Pol IV-Dependent siRNA Production is Reduced in Brassica rapa

Abstract
:1. Introduction
2. Experimental Section
3. Results and Discussion
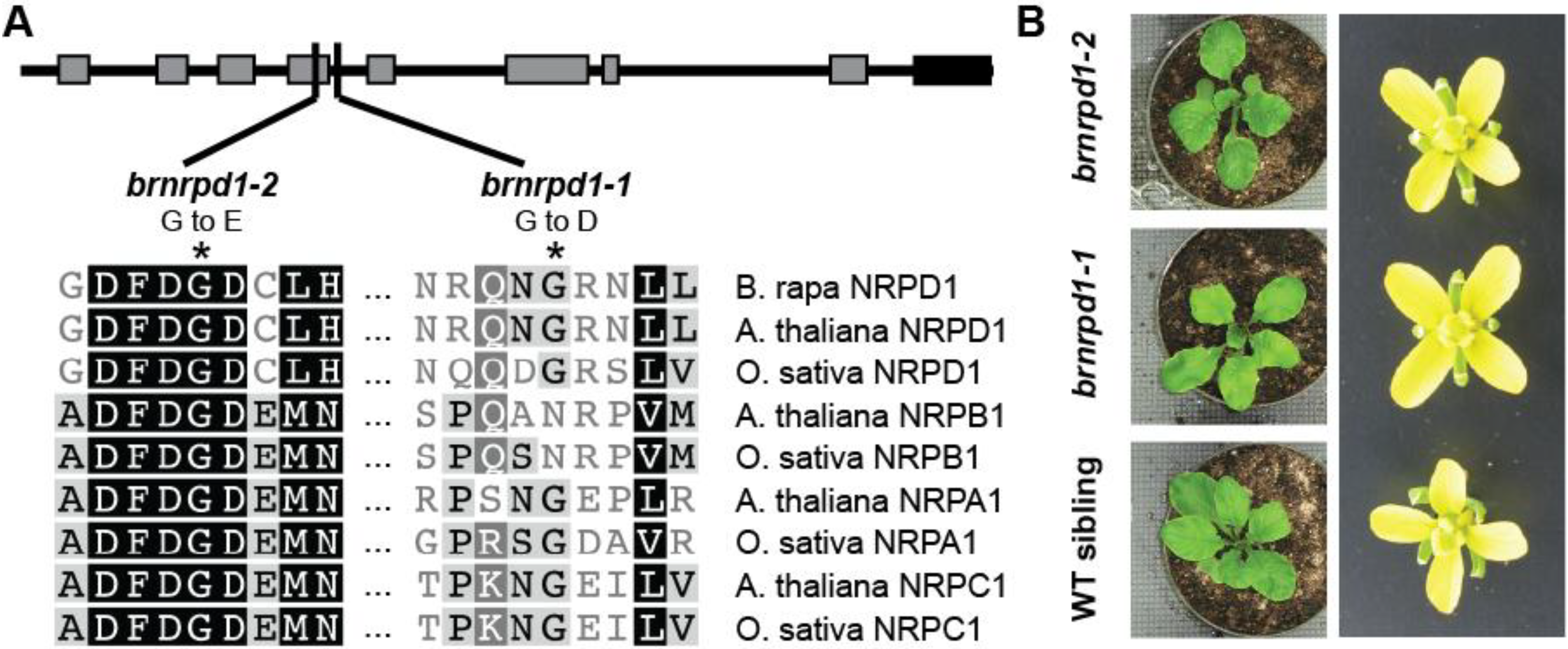
3.1. The Pol IV Pathway in Brassica Rapa

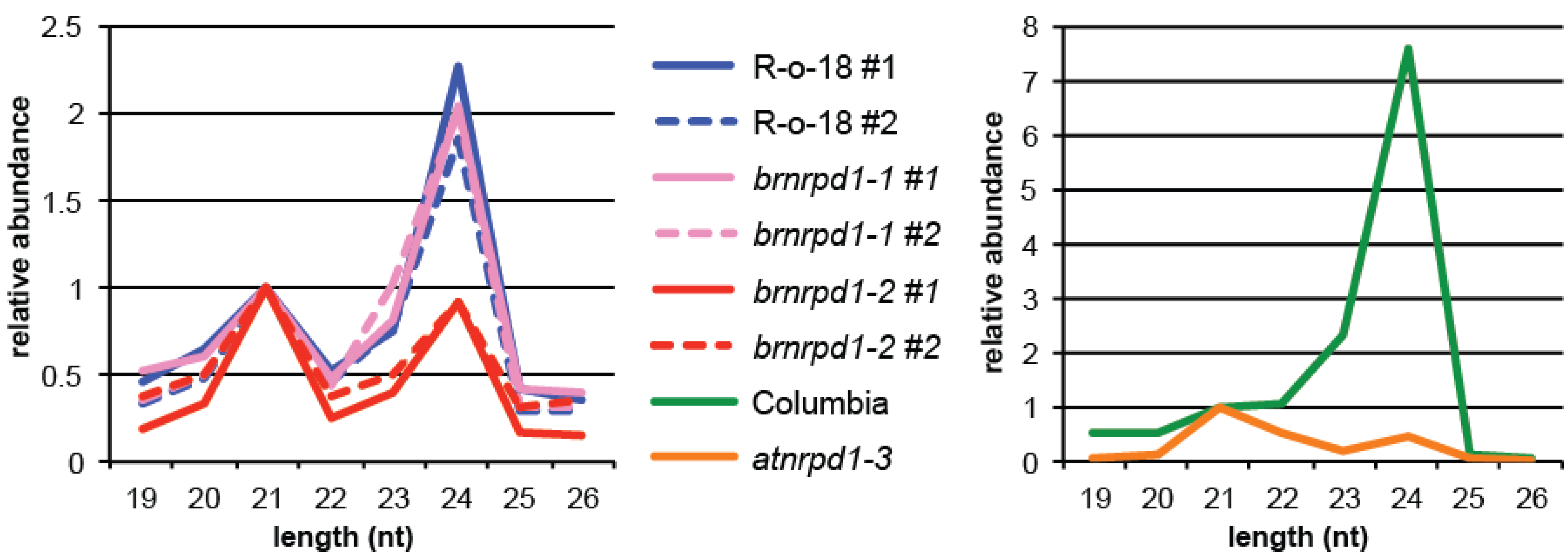
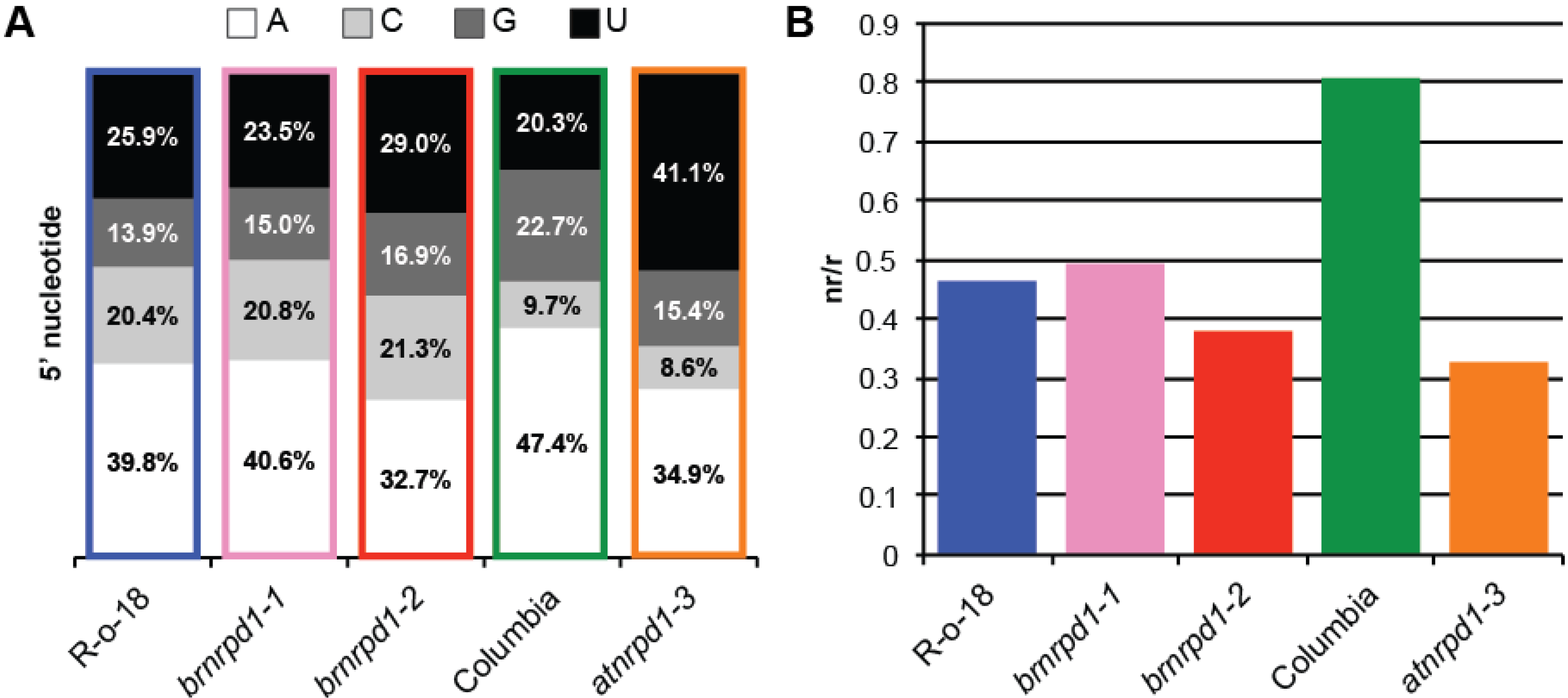
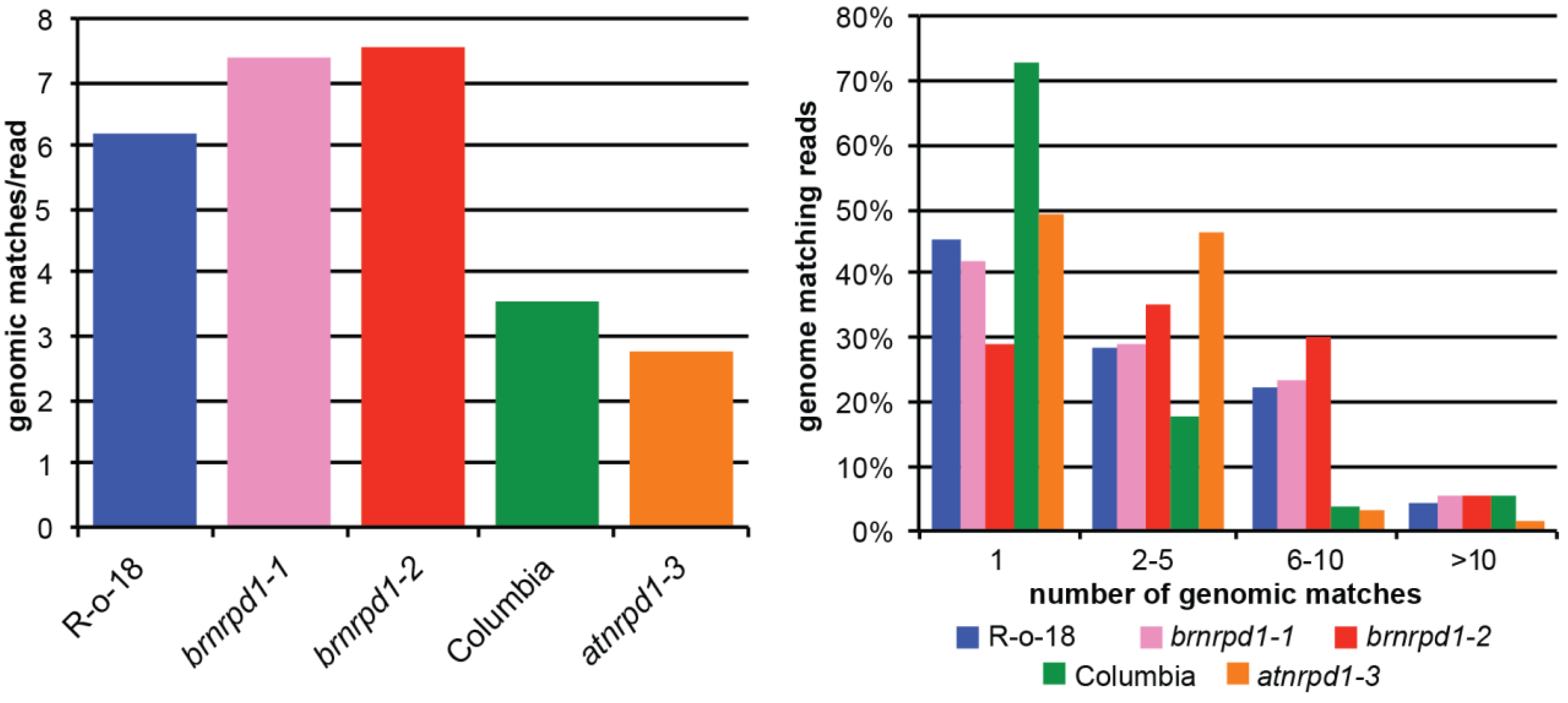
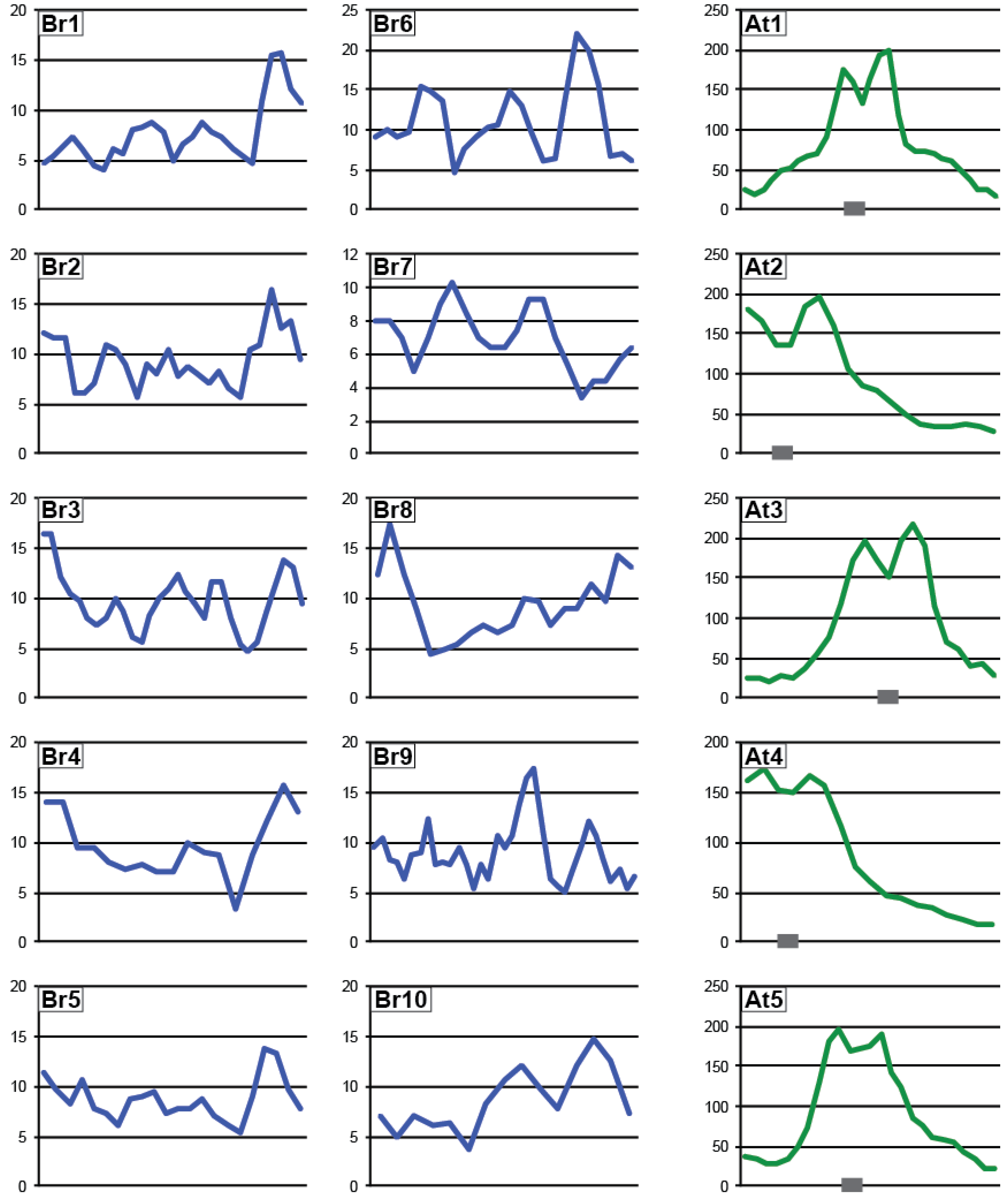
3.2. Small RNA Production in Brassica Rapa

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dataset | Total reads | Filtered | Genome matching | Non-redundant | |||
|---|---|---|---|---|---|---|---|
| R-o-18 #1 | 6,382,869 | 4,188,806 | 65.63% | 2,194,063 | 52.38% | 544,690 | 24.83% |
| R-o-18 #2 | 10,801,569 | 7,019,933 | 64.99% | 3,781,636 | 53.87% | 708,041 | 18.72% |
| brnrpd1-1 #1 | 11,043,788 | 7,183,286 | 65.04% | 3,860,502 | 53.74% | 943,725 | 24.45% |
| brnrpd1-1 #2 | 11,103,304 | 7,265,692 | 65.44% | 3,837,612 | 52.82% | 774,524 | 20.18% |
| brnrpd1-2 #1 | 6,967,097 | 4,676,438 | 67.12% | 2,290,659 | 48.98% | 481,686 | 21.03% |
| brnrpd1-2 #2 | 10,687,391 | 7,233,704 | 67.68% | 3,453,687 | 47.74% | 553,872 | 16.04% |
| Columbia | 11,777,408 | 6,452,321 | 54.79% | 3,879,919 | 60.13% | 1,525,519 | 39.32% |
| atnrpd1-3 | 10,531,269 | 3,873,131 | 36.78% | 1,830,698 | 47.27% | 365,031 | 19.94% |



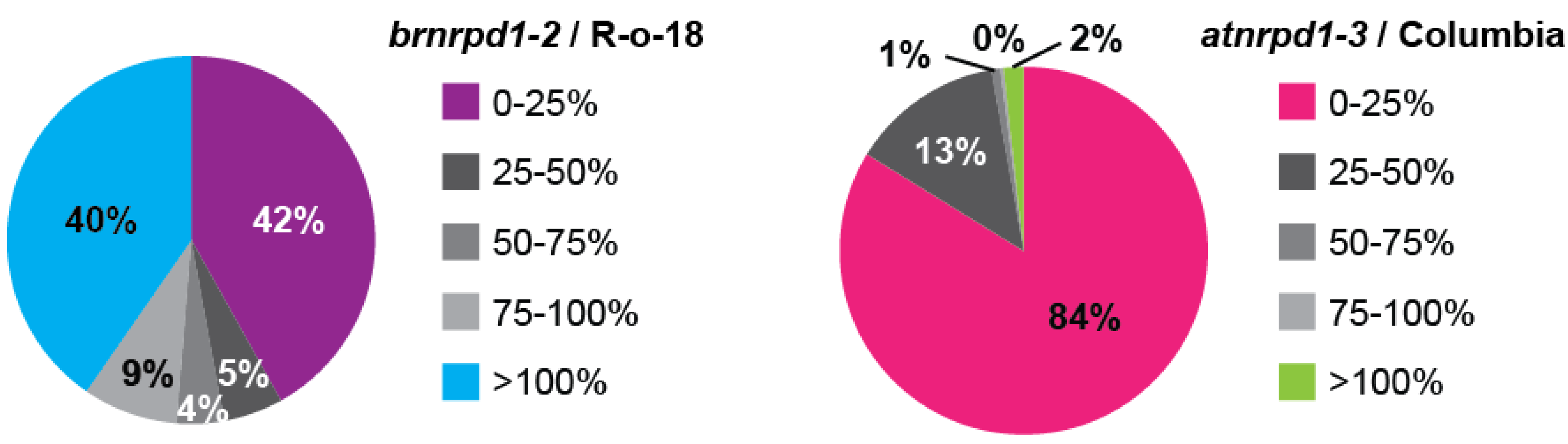
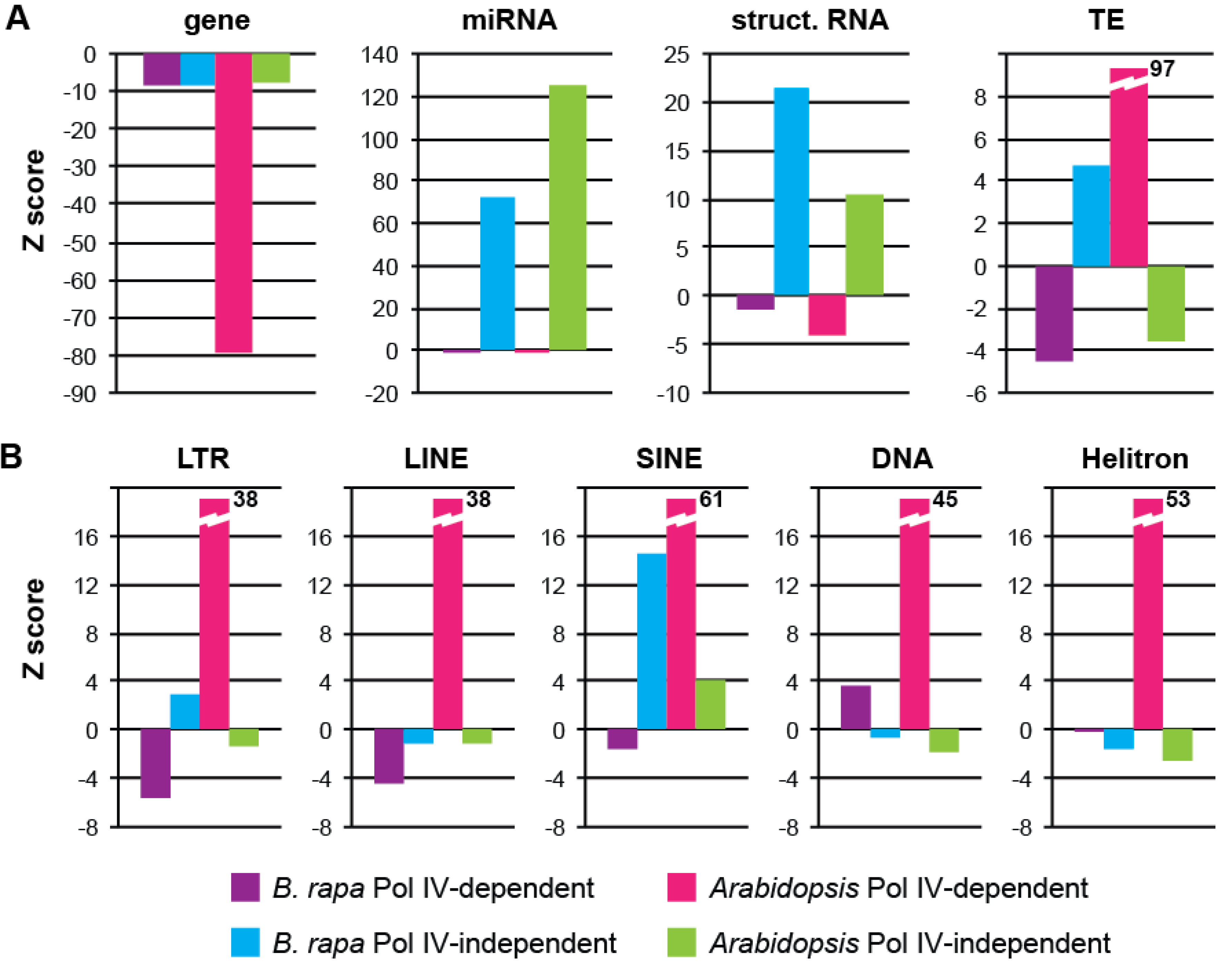
3.3. Pol IV-Dependent Loci in Brassica rapa



4. Conclusions
Supplementary Files
Acknowledgments
Conflicts of Interest
References
- Valencia-Sanchez, M.A.; Liu, J.; Hannon, G.J.; Parker, R. Control of translation and mRNA degradation by miRNAs and siRNAs. Gene Dev. 2006, 20, 515–524. [Google Scholar] [CrossRef]
- Moazed, D. Small RNAs in transcriptional gene silencing and genome defence. Nature 2009, 457, 413–420. [Google Scholar] [CrossRef]
- Mosher, R.A.; Schwach, F.; Studholme, D.; Baulcombe, D.C. PolIVb influences RNA-directed DNA-methylation independently of its role in siRNA biogenesis. Proc. Natl. Acad. Sci. USA 2008, 105, 3145–3150. [Google Scholar] [CrossRef] [Green Version]
- Haag, J.R.; Ream, T.S.; Marasco, M.; Nicora, C.D.; Norbeck, A.D.; Pasa-Tolic, L.; Pikaard, C.S. In vitro transcription activities of Pol IV, Pol V, and RDR2 reveal coupling of Pol IV and RDR2 for dsRNA synthesis in plant RNA silencing. Mol. Cell 2012, 48, 811–818. [Google Scholar] [CrossRef]
- Xie, Z.; Johansen, L.K.; Gustafson, A.M.; Kasschau, K.D.; Lellis, A.D.; Zilberman, D.; Jacobsen, S.E.; Carrington, J.C. Genetic and functional diversification of small RNA pathways in plants. PLoS Biol. 2004, 2, 642–652. [Google Scholar]
- Wierzbicki, A.T.; Ream, T.S.; Haag, J.R.; Pikaard, C.S. RNA polymerase V transcription guides ARGONAUTE4 to chromatin. Nat. Genet. 2009, 41, 630–634. [Google Scholar] [CrossRef]
- Law, J.A.; Jacobsen, S.E. Establishing, maintaining and modifying DNA methylation patterns in plants and animals. Nat. Rev. Genet. 2010, 11, 204–220. [Google Scholar] [CrossRef]
- Kasschau, K.D.; Fahlgren, N.; Chapman, E.J.; Sullivan, C.M.; Cumbie, J.S.; Givan, S.A.; Carrington, J.C. Genome-wide profiling and analysis of Arabidopsis siRNAs. PLoS Biol. 2007, 5, e57. [Google Scholar] [CrossRef]
- Zhang, X.; Henderson, I.R.; Lu, C.; Green, P.J.; Jacobsen, S.E. Role of RNA polymerase IV in plant small RNA metabolism. Proc. Natl. Acad. Sci. USA 2007, 104, 4536–4541. [Google Scholar]
- Herr, A.J.; Jensen, M.B.; Dalmay, T.; Baulcombe, D. RNA polymerase IV directs silencing of endogenous DNA. Science 2005, 308, 118–120. [Google Scholar] [CrossRef]
- Kanno, T.; Huettel, B.; Mette, M.F.; Aufsatz, W.; Jaligot, E.; Daxinger, L.; Kreil, D.P.; Matzke, M.; Matzke, A.J. Atypical RNA polymerase subunits required for RNA-directed DNA methylation. Nat. Genet. 2005, 37, 761–765. [Google Scholar] [CrossRef]
- Onodera, Y.; Haag, J.R.; Ream, T.; Nunes, P.C.; Pontes, O.; Pikaard, C.S. Plant nuclear RNA polymerase IV mediates siRNA and DNA methylation-dependent heterochromatin formation. Cell 2005, 120, 613–622. [Google Scholar] [CrossRef]
- Pontier, D.; Yahubyan, G.; Vega, D.; Bulski, A.; SaezVasquez, J.; Hakimi, M.-A.; Lerbs-Mache, S.; Colot, V.; Lagrange, T. Reinforcement of silencing at transposons and highly repeated sequences requires the concerted action of two distinct RNA polymerases IV in Arabidopsis. Gene Dev. 2005, 19, 2030–2040. [Google Scholar] [CrossRef]
- Chan, S.W.; Zilberman, D.; Xie, Z.; Johansen, L.K.; Carrington, J.C.; Jacobsen, S.E. RNA silencing genes control de novo DNA methylation. Science 2004, 303, 1336. [Google Scholar] [CrossRef]
- Perez-Hormaeche, J.; Potet, F.; Beauclair, L.; le Masson, I.; Courtial, B.; Bouche, N.; Lucas, H. Invasion of the Arabidopsis genome by the tobacco retrotransposon Tnt1 is controlled by reversible transcriptional gene silencing. Plant Physiol. 2008, 147, 1264–1278. [Google Scholar] [CrossRef] [Green Version]
- Mari-Ordonez, A.; Marchais, A.; Etcheverry, M.; Martin, A.; Colot, V.; Voinnet, O. Reconstructing de novo silencing of an active plant retrotransposon. Nat. Genet. 2013, 45, 1029–1039. [Google Scholar] [CrossRef]
- Matzke, M.; Kanno, T.; Daxinger, L.; Huettel, B.; Matzke, A.J. RNA-mediated chromatin-based silencing in plants. Curr. Opin. Cell Biol. 2009, 21, 367–376. [Google Scholar] [CrossRef]
- Pikaard, C.S.; Haag, J.R.; Ream, T.; Wierzbicki, A.T. Roles of RNA polymerase IV in gene silencing. Trends Plant Sci. 2008, 13, 390–397. [Google Scholar] [CrossRef]
- Nobuta, K.; Lu, C.; Shrivastava, R.; Pillay, M.; de Paoli, E.; Accerbi, M.; Arteaga-Vazquez, M.; Sidorenko, L.; Jeong, D.H.; Yen, Y.; et al. Distinct size distribution of endogeneous siRNAs in maize: Evidence from deep sequencing in the mop1–1 mutant. Proc. Natl. Acad. Sci. USA 2008, 105, 14958–14963. [Google Scholar] [CrossRef]
- Szittya, G.; Moxon, S.; Santos, D.M.; Jing, R.; Fevereiro, M.P.; Moulton, V.; Dalmay, T. High-throughput sequencing of Medicago truncatula short RNAs identifies eight new miRNA families. BMC Genomics 2008, 9, 593. [Google Scholar] [CrossRef]
- Mohorianu, I.; Schwach, F.; Jing, R.; Lopez-Gomollon, S.; Moxon, S.; Szittya, G.; Sorefan, K.; Moulton, V.; Dalmay, T. Profiling of short RNAs during fleshy fruit development reveals stage-specific sRNAome expression patterns. Plant. J. 2011, 67, 232–246. [Google Scholar] [CrossRef]
- Jeong, D.H.; Park, S.; Zhai, J.; Gurazada, S.G.; de Paoli, E.; Meyers, B.C.; Green, P.J. Massive analysis of rice small RNAs: Mechanistic implications of regulated microRNAs and variants for differential target RNA cleavage. Plant Cell 2011, 23, 4185–4207. [Google Scholar] [CrossRef]
- Aryal, R.; Yang, X.; Yu, Q.; Sunkar, R.; Li, L.; Ming, R. Asymmetric purine-pyrimidine distribution in cellular small RNA population of papaya. BMC Genomics 2012, 13, 682. [Google Scholar] [CrossRef]
- Ortiz-Morea, F.A.; Vicentini, R.; Silva, G.F.; Silva, E.M.; Carrer, H.; Rodrigues, A.P.; Nogueira, F.T. Global analysis of the sugarcane microtranscriptome reveals a unique composition of small RNAs associated with axillary bud outgrowth. J. Exp. Bot. 2013, 64, 2307–2320. [Google Scholar] [CrossRef]
- Sun, F.; Guo, W.; Du, J.; Ni, Z.; Sun, Q.; Yao, Y. Widespread, abundant, and diverse TE-associated siRNAs in developing wheat grain. Gene 2013, 522, 1–7. [Google Scholar] [CrossRef]
- Zhao, Y.T.; Wang, M.; Fu, S.X.; Yang, W.C.; Qi, C.K.; Wang, X.J. Small RNA profiling in two Brassica napus cultivars identifies microRNAs with oil production- and development-correlated expression and new small RNA classes. Plant Physiol. 2012, 158, 813–823. [Google Scholar] [CrossRef]
- Henderson, I.R.; Zhang, X.Y.; Lu, C.; Johnson, L.; Meyers, B.C.; Green, P.J.; Jacobsen, S.E. Dissecting Arabidopsis thaliana DICER function in small RNA processing, gene silencing and DNA methylation patterning. Nat. Genet. 2006, 38, 721–725. [Google Scholar] [CrossRef]
- Daxinger, L.; Kanno, T.; Bucher, E.; van der Winden, J.; Naumann, U.; Matzke, A.J.; Matzke, M. A stepwise pathway for biogenesis of 24-nt secondary siRNAs and spreading of DNA methylation. EMBO J. 2009, 28, 48–57. [Google Scholar] [CrossRef]
- Dunoyer, P.; Brosnan, C.A.; Schott, G.; Wang, Y.; Jay, F.; Alioua, A.; Himber, C.; Voinnet, O. An endogenous, systemic RNAi pathway in plants. EMBO J. 2010, 29, 1699–1712. [Google Scholar] [CrossRef]
- Erhard, K.F.; Stonaker, J.L.; Parkinson, S.E.; Lim, J.P.; Hale, C.J.; Hollick, J.B. RNA polymerase IV functions in paramutation in zea mays. Science 2009, 323, 1201–1205. [Google Scholar] [CrossRef]
- Stonaker, J.L.; Lim, J.P.; Erhard, K.F., Jr.; Hollick, J.B. Diversity of Pol IV function is defined by mutations at the maize rmr7 locus. PLoS Genet. 2009, 5, e1000706. [Google Scholar] [CrossRef]
- Warwick, S.I. Brassicaceae in Agriculture. Plant. Genet. Genomics 2011, 9, 33–65. [Google Scholar]
- Wang, X.; Wang, H.; Wang, J.; Sun, R.; Wu, J.; Liu, S.; Bai, Y.; Mun, J.H.; Bancroft, I.; Cheng, F.; et al. The genome of the mesopolyploid crop species Brassica rapa. Nat. Genet. 2011, 43, 1035–1039. [Google Scholar] [CrossRef]
- Stephenson, P.; Baker, D.; Girin, T.; Perez, A.; Amoah, S.; King, G.J.; Ostergaard, L. A rich TILLING resource for studying gene function in Brassica rapa. BMC Plant Biol. 2010, 10, 62. [Google Scholar] [CrossRef]
- Cheng, F.; Liu, S.; Wu, J.; Fang, L.; Sun, S.; Liu, B.; Li, P.; Hua, W.; Wang, X. BRAD, the genetics and genomics database for Brassica plants. BMC Plant Biol. 2011, 11, p. 136. Available online: http://www.brassicadb.org/ (accessed on 1 August 2013).
- Mosher, R.A.; Tan, E.H.; Shin, J.; Fischer, R.L.; Pikaard, C.S.; Baulcombe, D.C. An atypical epigenetic mechanism affects uniparental expression of Pol IV-dependent siRNAs. PLoS One 2011, 6, e25756. [Google Scholar]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 2009, 10, R25. [Google Scholar] [CrossRef]
- Birchler, J.A.; Veitia, R.A. The gene balance hypothesis: From classical genetics to modern genomics. Plant Cell 2007, 19, 395–402. [Google Scholar] [CrossRef]
- Haag, J.R.; Pikaard, C.S. Multisubunit RNA polymerases IV and V: Purveyors of non-coding RNA for plant gene silencing. Nat. Rev. Mol. Cell Biol. 2011, 12, 483–492. [Google Scholar] [CrossRef]
- Wierzbicki, A.T.; Cocklin, R.; Mayampurath, A.; Lister, R.; Rowley, M.J.; Gregory, B.D.; Ecker, J.R.; Tang, H.; Pikaard, C.S. Spatial and functional relationships among Pol V-associated loci, Pol IV-dependent siRNAs, and cytosine methylation in the Arabidopsis epigenome. Gene Dev. 2012, 26, 1825–1836. [Google Scholar] [CrossRef]
- Mi, S.J.; Cai, T.; Hu, Y.G.; Chen, Y.; Hodges, E.; Ni, F.R.; Wu, L.; Li, S.; Zhou, H.; Long, C.Z.; et al. Sorting of small RNAs into Arabidopsis argonaute complexes is directed by the 5' terminal nucleotide. Cell 2008, 133, 116–127. [Google Scholar] [CrossRef]
- Havecker, E.R.; Wallbridge, L.M.; Hardcastle, T.J.; Bush, M.S.; Kelly, K.A.; Dunn, R.M.; Schwach, F.; Doonan, J.H.; Baulcombe, D.C. The arabidopsis RNA-directed DNA methylation argonautes functionally diverge based on their expression and interaction with target loci. Plant Cell 2010, 22, 321–334. [Google Scholar] [CrossRef]
- Haag, J.R.; Pontes, O.; Pikaard, C.S. Metal A and metal B sites of nuclear RNA polymerases Pol IV and Pol V are required for siRNA-dependent DNA methylation and gene silencing. PLoS One 2009, 4, e4110. [Google Scholar]
- Pereira, V. Insertion bias and purifying selection of retrotransposons in the Arabidopsis thaliana genome. Genome Biol. 2004, 5, R79. [Google Scholar] [CrossRef] [Green Version]
- Lu, C.; Kulkarni, K.; Souret, F.F.; MuthuValliappan, R.; STej, S.S.; Poethig, R.S.; Henderson, I.R.; Jacobsen, S.E.; Wang, W.; Green, P.J.; et al. MicroRNAs and other small RNAs enriched in the Arabidopsis RNA-dependent RNA polymerase-2 mutant. Genome Res. 2006, 16, 1276–1288. [Google Scholar] [CrossRef]
- Mosher, R.A.; Melnyk, C.W.; Kelly, K.A.; Dunn, R.M.; Studholme, D.J.; Baulcombe, D.C. Uniparental expression of PolIV-dependent siRNAs in developing endosperm of Arabidopsis. Nature 2009, 460, 283–286. [Google Scholar] [CrossRef]
- Huettel, B.; Kanno, T.; Daxinger, L.; Aufsatz, W.; Matzke, A.J.M.; Matzke, M. Endogenous targets of RNA-directed DNA methylation and Pol IV in Arabidopsis. EMBO J. 2006, 25, 2828–2836. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Huang, Y.; Kendall, T.; Mosher, R.A. Pol IV-Dependent siRNA Production is Reduced in Brassica rapa. Biology 2013, 2, 1210-1223. https://doi.org/10.3390/biology2041210
Huang Y, Kendall T, Mosher RA. Pol IV-Dependent siRNA Production is Reduced in Brassica rapa. Biology. 2013; 2(4):1210-1223. https://doi.org/10.3390/biology2041210
Chicago/Turabian StyleHuang, Yi, Timmy Kendall, and Rebecca A. Mosher. 2013. "Pol IV-Dependent siRNA Production is Reduced in Brassica rapa" Biology 2, no. 4: 1210-1223. https://doi.org/10.3390/biology2041210