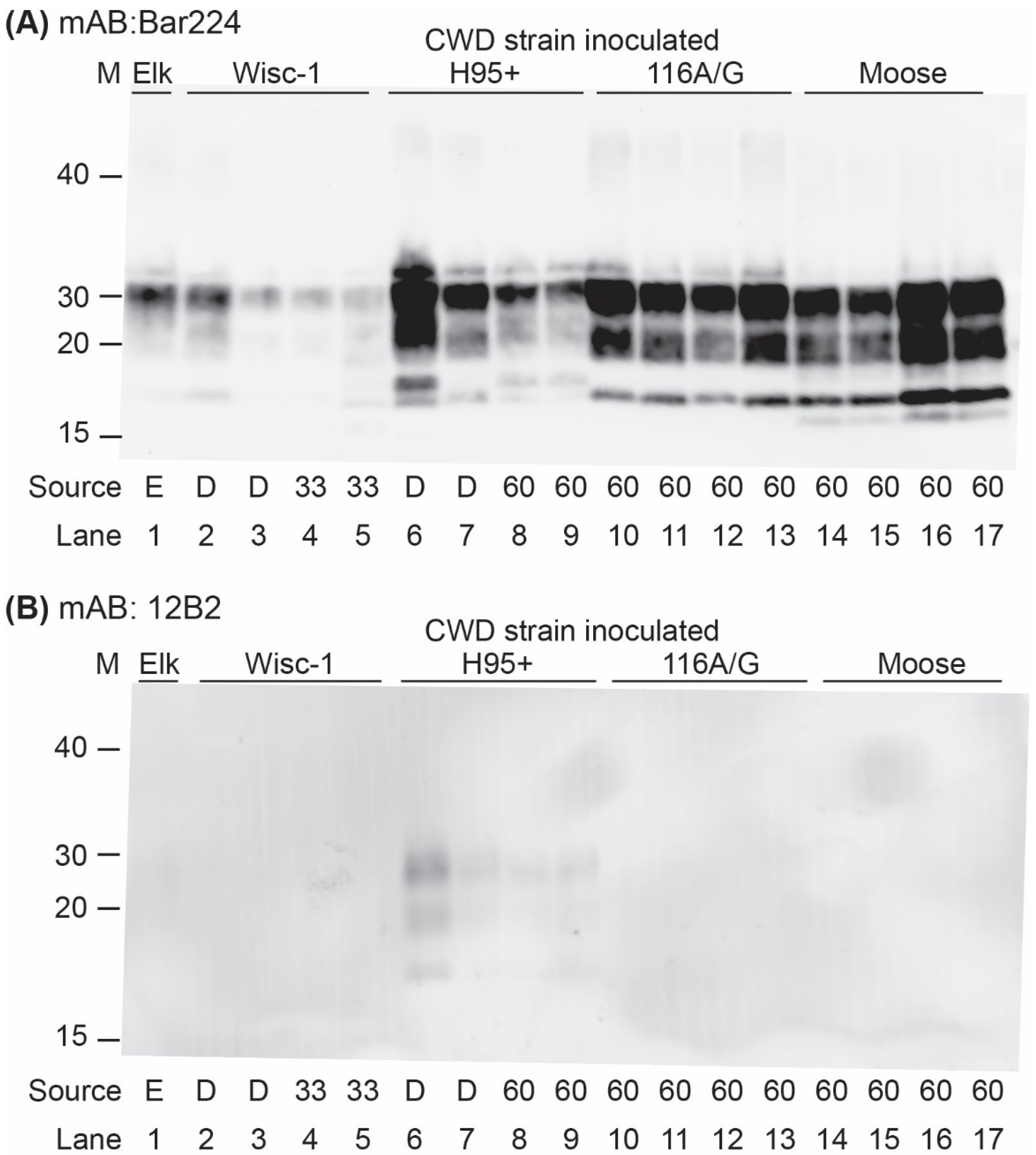
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Williams, E.S.; Young, S. Chronic wasting disease of captive mule deer: A spongiform encephalopathy. J. Wildl. Dis. 1980, 16, 89–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richardson, B. Distribution of Chronic Wasting Disease in North America. Available online: https://www.usgs.gov/centers/nwhc/science/expanding-distribution-chronic-wasting-disease (accessed on 8 February 2022).
- Kim, T.Y.; Shon, H.J.; Joo, Y.S.; Mun, U.K.; Kang, K.S.; Lee, Y.S. Additional cases of Chronic Wasting Disease in imported deer in Korea. J. Vet. Med. Sci. 2005, 67, 753–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benestad, S.L.; Mitchell, G.; Simmons, M.; Ytrehus, B.; Vikoren, T. First case of chronic wasting disease in Europe in a Norwegian free-ranging reindeer. Vet. Res. 2016, 47, 88. [Google Scholar] [CrossRef] [PubMed]
- Pirisinu, L.; Tran, L.; Chiappini, B.; Vanni, I.; Di Bari, M.A.; Vaccari, G.; Vikoren, T.; Madslien, K.I.; Vage, J.; Spraker, T.; et al. Novel Type of Chronic Wasting Disease Detected in Moose (Alces alces), Norway. Emerg. Infect. Dis. 2018, 24, 2210–2218. [Google Scholar] [CrossRef] [Green Version]
- Vikoren, T.; Vage, J.; Madslien, K.I.; Roed, K.H.; Rolandsen, C.M.; Tran, L.; Hopp, P.; Veiberg, V.; Heum, M.; Moldal, T.; et al. First Detection of Chronic Wasting Disease in a Wild Red Deer (Cervus elaphus) in Europe. J. Wildl. Dis. 2019, 55, 970–972. [Google Scholar] [CrossRef]
- Rivera, N.A.; Brandt, A.L.; Novakofski, J.E.; Mateus-Pinilla, N.E. Chronic Wasting Disease In Cervids: Prevalence, Impact And Management Strategies. Vet. Med. 2019, 10, 123–139. [Google Scholar] [CrossRef] [Green Version]
- Bartz, J.C. Environmental and host factors that contribute to prion strain evolution. Acta Neuropathol. 2021, 142, 5–16. [Google Scholar] [CrossRef]
- Duque Velasquez, C.; Kim, C.; Haldiman, T.; Herbst, A.; Aiken, J.; Safar, J.G.; McKenzie, D. Chronic wasting disease (CWD) prion strains evolve via adaptive diversification of conformers in hosts expressing prion protein polymorphisms. J. Biol. Chem. 2020, 295, 4985–5001. [Google Scholar] [CrossRef] [Green Version]
- Herbst, A.; Velasquez, C.D.; Triscott, E.; Aiken, J.M.; McKenzie, D. Chronic Wasting Disease Prion Strain Emergence and Host Range Expansion. Emerg. Infect. Dis. 2017, 23, 1598–1600. [Google Scholar] [CrossRef] [Green Version]
- Bruce, M.E.; Will, R.G.; Ironside, J.W.; McConnell, I.; Drummond, D.; Suttie, A.; McCardle, L.; Chree, A.; Hope, J.; Birkett, C.; et al. Transmissions to mice indicate that ‘new variant’ CJD is caused by the BSE agent. Nature 1997, 389, 498–501. [Google Scholar] [CrossRef]
- Bruce, M.E.; Dickinson, A.G. Biological Stability of Different Classes of Scrapie Agent; Prusiner, S.B., Hadlow, W.J., Eds.; Academic Press: New York, NY, USA, 1979; Volume 2, pp. 71–86. [Google Scholar]
- Kimberlin, R.H.; Cole, S.; Walker, C.A. Temporary and permanent modifications to a single strain of mouse scrapie on transmission to rats and hamsters. J. Gen. Virol. 1987, 68 Pt 7, 1875–1881. [Google Scholar] [CrossRef] [PubMed]
- Haley, N.J.; Seelig, D.M.; Zabel, M.D.; Telling, G.C.; Hoover, E.A. Detection of CWD prions in urine and saliva of deer by transgenic mouse bioassay. PLoS ONE 2009, 4, e4848. [Google Scholar] [CrossRef]
- Mathiason, C.K.; Powers, J.G.; Dahmes, S.J.; Osborn, D.A.; Miller, K.V.; Warren, R.J.; Mason, G.L.; Hays, S.A.; Hayes-Klug, J.; Seelig, D.M.; et al. Infectious prions in the saliva and blood of deer with chronic wasting disease. Science 2006, 314, 133–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamguney, G.; Miller, M.W.; Wolfe, L.L.; Sirochman, T.M.; Glidden, D.V.; Palmer, C.; Lemus, A.; DeArmond, S.J.; Prusiner, S.B. Asymptomatic deer excrete infectious prions in faeces. Nature 2009, 461, 529–532. [Google Scholar] [CrossRef] [Green Version]
- Angers, R.C.; Seward, T.S.; Napier, D.; Green, M.; Hoover, E.; Spraker, T.; O’Rourke, K.; Balachandran, A.; Telling, G.C. Chronic wasting disease prions in elk antler velvet. Emerg. Infect. Dis. 2009, 15, 696–703. [Google Scholar] [CrossRef]
- Tennant, J.M.; Li, M.; Henderson, D.M.; Tyer, M.L.; Denkers, N.D.; Haley, N.J.; Mathiason, C.K.; Hoover, E.A. Shedding and stability of CWD prion seeding activity in cervid feces. PLoS ONE 2020, 15, e0227094. [Google Scholar] [CrossRef] [Green Version]
- Brown, P.; Gajdusek, D.C. Survival of scrapie virus after 3 years’ interment. Lancet 1991, 337, 269–270. [Google Scholar] [CrossRef]
- Johnson, C.J.; Pedersen, J.A.; Chappell, R.J.; McKenzie, D.; Aiken, J.M. Oral transmissibility of prion disease is enhanced by binding to soil particles. PLoS Pathog. 2007, 3, e93. [Google Scholar] [CrossRef]
- Nichols, T.A.; Spraker, T.R.; Rigg, T.D.; Meyerett-Reid, C.; Hoover, C.; Michel, B.; Bian, J.; Hoover, E.; Gidlewski, T.; Balachandran, A.; et al. Intranasal inoculation of white-tailed deer (Odocoileus virginianus) with lyophilized chronic wasting disease prion particulate complexed to montmorillonite clay. PLoS ONE 2013, 8, e62455. [Google Scholar] [CrossRef] [Green Version]
- Smith, C.B.; Booth, C.J.; Pedersen, J.A. Fate of prions in soil: A review. J. Environ. Qual. 2011, 40, 449–461. [Google Scholar] [CrossRef]
- Hamir, A.N.; Miller, J.M.; Kunkle, R.A.; Hall, S.M.; Richt, J.A. Susceptibility of cattle to first-passage intracerebral inoculation with chronic wasting disease agent from white-tailed deer. Vet. Pathol. 2007, 44, 487–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamir, A.N.; Kunkle, R.A.; Cutlip, R.C.; Miller, J.M.; Williams, E.S.; Richt, J.A. Transmission of chronic wasting disease of mule deer to Suffolk sheep following intracerebral inoculation. J. Vet. Diagn. Investig. 2006, 18, 558–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cassmann, E.D.; Moore, S.J.; Greenlee, J.J. Experimental Oronasal Transmission of Chronic Wasting Disease Agent from White-Tailed Deer to Suffolk Sheep. Emerg. Infect. Dis. 2021, 27, 3156–3158. [Google Scholar] [CrossRef] [PubMed]
- Moore, S.J.; West Greenlee, M.H.; Kondru, N.; Manne, S.; Smith, J.D.; Kunkle, R.A.; Kanthasamy, A.; Greenlee, J.J. Experimental Transmission of the Chronic Wasting Disease Agent to Swine after Oral or Intracranial Inoculation. J. Virol. 2017, 91, e00926-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathiason, C.K.; Nalls, A.V.; Seelig, D.M.; Kraft, S.L.; Carnes, K.; Anderson, K.R.; Hayes-Klug, J.; Hoover, E.A. Susceptibility of domestic cats to chronic wasting disease. J. Virol. 2013, 87, 1947–1956. [Google Scholar] [CrossRef] [Green Version]
- Moore, S.J.; Smith, J.D.; Richt, J.A.; Greenlee, J.J. Raccoons accumulate PrP(Sc) after intracranial inoculation of the agents of chronic wasting disease or transmissible mink encephalopathy but not atypical scrapie. J. Vet. Diagn. Investig. 2019, 31, 200–209. [Google Scholar] [CrossRef] [Green Version]
- Moore, S.J.; Carlson, C.M.; Schneider, J.R.; Johnson, C.J.; Greenlee, J.J. Increased Attack Rates and Decreased Incubation Periods in Raccoons with Chronic Wasting Disease Passaged through Meadow Voles. Emerg. Infect. Dis. 2022, 28, 793–801. [Google Scholar] [CrossRef]
- Di Bari, M.A.; Nonno, R.; Castilla, J.; D’Agostino, C.; Pirisinu, L.; Riccardi, G.; Conte, M.; Richt, J.; Kunkle, R.; Langeveld, J.; et al. Chronic wasting disease in bank voles: Characterisation of the shortest incubation time model for prion diseases. PLoS Pathog. 2013, 9, e1003219. [Google Scholar] [CrossRef]
- Heisey, D.M.; Mickelsen, N.A.; Schneider, J.R.; Johnson, C.J.; Langenberg, J.A.; Bochsler, P.N.; Keane, D.P.; Barr, D.J. Chronic wasting disease (CWD) susceptibility of several North American rodents that are sympatric with cervid CWD epidemics. J. Virol. 2010, 84, 210–215. [Google Scholar] [CrossRef] [Green Version]
- Clements, G.M.; Hygnstrom, S.E.; Gilsdorf, J.M.; Baasch, D.M.; Clements, M.J.; Vercauteren, K.C. Movements of white-tailed deer in riparian habitat: Implications for infectious diseases. J. Wildl. Manag. 2011, 75, 1436–1442. [Google Scholar] [CrossRef] [Green Version]
- Geist, V.; Clausen, D.; Crichton, V.; Rowledge, D. The Challenge of CWD: Insidious and Dire; Living Legacy White Paper Series; Alliance for Public Wildlife, 2017; Available online: https://www.apwildlife.org/publications/ (accessed on 8 February 2022).
- Kay, C. Impact of native ungukates and beaver on riparian communities in the intermountain west. Nat. Resour. Environ. Issues 1994, 1, 6. [Google Scholar]
- Pritzkow, S.; Morales, R.; Moda, F.; Khan, U.; Telling, G.C.; Hoover, E.; Soto, C. Grass plants bind, retain, uptake, and transport infectious prions. Cell Rep. 2015, 11, 1168–1175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanger, F.; Coulson, A.R. A rapid method for determining sequences in DNA by primed synthesis with DNA polymerase. J. Mol. Biol. 1975, 94, 441–448. [Google Scholar] [CrossRef]
- Fischer, M.; Rulicke, T.; Raeber, A.; Sailer, A.; Moser, M.; Oesch, B.; Brandner, S.; Aguzzi, A.; Weissmann, C. Prion protein (PrP) with amino-proximal deletions restoring susceptibility of PrP knockout mice to scrapie. EMBO J. 1996, 15, 1255–1264. [Google Scholar] [CrossRef] [Green Version]
- Borchelt, D.R.; Davis, J.; Fischer, M.; Lee, M.K.; Slunt, H.H.; Ratovitsky, T.; Regard, J.; Copeland, N.G.; Jenkins, N.A.; Sisodia, S.S.; et al. A vector for expressing foreign genes in the brains and hearts of transgenic mice. Genet. Anal. Biomol. Eng. 1996, 13, 159–163. [Google Scholar] [CrossRef]
- Mercer, R.C.C.; Daude, N.; Dorosh, L.; Fu, Z.L.; Mays, C.E.; Gapeshina, H.; Wohlgemuth, S.L.; Acevedo-Morantes, C.Y.; Yang, J.; Cashman, N.R.; et al. A novel Gerstmann-Straussler-Scheinker disease mutation defines a precursor for amyloidogenic 8 kDa PrP fragments and reveals N-terminal structural changes shared by other GSS alleles. PLoS Pathog. 2018, 14, e1006826. [Google Scholar] [CrossRef]
- Creamer, J.S.; Oborny, N.J.; Lunte, S.M. Recent advances in the analysis of therapeutic proteins by capillary and microchip electrophoresis. Anal. Methods 2014, 6, 5427–5449. [Google Scholar] [CrossRef] [Green Version]
- Castle, A.R.; Daude, N.; Gilch, S.; Westaway, D. Application of high-throughput, capillary-based Western analysis to modulated cleavage of the cellular prion protein. J. Biol. Chem. 2019, 294, 2642–2650. [Google Scholar] [CrossRef] [Green Version]
- Angers, R.C.; Kang, H.E.; Napier, D.; Browning, S.; Seward, T.; Mathiason, C.; Balachandran, A.; McKenzie, D.; Castilla, J.; Soto, C.; et al. Prion strain mutation determined by prion protein conformational compatibility and primary structure. Science 2010, 328, 1154–1158. [Google Scholar] [CrossRef] [Green Version]
- Pushie, M.J.; Shaykhutdinov, R.; Nazyrova, A.; Graham, C.; Vogel, H.J. An NMR metabolomics study of elk inoculated with chronic wasting disease. J. Toxicol. Environ. Health Part A 2011, 74, 1476–1492. [Google Scholar] [CrossRef]
- Hannaoui, S.; Triscott, E.; Duque Velasquez, C.; Chang, S.C.; Arifin, M.I.; Zemlyankina, I.; Tang, X.; Bollinger, T.; Wille, H.; McKenzie, D.; et al. New and distinct chronic wasting disease strains associated with cervid polymorphism at codon 116 of the Prnp gene. PLoS Pathog. 2021, 17, e1009795. [Google Scholar] [CrossRef] [PubMed]
- Chandler, R.L. Encephalopathy in mice produced by inoculation with scrapie brain material. Lancet 1961, 1, 1378–1379. [Google Scholar] [CrossRef]
- Eklund, C.M.; Kennedy, R.C.; Hadlow, W.J. Pathogenesis of scrapie virus infection in the mouse. J. Infect. Dis. 1967, 117, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Herbst, A.; Ness, A.; Johnson, C.J.; McKenzie, D.; Aiken, J.M. Transcriptomic responses to prion disease in rats. BMC Genom. 2015, 16, 682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bessen, R.A.; Marsh, R.F. Identification of two biologically distinct strains of transmissible mink encephalopathy in hamsters. J. Gen. Virol. 1992, 73 Pt 2, 329–334. [Google Scholar] [CrossRef]
- Bartz, J.C.; Bessen, R.A.; McKenzie, D.; Marsh, R.F.; Aiken, J.M. Adaptation and selection of prion protein strain conformations following interspecies transmission of transmissible mink encephalopathy. J. Virol. 2000, 74, 5542–5547. [Google Scholar] [CrossRef] [Green Version]
- Race, B.; Meade-White, K.D.; Miller, M.W.; Barbian, K.D.; Rubenstein, R.; LaFauci, G.; Cervenakova, L.; Favara, C.; Gardner, D.; Long, D.; et al. Susceptibilities of nonhuman primates to chronic wasting disease. Emerg. Infect. Dis. 2009, 15, 1366–1376. [Google Scholar] [CrossRef]
- Meade-White, K.; Race, B.; Trifilo, M.; Bossers, A.; Favara, C.; Lacasse, R.; Miller, M.; Williams, E.; Oldstone, M.; Race, R.; et al. Resistance to chronic wasting disease in transgenic mice expressing a naturally occurring allelic variant of deer prion protein. J. Virol. 2007, 81, 4533–4539. [Google Scholar] [CrossRef] [Green Version]
- Jeffrey, M.; Martin, S.; Thomson, J.R.; Dingwall, W.S.; Begara-McGorum, I.; Gonzalez, L. Onset and distribution of tissue prp accumulation in scrapie-affected suffolk sheep as demonstrated by sequential necropsies and tonsillar biopsies. J. Comp. Pathol. 2001, 125, 48–57. [Google Scholar] [CrossRef]
- Yull, H.M.; Ritchie, D.L.; Langeveld, J.P.; van Zijderveld, F.G.; Bruce, M.E.; Ironside, J.W.; Head, M.W. Detection of type 1 prion protein in variant Creutzfeldt-Jakob disease. Am. J. Pathol. 2006, 168, 151–157. [Google Scholar] [CrossRef] [Green Version]
- Akhtar, S.; Wenborn, A.; Brandner, S.; Collinge, J.; Lloyd, S.E. Sex effects in mouse prion disease incubation time. PLoS ONE 2011, 6, e28741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saunders, S.E.; Bartelt-Hunt, S.L.; Bartz, J.C. Occurrence, transmission, and zoonotic potential of chronic wasting disease. Emerg. Infect. Dis. 2012, 18, 369–376. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Inoculum (Strain, Source) | Attack Rate | Incubation Period (mean ± SD) | ||
|---|---|---|---|---|
| CWD | Male | Female | Male | Female |
| Wisc1, Deer | 1/4 * | 1/4 * | - | - |
| Wisc1, tg33 | 4/6 | 4/6 | 333.3 ± 60.6 | 355.3 ± 76.7 |
| H95+, Deer | 4/4 | 6/7 | 388.5 ± 109.1 | 373.8 ± 83.5 |
| H95+, tg60 | 6/6 | 4/6 | 204.8 ± 24.8 | 291.3 ± 68.0 |
| CWD2, Elk | 1/5 * | 3/5 * | - | - |
| 116AG, tg60 | 3/6 | 7/7 | 398.3 ± 44.4 | 518.3 ± 37.6 |
| Moose, tg60 | 3/5 | 2/7 | 215.3 ± 24.0 | 497.5 ± 50.2 |
| Other prions | ||||
| CJD, tgHuman | 0/4 | 0/1 | - | - |
| RML, mouse | 1/1 | 4/4 | 122 | 260.0 ± 86.8 |
| Hy, hamster | 3/3 | 7/7 | 76.7 ± 16.3 | 115.9 ± 15.3 |
| Dy, hamster | 0/0 | 2/7 * | - | - |
| RAS, rat | 7/7 | 3/3 | 101.3 ± 1.5 | 139.1 ± 34.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Herbst, A.; Wohlgemuth, S.; Yang, J.; Castle, A.R.; Moreno, D.M.; Otero, A.; Aiken, J.M.; Westaway, D.; McKenzie, D. Susceptibility of Beavers to Chronic Wasting Disease. Biology 2022, 11, 667. https://doi.org/10.3390/biology11050667
Herbst A, Wohlgemuth S, Yang J, Castle AR, Moreno DM, Otero A, Aiken JM, Westaway D, McKenzie D. Susceptibility of Beavers to Chronic Wasting Disease. Biology. 2022; 11(5):667. https://doi.org/10.3390/biology11050667
Chicago/Turabian StyleHerbst, Allen, Serene Wohlgemuth, Jing Yang, Andrew R. Castle, Diana Martinez Moreno, Alicia Otero, Judd M. Aiken, David Westaway, and Debbie McKenzie. 2022. "Susceptibility of Beavers to Chronic Wasting Disease" Biology 11, no. 5: 667. https://doi.org/10.3390/biology11050667
APA StyleHerbst, A., Wohlgemuth, S., Yang, J., Castle, A. R., Moreno, D. M., Otero, A., Aiken, J. M., Westaway, D., & McKenzie, D. (2022). Susceptibility of Beavers to Chronic Wasting Disease. Biology, 11(5), 667. https://doi.org/10.3390/biology11050667